FOOD-RELATED INNOVATIVE BEHAVIORS OF
THE MACAQUES
ISLAMUL HADI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI SERTA
PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa disertas berjudul Food-Related Innovative Behaviors of The Macaques adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Mei 2013
Islamul Hadi
RINGKASAN
ISLAMUL HADI. Food-Related Innovative Behaviors of The Macaques. Dibim-bing oleh BAMBANG SURYOBROTO, KUNIO WATANABE, ACHMAD FA-RAJALLAH.
Perilaku inovatif dalam perilaku baru yang dilakukan oleh individu hewan dan terjadi pada frekuensi rendah dan tidak umum ditemukan pada kondisi normal. Perilaku inovatif memungkinkan hewan dalam menghadapi lingkungan baru dan mengeksploitasi sumber daya lebih luas. Perilaku inovatif telah diamati dan didokumentasikan pada berbagai hewan terutama burung dan primata. Hampir setengah dari perilaku inovatif ini berkaitan dengan perilaku makan termasuk di antaranya teknik ekstraksi makanan, jenis-jenis makanan baru, dan teknik meng-olah makanan.
Pertama, saya mengamati lima kelompok monyet ekor panjang (Macaca fascicularis) di ketinggian Taman Nasioanl Gunung Rinjani tahun 2008-2009. Saya mengamati salah satu kelompok monyet tersebut hidup di sekitar Peman-dian Air Panas Pengkereman yang memiliki suhu rendah dan miskin sumber makanan alami. Beberapa individu monyet di dalam kelompok Pengkereman menunjukkan perilaku mandi air panas. Perilaku tersebut merupakan perilaku ino-vasi yang baru di dalam kelompok tersebut. Perilaku ini berkembang sebagai tanggapan terhadap kondisi habitat mereka. Mereka mengambil sisa makanan dari manusia yang terendam di air panas. Mereka rasakan suhu yang lebih hangat saat mereka masuk ke air panas. Mereka mampu mengingat dan mengulangi perilaku ini karena monyet-monyet ini belajar dan mendapatkan keuntungan nutrisi dan termoregulasi dari lingkugan air panas.
Kedua, saya mengamati perilaku makan ikan yang dilakukan oleh bebe-rapa individu monyet ekor panjang di Taman Wisata Alam Pangandaran tahun 2010-2012. Perilaku ini pertama kali teramati di Kelompok Rengganis pada tahun 2010. Tiga monyet betina dewasa dan satu monyet betina muda makan ikan yang mereka dapatkan dari jaring yang tertinggal di perahu di Pantai Rengganis. Saat ini perilaku makan ikan hanya dilakukan oleh sejumlah individu monyet Kelom-pok Goa Parat dan K12. Perilaku makan ikan ini merupakan perilaku baru yang dilakukan oleh sejumlah individu monyet ekor panjang di Pangandaran karena adanya gangguan nutrisi. Kondisi ini mempengaruhi jumlah individu dan kelom-pok monyet yang ada di Taman Wisata Alam Pangandaran. Selain itu, kondisi tersebut berpengaruh terhadap perilaku makan monyet ekor panjang di areal ini. Penurunan jumlah makanan dari pengunjung Taman Wisata sejak krisis ekonomi Asia tahun 1998 mendorong monyet-monyet di sini untuk berpindah daerah jela-jah seperti yang dilakukan oleh Kelompok Rengganis atau mencari sumber makanan baru yang ada di wilyah jelajahnya seperti yang dilakukan oleh Kelom-pok Goa Parat. Perilaku makan ikan yang ditunjukkan oleh individu-individu monyet anggota Kelompok Goa Parat merupakan tanggapan terhadap perubahan yang terjadi di habitatnya.
merampas makanan yang ada di dalam mulut individu lain. Saya melakukan peng-amatan pada akhir Juni 2012. Pada pengpeng-amatan ini saya mengamatai Kelompok A yang terdiri dari 214 individu. Saya mencatat umur dan jenis kelamin individu-individu monyet yang melakukan perampasan makanan monyet-monyet yang menjadi targetnya. Saya juga melakukan pengamatan pengaruh distribusi dan jumlah makanan yang diberikan oleh penjaga Taman Monyet. Selama pengamat-an, saya mencatat 747 kali perampasan makanan yang dilakukan oleh tujuh monyet (satu jantan dewasa dan 6 betina dewasa). Perilaku ini hanya ditunjukkan pada saat pemberiaan makan oleh penjaga. Dari 747 peristiwa perampasan, 609 di antaranya berhasil. Target utama perilaku perampasan ini adalah monyet juvenile (650 kali perampasan; 578 kali berhasil). Selain itu, betina dewasa (93 kali peram-pasan; 30 kali berhasil) dan betina muda (empat kali peramperam-pasan; sekali berhasil) juga menjadi target perampasan makanan). Perampasan makanan dengan target utama monyet juvenil dikarenakan monyet ini secara alami lebih lemah dari monyet dewasa. Penyebaran dan jumlah makanan tidak berbengaruh terhadap frekuensi perampasan makanan, tetapi lama waktu makan sangat berpengaruh terhadap frekuensi perampasan. Lama waktu makan ini menunjukkan jumlah biji gandum yang tersisa di dalam kantung makanan di mulut monyet-monyet yang menjadi target perampasan.
SUMMARY
ISLAMUL HADI. Food-Related Innovative behavior of The Macaques. Super-vised by BAMBANG SURYOBROTO, KUNIO WATANABE, ACHMAD FA-RAJALLAH.
Innovative behavior is a new behavior performed by individuals, occurred in low frequencies and restricted to particular area. Innovative behavior may allow animals in coping new environment and exploit more resources. Innovation as product and process of learning behavior has been observed and documented in various numbers of animals, mainly birds and primates. Nearly a half of those records came from food-related behaviors including food extraction technique, novel food items, and food preparation. Many of those behaviors were recorded in anthropogenic disturbance habitat, where animals were challenged by rapidly changing environments.
First, I observed long-tailed macaques (Macaca fascicularis) in Pengke-reman hot-spring in the highland of Mt. Rinjani in Lombok Island in 2008-2009. They showed innovative behavior in utilizing hot-spring. Two individuals of long-tailed macaques were observed in September 5, 2004 incidentally to soak their limbs into the hot spring water. On August 20-22, 2008, during three consecutive day surveys, 4, 5, and 6 out of 26 individuals’ macaques immersed their bodies into the hot-spring stream and pool, respectively. However, this behavior not performed by individuals of the two groups living in adjacent areas, Pelawangan Sembalun and Pos 3 Senaru Groups and two others group of Sebau hot spring 20 km eastern of Pengkereman were never observed to soak their bodies in the water. The behavior was driven by scarcity of natural food source while the artificial foods from human tourist available in the area of hot-spring. The behavior fixed in some individuals who learn the thermoregulation advantages of the hot spring bathing in their habitat in highland with low ambient temperatures.
Second, I observed fish-eating behavior performed by long-tailed macaques in Pangandaran Recreation Park in 2010-2012. This behavior was firstly observed in Rengganis Group on November 2010, when three adult and a sub-adult females ate fishes obtained from unattended net. Recently the fish-eating behavior is performed only by individuals of Goa Parat and K12 groups of Pangandaran Recreational Park. The fish-eating behavior that performed by long-tailed maca-ques in Pangandaran might be newly established as the innovative behavior. After Asian economical crisis in 1998 long-tailed macaques in the Pangandaran is in condition of nutritional deterioration. This condition affectted number of indivi-duals and groups of long-tailed macaques in the Recreation Park and also the feeding behavior of the macaques live in park. Decreasing amount of food from visitors drove the macaques to move their home range like Rengganis Group or exploit new source in their home range like Goa Parat Group. The development of fish-eating behavior by individuals of Goa Parat Group could be the response to the challenge posed by the changing situation of the habitat.
