IDENTIFIKASI KERAGAMAN GEN
κ
-KASEIN
DAN HUBUNGANNYA DENGAN KUALITAS
SUSU KERBAU LOKAL
ANNISA OKTAVIA RINI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Identifikasi Keragaman Gen κ-Kasein dan Hubungannya dengan Kualitas Susu Kerbau Lokal adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, September 2014
Annisa Oktavia Rini
RINGKASAN
ANNISA OKTAVIA RINI. Identifikasi Keragaman Gen κ-Kasein dan Hubungannya dengan Kualitas Susu Kerbau Lokal. Dibimbing oleh CECE SUMANTRI dan EVY DAMAYANTHI.
Terdapat dua tipe kerbau lokal Indonesia, yakni kerbau sungai (riverine buffalo) dan kerbau rawa atau kerbau lumpur (swamp buffalo). Selain sebagai penghasil daging dan tenaga kerja, kerbau dapat dimanfaatkan sebagai penghasil susu. Susu merupakan salah satu pangan fungsional karena mempunyai nutrisi lengkap secara alami dan mudah diserap oleh tubuh.
Gen κ-Kasein dikenal berperan dalam mengontrol protein susu serta berperan dalam koagulasi dan pembentukan curd susu. Produk olahan susu kerbau di Indonesia masih bersifat tradisional, antara lain dadih, dali, dan danke. Salah satu produk olahan susu yang berbahan dasar susu kerbau dan memiliki nilai ekonomi tinggi adalah keju mozzarella. Hal tersebut merupakan peluang untuk meningkatkan nilai tambah susu kerbau lokal, namun kendala yang dihadapi adalah rendahnya produktivitas kerbau. Peningkatan produktivitas kerbau sebagai penghasil susu dapat didukung dengan peningkatan mutu genetik kerbau. Penelitian ini bertujuan untuk mengidentifikasi keragaman gen κ-Kasein pada kerbau lokal di Indonesia menggunakan teknik PCR-RFLP (polymerase chain reaction-restriction fragment length polymorphism) serta menganalisis hubungan antara keragaman genotipe dengan kualitas susu kerbau lokal.
Sampel yang digunakan berasal darah dan susu kerbau. Sampel darah sebanyak 290 ekor, terdiri atas 40 ekor kerbau sungai dan 250 ekor kerbau rawa. Sampel susu sebanyak 40 sampel, terdiri atas 13 sampel susu kerbau sungai dan 27 sampel susu kerbau rawa. Isolasi DNA menggunakan metode ekstraksi phenol-chloroform (Sambrook et al. 1989) yang dimodifikasi oleh Andreas et al. (2010). Pendeteksian keragaman gen κ-Kasein menggunakan metode PCR-RFLP dengan
EcoRV sebagai enzim restriksi. Analisis data menggunakan frekuensi alel, frekuensi genotipe, nilai heterozigositas, polymorphic informative content (PIC), dan perunutan susunan nukleotida. Hubungan antara varian genotipe gen κ-Kasein dengan kualitas susu dianalisa secara deskriptif.
SUMMARY
ANNISA OKTAVIA RINI. Identification of κ-Casein Gene Polymorphisms and Its Related with Local Buffalo Milk Quality. Supervised by CECE SUMANTRI and EVY DAMAYANTHI.
There were two types of local Indonesian buffalo, namely riverine buffalo and swamp buffalo. Besides as a meat producer and labor, it can be utilized as a milk producer. Milk is one of functional food because it has complete nutrition and easily absorbed by the body.
κ-Casein gene known as gene that plays a role in the controling milk protein and plays an important role in coagulation and curdling of milk. The processing of dairy buffalo products in Indonesia were still traditional, such as dadih, dali, and
danke. One of the dairy product was made from buffalo milk and had high economic value is the mozzarella cheese. It is an opportunity to enhance the value-added of local buffalo milk, but the constraint is the low productivity of buffalo to produce milk. Increasing the productivity of buffalo as a milk producer can be supported with increasing the genetic quality of buffalo. This study aimed to identifiy the polymorphisms of κ-Casein gene of the local buffalo in Indonesia using PCR-RFLP (polymerase chain reaction-restriction fragment length polymorphism) technique and analyze the association between genotype variation with local buffalo milk quality.
The sample used in this study were blood and milk of buffalo. The total of blood sample were used as much as 290 heads, consisting of 40 riverine buffaloes and and 250 swamp buffaloes. Milk samples were used as 40 samples, consisting of 13 riverine buffalo milk and 27 swamp buffalo milk. DNA isolation was done from blood using phenol-chloroform extraction method (Sambrook et al. 1989) as modified by Andreas et al. (2010). Identification of κ-Casein gene polymorphisms used PCR-RFLP, with EcoRV as the restriction enzyme. Data analysis were conducted using allele frequency, genotype frequency, heterozygosity, polymorphic informative content (PIC), and sequencing. The association between κ-Casein variant genotype with milk quality were analyzed descriptively.
Amplification of the κ-Casein gene resulted an amplicon with lenght of 157 bp, located in exon 4. Genotyping κ-Casein gene of riverine buffaloes produce three genotypes, with frequencies were CC (0.775), CT (0.200), and TT (0.025); while genotyping in swamp buffaloes produced only CC genotype (100%), meaning this gene was monomorphic. The observation heterozygosity of riverine (Ho = 0.200) and swamp buffaloes (Ho = 0.000) were low. The highest of PIC value was found in riverine buffalo at Siborong-borong Buffalo Breeding Center (PIC = 0.288). Sequencing of κ-Casein gene fragment of riverine buffalo showed two point mutations from Citosin (C) to Timin (T) at base position 23 bp (Thr/ACC to Ile/ATC) and 27 bp (silent mutation Thr/ACC to Thr/ACT) of PCR product. In general, swamp buffalo milk has higher percentage of fat, SNF (solid non fat), protein, and lactose than riverine buffalo. κ-Casein gene fragment in this study can not used as a genetic marker of buffalo milk quality.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Produksi dan Teknologi Peternakan
IDENTIFIKASI KERAGAMAN GEN
κ
-KASEIN
DAN HUBUNGANNYA DENGAN KUALITAS
SUSU KERBAU LOKAL
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
PRAKATA
Puji dan syukur Penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga penulisan hasil penelitian tesis ini berhasil diselesaikan dengan judul: “Identifikasi Keragaman Gen κ-Kasein dan Hubungannya dengan Kualitas Susu Kerbau Lokal”. Penulis menyadari bahwa proses penelitian dan penulisan dari tesis ini tidak akan berjalan tanpa adanya dukungan dari banyak pihak. Oleh sebab itu pada kesempatan ini penulis ingin menyampaikan ucapan terimakasih.
Kepada yang terhormat Prof Dr Ir Cece Sumantri MAgrSc dan Prof Dr Ir Evy Damayanthi MS selaku komisi pembimbing, penulis mengahanturkan ucapan terima kasih atas curahan waktu, arahan, bimbingan, dan motivasi mulai dari penyusunan proposal, pelaksanaan penelitian hingga penulisan tesis. Penulis juga menyampaikan ucapan terimakasih kepada Dr Epi Taufik SPt MVPH Msi selaku penguji luar komisi pada ujian sidang atas saran dan masukan yang diberikan.
Kepada Dr Jakaria SPt MSi dan Dr agr Asep Gunawan SPt MSc penulis mengucapkan terimakasih atas saran serta masukan selama studi. Penulis mengucapkan terimakasih kepada Dr Ir Salundik MSi selaku Ketua Program Studi Ilmu Produksi dan Teknologi Peternakan (ITP) serta jajarannya (Ibu Ade dan Mba Okta) di sekretariat Pasca ITP.
Terima kasih kepada Direktorat Jenderal Pendidikan Tinggi atas Beasiswa
Fresh Graduate yang diberikan kepada Penulis selama menempuh pendidikan pascasarjana dan kepada program Hibah KKP3N yang telah membiayai penelitian. Penulis sampaikan terima kasih kepada Kepala dan seluruh staf Balai Pembibitan Ternak Unggul Babi dan Kerbau Siborong-borong (BPTU-Siborong-borong) Tapanuli Utara-Provinsi Sumatera Utara dan Program Hibah Pascasarjana 2012 atas materi penelitian yang diberikan. Terima kasih kepada Laboratorium Genetika Molekuler Ternak (LGMT) bagian Pemuliaan dan Genetika Ternak dan Laboratorium Pengolahan Susu- Fapet IPB atas ijin melakukan penelitian dan penggunaan alat.
Penulis sampaikan terimakasih kepada teman-teman ITP angkatan 2012++, The Helix dan ABGSCi atas dukungan, kebersamaan, dan kasih sayang yang terjalin, semoga persahabatan ini akan tetap berjalan sampai waktu mendatang. Kepada Eryk Andreas SPt MSi, Irine SPt MSi, Ferdy Saputra SPt MSi, dan Shelvi Ssi, penulis mengucapkan terimakasih atas saran dan pendampingan selama di LGMT. Terimakasih kepada Heni Rizqiati SPt MSi dan Deni Alamsah SP, yang banyak membantu dan memberikan dukungan selama penelitian dilaksanakan.
Penulis ucapkan terimakasih yang tulus kepada keluarga tercinta Ayahanda Adi Suwardi (Almarhum), Ibunda Sulfanah Ma’ani, Kakak Andhika Fandrian Apriyandi, Amalia Wanda Riani dan Alfath Akmal Azzafran. Kepada Pria Sembada, terimakasih atas dukungan dan motivasi yang diberikan, salam sukses bermanfaat.
Semoga karya ilmiah ini bermanfaat.
