Bemisia tabaci
GENNADIUS (HEMIPTERA: ALEYRODIDAE)
PADA TANAMAN TOMAT (Lycopersicum esculentum
MILL.):
PANJANG ROSTRUM DAN SAYAP PADA BEBERAPA
KETINGGIAN TEMPAT SERTA PERIODE RETENSI
Tomato chlorosis virus
(ToCV)
HERLIE ARIFEBRIAWAN
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
ABSTRAK
HERLIE ARIFEBRIAWAN,Bemisia tabaciGennadius (Hemiptera: Aleyrodidae) pada Tanaman Tomat (Lycopersicum esculentumMill.): Panjang Rostrum dan Sayap pada Beberapa Ketinggian Tempat serta Periode Retensi Tomato chlorosis virus (ToCV). Dibimbing olehDEWI SARTIAMIdanGEDE SUASTIKA.
Bemisia tabacidigolongkan ke dalam ordo Hemiptera, subordo Sternorrhyncha, superfamili Aleyrodoidea, dan termasuk ke dalam famili Aleyrodidae. Faktor lingkungan dapat mempengaruhi ukuran panjang tubuh kutukebul ini. Serangga yang berada di dataran tinggi memiliki ukuran lebih besar, bobot tubuh lebih berat, dan warna lebih gelap dibandingkan dengan spesies serangga yang sama yang berada di dataran rendah. Sepanjang abad ke-20, B. tabaci telah menjadi vektor patogen-patogen tertentu baik di rumah kaca maupun di lahan terbuka di wilayah beriklim hangat. Penyakit Tomato chlorosis virus (ToCV) adalah salah satu penyakit yang ditularkan olehB. tabaci.Penyakit ToCV ini termasuk baru yang dikategorikan “new emerging disease” akibat pengaruh pemanasan global. Penelitian ini bertujuan untuk (1) mengetahui variasi panjang rostrum dan sayap B. tabaciberdasarkan ketinggian tempat hidupnya dan (2) mengukur periode retensi ToCV yang ditularkanB. tabaci. Penelitian ini dilakukan dengan pengukuran panjang rostrum dan panjang sayap B. tabacidari beberapa tempat dengan ketinggian berbeda. SampelB. tabaciberasal dari Ciawi (Bogor) ketinggian 573 m dpl (di atas permukaan laut), Pacet (Cianjur) ketinggian 1225 m dpl, Batu (Batu) ketinggian 675 m dpl dan Cikole (Sukabumi) ketinggian 1022 m dpl. Hasil penelitian menunjukkan bahwaB. tabaciyang memiki panjang rostrum dan panjang sayap yang berbeda-beda di setiap tempat. Panjang rostrum dari daerah Pacet 226.06±21.72 µm, Cikole 213.03±21.84 µm, Batu 211.21±18.60 µm, Ciawi 201.52±17.06 µm. Panjang sayap dari daerah Pacet 1031.33±95.66 µm, Cikole 1023.33±60.13 µm, Batu 928.67±67.40 µm, Ciawi 916.67±53.57 µm. Penelitian yang lain adalah mengakuisisi B. tabaci ke dalam kurungan serangga yang berisi tanaman tomat yang positif virus ToCV dan sebagian lagi dimasukkan ke dalam kurungan serangga yang berisi tanaman tomat yang sehat. Periode makan akuisisi selama 48 jam. Seekor imagoB. tabaciyang telah di akuisisi dipindahkan ke tanaman uji. Setiap 24 jamB. tabacitersebut dipindahkan ke tamanan uji berikutnya secara berseri selama tujuh hari. Percobaan dilakukan dengan 10 ulangan dari B. tabaci yang di-akuisisi ke tanaman tomat bervirus ToCV dan 10 ulangan dari B. tabaci yang di-akuisisi ke tanaman tomat sehat. Hasil penelitian menunjukkan bahwa B. tabaci viruliferus dalam seri inokulasi ke tanaman tomat mampu menularkan virus hingga hari ke-4, hal ini dilihat dari timbulnya gejala ToCV pada tanaman tomat tersebut. Untuk konfirmasi bahwa gejala klorosis yang muncul pada tanaman tomat uji disebabkan oleh infeksi ToCV dilakukan dengan RT-PCR.