June 2012 and observed one of two monkey troops, comprising 214 monkeys. I recorded the age classes and sexes of the individuals who performed the snatching behavior and were snatched from, and examined the effects of provisioned food distribution and quantity on the frequency of snatching trials and success. During the survey, I recorded 747 snatching trials, of which 609 were successful, all of which were performed by seven individuals: one adult male and six adult females. The snatching behavior occurred only during provisioning. The target animals were primarily juveniles (650 trials, 578 successful), while cases in which food was snatched from adult females (93 trials, 30 successful) and sub-adult females (4 trials, 1 success) were less frequent. Among the juveniles, small juveniles had food snatched more frequently than large juveniles. The higher frequency of snatching trials against juveniles was likely due to their subordinate nature. Neither the distribution nor quantity of the provisioned foods had significant effects on the number of snatching trials nor did the successes, while the time elapsed after provisioning have significant negative effects, attributed to a decrease in the number of wheat grains left within the mouth pouch of the potential target animals.
© Hak Cipta Milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Mayor Biosains Hewan
FOOD-RELATED INNOVATIVE BEHAVIORS OF
THE MACAQUES
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2013
Penguji pada Ujian Tertutup (17 Desember 2012): 1. Dr Entang Iskandar
(Peneliti pada Pusat Studi Primata LPPM Institut Pertanian Bogor) 2. Dr Yamato Tsuji
(Assistant Professor pada Primate Research Institute, Kyoto University, Japan)
Penguji pada Ujian Terbuka (23 Januari 2013): 1. Dr Ir Novianto Bambang Wawandono
(Direktur Konservasi Keanekaragaman Hayati, Dirjen PHKA Kementerian Kehutanan Republik Indonesia)
2. Dr Entang Iskandar
Judul Disertasi : Food-Related Innovative Behaviors of The Macaques Nama : Islamul Hadi
NIM : G362080011
Disetujui oleh Komisi Pembimbing
Dr Bambang Suryobroto Ketua
Prof Dr Kunio Watanabe Dr Ir Achmad Farajallah MSi
Anggota Anggota
Diketahui oleh
Ketua Mayor Biosains Hewan Dekan Sekolah Pascasarjana
Dr Bambang Suryobroto Dr Ir Dahrul Syah MSc Agr
PRAKATA
Disertasi ini berjudul Food-Related Innovative Behavior of The Macaques. Pengamatan perilaku innovasi terkait makanan pada monyet ekor panjang telah dilakukan di Pemandian Air Panas Pengkereman yang berada di Taman Nasional Gunung Rinjani, Lombok dan Taman Wisata Alam Pangandaran, Ciamis Jawa Barat. Pengamatan juga telah dilakukan Chosikei Monkey Park, Shodoshima Island, untuk melihat perilaku inovasi terkait makanan pada Monyet Jepang. Saya mengucapkan terima kasih kepada para dosen pembimbing Dr Bambang Suryobroto, Dr Achmad Farajallah and Prof Dr Kunio Watanabe atas bimbingan-nya. Terima kasih juga saya sampaikan kepada Dr Yamato Tsuji, Prof Yuzuru Hamada, Prof Hiroo Imai, Prof Akichika Mikami, (Alm) Prof Osamu Takenaka yang telah mensponsori saya dalam program kerjasama penelitian antara IPB and Primate Research Institute Kyoto University. Terima kasih yang setinggi-tinginya saya sampakan kepada seluruh staf pengajar di Biosains Hewan (Dra Taruni Sri Prawasti M Si, Dr. Dedy Duryadi Solihin, Tri Heru Widarto, M.Sc, Dr. Dyah Perwitasari, Dr. Rika Raffiudin, Dr. Tri Atmowidi, MSi) dan teman-teman di Zoo Corner (Dr Kanthi Arum Widayati,Dr Tetri Widiyani, Sarah Nila, SSi, Puji Rianti, MSi, Eneng Nunuz R, MSi, Elda Irma Kawulur, MSi, Andi Darmawan, M.Si, Rizqi Amar Absdussalam, SSi, Mbak Tini, Mbak Ani, Pak Adi Surachman) atas kebersamaannya selama masa pendidikan doktor ini. Terima kasih juga penulis sampaikan kepda anggota Section Ecology and Conservation, Social System Evolution, Evolutionary Morphology Section, Primate Research Institute Kyoto University Japan atas diskusi mingguan selama penulis belajar di PRI. Ucapan terima kasih kepada Dr Entang Iskandar dari PSSP LPPM IPB dan Dr Novianto Bambang Wawandono (Direktur KKH PHKA Kementerian Kehutanan RI) atas segala masukannya pada saat ujian.
Ucapan terima kasih juga tidak lupa saya sampaikan kepada Dedi Surachman SSi, Adi Nurfebrimansyah SSi, M. Sya’roni SSi, Heru Iswandi SSi, Faizan SSi, Abdul Basith Nasrianto dan staf Taman Nasional Gunung Rinjani, Lombok atas bantuannya selama penelitian di Lombok. Bapak Yana Hendrayana dan staf Taman Wisata Alam Pangandaran dan BBKSDA Jawa Barat atas bantuan dan ijin selama penelitian di Pangandaran. Terima kasih kepda staf Chosikei Monkey Park, Shodoshima, Japan (Mr. Ryuji Hida, Akihiro Nishio, Mrs. Chiyoko Saeki, and Mr. Mitsuyuki Ishii) dan Cooperative Research Program 2012 Primate Research Institute Kyoto University kepada penulis dan Grant-in-Aid from the Department of Academy and Technology of Japan (Japanese Society for Promotion of Science, no. 23780160; 2011–2013 kepada Dr. Yamato Tsuji) atas segala bantuannya selama penelitian di Jepang. Salah satu bagian dari disertasi ini yang berjudul “Food snatching of free ranging Japanese macaques observed on Shodoshima Island : prelimary report” telah dipublikasikan pada Jurnal Primates pada awal tahun ini.
untuk mengikuti the 2010 Pre IPS and IPS congress di Inuyama and Kyoto dan presentasi poster “Hot-Spring Bathing Of Long-Tailed Macaques Observed in The High-land of Mt. Rinjani, Lombok, Indonesia” dan makalah berjudul “ Population status of long-tailed macaque (Macaca fascicularis) and lutung (Trachypithecus auratus) in Lombok Island, Indonesia.
Akhirnya, terima kasih atas segala dukungan yang diberikan oleh keluarga besar penulis, Irma Sofiana Afif dan Reyqa Rinjania atas segala dukungan dan kesabarannya selama penulis mengikuti pendidikan doctor.
Bogor, Mei 2013
DAFTAR ISI
INNOVATIVE BEHAVIOR OBSERVED IN THE HIGHLAND OF MT. RINJANI, LOMBOK, INDONESIA
III FISH-EATING BEHAVIOR OF LONG-TAILED MACAQUE FROM PANGANDARAN RECREATION PARK
DAFTAR TABEL
II.1 Climatic data of Pengkereman area measured during two observation-al
periods 4
II.2 Number of groups and individuals of long-tailed macaques in high-land
of Mt. Rinjani observed in 2004-2009 8
III.1 Temporal change of number of group and individuals of long-tailed macaques inhabited Pangandaran Recreation Park 16 III.2 Number of individuals involved on fish-eating during the experiment
conducted in November 9, 2010 17 III.3 Date, location, group and individual numbers performed fish-eating
behavior 19
IV.1 Daily change on food distribution designed during the study period 27 IV.2 Indviduals who conduct the snatching behaviour and details of their
target animals, and frequencies and success of the snatching 28
DAFTAR GAMBAR
II.1 Research location on Pengkeraman Area of Mount Rinjani National
Park (1980 m asl) 6
II.2 Yearly cumulative number of visitor and monthly distribution of average number visitor to Rinjani National Park from Gate of Senaru and Sembalun during (2001-2009)
7
II.3 Some postures of individuals during bathing activities in the hot-spring
area 9
III.1 Research site of Pangandaran Peninsula which designated as Nature
Reserved and Recreation Park under decree of Ministry of Agriculture 14 III.2 The Pangandaran Recreation Park and home ranges of Rengganis, Goa
Parat and K12 Groups 15
III.3 Temporal change of the visitor number to Pangandaran Recreation Park
during period of 1999-2012 15
I
INTRODUCTION
Innovative behavior may allow animals coping with environmental change, to exploit a wide range of habitat type and to increase their geographical range. Behavioral innovation may also be a good indicator of general behavioral flexibil-ity, which has long been predicted to be an important determinant among eco-logical variables (Reader & Macdonald 2005). Behavioral scientists have long observed that animals in various phylogeny performed behavioral innovation and differences in innovative tendency (Lefebvre et al. 1997). Classical example of behavioral innovation concerned with food extraction, preparation, and process-ing; for example: wheat and potato washing performed by Japanese macaque (Macaca fuscata) (Kawai 1965; Hirata et al. 2001), novel tool-use by chimpan-zees (Pan troglodytes) (Goodall 1964), and milk-bottle-top opening by British titmice (Parus caeruleus) (Fischer & Hinde 1949). Beside some benefits, inno-vative behavior also carry ecological and physiological cost such as consuming hazardous food items, risk from detected by predator, and investment of time and energy involved in exploiting a new resource.