Bogor, September 2014
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 3
Manfaat Penelitian 3
2 TINJAUAN PUSTAKA 3
Kerbau 3
Susu Kerbau 4
Komposisi Susu 5
Faktor yang Berpengaruh terhadap Produksi Susu 6
κ-Kasein 7
Penanda Molekuler 8
Keragaman Genetik 10
3 METODE 10
Lokasi dan Waktu 11
Materi 11
Prosedur Analis15is 11
Prosedur Analisis Data 13
4 HASIL DAN PEMBAHASAN 15
Amplifikasi Gen κ-Kasein 15
Keragaman Gen κ-Kasein dengan Metode PCR-RFLP 15
Frekuensi Alel dan Genotipe Gen κ-Kasein 16
Derajat Polimorfisme Gen κ-Kasein|EcoRV 19
Runutan Ruas Gen κ-Kasein 20
Polimorfisme Gen κ-Kasein|EcoRV dengan Kualitas Susu 21 Karakteristik Kualitas Susu Kerbau Sungai dan Rawa 23
5 SIMPULAN DAN SARAN 26
Simpulan 26
Saran 26
DAFTAR PUSTAKA 26
LAMPIRAN 32
DAFTAR TABEL
1 Komposisi susu kerbau dan sapi 4
2 Performa reproduksi kerbau 7
3 Frekuensi Gen κ-Kasein|EcoRV pada kerbau sungai dan rawa 17 4 Frekuensi Gen κ-Kasein ekson 4 pada kerbau hasil penelitian dari
sumber lain 18
5 Nilai heterozigositas pengamatan (Ho), Harapan (He), dan PIC 19 6 Karakteristik kualitas susu kerbau sungai dengan genotipe berbeda 22 7 Karakteristik kualitas susu kerbau sungai dan rawa bergenotipe CC 24
DAFTAR GAMBAR
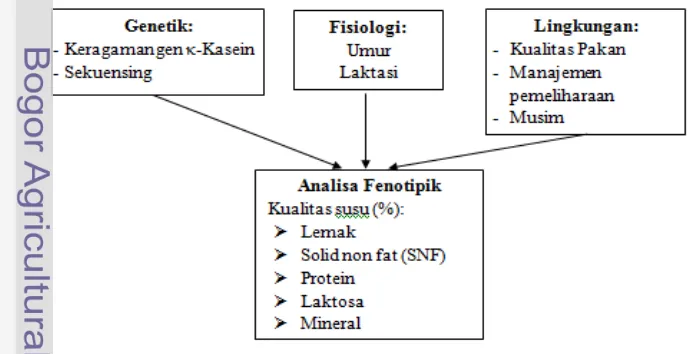
1 Kerangka pemikiran penelitian 2
2 Kerbau lokal Indonesia 3
3 Rekonstruksi struktur gen κ-Kasein berdasarkan sekuen di GenBank
nomor akses AM900443 (Masina et al. 2007) 8
4 Posisi penempelan primer pada sekuen gen κ-Kasein sebagian ruas
ekson 4 11
5 Visualisasi elektroforesis amplifikasi gen κ-Kasein pada gel agarosa
1.5% 15
6 Hasil pemotongan gen κ-Kasein pada kerbau sungai menggunakan metode PCR-RFLP dengan enzim restriksi EcoRV pada gel agarosa 2% 16
7 Runutan nukleotida ruas gen κ-Kasein 20
8 Perbandingan rataan kualitas susu kerbau sungai dan rawa lokal
genotipe CC 24
DAFTAR LAMPIRAN
1
PENDAHULUAN
Latar Belakang
Kerbau merupakan salah satu ternak yang bisa beradaptasi dengan lingkungan tropis di Indonesia yang tersebar hampir di setiap pulau, namun perkembangan populasi kerbau di Indonesia mengalami penurunan. Berdasarkan data Badan Pusat Statistik, tahun 2007 populasi kerbau diperkirakan sebanyak 2.086.000 ekor, sedangkan populasi kerbau di Indonesia pada tahun 2012 diperkirakan sebanyak 1.438.000 ekor (BPS 2012). Kerbau memiliki keunggulan untuk dikembangkan karena mampu bertahan hidup dengan pakan berkualitas rendah, mampu bertahan hidup pada tekanan iklim setempat, dan daya tahan yang tinggi terhadap penyakit dan parasit. Terdapat dua tipe kerbau lokal Indonesia, yakni kerbau sungai (riverine buffalo) dan kerbau rawa atau kerbau lumpur (swamp buffalo). Selain sebagai penghasil daging dan tenaga kerja, kerbau dapat dimanfaatkan sebagai penghasil susu. Susu merupakan salah satu pangan fungsional karena mempunyai nutrisi lengkap secara alami dan mudah diserap oleh tubuh. Sebagai salah satu bahan pangan terbaik, susu memiliki fungsi fisiologis untuk pencegahan penyakit seperti osteoporosis, obesitas, hipertensi, kanker, serta meningkatkan sistem imunitas tubuh terhadap berbagai penyakit sehingga tercapai kesehatan yang optimal.
Kualitas susu merupakan salah satu faktor penting bagi konsumen dan beberapa industri olahan susu. Susu kerbau mengandung bahan padat yang tinggi dan kadar air yang lebih rendah daripada susu sapi. Kadar lemak susu kerbau lebih tinggi yaitu antara 7-8% sedangkan susu sapi hanya sekitar 3.5%, protein susu kerbau juga lebih tinggi yaitu 5-6% sedangkan pada sapi 3.3%. κ-Kasein adalah protein susu yang menyusun sekitar 12 - 15% dari total kasein pada susu sapi dan bertindak sebagai stabilisasi, yaitu mempertahankan seluruh kompleks kasein dalam suspensi koloidal yang memberikan warna putih susu (Soeparno 1992) serta berperan dalam menentukan ukuran dan fungsi dari butiran susu (Yahyaoui et al. 2003). Keragaman gen kasein berhubungan dengan perbedaan komposisi, pengolahan dan kualitas, serta nilai karakteristik susu. Penelitian mengenai keragaman gen κ-Kasein telah banyak dilakukan pada sapi (Rijnkels et al. 1997). Sumantri et al. (2005) melaporkan keragaman gen κ-Kasein pada ternak sapi perah di BPTU Baturraden berpengaruh nyata terhadap kualitas susu. κ -Kasein alel B diketahui berasosiasi berhubungan dengan kadar lemak, protein, dan kasein yang tinggi, serta waktu koagulasi rennet yang lebih baik sehingga efisien untuk pengolahan keju.
2
dan rendah kolesterol. Peningkatan produktivitas kerbau sebagai penghasil susu dapat didukung dengan peningkatan mutu genetik ternak kerbau salah satunya adalah seleksi kualitas susu melalui pendekatan molekuler menggunakan marka genetik gen κ-Kasein. Faktor lain yang dapat mempengaruhi kualitas susu antara lain bangsa, genetik, umur, laktasi, musim, lingkungan, manajemen pemeliharaan. Potensi keanekaragaman sumberdaya genetik kerbau perlu dimanfaatkan secara berkelanjutan untuk meningkatkan produktivitas serta mewujudkan kesejahteraan masyarakat, ketersediaan bahan pangan, terciptanya lapangan kerja dan peningkatan devisa negara. Kendala yang dihadapi saat ini adalah informasi mengenai karakteristik kerbau lokal belum banyak diketahui. Informasi tersebut sangat penting dan diperlukan bagi pengembangan dan perbaikan mutu genetik ternak kerbau secara nasional. Penelitian keragaman gen κ-Kasein pada kerbau di Indonesia masih minim dilakukan. Penelitian ini betujuan untuk diidentifikasi keragaman gen κ-Kasein pada kerbau lokal di Indonesia menggunakan teknik
polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) serta menganalisis hubungan antara variasi genotipe dengan kualitas susu kerbau lokal.
Perumusan Masalah
Indonesia memiliki dua tipe kerbau, yaitu kerbau sungai dan rawa yang tersebar hampir di seluruh wilayah bagian Indonesia. Kerbau sungai yang merupakan tipe tipe perah (penghasil susu) umumnya hanya ditemui di wilayah Sumatera Utara. Kerbau rawa banyak digunakan sebagai tipe pekerja dan potong. Karakteristik fenotipe yang diamati dalam penelitian ini adalah mengukur kualitas susu. Penilaian kualitas susu mewakili persentase lemak, bahan kering tanpa lemak (BKTL), protein, laktosa, dan mineral. Laporan tentang adanya pengaruh dari faktor genetik, fisiologi, dan lingkungan terhadap sifat fenotipe telah banyak dilaporkan pada sapi, sedangkan pada kerbau jarang ditemukan. Penelitian ini dirancang untuk mendapatkan informasi keragaman gen κ-Kasein dan hubungannya dengan kualitas susu kerbau lokal. Penelitian ini dirancang berdasarkan tinjauan pustaka yang terkait dengan topik penelitian. Kerangka pemikiran diatas kemudian diringkas dalam Gambar 1.
3 Tujuan Penelitian
Penelitian ini bertujuan untuk mengidentifikasi keragaman gen κ-Kasein pada kerbau lokal di Indonesia menggunakan teknik PCR-RFLP serta menganalisis hubungan antara keragaman genotipe dengan kualitas susu kerbau lokal.
Manfaat Penelitian
Hasil identifikasi gen κ-Kasein kerbau lokal ini akan dijadikan sebagai informasi dan dasar penentuan seleksi mendapatkan kerbau dengan kualitas protein tinggi dalam penerapan marker assisted selection (MAS) serta mengetahui parameter kualitas susu kerbau sungai dan rawa lokal.
2
TINJAUAN PUSTAKA
Kerbau
Kerbau termasuk ke dalam family Bovidae, genus Bubalus yang berkembang di banyak bagian dunia dan diduga berasal dari daerah India. Kerbau domestikasi atau water buffalo yang terdapat saat ini berasal dari spesies Bubalus arnee. Spesies kerbau lainnya yang masih liar adalah Bubalus mindorensis,
Bubalus depressicornis dan Bubalus caffer (Hasinah dan Handiwirawan 2006). Kerbau domestik (Bubalus bubalis) terdiri atas dua tipe yaitu tipe sungai (riverine buffalo) dan tipe rawa (swamp buffalo). Kedua tipe kerbau ini dibedakan berdasarkan pada penampakan, tingkah laku, pemanfaatan dan habitatnya. Kerbau sungai mempunyai bentuk tanduk melingkar ke bawah dan kerbau rawa mempunyai tanduk melengkung ke belakang. Kerbau sungai menunjukkan kesenangan terhadap air mengalir yang bersih, sedangkan kerbau rawa suka berkubang dalam lumpur, rawa-rawa dan air menggenang. Kerbau rawa dipelihara terutama sebagai ternak kerja dan untuk produksi daging, namun di beberapa daerah kerbau ini juga diperah (Bahri dan Talib 2007; Wirdahayati 2008). Sebaliknya kerbau sungai adalah termasuk tipe perah, dan salah satu diantaranya adalah kerbau Murrah.
a b
4
Kerbau sungai banyak terdapat di Sumatera Utara yaitu kerbau Murrah yang dipelihara oleh masyarakat keturunan India dan digunakan sebagai penghasil susu. India memiliki jenis kerbau sungai yang beragam dengan populasi kerbau sungai sebesar 95 juta ekor dan hampir 56% dari total produksi susu nasionalnya berasal dari susu kerbau. Produksi susu kerbau Murrah India dapat mencapai 1.029-2.565 kg laktasi-1 (Dhanda 2006); sedangkan pada kerbau mediterranean di Italia produksi susu dapat mencapai 5.000 kg laktasi-1 (ANASB 2003). Produksi susu kerbau rawa di China rata-rata sebesar 2.6 kg hari-1 (Chunxi Z dan Zhongquan 2001), Vietnam sebesar 1.50 kg hari-1 (Nguyen van Thu 2000a). Perbadaan kerbau rawa dan sungai bila dilihat dari tingkat kromosom, kerbau rawa mempunyai 24 pasang kromosom (48 kromosom), sedangkan kerbau sungai 25 pasang (50 kromosom).