Bemisia tabaci
GENNADIUS (HEMIPTERA: ALEYRODIDAE)
PADA TANAMAN TOMAT (Lycopersicum esculentum
MILL.):
PANJANG ROSTRUM DAN SAYAP PADA BEBERAPA
KETINGGIAN TEMPAT SERTA PERIODE RETENSI
Tomato chlorosis virus
(ToCV)
HERLIE ARIFEBRIAWAN
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Pertanian di Departemen Proteksi Tanaman,
Fakultas Pertanian, Institut Pertanian Bogor
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
Judul : Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) pada Tanaman Tomat (Lycopersicum esculentum Mill.): Panjang Rostrum dan Sayap pada Beberapa Ketinggian Tempat serta Periode RetensiTomato chlorosis virus(ToCV)
Nama : Herlie Arifebriawan
NRP : A34061206
Menyetujui,
Pembimbing I Pembimbing II
Dra. Dewi Sartiami, M.Si. Dr. Ir. Gede Suastika, M.Sc.
NIP. 19641204 199103 2 001 NIP. 19620607 198703 1 003
Mengetahui,
Ketua Departemen Proteksi Tanaman
Dr. Ir. Dadang, M.Sc. NIP. 19640204 199002 1 002
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 01 Februari 1988, sebagai anak ketiga dari tiga bersaudara pasangan Suherijono dan Lies Sumerdulinah. Penulis menyelesaikan pendidikan Sekolah Dasar di SD Islam Al-Azhar 4 Kebayoran Lama pada tahun 2000. Penulis melanjutkan studi ke SLTP Islam Al-Azhar 3 Bintaro dan lulus tahun 2003. Kemudian pada tahun 2006, penulis menyelesaikan pendidikan di SLTA Islam Al-Azhar 1 Kebayoran Baru. Selama SLTP dan SMA penulis aktif dalam kegiatan ekstrakulikuler, seperti sepak bola, basket dan OSIS.
PRAKATA
Puji serta syukur penulis panjatkan atas ke hadirat Allah SWT yang telah memberikan rahmat dan karunia-Nya, sehingga penulis dapat menyelesaikan penulisan skripsi dengan judul “Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) pada Tanaman Tomat (Lycopersicum esculentum Mill.): Panjang Rostrum dan Sayap pada Beberapa Ketinggian Tempat serta Periode Retensi Tomato chlorosis virus (ToCV)”. Penelitian ini bertujuan untuk (1) mengetahui variasi panjang rostrum dan sayap B. tabaci berdasarkan ketinggian tempat hidupnya dan (2) mengukur periode retensi ToCV yang ditularkannya., yang hasilnya diharapkan dapat memberikan informasi dasar mengenai perbandingan morfologi yang mencakup panjang rostrum dan sayap kutukebul B. tabaci pada beberapa ketinggian tempat yang berbeda dan lama periode retensi kutukebul B. tabaci yang berperan sebagai vektor ToCV dalam tanaman tomat. Skripsi ini disusun sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian pada Fakultas Pertanian, Institut Pertanian Bogor. Penelitian dilaksanakan di Laboratorium Biosistematika Serangga dan Laboratorium Virologi, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor dari April sampai November 2010.
Pada kesempatan ini, penulis mengucapkan terima kasih yang sebesar-besarnya kepada Dra. Dewi Sartiami, M.Si. dan Dr. Ir. Gede Suastika, M.Sc. sebagai dosen pembimbing skripsi dan telah memberikan arahan kepada penulis sehingga skripsi ini dapat terselesaikan dengan baik. Terima kasih juga penulis sampaikan kepada Dr. Ir. Abdul Munif, M.Sc sebagai dosen penguji, Dr. Ir. Swastiko Priyambodo, M.Si sebagai dosen pembimbing akademik, serta kepada kedua orang tua (Suherijono dan Lies Sumerdulinah), dan kakak-kakakku Rilie Adisetiawan dan Hersye Nurauliawati atas doa dan semangatnya.