Kummer and Goodall (1985), in their review on the primate behavioral innovation, have put the landmark to stimulate the general interest in topic of animal behavioral innovation. They suggested some behavioral innovations was derived from ability of the individuals to profit from an accidental happening, while others resulted from the use of existing behavior pattern for new purposes. Frequency of occurrence of behavioral innovation correlates with relative brain size of animals in both birds and primates (Lefebvre et al. 1997; Reader & Laland 2002). For example, over 500 cases of behavioral innovation have been reported in non- human primates, while over 2200 reports in birds (Reader & Laland 2002, Lefebvre & Bolhuis 2005). Nearly half of the innovation in non-human primates relates to foraging context including food extraction and novel food items. According to Reader and Laland (2005), some potential processes underlying the behavioral innovation were: 1) neophilia and neophobia, 2) exploration and curio-sity, 3) asocial learning and problem solving, 4) insight, 5) creativity 6) behav-ioral flexibility, 7) social processes.
2
I have engaged in the socio-behavioral studies long-tailed macaques living in Java, Bali and Lombok since 1999. In 2012, further, I got an opportunity to visit Japan and observe Japanese macaques. During those studies I got several cases of possibly newly acquired innovative behaviors; that is, hot-spring bathing and fish-eating of long-tailed macaques (part II and III) and snatching behavior of Japanese macaques (part VI). In this dissertation, I also describe these three innovative behaviors and discuss about the possible function of those behavioral innovations. I also discuss how these behaviors could be realized in what kind of the circumstances to make clear the key factors of causation of the behaviors and capabilities of the subject primate species.
References
Fischer J, Hinde RA.1949. The opening milk bottles by bird. British Bird 42: 347-357
Fooden J.1976. Provisional classification and key to living species of macaques. Folia Primatol 25 : 225-236
Fooden J. 1995. Systematic review of Southeast Asian longtail macaques, Macaca fascicularis (Raffles, [1921]). Fieldiana Zoology 81: 1-206.
Goodall J. 1964. Tool-using aimed throwing in a community of free-living chimp-anzee. Nature 201: 1264-1266
Hirata S, Watanabe K, Kawai M. 2001. Sweet potato washing revisited. In: Matsuzawa T, editor. Primate Origins of Human Cognition and Behavior. Tokyo (JP): Springer. p. 487-508
Kawai M. 1965. Newly-acquired pre-cultural behavior of the natural Troop of Japanese monkeys on Koshima Islet. Primates 6: 1-30
Kummer H, Goodall J. 1985. Conditions of innovative behavior in primates. Phil Trans R Soc Lond B 308: 203-214
Lefebvre L, Bolhuis JJ. 2005. Positive and negative correlates of feeding innovations in birds: evidence for limited modularity. In: Reader SM, Laland KN, editors. Animal Innovation. New York (US): Oxford Univer-sity Pr. p 39-61
Lefebvre L, Whittle P, Lascaris E, Finklestein A. 1997. Feeding innovations and forebrain size birds. Anim Behav 53: 1077-1097
Reader SM, Laland KN. 2005. Animal innovation: an introduction. In: Animal Innovation. Reader SM, Laland KN, editors. New York (US): Oxford University Pr. p 3-35
Reader SM, Laland KN. 2002. Social intelligence, innovation and enhanced brain size in primates. Proc Nat Acad Sci 99: 4436-4441
Reader SM, Laland KN. 2001. Primate innovation: sex, age, and social; rank differences. In J Primatol 22: 787-805
3
II
HOT-SPRING BATHING OF LONG-TAILED MACAQUES
OBSERVED IN THE HIGHLAND OF MT. RINJANI,
LOMBOK, INDONESIA
Abstract
Along the survey on the population of long-tailed macaques (Macaca fascicularis) in highland of Mt. Rinjani, Lombok, I found individuals of Pengke-reman Group performed hot spring bathing behavior in hot spring pool and stream of Pengkereman area (1982 m asl). Two individuals of long-tailed maca-ques were observed in September 5, 2004 incidentally to soak their limbs into the hot spring water. On August 20-22, 2008, during three consecutive day surveys, 4, 5, and 6 out of 26 individuals macaques immersed their bodies into the hot spring stream and pool, respectively. However, this behavior not performed by indiv-iduals of the two groups living in adjacent areas, Pelawangan Sembalun and Pos 3 Senaru Groups and two others group of Sebau hot spring 20 km eastern of Peng-kereman were never observed to soak their bodies in the water. The behavior was driven by scarcity of natural food source while the artificial foods from human tourist available in the area of hot spring. The behavior fixed in some individuals who learn the advantages of the hot spring bathing in their habitat in highland with low ambient temperatures.
Keywords: long-tailed macaque, Macaca fascicularis, Hot-spring bathing, Innovative behavior, Mt. Rinjani, Lombok
Introduction
The behavioral patterns that are non-universal or present at low occurrence in few populations are said as innovative behaviors (Van Schaik et al. 2006). Reader and Laland (2005) regarded innovative behavior as a new or modified behavioral pattern not previously found in the population.
4
the thermoregulation behavior. However, historically Japanese macaques in Jigokudani Monkey Park were introduced to the man-made hot spring pool by providing them with food immersed to water (Zhang et al. 2007).
Along the survey of tailed macaques in Lombok Island, I found long-tailed macaques performed hot spring bathing in highland of Mt. Rinjani National Park. The bathing was observed only in a group of macaques live in Pengkereman area (1982 m above sea level) while other groups were not doing it. I had opportunity to follow the emergence of the hot spring bathing from 2004 to 2009. This paper describes this newly established innovative behavior related with living in highland with low ambient temperature, scarcity of natural food source and availability of artificial food source in hot spring.
Methods
Mount Rinjani at northern Lombok Island is the highest mount in the island (3726 m asl) (BTNGR 2009) (Fig. II.1). The mountain is an active volcano. In the caldera about 2000 m asl there is a lake named Danau Segara Anak (Lake of Sea Child in the native Sasak language). Hot springs can be seen at several places around the mountain. The hot spring Aik Kalak (hot water) is located at the Pengkereman (place for bathing, 08°23′22.7″S, 116°25′11.9″E) at 1982m asl on the north of Danau Segara Anak (Fig. II.1). The hot-springs flow out to form part of river Kokok Putih (White River). The Pengkereman area is relatively flat and provides good bathing places; that is a segment of about 300m of Kokok Putih River contains several water pools and slow streams and large waterfalls (Fig. 1). Table II.1 Climatic data of Pengkereman area measured during two observational
periods
Temperature measured in (oC)
April 21-22, 2009 August 20-22, 2008
Mean Min Max n Mean Min Max n
Water 21 18 24 3 36.8 34 39 5
Ambient (daytime) 14.3 11 17 4 19.6 10 28 5
Ambient (midnight) 13 13 13 2 2.3 1 4 3
The climatic data of Pengkereman area at two observational periods are presented in Table II.1. April and August correspond to the early and mid of dry season based on the climatic data measured in the lowland of Lombok during 2001-2008 (Nandini & Narendra 2011). However, in April to early of May rain could occur in the highland of Mt. Rinjani. In April 21-22, 2009, water from Lake of Segara Anak overflowed to the hot spring to make the temperature decreased (see Table II.1).
5 It is just recently that hot spring bathing at the Pengkereman area became popular and many tourists foreign and native visit the area. Tourists immerse in these hot spring believing to heal the skin diseases and rheumatism (BTNGR 2009). The west bank of upper area of Pengkereman is designated as camping site for tourist.