Susu Kerbau
Susu merupakan suspensi koloidal yang mengandung lemak, protein, karbohidrat, mineral, vitamin, dan enzim. Secara umum, komposisi susu setiap spesies hampir sama, hanya saja terdapat perbedaan dari segi kualitas dan kuantitas (Huppertz dan Kelly 2009). Susu kerbau mudah dikenal karena memiliki ciri-ciri warna lebih putih, lebih kaya lemak, globula lemak susu lebih kecil dan teremulsi dengan susu. Lemaknya lebih mudah dicerna dan mengandung mineral yang lengkap. Dadih merupakan salah satu produk susu kerbau fermentasi yang berasal dari Sumatera Barat yang merupakan bahan pangan tradisional (Surono et al. 2008) yang memiliki kandungan protein dan lemak lebih tinggi dibandingkan dadih susu sapi dan kambing (Sunarlim 2009). Susu kerbau, seperti halnya susu sapi dapat diproses menjadi berbagai macam produk seperti es krim, yoghurt, dan
buttermilk. Untuk membuat 1 kg keju dibutuhkan 8 kg susu sapi, tetapi dengan susu kerbau hanya 5 kg, sedangkan untuk membuat mentega yang dengan susu sapi dibutuhkan 14 kg, dengan susu kerbau hanya membutuhkan 10 kg (Hasinah dan Handiwirawan 2007). Tabel 1 menunjukkan variasi komposisi susu kerbau dan bangsa sapi.
Tabel 1 Komposisi susu kerbau dan sapi
Ternak
Kualitas susu (%) Sumber
Protein Lemak Laktosa Abu Padatan Total
Kerbau 4.40 7.10 5.20 8.00 17.50 Ahmad et al.
(2008) 4.60 7.30 5.60 - 17.60 Menard et al.
(2010) 5.00 7.10 4.60 9.00 17.70 Han et al.
(2012) Sapi
Jersey 3.31 5.77 4.67 0.57 - Fayeye et al.
(2013)
5 Beberapa penelitian menunjukkan bahwa produksi susu dari setiap ternak kerbau rawa yang diperah berkisar antara 1.50-2.50 liter ekor-1 hari-1 dengan lama pemerahan sekitar 7 bulan (Zulbadri 2002). Kadar lemak susu kerbau pada umumnya (tipe perah dan tipe pedaging) antara 6.6 – 9.0% diatas kadar lemak susu sapi 3.6 – 4.9% (Dhanda 2006) yang antara lain dipengaruhi oleh bangsa ternak dan faktor pakan.
Komposisi Susu
Kemampuan proses dan fungsi susu dan produk susu ditentukan oleh sifat dan konsentrasi komponen utamanya, yaitu protein, lemak, laktosa, dan garam. Konsentrasi dan sifat komponen penyusun susu dapat bervariasi. Perbedaan tersebut dapat terjadi akibat pengaruh spesies, bangsa, kondisi kesehatan, status nutrisi, tingkat laktasi dan umur yang berbeda (Fox 2003).
Susu sapi mengandung 6 protein susu spesifik : 4 kasein, antara lain αs1-, αS2, β- dan κ-, masing-masing sebesar 38%, 10%, 36%, dan 15% dari keseluruhan
kasein; serta β-laktoglobulin dan α-laktalbumin, masing-masing sebesar 40% dan 20% dari total whey protein. Selain itu juga mengandung beberapa whey protein minor, termasuk bovine serum albumin (BSA) dan immunoglobulins (Ig) yang di transfer dari darah ; masing-masing sebesar 10% dari whey protein dari susu sapi dewasa. Kandungan protein susu relatif tetap selama laktasi. Protein susu yang berupa kasein, β-laktoglobulin dan α-laktalbumin disintesis di dalam kelenjar ambing yang dikontrol oleh gen, sedangkan sisanya diabsorpsi dari darah (Fox 2003).
Kandungan lemak susu sapi serupa dengan kandungan lemak susu spesies lain yang mengandung banyak triacylglycerol dan sejumlah kecil diacylglyserol, monoacylglycerol, asam lemak bebas, phospolipid, dan sterol. Sejumlah vitamin larut dalam lemak, yaitu A, D, E, K (Hasim dan Martindah 2012) dan β-karoten terdapat di dalam lemak susu sapi. Triacylglycerol menyusun sebanyak 98% dari lemak total, sehingga memiliki efek secara langsung terhadap sifat lemak susu, contohnya: hidrofobisitas, berat jenis, sifat leleh. (MacGibbon dan Taylor 2006).
Karbohidrat utama dari susu adalah laktosa yang terdapat dalam bentuk alfa dan beta. Laktosa terlarut di dalam susu sehingga mempengaruhi stabilitas titik beku, titik didih dan tekanan osmosis dari susu. Kadar laktosa dalam susu sapi adalah 4.8 (g 100 g-1) (Huppertz dan Kelly 2009). Kadar laktosa relatif tetap, namun produksi laktosa meningkat sejalan dengan peningkatan produksi susu. Fluktuasi kadar laktosa terjadi sesuai dengan dinamika produksi susu selama laktasi (Philips 2010).
Mineral merupakan fraksi kecil dari susu (8-9 g.L-1) yang mengandung ion kation (kasium, magnesium, sodium, dan potassium) dan anion (fosfat anorganik, sitrat, dan klorida). Ion berperan dalam struktur dan kestabilan misel kasein. Ion tersebut dapat lebih atau kurang berhubungan antara ion itu sendiri dan dengan protein. Ion dapat bersifat diffusible (natrium, kalium, dan klorida) atau sebagian dapat berhubungan dengan molekul kasein (kalsium, magnesium, fosfat, dan sitrat) untuk membentuk partikel koloid besar yang disebut dengan misel kasein. Kandungan kalsium dan fosfat ditemukan tinggi pada susu sapi yang kaya akan protein (Gaucheron 2005).
6
Faktor yang Berpengaruh terhadap Komposisi Susu
Huppertz dan Kelly (2009) menyatakan bahwa perubahan komposisi susu dapat dipengaruhi oleh berberapa faktor, antara lain: faktor genetik (bangsa dan tipe), tingkatan laktasi, status kesehatan/fisiologi, dan lingkungan (pakan, iklim, dan pemerahan). Perubahan komposisi susu selama laktasi mencerminkan perubahan kualitas susu, keseimbangan energi dan tingkat konsumsi pakan. Pada minggu-minggu awal laktasi ternak sapi, kadar lemak susu akan menurun secara cepat sejalan dengan peningkatan produksi susu. Setelah minggu keempat, kadar lemak susu akan meningkat dengan perlahan. Turunnya kadar protein susu terjadi pada 12 minggu pertama laktasi, kemudian akan meningkat sampai masa laktasi berakhir. Kadar laktosa meningkat ketika susu mulai diproduksi menggantikan kolostrum serta konsentrasi laktosa cenderung stabil selama masa laktasi. Penurunan laktosa dalam susu terjadi pada akhir masa laktasi ketika dilakukan pembatasan pakan (Mariana 2010). Gurler et al. (2013) menyatakan bahwa produksi susu berbanding terbalik dengan kadar lemak, protein, dan abu.
Bangsa sapi yang berbeda akan menghasilkan komposisi susu yang berbeda, misalnya sapi perah FH menghasilkan susu dengan kandungan lemak lebih rendah apabila dibandingkan sapi Jersey (Palladino et al. 2010). Faktor-faktor lain yang mempengaruhi komposisi susu ialah keragaman akibat musim, hal ini terutama terjadi pada daerah beriklim subtropis dimana kandungan lemak akan menurun pada akhir musim semi dan akan meningkat menjelang musim dingin. Perbedaan tersebut biasanya dihubungkan dengan adanya perubahan pakan ternak dari biji-bijian pada musim dingin menjadi rumput-rumputan pada musim semi (Muchtadi 2009).
Faktor lainnya ialah umur. Umur sapi berpengaruh kecil sekali terhadap komposisi susu. Selama jangka waktu 10 tahun, rata-rata kandungan lemak susu menurun sekitar 0.2%. Penyakit juga dapat mempengaruhi komposisi susu. Penyakit pada sapi dapat mengacaukan keseimbangan komponen-komponen di dalam susu, hal tersebut menyebabkan terjadi kenaikan kadar lemak dan garam-garam mineral serta penurunan kadar laktosa (Muchtadi 2009).
Pakan berpengaruh terhadap komposisi susu. Kurangnya pemberian pakan akan mengurangi produksi susu. Keragaman cukup besar yang terjadi dalam kandungan protein dan karbohidrat dalam pakan tidak akan banyak mempengaruhi komposisi susu, akan tetapi pakan yang banyak mengandung lemak atau pakan tersebut secara sengaja dicampuri lemak atau minyak, pengaruhnya akan terlihat jelas kadar dan komposisi lemak susu. Komposisi susu dapat dipengaruhi oleh faktor-faktor dari luar, misalnya pemalsuan dengan cara menambahkan air atau bahan lain, kegiatan mikroba, kurangnya homogenisasi dalam pengambilan sampel, dan lain lain (Muchtadi 2009).
7 produktivitas ternak kerbau perlu ditingkatkan. Hal tersebut ditunjukkan pada Tabel 2.