Tak lupa juga penulis mengucapkan terima kasih kepada rekan-rekan Laboratorium Biosistematika dan Musium Serangga, Ibu Aisyah, Mbak Lia, Mbak Elsa, Mbak Atik, Fitrah, Vani. Rekan-rekan di Laboratorium Virologi, Mbak Tuti, Bu Ifa, Bu Rita, Pak Irwan, Ka Aceu, Ita, Amel, Lara, Dillah, Laras, Anto dan rekan-rekan PTN 43 lainnya yang tak dapat disebutkan satu persatu.
Bogor, Maret 2011
DAFTAR ISI
Halaman
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... ix
DAFTAR LAMPIRAN... x
PENDAHULUAN ... 1
Latar Belakang... 1
Tujuan Penelitian ... 3
Manfaat Penelitian ... 3
TINJAUAN PUSTAKA ... 4
Bemisi tabaci... 4
Tomato chlorosis virus(ToCV) ... 5
Hubungan Virus dengan Serangga Vektornya ... 6
BAHAN DAN METODE ... 8
Tempat dan Waktu Penelitian... 8
Metode Penelitian ... 8
Survei dan Pengambilan Sampel Kutukebul dan Tanaman Tomat Sumber ToCV... 8
Pembuatan Preparat dan IdentifikasiB. tabaci... 8
Pengukuran Panjang Rostrum dan SayapB. tabaci... 9
Pengukuran Periode Retensi ToCV pada Tubuh Kutukebul ... 10
Analisis Data ... 12
HASIL DAN PEMBAHASAN ... 13
Identitas Kutukebul Pengkoloni Tomat... 13
Variasi Panjang Rostrum dan Panjang SayapB. tabaci... 14
Periode Retensi ToCV dalam Tubuh Kutukebul ... 15
KESIMPULAN DAN SARAN ... 20
Kesimpulan ... 20
Saran... 20
DAFTAR PUSTAKA ... 21
DAFTAR TABEL
Halaman
1. Panjang rostrum dan sayapB. tabaci... 15 2. Masa infektif B. tabaci dalam penularan berseri Tomato chlorosis
DAFTAR GAMBAR
Halaman
1. PengukuranB. tabaci... 10 2. Tanaman tomat untuk pengujian retensi ToCV ... 10
3. MorfologiB. tabaci... 13 4. Tanaman tomat uji yang memperlihatkan gejala klorosis setelah
diinokulasi ToCV melalui B. tabaci (kiri) dan yang tidak
memperlihatkan gejala (kanan)... 16
5. Hasil elektroforesis menggunakan pasangan primer spesifik
DAFTAR LAMPIRAN
Halaman
1. Panjang rostrum dan panjang sayap B. tabacidari Kecamatan Pacet,
Kabupaten Cianjur ... 26
2. Panjang rostrum dan panjang sayapB. tabacidari Kecamatan Cikole,
Kota Sukabumi ... 27
3. Panjang rostrum dan panjang sayap B. tabaci dari Kecamatan Batu,
Kota Batu... 28
4. Panjang rostrum dan panjang sayap B. tabacidari Kecamatan Ciawi,
Kabupaten Bogor ... 29
PENDAHULUAN
Latar Belakang
Bemisia tabaci digolongkan ke dalam ordo Hemiptera, subordo Sternorrhyncha, superfamili Aleyrodoidea, dan termasuk ke dalam famili
Aleyrodidae (Borror et al. 1992). Spesies B. tabaci merupakan kutukebul yang memiliki kisaran inang luas. Kalshoven (1981), mengelompokkan tanaman inang
dari serangga ini meliputi beberapa famili, yaitu famili Compositae, Cucurbitaceae,
Cruciferae, dan Solanaceae. Menurut Hill (1987), tanaman inang utama B. tabaci
adalah kapas, tembakau, tomat, ubi jalar, ubi kayu, beberapa jenis gulma, serta
tanaman lain yang dapat menjadi inang alternatif. Hal ini menyebabkan banyaknya
nama umum B. tabaci yang dikenal luas, diantaranya adalah kutukebul kapas (cotton whitefly), kutukebul tembakau (tobacco whitefly), dan kutukebul ubi jalar (sweetpotato whitefly) (Kalshoven 1981).