The number of visitor, especially after the Rinjani National Park was established in 1992, is rapidly increasing from 1920 people in 2001 to 8469 in 2008 (Fig.II.2). The tourist averaged 586.3 individuals per month (min = 13 in April 2003, max = 1433 in August 2008) during tracking season (April to December, Fig. II.2).They frequently left food garbages and these provide foods for macaques. This might be the reason that groups of long-tailed macaques visit the camping sites, not only the Pengkereman area but also at several others. At present, we may find macaque groups up to 2700 m asl on the mountain slope where the uppermost camping site is located (Hadi 2005). In the rainy season (November to March) the trekking in the area can be very dangerous and peoples rarely climb up the mountain. Eruption of the volcano, further, may occur at unexpected time.
The hot spring bathing of long-tailed macaques at Pengkereman was first observed incidentally on September 2004 along with the population survey being carried out (Islamul Hadi, unpublished data). I visited again the area three times; April 30 to May 1, August 20 to 22 2008, and April 21 to 22, 2009. After May 4, 2009 the National Park stopped tracking activities because of the eruption of Mt. Rinjani and unstable conditions succeeded thereafter.
While in August 2008, I stayed at the camping site from 6:30 of 20th to 11:30 of 22nd and observed the macaques. When they took hot spring bathing the activity of the party is recorded in terms of time, duration and their detailed behavior. Macaques did not care much on the presence of observers and it was possible to record all in video.
6
Figure II.1 Research location on Pengkeraman Area of Mount Rinjani National Park (1980 m asl). Point A, B, C are the point where macaques took bath, A = stream with approximately 30 cm depth, B = stream with approximately 15 cm depth, C = pool with 3 x 10 m and 90 cm depth
Lombok Island
380000
380000
400000
400000
420000
420000
440000
440000
460000
460000
20000 9020000
40000 9040000
60000 9060000
80000 9080000
Road Mt. Rinjani National Park
Lake
N
E W
Figure II.2 (a) Yearly cumulative number of visitor
covered by volcanic sand with sparsely distributed shrubs and pine trees. are 13-18 groups of long
Rinjani National Park (Hadi of Mt Rinjani (Table
Observation in 2004
In 2004 three groups were found living on first group is at Pengkereman
consisting of 16 individuals by 22 individuals was km from Pengkereman
asl), consisting of 8 individuals.
pine trees. Since the food trees are very scarce in the area they might be unable to survive without leftover foods of huma
groups were only recently In September 5 the stream (point B in the stream by stepping
macaques stayed in the stream
moved away. Though observation was very limited it seemed that most monkeys did not take much attention to
thermometer to measure the stream’s temperature but was warm.
(a) Yearly cumulative number of visitor and (b) monthly distribution of average number visitor to Rinjani National Park from Gate of Senaru and Sembalun during (2001-2009) (source: Rekap Pengunjung Taman Nasional Gunung Rinjani 2001-2009, Balai Taman Nasional Gunung Rinjani, Mataram-Lombok.
Results
Most of the part of the highland of Mt. Rinjani beyond
covered by volcanic sand with sparsely distributed shrubs and pine trees. of long-tailed macaques observed in the whole
Rinjani National Park (Hadi et al. 2012). Five of them were found in the highland of Mt Rinjani (Table II.2).
Observation in 2004
In 2004 three groups were found living on the highland of Mt Rinjani. Pengkereman hot spring area found on September 4 16 individuals. The second group, Pos 3 Senaru Group, individuals was found at the northern side of Segara Anak
Pengkereman. The third groups lived at Pelawangan Sembalun (2700 m asl), consisting of 8 individuals. The two areas are covered by grass with scattered Since the food trees are very scarce in the area they might be unable to survive without leftover foods of human beings. It is reasonable to expect that the
were only recently appeared in the area after recurrent visit of tourist In September 5, 2004 I found two juveniles with their limbs immersed
in Fig. II.1) of Pengkereman, while other macaques ing on the stone after foraging leftover food
in the stream for a few minutes without any motion and soon Though observation was very limited it seemed that most monkeys did not take much attention to the streams. In this initial
thermometer to measure the stream’s temperature but perceptionally
7 covered by volcanic sand with sparsely distributed shrubs and pine trees. There the whole area of Mt. were found in the highland
8
In this year I could also observed Pos 3 Senaru and Pelawangan Senaru Group at the observational period (Table II.2). However, no macaques were seen to enter to the fresh water streams or pool.
Table II.2 Number of groups and individuals of long-tailed macaques in highland of Mt. Rinjani observed in 2004-2009
Location Year No.Group
Number of Individuals
AM AF SM SF JU IN TO
Pengkereman 2004 1 2 7 0 0 6 1 16
(hot spring) 2008(1) Macaques not appeared in the area
2008(2) 1 8 8 4 0 4 2 26
2009 Macaques not appeared in the area
Pos 3 Senaru 2004 1 4 8 0 1 4 5 22
(hot spring) 2008(1) Not observed
2008(2) Not observed
2009 2 2 5 0 0 3 1 11
1 2 0 0 2 0 5
Note : AF = adult male, AF = adult female, SM = sub-adult male, SF=sub adult female JU = juvenile , IN = infant, TO = Total
2008 (1) = observed in April-May 2008, 2008 (2) observed in August 2008
Observation in 2008
I revisited the Pengkereman hot spring site on April 30–May 1, 2008, but found no monkeys there. At the time no visitor sheltered in the area while the ambient temperature was cold (see Table II.1) and the pool and stream was cover-ed with cold water overflowcover-ed from the Segara Anak Lake (see Table II.1). At Pos 3 Senaru I found 12 individuals but no monkeys entering the cold water. The Pelawangan Sembalun Group was found consisting of 13 individuals and they too did not enter the pool.
9 males; two sub-adults, two juveniles) again immersed their bodies in the hot spring pool (point C) from 10:00-10:11. Macaques often jumped down from the heights of big rocks, swam and/or dived in the hot water. They often played with or groomed with each other in the hot water. When they took bathe in the hot water pool (Point C) individuals exhibited various activities including moving, resting, and aggressive behaviors (drag, bite, chase) as Japanese macaques did in Jigokudani Monkey Park (Zhang et al. 2007) (Fig. II.3(a)). In the stream (point A), most individuals immersed their bodies simultaneously and eat remains of human meal left as garbage inside the hot water ((Fig. II.3(b)). The posture of them as well as the duration of immersing their bodies into the hot water resemble to Japanese macaques in the Jigokudani Monkey Park in Japan and human beings enjoying hot-spring bathing. Thus, in this observation period of 2008 many ma-caques of Pengkereman Group have already used to enter and immerse themselves in the hot water stream and pools.
The Pos 3 Senaru group was seen at the same location where they first found. However, no macaques of the group found to enter fresh water stream. In this period, I could not found Pelawangan Sembalun group, possibly because of our time constraint. Another unidentified group, however, was sighted near to the Pengkereman hot spring area (about one kilometer to the south).
Observation in 2009
I revisited the Pengkereman hot spring on April 21-22, 2009. I found no monkeys there. I also found no tourist sheltered in the camping site. I could only found the group of Pos 3 Senaru about 3 kms away from the area. The group con-sisting of 24 individuals. Here many tourists sheltered.
On March 10-12, 2009, I visited Sebau hot spring. Two groups of long-tailed macaques were found in the area. They were five and 11 individuals (Table II.2), respectively. Those individuals of macaques was not well habituated to human since they kept distance of more than 15 meters even though I served them with food. During three days of observation, only human tourists occupied the hot spring pool (size approximately 3 m x 3 m).
(a) (b)
10
Discussion
Bathing is a commonly observed behavior of long-tailed macaque in the tropical area. For examples, it was seen in Java (Pangandaran, Cikakak, Solear) and several places in Bali (Sangeh and Ubud) and Lombok (Suranadi, Gunung Pengsong and Sekotong) (Islamul Hadi, unpublished data). Fooden (1995) regard-ed this species as excellent swimmer. All of those activities were recordregard-ed in the lowland. However, hot-spring bathing of long-tailed macaque in lowland and highland has not been reported yet.