Tabel 2 Performa reproduksi kerbau (Abubakar 2010)
Karakteristik Selang waktu
Jarak beranak > 20 bulan
Umur beranak pertama 3.5 – 4 tahun
Estrus post partum 6 – 12 bulan
Dewasa kelamin 3 tahun
Lama kebuntingan 330 – 334 hari
κ – Kasein
Caroli et al. (2009) melaporkan bahwa 6 protein utama pada susu (αs1-. β-,
αs2-, κ-Kasein, α-LA, dan α-LG) menunjukkan polimorfisme genetik. Fitzgerald
(1997) menambahkan bahwa variasi genetik protein berhubungan dengan komposisi susu terutama kasein. Susu sapi umumnya mengandung 3.5% protein dengan pembagian sekitar 80% kasein dan 20% whey (Narayan 2013). Protein susu sebagian besar (sekitar 72-82%) berupa kasein yang dikontrol oleh empat lokus saling berdekatan (haplotipe), dengan runutan genom αs1-. β-, αs2- dan κ
-Kasein pada panjang 250 pb terletak di kromosom 6/BTA 6q31 (Rijnkels et al.
1997). Keterkaitan lokasi yang sangat dekat antara keempat gen kasein dan diwariskan lebih sebagai klaster, menjadikannya potensial untuk dieksplorasi sebagai marka pembantu seleksi sifat protein susu dan kualitas susu (Azevedo et al. 2008).
Protein κ-Kasein pada sapi tersusun atas 169 asam amino, sedangkan pada kambing tersusun atas 171 asam amino (Yahyaoui et al. 2003) dengan berat molekul 19 kD. κ-Kasein terletak pada kromosom 6 pada kelompok bovine
(Azevedo et al. 2008) dan kromosom 7 pada kerbau (Iannuzzi 1998). Variasi tipe B (alel B) gen κ-Kasein dihasilkan dari titik mutasi (T/C) pada posisi ekson 4, mempengaruhi peningkatan efisiensi produksi keju. Pada sperma, genotipe BB atau AB adalah indikator genotipe ideal untuk produksi susu dan pembuatan keju, dan juga menurunkan waktu koagulasi dan meningkatkan kestabilan curd. (Abbasi
et al. 2009).
Mitra et al. (1998) melaporkan bahwa pasangan primer (K1 dan K2) gen CSN3 menghasilkan fragmen sepanjang 400 pb. Penggunaan teknik RFLP dengan enzim endonuklease HindIII dan HinfI yang sebelumnya pernah digunakan untuk identifikasi keragaman gen CSN3 pada kerbau Murrah dan Nili-Ravi menghasilkan pita monomorfik yang menunjukkan hanya adanya alel CSN3 B.
Beneduci et al. (2010) melaporkan bahwa gen CSN3 (κ-Kasein) pada kerbau Murrah Bulgarian di ekson 4 memiliki fragmen sepanjang 221 pb. Sekuen yang berurutan dianalisa menggunakan Bioedit v 7.0.5 dan terdeteksi perubahan dari Ile menjadi Thr pada posisi kodon 135. Selain itu, terjadi silent mutation pada Thr136 (ACC/ACT), namun tidak terjadi variasi genetik pada Ala148. Mitra et al.
8
B adalah 0.76 dan 0.24. Simpangan antara frekuensi genotipe yang diamati dengan harapan menggunakan keseimbangan Hardy Weinberg ditemukan tidak signifikan (P>0.05), dimana populasi kerbau Murrah Bulgarian berada pada keadaan seimbang pada lokus CSN3.
Keterangan :
Lokus = AM900443 Panjang = 14692 pb
Gen = 1058–1121, 3559–3620, 9527–9559, 11669 –12185, 14045–14216
Sekuen depan = 1057 = 1057 pb
Exon 1 = 1058–1121 = 64 pb Intron 1 = 1122–3558 = 2437 pb Exon 2 = 3559–3620 = 62 pb Intron 2 = 3621–9526 = 5906 pb Exon 3 = 9527–9559 = 33 pb Intron 3 = 9560–11668 = 2109 pb Exon 4 = 11669–12185= 517 pb Intron 4 = 12186–14044 = 1859pb Exon 5 = 14045–14216= 172 pb Sekuen ujung = 14217 pb
Gambar 3 Rekonstruksi struktur gen κ-Kasein berdasarkan sekuen di GenBank
nomor akses AM900443 (Masina et al. 2007)
Penelitian mengenai kerbau menggunakan metode PCR-RFLP dengan enzim restriksi HindIII, HinfI, dan TaqI (Abassi et al. 2009 dan Dayem et al. 2009) dapat mendeteksi keragaman CSN3 pada sapi (variasi alel A dan B, dengan situs mutasi T136I dan D148A), namun tidak terdeteksi pada populasi kerbau (Beneduci et al. 2010). Keragaman gen pada susu berhubungan erat dengan sifat koagulasi, sehingga berpengaruh dalam proses produksi dan komposisi hasil olahan susu. Pada variasi B berhubungan dengan tingginya kadar lemak, protein, dan kasein yang terdapat di dalam susu sapi, dan hal tersebut sangat mempengaruhi pembuatan keju dan koagulasi rennet yang lebih baik dibandingkan dengan variasi A (Heck et al. 2009).
Penanda Molekuler
Penanda molekuler merupakan pemanfaatan dari keragaman pada tingkat
deoxyribonucleic acid (DNA). Penanda molekuler memiliki peranan penting dalam genetika ternak. Hal tersebut merupakan salah satu faktor utama yang mendasari proses seleksi (Vignal et al. 2002). Tipe dasar perubahan dalam DNA meliputi subtitusi, delesi, insersi, dan inversi (Nei dan Kumar 2000).
9
Single nucleotide polymorphism (SNP) merupakan penanda yang memiliki perbedaan satu nukleotida dalam sekuen DNA (Vignal et al. 2002) frekuensi mutasi dan stabilitas yang tinggi menyebabkan SNP sering digunakan sebagai penanda molekuler dalam penelitian tentang genetika populasi dan pemetaan gen untuk penyakit kompleks (Ye et al. 2001). Metode umum yang digunakan dalam menganalisis adanya SNP antara lain PCR-RFLP dan PCR-SSCP.
Polymerase chain reaction (PCR) merupakan suatu teknik untuk menggandakan jumlah molekul DNA pada ruas-ruas tertentu dan monomer-monomer nukleotida yang dilakukan secara in vitro. Proses ini berjalan dengan bantuan primer dan enzim polymerase. Primer merupakan oligonukleotida spesifik pada DNA template. Enzim polymerase merupakan enzim yang dapat mencetak urutan DNA baru. Hasil dari proses PCR dapat divisualisasikan dengan elektroforesis (Williams 2005). Muladno (2002) menyatakan bahwa secara umum, reaksi yang terjadi dalam mesin PCR dapat dibagi menjadi tiga tahap, yaitu tahap denaturasi (pemisahan untai ganda DNA), tahap annealing (penempelan primer), dan tahap extension (pemanjangan primer).
Polymerase chain reaction-restriction fragment length polymorphisms atau RFLP merupakan metode analisis lanjutan dari produk PCR. Metode PCR-RFLP memanfaatkan perbedaan pola pemotongan enzim pemotong yang berbeda-beda pada tiap-tiap mikroorganisme. Analisis RFLP sering digunakan untuk mendeteksi lokasi genetik dalam kromosom yang menyandikan penyakit yang diturunkan (Orita et al. 1989), ataupun untuk mendeteksi adanya keragaman pada gen yang berhubungan dengan sifat ekonomis, seperti produksi dan kualitas susu. Metode PCR-RFLP memvisualisasikan perbedaan pada level DNA yang didasarkan pada penggunaan enzim pemotong yang dapat memotong DNA pada tempat sekuens nukleotida spesifik (Montaldo dan Herrera 1998). Li dan Graur (1991) menyatakan bahwa enzim pemotong yang dapat mengenal sekuens DNA spesifik disebut recognition sequences dan biasanya memiliki panjang empat sekuens basa atau lebih dan bersifat palindrome.
Metode PCR-RFLP biasa digunakan untuk mencari keragaman genotipe (Yahyaoui et al. 2001). Penyisipan (insersi), penghilangan (delesi), maupun subtitusi nukleotida yang terjadi pada daerah rekognisi suatu enzim restriksi menyebabkannya tidak lagi dikenalinya situs pemotongan enzim restriksi dan terjadinya menyebabkannya perbedaan pola pemotongan DNA (Lewin 1994). Teknik ini digunakan secara luas untuk mendeteksi variasi DNA baik pada daerah yang bersifat penyandi (coding region) pada genom maupun pada daerah yang tidak penyandi atau daerah non-coding (Vasconcellos et al. 2003).
10
Keragaman Genetik
Identifikasi keragaman genetik dalam suatu populasi digunakan untuk mengetahui dan melestarikan bangsa-bangsa dalam populasi terkait dengan penciri suatu sifat khusus. Informasi keragaman genetik suatu bangsa akan sangat bermanfaat bagi keamanan dan ketersediaan bahan pangan yang berkesinambungan (Blott et al. 1998). Nei dan Kumar (2000) menyatakan bahwa populasi dinilai beragam jika memiliki dua atau lebih alel dalam satu lokus dengan frekuensi yang cukup (biasanya lebih dari 1%). Hukum Hardy-Weinberg menyatakan bahwa frekuensi genotipe suatu populasi yang cukup besar tidak akan berubah dari satu generasi ke generasi lainnya jika tidak ada seleksi, migrasi, mutasi, dan genetic drift. Keadaan populasi yang demikian disebut dalam keadaan equilibrium (seimbang) (Noor 2008). Selain itu silang dalam dan silang luar juga dapat mempengaruhi frekuensi genotipe. Keragaman genetik dapat digunakan sebagai parameter dalam mempelajari genetika populasi dan genetika evolusi. Tingkat keragaman dalam populasi dapat digambarkan dari frekuensi alel. Frekuensi alel merupakan rasio relatif suatu alel terhadap keseluruhan alel yang ditemukan dalam satu populasi (Nei dan Kumar 2000).