Imago B. tabaciberwarna kuning dengan panjang tubuh 1-1.5 mm dan sayap yang tertutup oleh tepung berwarna putih. Lama hidup imago bervariasi tergantung
faktor lingkungan. Lama hidup imago betina hingga 16 hari, sedangkan imago
jantan umurnya lebih singkat dibandingkan imago betina, yaitu sekitar 9-15 hari
(CABI 2005).
Suhu tubuh serangga sangat dipengaruhi oleh suhu lingkungan tempat
serangga tersebut hidup. Serangga beradaptasi dengan suhu lingkungan untuk
menjaga kebugaran tubuhnya. Salah satu bentuk adaptasi serangga adalah dengan
adanya perbedaan ukuran tubuh (Kingsolver 2008). Serangga yang hidup pada suhu
rendah memerlukan nutrisi yang lebih banyak dan memiliki metabolisme yang lebih
lambat dibandingkan serangga yang hidup pada suhu tinggi. Serangga yang
memiliki metabolisme rendah mempunyai ukuran tubuh lebih panjang (Prado
2010). Menurut Oliveira et al. (2004) serangga yang berada di dataran tinggi memiliki ukuran lebih besar, bobot tubuh lebih berat, dan warna lebih gelap
dibandingkan dengan spesies serangga yang sama yang berada di dataran rendah.
Data mengenai variasi morfologi B. tabaci berdasarkan ketinggian tempat belum ada, sehingga diperlukan penelitian mengenai pengetahuan variasi morfologi
Beberapa tahun belakangan,B. tabacitelah menjadi masalah utama bagi para petani di seluruh dunia. WalaupunB. tabacidianggap sebagai grup serangga tropis, spesies berbahaya ini banyak sekali ditemukan di seluruh belahan lain dunia,
terutama di daerah beriklim subtropis. Sepanjang abad ke-20, B. tabaci telah menjadi vektor patogen-patogen tertentu baik di rumah kaca maupun di lahan
terbuka di wilayah beriklim hangat (Martin et al. 2000). Stadia nimfa dan imago kutukebul merupakan stadia yang menyebabkan kerusakan tanaman (Morales
2001).
Tomato chlorosis virus (ToCV) adalah salah satu virus tanaman yang ditularkan oleh B. tabaci. ToCV diketahui tidak dapat ditularkan melalui cairan perasan tanaman sakit ataupun melalui benih. ToCV ditularkan ke dalam jaringan
tanaman oleh kutukebul Bemisia tabaci biotipe A dan B, Trialeurodes abutilonea (Wisler et al. 1998b), dan T. vaporariorum (Wintermantel & Wisler 2006).
Menurut Wisler & Duffus (2001) B. tabaci merupakan kutukebul yang memiliki efisiensi menularkan ToCV lebih baik dibanding kutukebul yang lainnya.
ToCV ini termasuk baru yang dikategorikan “new emerging disease” akibat
pengaruh pemanasan global (Segev et al. 2004). ToCV pertama kali tersebar di negara bagian Florida, USA sejak tahun 1989. Virus ini dengan cepat menyebar ke
seluruh dunia dan sampai saat ini keberadaannya telah dilaporkan di banyak negara
seperti di Perancis (Masseet al. 2008), Spanyol (Navas-Castilloet al. 2000; Lozano et al.2006), Taiwan (Tsaiet al.2004), dan Yunani (Katanyaet al.2008). Menurut Hartono & Wijonarko (2007), penyakit ini pada tanaman tomat telah menyebar di
sentra pertanaman tomat di Magelang, Jawa Tengah dan Purwakarta, Jawa Barat.