In highland of Mt. Rinjani there are 5 groups of long-tailed macaques (see Table 2). The area is covered with sparsely distributed shrubs and pine trees that could not provide enough foods for them. The groups began to forage around the highland when recurrent tourists visited the area starting from 2003. Macaques explored the leftover food and this might be the main reason of their presence in the highland area. Moreover, foods (and/or its leftover) are frequently thrown into the water of pool and stream
Hot spring bathing of long-tailed macaques seems to appear only recently. Individuals crossing into hot stream water were first seen in 2004 at Pengkereman. The hot spring bathing behavior could be driven by their activities in collecting remains of human meals immersed in shallow hot water pool and stream. Further, after experiencing the hot water, the behavior could be fixed in some individuals of the group as the response to the environmental condition, especially to low ambient temperatures on the highland. The thermoregulation behavior in various primate populations have been reported, for example, baboons sheltered in cave in the cold day (Barret et al. 2004). The famous Japanese monkeys in the Jigokudani Monkey Park also use hot spring bathe more often in winter and less in summer (Zhang et al. 2007).
The case of Pengkereman macaques may be an innovative behavior since the behavior is non-universal and occur in low frequency (van Schaik et al. 2006) and only recently emerge in the population (Reader & Laland 2005). Of those seven processes underlying innovation (Reader & Laland 2005), exploration and behavior flexibility may play roles in the emergence of hot spring bathing. Macaques explored leftover food in hot spring area since the scarcity of natural food. The immersed food in hot water might attract macaques to enter the hot spring and experience hot water. Behavioral flexibility in large- and complex-brained taxa, including macaques, might allowed them to modify their behavior when they exposed to special circumstance (for macaques of Pengkereman Group, the circumstances are living in highland with low temperatures, scarcity of natur-al food source and availability of artificinatur-al food source in hot spring area). Some individuals may use the ability to adopt it for their better lives.The adoption mechanism may include a trigger (that is, food in the hot water) and fixation of the behavior rewarded by thermoregulation advantages. There are possibilities that at anywhere the same situation is available the similar innovative behavior may occur.
11 wide and there is only one pool available. The pool is occupied intensively by human. It is very difficult for macaques to share it with human. If, however, macaques could find food immersed in the hot spring pool and in doing so they conditionally learn the thermoregulation advantage of the hot water, it can be expected that individuals of Sebau Group might eventually bath in the hot spring.
Conclussion
I conclude that the hot-spring bathing by long-tailed macaques only found in Pengkereman Hot-Spring in highland of Mt Rinjani National Park. The behav-ior is an innovative behavbehav-ior performed by members of Pengkereman Group. The behavior emerge because of the macaques is attracted to enter hotspring to obtain the food. They get nutritional and thermoregulation advantages from hot-spring.
References
Barrett L, Gaynor D, Rendall D, Mitchell D, Henzi SP. 2004. Habitual cave use and thermoregulation in chacma baboons (Papio hamadryas ursinus). J Hum Evol 46: 215–222
[BTNGR] Balai Taman Nasional Gunung Rinjani. 2008. Buku Informasi Taman Nasional Gunung Rinjani (in Indonesian). Mataram (ID): Balai Taman Nasional Gunung Rinjani. p 1-25
Carpenter A. 1887. Monkeys opening oysters. Nature 36:53.
Fooden J.1976. Provisional classification and key to living species of macaques. Folia Primatol 25 : 225-236
Hadi I. 2005. Distribution and present status of long-tailed macaques (Macaca fascicularis) in Lombok Island, Indonesia. Nat Hist J Chula Univ Supl 1: 90
Hadi I, Suryobroto B, Perwitasari D. 2007. Food preference of semi-provisioned macaques based on feeding duration and foraging party size. Hayati J Biosci 14 : 13-17
Malaivijitnond S, Lekprayoon C, Tandavanitj N, Panha S, Cheewatham C, Hamada Y. 2007. Stone-tool usage by Thai long-tailed macaques (Macaca fascicularis). Am J Primatol 69:227–233.
Minakata K. 1984. Ethnography and Folklore on Monkeys (in Japanese).Tokyo (JP) : Heibonsha. p 1-364
Nandini R, Narendra BH, 2011. Kajian perubahan curah hujan, suhu dan tipe iklim pada zone ekosistem di Pulau Lombok (abstract in English). J Analisis Kebijakan Kehutanan 8: 228-244
Reader SM, Laland KN. 2005. Animal innovation: an introduction. In: Animal Innovation. Reader SM, Laland KN, editors. New York (US): Oxford University Pr. p 3-35
12
Van der Kraan A. 2009. Lombok: Penaklukan, Penjajahan dan Keterbelakangan. Supanra MD, translator. Mataram (ID): Lengge. Translation from: Lom-bok: Conquest, Colonization, and Underdevelopment, 1870-1940. p1-502 Van Schaik CP, van Noordwijk M, Wich, SA. 2006. Innovation in wild Bornean
orangutans (Pongo pygmaeus wurmbii). Behaviour 143:839 – 876
13
III
FISH-EATING BEHAVIOR OF LONG-TAILED MACAQUES
OBSERVED FROM PANGANDARAN RECREATION PARK
Abstract
I observed fish-eating behavior performed by long-tailed macaques in Pangandaran Recreation Park. This behavior firstly observed in group of Reng-ganis on November, 2010, while three adult females and a sub-adult female ate fishes obtained from unattended net left in docked boat. Recently the fish-eating behavior is performed only by individuals of Goa Parat and K12 groups of Pangandaran Recreational Park. The fish-eating behavior that performed by long-tailed macaques in Pangandaran might be newly established as the innovative behavior. After Asian economical crisis 1998 long-tailed macaques in the Pangandaran is in condition of nutritional deterioration. This condition affected number of individuals and groups of long-tailed macaques in the Recreation Park and also the feeding behavior of the macaques live in park. Decreasing amount of food from visitors drove the macaques to move their home range like Rengganis Group or exploit new source in their home range like Goa Parat Group. The development of fish-eating behavior by individuals of Goa Parat Group could be the response to the challenge posed by the changing situation of the habitat. Key words: Pangandaran, long-tailed macaque, Macaca fascicularis, fish-eating,
innovative behavior
Introduction
The long-tailed macaques (Macaca fascicularis) are flexible in their food choices. They are regarded as frugivorous because in non-provisioned habitats their diet is dominated by fruit (Hasanbahri et al. 1996; Yeager 1996). In human-altered places (for example, recreation parks and temples), they tend to be omni-vorous by shifting their diet to include food items introduced by human (Wheatly 1989; Son 2003; Hadi et al. 2007).
14
addition to macaques, some researchers reported fish eating in chacma baboons (Papio ursinus; Hamilton & Tilson 1985) and olive baboons (P. anubis; Ransom 1981).
Along the routine work in 2009-2012 on the population monitoring of long-tailed macaques in Pangandaran Recreation Park we found individuals consuming various species of fish originally discarded by fishermen. They also stole fishes from docked boats and from unattended fishing nets. We call this as “fish-eating behavior”. This behavior seemed to be established recently. Here we describe the fish-eating as an innovative behavior of the long-tailed macaques in response to changes in their environment. This innovative behavior may allow Java, Indonesia (Sumardja & Kartawinata 1977). The Reserve partially includes 37.7 Ha of Recreation Park where visitor can enter freely (Fig. III.1 & III.2). The Park is planted forest dominated by teakwood (Tectona grandis), bayur (Ptero-spermum javanicum) and mahogany (Swietenia macrophyla).
During 1999-2009, management of the Park was handed over from West Java Nature Conservation Office to forestry-related company Perhutani. The management allowed the fishermen to enter freely and now they settle at beaches of Goa Panggung, Wisma Wana and Cirengganis (see Fig III.2). They park their fish-ing boat on the sandy beaches after their daily fishing sessions. On the parked boat fishermen collect fishes from fishing nets and throwing out undesirable fish and/or other materials trapped. Some fishing nets also put abandoned in the area.
Figure III.1 Research site of Pangandaran Peninsula which designated as Nature Reserved and Recreation Park under decree of Ministry of Agricul-ture Nomor : 34/KMP/1961 and Nomor : 170/Kpts/Um/3/1978.