Derajat heterozigositas, menurut Nei (1987), merupakan rataan persentase lokus heterozigositas tiap individu atau rataan persentase individu heterozigot dalam populasi. Suatu alel dikatakan polimorfik jika memiliki frekuensi alel sama dengan atau kurang dari 0.99. Hartl dan Clark (1997) menyatakan bahwa polimorfisme genetik dalam suatu populasi dapat digunakan dalam menentukan hubungan antar subpopulasi yang terfragmentasi dalam suatu spesies. Perhitungan keragaman genetik dalam populasi secara kuantitatif dapat diperoleh melalui dua ukuran keragaman variasi populasi yaitu proporsi lokus polimorfisme dalam populasi dan rata-rata proporsi individu heterozigot dalam setiap lokus (Nei dan Kumar 2000). Keragaman genetik antara subpopulasi dapat diketahui dengan melihat persamaan dan perbedaan frekuensi alel di antara subpopulasi (Li et al. 2000). Javanmard et al. (2005) menyatakan bahwa nilai heterozigositas di bawah 0.5 (50%) mengindikasikan rendahnya variasi suatu gen dalam populasi dan jika nilai Ho (heterozigositas pengamatan) lebih rendah dari He (heterozigositas harapan) maka dapat mengindikasikan adanya proses seleksi yang intensif (Tambasco et al. 2003). Avise (1994) juga menyatakan bahwa semakin tinggi derajat heterozigositas suatu populasi maka daya tahan hidup populasi tersebut akan semakin tinggi. Seiring dengan menurunnya derajat heterozigositas akibat dari silang dalam dan fragmentasi populasi, sebagian besar alel resesif yang bersifat letal semakin meningkat frekuensinya.
3 METODE
Sebagian data penelitian berasal dari kegiatan Kerjasama Kemitraan Penelitian dan Pengembangan Pertanian Nasional (KKP3N) nomor kontrak 720/LB.620/I.1/2/2013 tanggal 25 Februari 2013 dengan judul “Pengembangan Teknologi Pengolahan Susu Kerbau Di Sumatera Utara Menjadi Produk Keju
11 Lokasi dan Waktu
Penelitian ini dilaksanakan di Laboratorium Genetika Molekuler Ternak (LGMT) Bagian Pemuliaan dan Genetika dan Laboratorium Pengolahan Susu Fakultas Peternakan, Institut Pertanian Bogor. Penelitian dilakukan pada bulan Mei hingga April 2014.
Materi Sampel Darah dan Susu
Total sampel darah kerbau yang digunakan sebanyak 290 dengan jenis kerbau sungai berasal dari daerah Sumatera Utara (40 ekor); dan kerbau rawa sebanyak 250 ekor yang berasal dari Banten (30 ekor), Semarang (30 ekor), Mataram (30 ekor), Sulawesi Selatan (30 ekor), Aceh (67 ekor), Riau (25 ekor), Sumatera Utara (24 ekor), dan Sumatera Barat (14 ekor).
Sampel susu yang digunakan untuk pengujian kualitas susu merupakan hasil pemerahan pada pagi hari. Total sebanyak 40 sampel yang terdiri dari 13 sampel susu kerbau sungai yang berasal dari BPTU Siborong-borong dan Deli Serdang, serta 27 sampel susu kerbau rawa yang berasal dari BPTU Siborong-borong, Deli Serdang, Pematang Panjang, Batu sangkar, dan Kampar. Lokasi peternakan kerbau di Deli Serdang, Pematang Panjang, Batu Sangkar, dan Kampar merupakan peternakan rakyat, sedangkan lokasi BPTU Siborong-borong merupakan balai perbibitan babi dan kerbau milik pemerintah.
Primer
Primer gen κ-Kasein yang digunakan dalam penelitian ini dimodifikasi menggunakan teknik PIRA-PCR dengan runutan primer κ-Kasein : forward 5’
GTT GAG CCT ACA AGT ACA gaT A 3’, dan reverse5’ TGT CTT CTT TGA
TGT CTC CTT AGA G 3’, nomor akses GenBank AM900443, Bubalus bubalis
κ-Casein gene, exon 1-5 (Masina et al. 2007).
Gambar 4 Posisi penempelan primer pada sekuen gen κ-Kasein sebagian ruas ekson 4
Prosedur Analisis Penilaian Kualitas Susu Kerbau
Penilaian kualitas susu meliputi persentase lemak, BKTL (Bahan Kering Tanpa Lemak), protein, laktosa, dan mineral. Prosedur pengukuran parameter tersebut menggunakan alat milkotester. Volume sampel yang digunakan sebanyak ±40 ml.
Forward
12001 taccatcaat accattgtta gtgttgagcc tacaagtaca(ga)tatcactg aagcaataga
12061 gaacactgta gctactctag aagcttcctc agaagttatt gagagtgtac ctgagaccaa
12121 cacagcccaa gttacttcaa ccgtcgtcta aaaactctaa ggagacatca aagaagacaa
12
Ekstraksi Sampel Darah dan Susu
Sampel darah diambil sebanyak 5 mL melalui vena jugularis di leher kerbau menggunakan jarum vennoject dan tabung vaccutainer mengandung EDTA. Selanjutnya sampel darah ditambahkan alkohol (1:2) dikocok dan disimpan pada suhu ruang sampai siap untuk diekstraksi DNA; sedangkan untuk sampel susu diambil sebanyak 40 mL dan segera disimpan pada suhu refri untuk segera dianalisa.
Ekstraksi DNA dilakukan dari darah dan susu kerbau menggunakan prosedur ekstraksi mengikuti metode phenol-chloroform (Sambrook et al. 1989) yang telah dimodifikasi oleh Andreas et al. (2010):
Preparasi Sampel. Sampel sebanyak 200 µL dimasukkan ke dalam tabung 1.5 mL. Sampel kemudian ditambahkan air destilasi sampai 1000 µ L. Sampel diendapkan dengan sentrifugasi pada kecepatan 8000 rpm selama lima menit, supernatan dibuang.
Degradasi Protein. Sampel yang ditambahkan 1xSTE (sodium tris EDTA) sampai volume 350 µ L, 40 µ L SDS 10% dan 10 µ L proteinase K 5 mg/mL. Campuran diinkubasi pada suhu 55 ˚C selama dua jam digoyang pelan.
Degradasi Bahan Organik. Ditambahkan 400 µ L larutan phenol, 400 µ L
chloroform:isoamyl alcohol (24:1) dan 40 µ L NaCl 5M. Campuran digoyang pada suhu ruang selama satu jam.
Presipitasi DNA. Sampel disentrifugasi pada kecepatan 12000 rpm selama lima menit hingga fase air terpisah dengan fase phenol. Fase air dipindahkan ke tabung baru sebanyak 400 µ L. Molekul DNA diendapkan dengan menambahkan 800 µ L alkohol absolut dan 40 µ L NaCl 5M. Campuran diinkubasi pada suhu -20 ˚C semalaman. Pengendapan DNA dilakukan dengan sentrifugasi pada kecepatan 12000 rpm selama lima menit. Endapan yang diperoleh dicuci dengan 800 µ L alkohol 70%, kemudian diendapkan lagi dan dikeringkan dalam keadaan terbuka sampai alkohol hilang. Endapan DNA yang telah bersih dari alkohol dipulihkan kembali dengan menambahkan 100 µL TE 80% (Elution Buffer). Sampel DNA disimpan pada suhu -20 ˚C dan siap untuk digunakan.
Amplifikasi Ruas Gen κ-Kasein
Amplifikasi ruas gen κ-Kasein dilakukan dengan metode PCR (Polymerase Chain Reaction). Pereaksi yang digunakan untuk amplifikasi gen κ-Kasein merupakan campuran yang terdiri atas sampel DNA 0.5 µL, destilated water 5.85 µL, primer 0.3 μL, Taq phyre 0.05 µ L, buffer 2x PCR (phyre) 7.5 µ L. Campuran tersebut kemudian diinkubasi dalam mesin thermocycler dengan kodisi suhu pradenaturasi 95 °C selama lima menit, denaturasi 95 °C selama 20 detik, anneling60 °C selama 30 detik, ekstensi awal72 °C selama 40 detik, dan ekstensi akhir 72 °C selama 5 menit. Siklus PCR yang digunakan sebanyak 35 siklus. Analisis PCR-RFLP
Enzim pemotong yang digunakan untuk ruas gen target adalah EcoRV atau
Eco123. Produk PCR sebanyak 4 µ L dipindahkan ke tabung 0.5 mL, ditambahkan
13 pada suhu 37 °C selama 16 jam. Enzim EcoRV mengenali situs pemotongan sekuen GAT|ATC.
Elektroforesis dan Pendeteksian Keragaman DNA
Penentuan genotipe masing-masing individu dilakukan dengan pendekatan
Restriction Fragment Length Polymorphism (RFLP) yang divisualisasikan pada gel agarosa 2% dengan TBE 0.5 x (Tris Borat EDTA) pada 100 volt selama 40 menit. Gel diwarnai dengan 2.5 µ L bromida yang divisualisasikan pada UV transiluminator.
Setelah elektroforesis dengan gel agarose, akan muncul pita-pita DNA. Setiap pita DNA yang muncul dibandingkan dengan marker untuk diketahui panjang fragmennya. Setiap pita DNA dari setiap sampel dibandingkan untuk menentukan genotip pita DNA. Satu posisi migrasi yang sama dianggap sebagai satu tipe atau alel.
Prosedur Analisis Data
Analisis data dibagi menjadi dua bagian, yaitu analisis keragaman gen κ -Kasein dan analisis kualitas susu. Analisis keragaman genetik dilakukan dengan menghitung (1) frekuensi alel dan genotipe, (2) heterozigositas, (3) Polymorphic Informative Content (PIC), dan (4) kesamaan sekuens gen κ-Kasein (homology). Hubungan antara genotipe gen κ-Kasein dengan sifat kualitas susu dianalisa secara deskriptif. Peubah yang diamati pada analisa kualitas susu : Lemak, BKTL, Protein, Laktosa, dan Mineral.