(a) mainland of Java, (b) recreation park, (c) nature reserve b
a
15
Figure III.2 The Pangandaran Recreation Park (part b of Fig. 1) and home range of Rengganis, K12 and Goa Parat Groups and boat parking areas at Goa Panggung (A), Wisma Wana (B), Cirengganis (C)
Since the Asian economical crisis occurred in 1998, number of visitors to the Pangandaran Recreation Park increased significantly in 1999 but suddenly decreased drastically more than 8 folds (Fig III.3). It was 223591 people in 2000 to 27092 in 2006 and increasing slightly to 110530 in 2012.
16
Animal Subject
During the field works within 2009-2012, I found 5-6 groups of long-tailed macaque inhabited the Park. I could compare the number of groups and individuals of long-tailed macaques from the previous observation in the literatures. The number of individuals and groups fluctuated year by year (Table III.1). Group of Rengganis, Goa Parat, K12 and Parkir occupied east coast of Pangandaran Recreation Park (Fig. III.2), while Goa Jepang and Cikamal groups occupied western coast. Their home ranges were highly overlapped.
Table III.1 Temporal change of number of group and individuals of long-tailed macaques inhabited Pangandaran Recreation Park.
Year Group Individuals Sources 1976 7 100 Watanabe et al. 2009 macaques in this area, I incidentally found four macaques (three adult females and one sub-adult female) of Rengganis group ate fishes and crab. They took fishes and crabs from unattended net in the boat harbored in Rengganis beach. Hence, I called this as fish-eating behavior. I followed the group and recorded sex and age class of participants of fish-eating. I also recorded time and location of macaques eating fish and also the local name of fishes eaten by macaques. Whenever possible, I also identified individuals who involved on fish eating.
In November 9, I conducted experiment to check whether other individuals of the Rengganis group would ate fish. I offered hair-tailed fish (Trichiurus leptu-rus) and island mackerels (Rastelliger sp.). I also treated the same fishes to other group of macaques living in the Park. Sex, age class and responses of group mem-bers to the fishes were recorded.
I revisited Pangandaran Recreation Park on March, April, November and December 2011, January, March and April 2012. To collect more information of the history of fish-eating in the Pangandaran, I also interviewed staffs of the park and the fishermen who harbored their boat in Recreation Park. The fishes eaten by monkeys were identified following Allen (1999) and Warmbrunn (2009).
17
Results
Observation and Experiment in 2010
On November 7, 2010, I found incidentally three adult and one sub-adult females of Rengganis Group ate unidentified fishes and sea crab (Portunus pelagicus) in Rengganis Beach. They took fishes and sea crab from unattended net left in docked boat in Rengganis beach (Point C in Fig III.2).
On November 9, I recorded eight individuals of this group responded positively to eat fish served by me. They were two adult males including alpha-male (Faisal), a sub-adult alpha-male, four adult fealpha-males and an unidentified sex juvenile, respectively. A late infant also observed to take remaining fish from her mother. Two adult males of the neighboring group, Goa Parat, also performed fish-eating after being served by me. I also observed alpha male (Romy) of the Goa Parat Group and an adult male hold and smelt fish, while four juveniles observed him. An adult female also observed to smell the fish. On the other hand, no individuals of Parkir, Goa Jepang, and Cikamal groups recorded to eat fishes. Number of individuals who responded positively to fishes offered is shown in Table III.2.
Table III.2 Number of individuals involved on fish-eating during the experiment conducted in November 9, 2010 fishermen could manage to fish in nearby shores, the fishes were treated as high-economical value. Therefore, no fishes were discarded. Without availability of fish, I could not observe the fish-eating behavior. Coincidentally, I found the group of Rengganis disappeared from the park. It was probably moved to the reserved forest. In this period, number of individuals of Goa Parat Group decreas-ing from 57 to 46. Eleven individuals moved from their original group to form new group of K12 which ranging in overlap with Goa Parat Group.
18
dussumeri). On 08:12-08:33, Goa Parat group members moved to Wisma Wana Beach. Twelve individuals (an adult male; a sub adult male; four adult females; three sub-adult females; two juveniles) ate fusilier fish (Caesio sp.) and sea crab (Portunus pelagicus). Then, Goa Parat group moved back to Goa Panggung. Eleven individuals (an adult male; a adult male; four adult females; three sub-adult females; two juveniles) were recorded to eat croaker fish, crab and shrimp (Penaeus merguinensis).
On November 16, Goa Parat Group foraged in Goa Panggung. Some indiv-iduals searched left-over fish on chariot and fish net in the area. No indivindiv-iduals recorded to eat fish. Then, the group moved to Cirengganis, while K12 group visited Goa Parat. Four individuals of K12 (an adult male, three adult females) recorded to eat fish in 10:36-10:40. They ate gabel fish (unidentified), long-tongue (Cynoglossus lingua) bearded croaker fish, moonfish (Leighnathus equulus),
mantis shrimp (Odontodactylus scyllarus) and sea crab. Goa Parat group also visited the area in the afternoon (16:23-17:54). Six individuals (an adult male; a sub-adult male; two adult females; a sub-adult female; a juvenile) observed to eat gabel fish and sea crab. I found a sub-adult male ate puffer fish (Tetraodon luna-ris) and the remains also eaten by a sub-adult female. This fish is well known by fisherman as the poisonous fish in the area.
I observed again the group of Goa Parat in November 17. At 10:07-10:58 I observed four individuals (an adult male; two adult females; a juvenile) of Goa Parat Group ate moonfish, sea crabs, and mantis shrimps at Goa Panggung. In the evening at the same day (16:26-16:45), I observed four individuals (an adult male, two adult females, a juvenile) of K12 Group ate bearded croaker fish, jeprox fish (Terapon puta).
On December 18, I observed Goa Parat Group in Cirengganis (08:09-08:39). However, no macaques ate fish in this area. At 15:09-16:18, I observed 20 macaques (two adult males, four adult females, 14 juveniles) of Goa Parat Group ate fish in Wisma Wana. They ate abandoned bearded croaker fish in the seashore.
Observation in 2012
I visited Pangandaran Recreation Park on January 22-23. I also visited the park on March 23-25 and April 19-22. These periods were west wind season. The raw fishes from the fishermen were unavailable. On January 22, I observed Goa Parat Group foraged in Goa Panggung Beach. At 08:37-09:06, I found an adult male macaque ate grilled hair-tailed fish and the remain was taken by a juvenile. This human-cooked fish might came from visitor garbage.
Interview
19 They argued that the group started to eat fish because of scarcity of food. At the time visitors were decreasing and the Management prevented the macaques to contact with them. The fruits were also lacking in the Park.
Table III.3 Date, location, group and individuals numbers performed fish-eating behavior
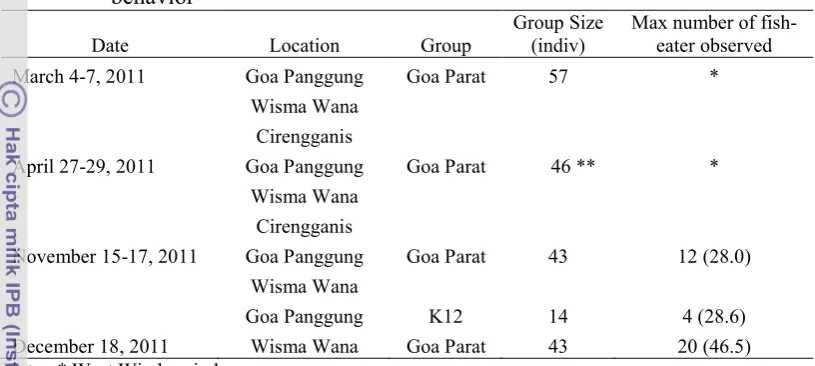
Date Location Group Group Size (indiv) Max number of fish-eater observed
March 4-7, 2011 Goa Panggung Goa Parat 57 * disappeared from the Park (see Table 2). Number in parenthesis in column of maximum number of fish-eater represents percentage of individuals performed fish-eating behavior in each period observation.
Discussion
Fish-eating is newly established behavior of long-tailed macaques in Pangandaran Recreation Park, even though they were exposed to aquatic environment which provide them chance to exploit aquatic food resources (e.g. fishes, crustacean and mollusks). Hadi (2001) who observed food habit of the same groups on period of 1999-2000 had reported no fishes in their food items. Erawati (2004) and Rotua (2004) who observed long-tailed macaques in this area were also not found this behavior. Son (2003) also did not confirm long-tailed macaques in Vietnam to consume raw fish even though they observed to live in aquatic environment (Mangrove Forest Park). Recently the case of fishing and eating fish by long-tailed macaque was reported in Sumatera and Kalimantan (Stewart et al. 2007).
The start of the behavior in Pangandaran Recreation Park might be within 2010 in Rengganis group. No macaques were reported to eat fish in the Park be-fore the finding in November 2010. After the Rengganis Group moved out of the Park, only members of Goa Parat and K12 in various sex and age class perform fish-eating behavior (Table III.2 & III.3).
20
seemed to be driven by the exposition of fish in the period of nutritional deterio-ration.
The same condition occurred recently in Pangandaran Recreational Park. During period of 1999-2006, visitor of the Pangandaran Park decreased drastically. Moreover, the Pangandaran Recreation Park had zero visitors within January-September 2006 after Tsunami disaster. Even though number of visitors increased every year after 2006, the policy of Park Management did not allow visitors to provide food for macaques. They prevented the macaques to contact with visitor in Park area. This condition lead to decrease in the availability of food since macaques of Pangandaran is highly dependent to human-provisioned food (Hadi 2001). In other word, after 1998 long-tailed macaques in the Pangandaran is in condition of nutritional deterioration. This condition affected number of individu-als and groups of long-tailed macaques in the Recreation Park and individu-also the feeding behavior of the macaques live in park. Decreasing amount of food from visitors drove the macaques to move their home range like Rengganis Group or exploit new source in their home range like Goa Parat Group. The development of fish-eating behavior by individuals of Goa Parat Group could be the response to the challenge posed by the changing situation of the habitat.
Once the group learned to eat fish, it is persisted and was not affected by season. During the West Wind period, when most of fishermen temporarily stop to fish, raw fishes were not available for macaques. However, when they found left-over human-cooked fish, they remember the fish and ate it. In any case, the season affected the frequencies of fish-eating behavior and it was performed more frequently in non-West Wind rather than in West Wind period.
Even though my first finding on fish-eating in Rengganis group was performed by adult females and sub-adult females, I suggested the adult males could be the innovator of the behavior in Goa Parat Group. The adult males might copied the behavior from their neighbor Rengganis. However, adult females and juveniles were more frequent to be seen involved on fish-eating behavior. This is reasonable since juveniles and adult females need more food to meet their nutriti-onal requirement for growth and reproduction.
Recently the fish-eating behavior is performed only by individuals of Goa Parat and K12 groups of Pangandaran Park. Given the overlapping home ranges, I predict other groups of the macaques could learn from Goa Parat to get experi-ences to taste fish and perform the fish-eating behavior. Further observation would be addressed on the intensive individuals identification to construct the possible lineage on the mechanism of fish-eating behavior transmission.
Conclussion
21
References
Allen G. 1999. Marine Fishes of South-East Asia. Singapore (SG): Periplus. p 1-302
Erawati E. 2004. Populasi dan distribusi monyet ekor panjang (Macaca fasci-cularis, Raffles 1821) di Taman Wisata Alam Pananjung Pangandaran. (abstract in English) [minithesis]. Bandung (ID): Universitas Padjajaran. Fooden J. 1995. Systematic review of Southeast Asian longtail macaques, Macaca
fascicularis (Raffles, [1921]). Fieldiana Zoology 81: 1-206.
Hadi I. 2001. Pemilihan makanan oleh monyet karier butawarna (abstract in Eng-lish) [minithesis]. Bogor (ID). Institut Pertanian Bogor
Hadi I, Suryobroto B, Perwitasari D. 2007. Food preference of semi-provisioned macaques based on feeding duration and foraging party size. Hayati 14 : 13-17
Hamilton WJ, Tilson RL. 1985. Fishing baboons at desert waterholes. Am J Pri-matol 8: 255 –257.
Hasanbahri S, Djuwantoko, Ngariana IN. 1996. Komposisi jenis tumbuhan pakan kera ekor panjang (Macaca fascicularis) di habitat hutan jati. (abstract in English). Biota 1:1-8
Leca JB, Gunst N, Watanabe K, Huffman M. 2007. A new case of fish-eating in Japanese macaques: implications for social constraints on the diffusion of feeding innovation. Am J Primatol 69:821–828
Majumder J, Lodh R. Agarwala BK. 2012. Fish feeding adaptation by Rhesus macaque Macaca mulatta (Cercopithecidae) in the Sundarban mangrove swamps, India. J Threat Taxa 4: 2539-2540.
Purnama AR. 1998. Strategi makan kelompok kera ekor panjang (Macaca fascicularis, [Raffles 1821]) di Taman Wisata Alam-Cagar Alam Panan-jung Pangandaran Jawa Barat. (abstract in English) [minithesis]. Bandung (ID): Universitas Padjajaran.
Ransom TW. 1981. Beach troop of the Gombe. Lewisburg PA (US): Bucknell University Pr. p 1-319
Rotua MY. 2004. Pola akivitas harian dan perilaku monyet ekor panjang panjang (Macaca fascicularis, Raffles 1821) di Taman Wisata Alam Pananjung Pangandaran. (abstract in English) [minithesis]. Bandung (ID): Univer-sitas Padjajaran.
Son VD. 2003. Diet of Macaca fascicularis in a mangrove forest Vietnam. Lab Primate News 42:1–5.
Stewart AE, Gordon CH, Wich SA, Schroor P, Meijaard E. 2008. Fishing in Macaca fascicularis: a rarely observed innovative behavior. Int J Prima-tol 29:543–548
Sumardja EA, Kartawinata K. 1977. Vegetation analysis of the habitat of banteng (Bos javanicus) at the Pananjung-Pangandaran Nature Reserve, West Java. Biotrop Bull 13: 3-43
Warmburnn A. 2009. The livelihoods of Sundanese fishermen in Cijulang, West Java and their implications for fisheries management [thesis]. New South Wales (AU) University of Newcastle.
22
Watanabe K, Mitani M, Suryobroto B, Hadi I, Widayati KA, Megantara EN, Gurmaya KJ, Wedana M, Dirgayusa IW, Purnama AR, Brotoisworo E.2009. Population trends of Tracypithecus auratus and Macaca fascicularis in the Pangandaran Nature Reserve, Indonesia. In Lohman D, editor. ATBC Asia-Pasific Chapter Annual Meeting: Assesing and Restor-ing Biodiversity in a Human Dominated Landscape. 2007. Feb 12-15; Chiang Mai, Thailand. Chiang Mai (TH): ATBC. p 106
Wheatly BP. 1989. Diet of balinese temple monkeys, Macaca fascicularis. Kyoto Univ Overseas Res Rep Study Asian non-Human Primates 7: 62-75.
23
IV
FOOD- SNATCHING BEHAVIOR OF FREE-RANGING
JAPANESE MACAQUES OBSERVED ON SHODOSHIMA
ISLAND: A PRELIMINARY REPORT*
__________________________________________________________________ * The chapter has been published in Primates 54: 153-158, received: 18 October
31
V
GENERAL DISCUSSION
Behavioral innovation has suggested as complex mechanism that involved cognition process. However, in fact many behavioral innovations reported so far are relatively simple behavior pattern such as eating novel food. For example, chimpanzees eating mangoes (Takahasi 1983), blue monkeys (Cercopithecus mitis) eating a flying squirrel (Fairgrieve 1997), black howler monkeys (Alouatta caraya) eating marsh plant roots (Rodrigues & Maricho-Filho 1995), common squirrel monkeys (Saimiri sciureus) eating bat (Sarsa et al. 1997) and ring-tailed lemurs (Lemur catta) eating chameleon (Oda 1996), fish-eating by Japanese macaques (Watanabe 1989) and rhesus macaque (Mamjuder et al. 2012). Another examples from non-novel food eating are ring-tailed lemur immersing own tail to drink water (Hosey et al. 1997) and using stepping stick on walking chimpanzee to protect against the spiny trees of kapok (Ceiba pentandra) (Alp 1997). Recent-ly, rhesus macaques also reported to use pro-tools (bordering) to open the coco-nuts (Comins et al. 2012)
32
macaques (Yamada, 1966; Zhang & Watanabe 2007). The stressful conditions that often caused by the alteration of environment by human can be important for the appearances of innovative behaviors.