Frekuensi Genotipe dan Frekuensi Alel
Frekuensi genotipe merupakan rasio dari jumlah suatu genotipe terhadap jumlah populasi. Keragaman genotipe pada masing-masing individu ternak dapat ditentukan melalui pita-pita DNA yang ditemukan. Frekuensi genotipe dapat diketahui dengan menghitung perbandingan jumlah genotipe tertentu pada setiap populasi, dengan rumus Nei dan Kumar (2000) sebagai berikut:
Frekuensi alel merupakan rasio suatu alel terhadap keseluruhan alel pada suatu lokus dalam populasi. Frekuensi alel ( ) gen κ-Kasein|EcoRV dapat dihitung berdasarkan rumus Nei dan Kumar (2000), sebagai berikut:
∑
Keterangan:
= frekuensi genotipe ke-ii
= frekuensi alel ke-i
nii = jumlah individu bergenotipe ii
nij = jumlah individu bergenotipe ij
14
Heterozigositas
Keragaman genetik dapat diketahui melalui estimasi frekuensi heterozigositas pengamatan yang diperoleh dari masing-masing lokasi, dengan menggunakan rumus Weir (1996) sebagai berikut:
∑
Keterangan:
Ho = heterozigositas pengamatan nij = jumlah individu heterozigot
N = jumlah individu yang diamati
Heterozigositas harapan (He) berdasarkan frekuensi alel dihitung menggunakan rumus Nei dan Kumar (2000) sebagai berikut:
∑
Keterangan:
He = nilai heterozigositas harapan
= frekuensi alel q = jumlah alel
Polimorphic Informative Content (PIC)
Tingkat informative suatu alel dihitung menggunakan pendekatan nilai Polymorphic Informative Content (PIC) (Bostein et al. 1980):
∑
∑
∑
Keterangan:
Pi = frekuensi alel ke-i n = jumlah alel
Perunutan Susunan Nukleotida
Pasangan primer yang digunakan untuk analisa sekuensing menggunakan runutan dari GenBank nomor akses AM900443 (Masina et al. 2007) yang tidak dimodikasi. Hasil sekuens fragmen gen κ-Kasein|EcoRV pada kerbau sungai dan rawa dianalisis kesamaannya (homologi) dengan sekuens yang terdapat di
15 Hubungan Keragaman Gen κ-Kasein Terhadap Kualitas Susu
Analisis kualitas susu menggunakan milkotester. Data karakteristik parameter susu dianalisis untuk mendapatkan nilai rataan dan simpangan baku pada masing-masing populasi. Pengaruh keragaman gen κ-Kasein terhadap parameter kualitas susu dianalisa secara deskriptif.
4
HASIL DAN PEMBAHASAN
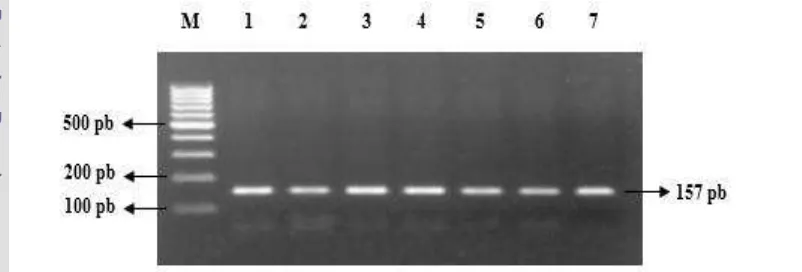
Amplifikasi Gen κ-Kasein
Amplifikasi fragmen gen κ-Kasein ekson 4 pada kerbau menghasilkan panjang produk PCR sebesar 157 pb (Gambar 5). Amplifikasi fragmen gen κ -Kasein dilakukan pada suhu annealing 60 ºC selama 30 detik. Pelt-Verkuil et al.
(2008) menyatakan bahwa waktu annealing yang dibutuhkan primer dapat berkomplemen dan menempel dengan target bergantung pada kapasitas pemanasan mesin thermocycler yang digunakan, volume campuran PCR serta konsentrasi primer dan gen target.
Keragaman Gen κ-Kasein dengan Metode PCR RFLP
Identifikasi keragaman gen κ-Kasein pada kerbau sungai dan rawa dilakukan dengan mengidentifikasi ada tidaknya situs pemotongan oleh enzim
EcoRV, dimana sekuens target telah diamplifikasi menggunakan pasangan primer yang telah dimodifikasi menggunakan teknik PIRA (Ke et al. 2001), sehingga jika terdapat pemotongan hal ini mengindikasikan bahwa gen κ-Kasein pada kerbau tersebut memiliki situs enzim EcoRV yaitu GAT|ATC.
Keragaman gen κ-Kasein|EcoRV pada kerbau sungai ditemukan pada ekson 4, yaitu pada sekuen ke-12043 pb atau pada posisi basa ke-21 produk PCR. Hasil pemotongan enzim EcoRV menghasilkan tiga fragmen dengan panjang berbeda yaitu 157 pb (alel C), 136 dan 21 pb (alel T). Dengan demikian, suatu individu dikatakan memiliki genotipe CC apabila proses restriksi menghasilkan satu fragmen pita (157 pb), genotipe CT dengan tiga pita (157, 136, dan 21 pb), dan genotipe TT dengan dua pita (136 dan 21 pb) yang ditampilkan pada Gambar 6.
16
Hasil tersebut membuktikan bahwa teknik PIRA efektif digunakan dalam mengidentifikasi keragaman gen κ-Kasein pada kerbau. Mitra et al. (1998) melaporkan bahwa gen κ-Kasein|HinfI pada kerbau Murrah dan Nili-Ravi di India memiliki panjang produk PCR 379 pb. Panjang produk PCR beberapa penelitian gen κ-Kasein pada kerbau ditemukan bervariasi, antara lain ditemukan produk PCR sepanjang 453 pb (Abbasi et al. 2009), 221 pb (Beneduci et al. 2010), 350 pb (Gangaraj et al. 2008). Perbedaan panjang fragmen disebabkan primer yang digunakan berbeda. Berbeda dengan identifikasi keragaman gen κ-Kasein pada beberapa penelitian yang sudah dipublikasi menggunakan penamaan alel A dan B, pada penelitian ini menggunakan penamaan alel C dan T berdasarkan perubahan basa yang terjadi namun memiliki situs perubahan basa yang sama (alel A dan C memiliki asam amino ACC/Threonin; sedangkan alel B dan T memiliki asam amino ATC/isoleusin).
Masina et al. (2007) dengan studinya menggunakan kerbau sungai melaporkan bahwa pada ekson 1, 2, 3, dan 5 tidak ditemukan keragaman. Pada ekson 4, ditemukan dua keragaman basa nukleotida (T ke C pada posisi 135 yang merubah asam amino dan silent mutation T ke C pada posisi 136).
Gen κ-Kasein|EcoRVpada kerbau rawa tidak ditemukan variasi alel. Alel yang muncul hanya alel C yang menunjukkan hanya terdapat genotipe CC. Tidak adanya variasi gen κ-Kasein pada kerbau juga ditemukan pada beberapa penelitian yang dilakukan oleh Gangaraj et al. (2008), El-Rafey dan Darwish (2008) serta Otaviano et al. (2005) juga menemukan hanya genotipe BB saja yang muncul (monomorfik) dengan teknik PCR-RFLP menggunakan enzim Hinf, HindIII, dan
AluI.
Frekuensi Alel dan Frekuensi Genotipe Gen κ-Kasein
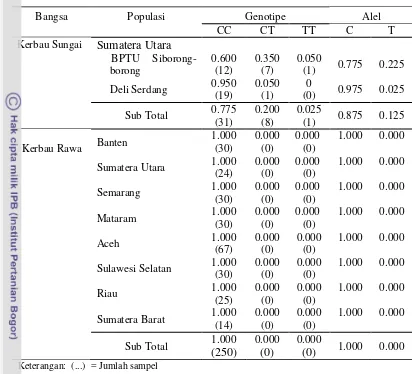
Perubahan genetik pada sebuah populasi biasanya digambarkan dengan adanya perubahan pada frekuensi alel (Nei dan Kumar, 2000). Frekuensi genotipe dan alel gen κ-Kasein pada kerbau sungai dan rawa ditampilkan pada Tabel 3.
17
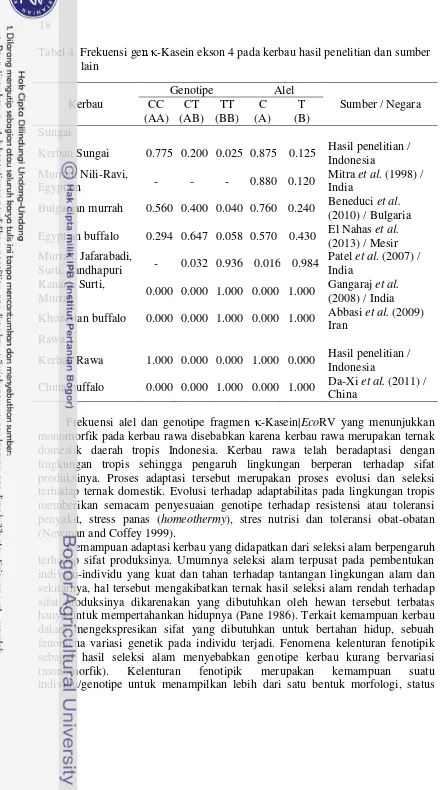
Frekuensi genotipe homozigot CC menunjukkan nilai tertinggi pada seluruh lokasi, dibandingkan dengan genotipe CT dan TT. Frekuensi genotipe CC ditemukan tinggi pada daerah Deli Serdang (0.950), frekuensi genotipe CT tertinggi ditemukan pada daerah BPTU Siborong-borong (0.350), sedangkan genotipe TT hanya ditemukan pada satu ekor kerbau sungai di BPTU Siborong-borong (0.050). Frekuensi alel C dan T kerbau sungai adalah 0.875 dan 0.125. Adanya variasi alel menunjukkan gen κ-Kasein pada kerbau sungai polimorfik. Suatu alel dapat dikatakan bersifat polimorfik apabila frekuensi alel sama dengan atau kurang dari 0.99 (Nei 1987). Berbeda dengan kerbau sungai, pada kerbau rawa hanya ditemukan genotipe CC (100%) dengan nilai frekuensi genotipe masing-masing tempat adalah sama (CC = 1, CT = 0, dan TT = 0), hal tersebut mengarah kepada terjadinya fiksasi alel C (1.00) atau dikenal dengan alel monomorfik. Polimorfisme genetik fragmen gen κ-Kasein ekson 4 yang dilakukan beberapa peneliti ditampilkan pada Tabel 4, yang menunjukkan bahwa keragaman pada kerbau adalah rendah. Kejadian monomorfisme juga ditemukan terjadi pada kerbau sungai di India dan Iran serta kerbau rawa di China.
Tabel 3 Frekuensi gen κ-Kasein|EcoRV pada kerbau sungai dan rawa
Bangsa Populasi Genotipe Alel
18
Tabel 4 Frekuensi gen κ-Kasein ekson 4 pada kerbau hasil penelitian dan sumber lain
Frekuensi alel dan genotipe fragmen κ-Kasein|EcoRV yang menunjukkan monomorfik pada kerbau rawa disebabkan karena kerbau rawa merupakan ternak domestik daerah tropis Indonesia. Kerbau rawa telah beradaptasi dengan lingkungan tropis sehingga pengaruh lingkungan berperan terhadap sifat produksinya. Proses adaptasi tersebut merupakan proses evolusi dan seleksi terhadap ternak domestik. Evolusi terhadap adaptabilitas pada lingkungan tropis memberikan semacam penyesuaian genotipe terhadap resistensi atau toleransi penyakit, stress panas (homeothermy), stres nutrisi dan toleransi obat-obatan (Newman and Coffey 1999).
19 fisiologi dan tingkah laku sebagai respon terhadap perubahan lingkungan (Noor 2008).
Derajat Polimorfisme Gen κ-Kasein|EcoRV
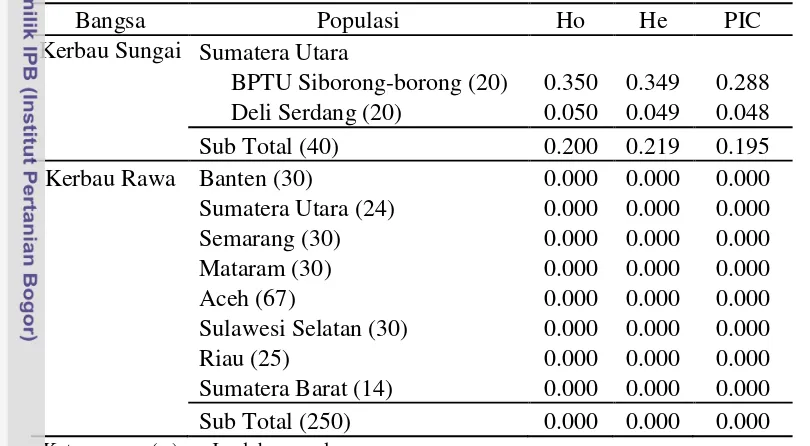
Pendugaan nilai heterozigositas pengamatan (Ho), heterozigositas harapan (He), dan polymorphic informative content (PIC) fragmen gen κ-Kasein|EcoRV pada kerbau sungai dan rawa ditampilkan pada Tabel 5. Nilai heterozigositas pengamatan (Ho) fragmen κ-Kasein|EcoRV menunjukkan bahwa kerbau sungai di BPTU Siborong-borong memiliki nilai heterozogositas tertinggi (0.350) dibandingkan dengan Deli Serdang (0.050), sedangkan nilai heterozigositas kerbau rawa pada seluruh populasi adalah 0.000.
Nilai heterozigositas harapan (He) fragmen κ-Kasein|EcoRV menunjukkan tidak jauh berbeda dengan nilai heterozigositas pengamatan, dimana kerbau sungai di BPTU Siborong-borong memilki nilai heterozigositas tertinggi (0.349) dibandingkan dengan Deli Serdang (0.049), sedangkan nilai heterozigositas kerbau rawa pada seluruh populasi adalah 0.000. Nilai heterozigositas pengamatan dan harapan didapatkan rendah (<0.5) dan tidak jauh berbeda, hal tersebut kemungkinan disebabkan belum adanya seleksi intensif terhadap kualitas susu berdasarkan gen κ-Kasein.
Nei (1987) menyatakan bahwa salah satu cara mengetahui nilai variasi genetik adalah dengan menghitung nilai heterozigositas. Javanmard et al. (2005) menyatakan bahwa nilai heterozigositas di bawah 0.5 (50%) menunjukkan rendahnya variasi suatu gen dalam populasi. Keragaman genetik yang sangat rendah pada kerbau disebabkan kemampuan adaptasi yang didapatkan dari seleksi alam daerah tropis. Kemampuan hidup dalam kondisi yang terbatas (nutrisi dan iklim) memberikan dampak kerbau harus mengekspresikan sifat yang sesuai dengan kebutuhannya dalam mempertahankan hidup.
Tabel 5 Nilai Heterozigositas pengamatan (Ho), harapan (He) dan PIC
Bangsa Populasi Ho He PIC
Kerbau Sungai Sumatera Utara
BPTU Siborong-borong (20) 0.350 0.349 0.288
Deli Serdang (20) 0.050 0.049 0.048
Sub Total (40) 0.200 0.219 0.195
Kerbau Rawa Banten (30) 0.000 0.000 0.000
Sumatera Utara (24) 0.000 0.000 0.000
Semarang (30) 0.000 0.000 0.000
Mataram (30) 0.000 0.000 0.000
Aceh (67) 0.000 0.000 0.000
Sulawesi Selatan (30) 0.000 0.000 0.000
Riau (25) 0.000 0.000 0.000
Sumatera Barat (14) 0.000 0.000 0.000
Sub Total (250) 0.000 0.000 0.000
20
Pendugaan nilai PIC dilakukan juga untuk mengetahui tingkat keragaman suatu penciri genetik. PIC memiliki kisaran nilai 0-1, dimana nilai PIC=0 apabila hanya ditemukan satu alel, sedangkan PIC=1 apabila terdapat jumlah alel yang tak terhingga. Apabila satu gen hanya memiliki dua alel maka akan dihasilkan nilai PIC maksimum sebesar 0.375 (Hildebrand et al. 1992). Tingginya nilai PIC menunjukkan suatu penciri genetik memiliki keragaman yang tinggi, sedangkan jika nilai PIC rendah menunjukkan suatu penciri genetik memiliki tingkat keragaman yang rendah (homozigot). PIC menggambarkan nilai heterozigositas yang terkoreksi oleh informasi perkawinan sebagian (partially informative mating), sehingga nilai PIC selalu memiliki nilai yang lebih rendah dibandingkan dengan nilai heterozigositas (Hildebrand et al. 1992). Tabel 5 menunjukkan secara umum, nilai PIC kerbau sungai dan rawa di seluruh populasi adalah rendah. Nilai PIC kerbau sungai tertinggi ditemukan di BPTU Siborong-borong (PIC = 0.288) dibandingkan di Deli Serdang (PIC = 0.048). Pada kerbau rawa diseluruh populasi nilai PIC=0 karena gen κ-Kasein|EcoRV bersifat monomorfik hanya ditemukan satu alel, yaitu alel C.
Runutan DNA Ruas Gen κ-Kasein
Perunutan nukleotida dilakukan pada ruas sebagian ekson 4 gen κ-Kasein kerbau sungai dengan genotipe yang berbeda, yaitu genotipe homozigot CC dan TT. Analisis kesamaan runutan DNA hasil sekuensing dengan runutan nukleotida gen κ-Kasein pada sapi dan kerbau yang ada di GenBank dilakukan dengan pendekatan BLAST (basic local alignment search tool) (http://blast.ncbi.nlm.nih.gov/Blast.cgi), dan ditampilkan pada Gambar 7.
21 Kesamaan sekuen fragmen gen κ-Kasein memberikan keyakinan bahwa fragmen gen κ-Kasein|EcoRV yang diamplifikasi pada bangsa kerbau yang diteliti merupakan fragmen gen κ-Kasein. Selain untuk mendapatkan nilai kesamaan gen κ-Kasein, analisis sekuensing fragmen gen κ-Kasein dilakukan untuk mendapatkan gambaran mutasi atau perubahan basa yang terjadi pada lokus
EcoRV. Runutan nukleotida gen κ-Kasein dalam penelitian ini memiliki tingkat kesamaan yang tinggi dengan kerbau yang ada pada GenBank, yaitu mencapai 98-100% sedangkan pada sapi (GenBank AY380229 dan AY380228) hanya 90-91%. Berdasarkan hasil analisis sekuensing ditemukan 2 mutasi atau perubahan basa nukleotida pada fragmen gen κ-Kasein|EcoRV, yakni basa C (Citosin) menjadi T (Timin) pada posisi ke-23 pb dan ke-27 pb produk PCR. Hanya perubahan basa ke-23 pb dari produk PCR yang dikenali oleh enzim restriksi EcoRV.
Situs mutasi pada posisi ke-23 pb dan 27 pb produk PCR atau pada posisi 12045 pb dan 12049 pb sesuai sekuen gen κ-Kasein di GenBank (kode akses AM900443) yang ditemukan pada penelitian ini sesuai dengan yang ditemukan pada Masina et al. (2007) dan Beneduci et al. (2010) yang melaporkan bahwa terjadi perubahan basa C menjadi T pada kodon 135 sehingga terjadi perubahan asam amino Threonin (ACC) menjadi Isoleusin (ATC), dan silent mutation pada kodon 136 merubah basa C menjadi T tanpa mengubah asam amino Threonin (ACC) menjadi Threonin (ACT).
Berdasarkan tipe mutasi yang terdapat pada penciri PCR-RFLP fragmen gen κ-Kasein|EcoRV, mutasi tersebut termasuk dalam tipe mutasi transisi. Sebagaimana dinyatakan Li dan Graur (1991), mutasi tipe transisi yaitu pertukaran atau subtitusi antara basa purin (A = Adenin, G = Guanin) atau basa pirimidin (C = Citosin, T = Timin).
Polimorfisme Gen κ-Kasein|EcoRV dengan Kualitas Susu
Pemeriksaan produksi dan kualitas susu kerbau dilakukan untuk setiap kondisi pemeliharaan berbeda namun dengan kondisi agroekosistem yang tidak jauh berbeda. Data produksi susu merupakan data rataan hasil pemerahan seluruh kerbau setiap lokasi selama satu hari dengan kisaran periode laktasi ketiga sampai keempat. Pemeriksaan produksi susu serta pengaruh varian genotipe κ-Kasein terhadap protein dan kualitas susu lainnya ditampilkan pada Tabel 6.
Produksi susu kerbau sungai di lokasi Deli Serdang (6-10 liter ekor-1 hari-1) lebih tinggi dibandingkan BPTU Siborong-borong (4-6 liter ekor-1 hari-1). Hal tersebut kemungkinan dipengaruhi oleh kondisi laktasi kerbau di Deli Serdang yang memasuki laktasi ke-4 dan manajemen pemberian pakan. De Roos et al. (2004), melaporkan bahwa kapasitas produksi susu sapi perah akan meningkat terus menerus dan mencapai puncak produksi pada periode laktasi ketiga atau keempat kemudian turun berlahan-lahan.
22
brachiaria decumbens) dan pakan konsentrat sebanyak 3-4 kg hari-1 diberikan 2 kali (pagi dan sore). Tyler dan Ensminger (2006) menyatakan bahwa frekuensi pemberian pakan yang lebih dari dua kali akan dapat meningkatkan konsumsi bahan kering pakan, kadar lemak susu, dan produksi susu. Kecukupan pakan secara kuantitas dan kualitas menentukan optimalitas produksi susu. Adanya perbedaan jumlah produksi susu diduga karena perbedaan periode laktasi dan manajemen pemeliharaan.