Next, all these macaques could get the appropriate materials for their behavioral innovations. Lombok long-tailed macaques could find the hot water pools/streams that might provide for monkeys to take warm in the cold temp-erature of highland. It is extremely important that the water temptemp-erature is being kept optimal for hot water bathing during the dry season so as many human beings enjoy it. Further, long-tailed macaques in the Pengkereman got artificial food source in the hot spring area which attract them to the hot spring and experience the hot water. Pangandaran macaques got new foods (raw fishes) after the food supplies for them decreased much. It is noteworthy that many fishes abundant were present every morning in front of monkeys at the Rengganis, Wisma Wana, and Goa Panggung area, which make them familiar with raw fishes and give opportunity to try it. Japanese macaques on the Koshima Island also started raw-fish eating about seven years after the provision for macaques drastically decreased (Watanabe 1989). It may take times before macaques add raw fishes in their daily diets. Snatching behavior of Japanese macaques is rarely observed in the limited populations. The frequency occurred in the Choshikei Groups is apparently very high among current population of Japanese macaques. When the target monkeys resist to the snatchers and/or the family members such as mothers or sisters protest them the snatching will be very difficult. The target macaques may learn the counter strategy and become to avoid from those macaques who take foods from the cheek pouch of them. Hence, it is known that Choshikei macaques are very gregarious and much more tolerant to the other individuals (Yamada 1966; Zhang & Watanabe 2007). The inter-individual distance is very short among them and often they moved around on the heads of other individuals when provisions are offered. In such a crowded situation macaques may be able to catch targets and snatch foods with less resistance.
33 America. Recently, they found and survive in the human-altered environment and are commensally animals.
The provisioned groups can develop to the larger groups when comparing with pure wild groups. Monkeys in the larger groups need to adjust themselves to the different condition that will be caused by many individuals, which is not experienced by them when they live in the pure wild condition. They need to thrive well within the entangled relationships that are created by many individuals involved in the group. The ability to form larger groups may also indicate the higher flexibility of monkeys adjusting themselves to more complicated social relationships.
References
Alp R. 1997. ‘Stepping-sticks’ and ‘seat-sticks’: new types of tools used by wild chimpanzee (Pan troglodytes) in Sierra Leone. Am J Primatol 41: 45-52 Comins JA, Russ BE, Humbert KA, Hausser MD. 2011. Innovative
coconut-opening in semi free-ranging rhesus monkey (Macaca nulatta): a case of behavioral propensities. J Ethol 29: 187-189
Fairgrieve C. 1997. Meat-eating by blue monkey (Cercopithecus mitis stuhl-manni): predation of a flying squirrel (Ananchanus derbiatus jacksani). Folia Primatol 68: 354-356
Fooden J. 1995. Systematic review of Southeast Asian longtail macaques, Macaca fascicularis (Raffles, [1921]). Fieldiana Zoology 81: 1-206.
Hosey GR, Jacques M, Pitts A. 1997. Drinking from tails: social learning of a novel behavior in a group of ring-tailed lemur (Lemur catta). Primates 38: 415-422
Kawai M. 1965. Newly-acquired pre-cultural behavior of the natural Troop of Japanese monkeys on Koshima Islet. Primates 6: 1-30
Majumder J, Lodh R. Agarwala BK. 2012. Fish feeding adaptation by Rhesus macaque Macaca mulatta (Cercopithecidae) in the Sundarban mangrove swamps, India. J Threat Taxa 4: 2539-2540.
Oda R. 1996. Predation on chameleon by a ring-tailed lemur (Lemur catta) in the Berenty Reserve, Madagascar. Folia Primatol 67 : 40-43
Rodrigues FHG, Maricho-Filho J. 1995. Feeding on a marsh-living herbaceous plant by black howler monkeys (Alouatta caraya) in Central Brazil. Folia Primatol 6 : 115-117
Sarsa LL, Ferrari SF, Dina ALCB. 1997. Feeding behavior and predation of a bat by Saimiri sciureus in a semi-natural Amazonian environment. Folia Primatol 68: 194 – 198
Takahasi K. 1983. Mahale chimpanzee taste mangoes toward acquisition of a new food item? Primates 24 : 273-275
Watanabe K. 1989. Fish: new addition to the diet of Japanese macaques on Koshima Island. Folia Primatol 52: 124 -131.
34
ATBC Asia-Pasific Chapter Annual Meeting: Assesing and Restoring Biodiver-sity in a Human Dominated Landscape. 2007. Feb 12-15; Chiang Mai, Thailand. Chiang Mai (TH): ATBC
Yamada M. 1966. Five nature troops of Japanese macaques in Shodoshima Island (I): distribution and social organization. Primates 7: 315–362
35
VI
GENERAL CONCLUSSION
37
VII
REFERENCES
Allen G. 1999. Marine Fishes of South-East Asia. Singapore (SG): Periplus. p 1-302
Alp R. 1997. ‘Stepping-sticks’ and ‘seat-sticks’: new types of tools used by wild chimpanzee (Pan troglodytes) in Sierra Leone. Am J Primatol 41: 45-52 Barrett L, Gaynor D, Rendall D, Mitchell D, Henzi SP. 2004. Habitual cave use
and thermoregulation in chacma baboons (Papio hamadryas ursinus). J Hum Evol 46 : 215–222
Brockmann HJ, Barnard CJ. 1979. Kleptoparasitism in birds. Anim Behav 27:487– 514.
Brostrom G, Holmberg H. 2011. Generalized linear models with clustered data: fixed and random effects models with glmmML. Comp Stat Data Anal 55 : 3123–3134.
Brown GR. 2004. Tolerated scrounging in non-human primates. Behav Brain Sci 7: 562–563.
[BTNGR] Balai Taman Nasional Gunung Rinjani. 2008. Buku Informasi Taman Nasional Gunung Rinjani (in Indonesian). Mataram (ID): Balai Taman Nasional Gunung Rinjani. p 1-25
Carpenter A. 1887. Monkeys opening oysters. Nature 36:53.
Comins JA, Russ BE, Humbert KA, Hausser MD. 2011. Innovative coconut-opening in semi free-ranging rhesus monkey (Macaca nulatta): a case of behavioral propensities. J Ethol 29 : 187-189
Erawati E. 2004. Populasi dan distribusi monyet ekor panjang (Macaca fascicula-ris, Raffles 1821) di Taman Wisata Alam Pananjung Pangandaran. (abs-tract in English) [minithesis]. Bandung (ID): Universitas Padjajaran. Fairgrieve C. 1997. Meat-eating by blue monkey (Cercopithecus mitis
stuhlman-ni): predation of a flying squirrel (Ananchanus derbiatus jacksani). Folia Primatol 68: 354-356
Feistner ATC, McGrew WC. 1989. Food sharing in primates: a critical review. In: Seth K, Seth S, editors. Perspectives in Primate Biology, vol 3. New Delhi (IN): Today and Tomorrow’s Pr. p 21–36.
Fischer J, Hinde RA.1949. The opening milk bottles by bird. British Bird 42: 347-357
Fooden J.1976. Provisional classification and key to living species of macaques. Folia Primatol 25 : 225-236
Fooden J. 1995. Systematic review of Southeast Asian longtail macaques, Macaca fascicularis (Raffles, [1921]). Fieldiana Zoology 81: 1-206.
Gilby IC. 2006. Meat shearing among the Gombe chimpanzees: harassment and reciprocal exchange. Anim Behav 71: 953–963.
Goodall J. 1964. Tool-using aimed throwing in a community of free-living chimp-anzee. Nature 201: 1264-1266
Hadjichrysanthou C, Broom M. 2012. When should animals share food? Game theory applied to kleptoparasitic populations with food sharing. Behav Ecol 23: 977–991.