Tabel 6 Karakteristik kualitas susu kerbau sungai dengan genotipe berbeda
Lokasi Produksi Susu Kualitas
Susu (%) Genotipe
BPTU Siborong-borong CC (4) CT (0) TT (1)
Laktasi ke-3 4-6 liter Lemak 7.758 ± 0.247 0 6.94 BKTL 11.333 ± 0.186 0 10.61 Protein 5.063 ± 0.094 0 4.725 Laktosa 4.539 ± 0.086 0 4.575 Mineral 0.880 ± 0.025 0 0.87
Deli Serdang CC (7) CT (1) TT (0)
Laktasi ke-4 6-10 liter Lemak 7.350 ± 0.876 7.435 0 BKTL 11.064 ± 0.625 10.915 0 Protein 4.936 ± 0.271 4.875 0 Laktosa 5.257 ± 0.296 5.185 0 Mineral 0.859 ± 0.067 0.84 0
Keterangan: (...) = jumlah sampel; BKTL = Bahan Kering Tanpa Lemak; Genotipe CT dan TT tidak dimasukkan dalam uji analisis hubungan antara genotipe dengan fenotipe kualitas susu
Parameter kualitas susu yang dikelompokkan berdasarkan genotipe gen κ -Kasein (Tabel 6) ditemukan tidak jauh berbeda pada parameter kadar lemak, BKTL, protein, laktosa, dan mineral pada lokasi di BPTU Siborong-borong dan Deli Serdang.
Kadar Protein Susu
Kadar protein susu berdasarkan varian genotipe gen κ-Kasein di masing-masing lokasi (Tabel 6) memperlihatkan nilai rataan protein susu kerbau genotipe CC, CT dan TT tidak jauh berbeda. Namun, terdapat kecenderungan kerbau genotipe CC atau alel C (ACC) berpengaruh terhadap kadar protein yang lebih tinggi. Beberapa penelitian keragaman gen κ-Kasein pada kerbau menyatakan bahwa alel B berpengaruh terhadap kualitas protein dan koagulasi rennet yang lebih baik, namun pada hasil penelitiannya tidak ditemukan keragaman sehingga tidak dapat diasosiasikan (Gangaraj et al. 2008; Abbasi et al. 2009). Hal tersebut dikarenakan kebanyakan peneliti mengacu pada genotipe BB gen κ-Kasein sapi yang terbukti berpengaruh terhadap protein dan produksi susu (Caroli et al. 2004; Azevedo et al. 2008). Terdapat perbedaan signifikan pada sekuens gen κ-Kasein antara kerbau (Bubalus bubalis) dengan sapi (Bos taurus) (Gambar 7).
23 rendah sehingga asosiasi antara keragaman gen dengan sifat kualitas susu sulit dilakukan. Produksi susu merupakan sifat kuantitatif yang dikendalikan banyak gen, dan diwariskan serta dipengaruhi oleh faktor lingkungan (Noor 2008). Roginski (2003) menyatakan bahwa protein susu dikontrol oleh gen kasein komplek yang terdiri atas empat jenis, yaitu αS1-Kasein (39-46% dari total
kasein), β-Kasein (25-35% dari total kasein), αS2-Kasein (8-11% dari total kasein)
dan κ-Kasein (8-15% dari total kasein). Kontrol gen kasein terhadap protein susu sebesar 80%, sedangkan 20% sisanya dikontrol oleh gen pengontrol whey susu yang terdiri atas β-laktoglobulin, α-laktalbumin, serum albumin, glikomakropeptida dan protein antimikrobia yang berupa immunoglobulin, laktoferin, laktoperoksidase dan lisozim (Eigel 1984). Adapun Bonfatti et al.
(2012) menyatakan bahwa pengaruh satu gen aditif tidak berhubungan terhadap sifat produksi susu. Pemeriksaan dua atau lebih gen kasein secara bersamaan bermanfaat bisa memperhitungkan pengaruh interaksi yang mungkin terjadi antara dua atau lebih gen haplotipe (Anggraeni et al. 2012).
Parameter Kualitas Susu yang Lain
Pemeriksaan pengaruh varian genotipe gen κ-Kasein terhadap lemak susu menunjukkan bahwa genotipe yang muncul pada setiap lokasi tidak memiliki konsistensi dalam menghasilkan kadar lemak susu. Hal tersebut menjelaskan bahwa kadar lemak tidak dipengaruhi oleh polimorfisme gen κ-Kasein namun sangat dipengaruhi oleh lingkungan.
Pengamatan pada komponen susu lainnya menunjukkan bahwa genotipe CC memiliki nilai rataan kadar BKTL dan mineral yang lebih tinggi dibandingkan genotipe CT dan TT, serta tidak ditemukan varian genotipe mempengaruhi kadar laktosa susu.
Karakteristik Kualitas Susu Kerbau Sungai dan Rawa
Pemeriksaan parameter kualitas susu kerbau sungai dan rawa dilakukan pada beberapa lokasi di pulau Sumatera. Susu kerbau sungai didapatkan dari provinsi Sumatera Utara (lokasi BPTU Siborong-borong dan Deli Serdang); sedangkan susu kerbau rawa didapatkan dari provinsi Sumatera Barat (lokasi Pematang Panjang dan Batu Sangkar), dan provinsi Riau (lokasi Kampar) yang ditampilkan pada Tabel 7. Data kualitas susu diperoleh dari pengujian kualitas hasil pemerahan selama satu hari pada kisaran bulan laktasi pertama sampai bulan laktasi keenam pada periode laktasi kesatu sampai kelima.
24
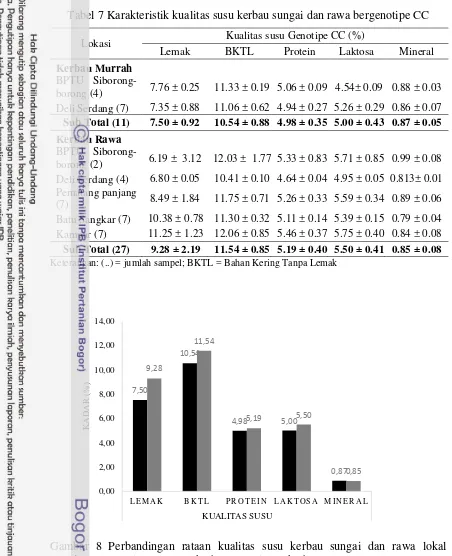
Tabel 7 Karakteristik kualitas susu kerbau sungai dan rawa bergenotipe CC
Lokasi Kualitas susu Genotipe CC (%)
Lemak BKTL Protein Laktosa Mineral
Kerbau Murrah
Keterangan: (..) = jumlah sampel; BKTL = Bahan Kering Tanpa Lemak
Gambar 8 Perbandingan rataan kualitas susu kerbau sungai dan rawa lokal genotipe CC ( = kerbau sungai; = kerbau rawa)
Kadar Lemak
25 kerbau rawa diduga dipengaruhi oleh manajemen pemeliharaan yang berbeda. Kadar lemak kerbau rawa di lokasi Deli Serdang (6.80 ± 0.05%) dan BPTU Siborong-borong (6.19 ± 3.12%) lebih rendah dibanding lokasi Pematang Panjang (8.49 ± 1.84%), Batu Sangkar (10.38 ± 0.78%), dan Kampar (11.25 ± 1.23%). Pemberian pakan berupa rasio konsentrat yang lebih banyak dibandingkan hijauan membuat penurunan kadar lemak susu kerbau rawa di lokasi Deli Serdang BPTU Siborong-borong. Sebaliknya, pada kerbau rawa di lokasi Pematang Panjang, Batu Sangkar, dan Kampar proporsi pemberian hijauan lebih besar dibandingkan konsentrat. Tyler dan Ensminger (2006) menyatakan bahwa tingginya proporsi hijauan dalam pakan ternak perah menyebabkan peningkatan kadar lemak susu yang dihasilkan. Pakan hijauan merupakan sumber serat. Kekurangan pakan serat akan menghasilkan susu dengan kadar lemak rendah. Hal ini disebabkan hijauan dalam rumen merupakan sumber asam asetat yang merupakan bahan baku pembentuk berbagai asam lemak dari lemak susu, yaitu butirat, kaproat, kaprilat, kaprat, laurat, miristat, palmitat, oleat, stearate, dan linoleat. Semakin banyak produksi asetat, semakin banyak sintesis asama lemak yang kemudian menyebabkan peningkatan kadar lemak susu. Kandungan lemak dalam susu adalah komponen terpenting disamping protein yang mempengaruhi harga jual susu (Zurriyati 2011).
Gambar 8 menunjukkan bahwa rataan kadar lemak susu kerbau sungai lebih rendah (7.50%) dibandingkan kerbau rawa (9.28%). Beberapa penelitian yang mengukur kadar lemak susu kerbau sungai dan rawa melaporkan bahwa kadar lemak susu kerbau sungai berkisar pada 3.37-14.42% (Meena et al. 2007), dan 7.07% (Misra et al. 2000); sedangkan kadar lemak susu kerbau rawa berkisar 7.1% Han et al. (2012). Hal tersebut membuktikan bahwa selain manajemen pemeliharaan, bangsa ternak berperan penting dalan komposisi lemak. Ruskin (1981) melaporkan bahwa kadar lemak susu kerbau berkisar 6-8%, namun dapat lebih tinggi pada peternakan kerbau perah dengan manajemen pakan yang baik dan pada kerbau rawa (karena jarang diperah).
Kadar Protein
Tingkat kadar protein susu merupakan karakter yang diturunkan dan tingkat kadarnya hampir selalu konstan (stabil). Rataan kadar protein susu kerbau sungai pada penelitian ini didapatkan sebesar 4.98%, sedangkan pada susu kerbau rawa sebesar 5.19%. Sedikit perbedaan ditemukan oleh Hogberg dan Lind (2003) yang menyatakan bahwa kadar protein susu kerbau sungai sebesar 4.6%; sedangkan pada kerbau rawa sebesar 3.968% (Khan et al. 2007).
Kadar Kualitas Susu yang Lainnya
Rataan kadar BKTL kerbau sungai yang didapatkan pada penelitian adalah sebesar 10.54%, sedangkan pada kerbau rawa didapatkan sebesar 11.54%. Hasil tersebut bersesuaian dengan penelitian Meena et al. (2007) yang menyatakan bahwa BKTL kerbau sungai berkisar pada 5.2-10.59, dan 10.1% (Misra et al.
2000); sedangkan pada kerbau rawa sebesar 9.48% (Khan et al. 2007).