Penyakit ToCV ini mempunyai gejala menguning pada bagian interval daun
(Duffuset al. 1996, Hirotaet al. 2010), bintik-bintik nekrotik kecil (Wintermantel & Wisler 2006), mengeriting (Hirota et al. 2010), dan gejala lanjutan akan menyebabkan daun tampak berwarna merah kecoklatan (Wisler et al. 1998a). Menurut Wintermantel et al. (2005), ToCV adalah virus yang berbentuk panjang
lentur (flexuous filamentous) dengan ukuran diameter 12 nm dan panjang rata-rata 800-850 nm.
perlu dilakukan untuk mengetahui lama waktu retensi virus ToCV dalam tubuh
serangga vektorB. tabaci.
Tujuan Penelitian
Penelitian ini bertujuan untuk (1) mengetahui variasi panjang rostrum dan
sayapB. tabaciberdasarkan ketinggian tempat hidupnya dan (2) mengukur periode retensi ToCV yang ditularkannya.
Manfaat Penelitian
Hasil penelitian diharapkan dapat memberikan informasi dasar mengenai
perbandingan morfologi yang mencakup panjang rostrum dan sayap kutukebul
B. tabaci pada beberapa ketinggian tempat yang berbeda dan lama periode retensi kutukebul B. tabaci yang berperan sebagai vektor ToCV dalam tanaman
TINJAUAN PUSTAKA
Bemisi tabaci
Kutukebul B. tabaci digolongkan ke dalam ordo Hemiptera, subordo Sternorrhyncha, superfamili Aleyrodoidea, dan termasuk kedalam famili Aleyrodidae
(Bororet al. 1992). SeranggaB. tabacimerupakan spesies kutukebul yang memiliki kisaran inang luas. Kalshoven (1981), mengelompokkan tanaman inang dari serangga
ini meliputi beberapa famili, yaitu famili Compositae, Cucurbitaceae, Cruciferae, dan
Solanaceae. Menurut Hill (1987), tanaman inang utama B. tabaci adalah kapas,
tembakau, tomat, ubi jalar, ubi kayu, beberapa jenis gulma, serta tanaman lain yang
dapat menjadi inang alternatif. Hal ini menyebabkan banyaknya nama umum
B. tabaci yang dikenal luas, diantaranya adalah kutukebul kapas (cotton whitefly), kutukebul tembakau (tobacco whitefly), dan kutukebul ubi jalar (sweetpotato whitefly) (Kalshoven 1981).
Imago B. tabacimenurut Usman (2003) lebih menyukai daun tanaman tomat dibandingkan dengan daun tanaman cabai. Permukaan daun tanaman tomat secara
genetik memiliki rambut-rambut daun yang banyak sehingga dapat meningkatkan
suhu dan kelembaban mikro dan mengakibatkan pertumbuhan dan perkembangan
serta kehidupan serangga ini lebih kondusif. Selain itu, kondisi permukaan daun
tanaman tomat tersebut lebih memberi kenyamanan serangga ini untuk melakukan
aktivitas makan dan bertelur karena serangga ini lebih mudah menjangkarkan
tungkai-tungkainya dibandingkan pada tanaman cabai yang memiliki permukaan
daun yang lebih licin.
Menurut Martin (2000), ciri morfologi B. tabaci adalah sebagai berikut: Telur
yang baru diletakkan berwarna kekuningan dan biasanya tertutup lilin, warna telur akan
berubah setelah 24 jam menjadi berwarna coklat. Nimfa instar satu berbentuk bulat
panjang, berwarna hijau cerah dengan panjang tubuh 0.22 mm dan lebar 0.13 mm.
Nimfa instar dua berwarna hijau gelap dengan panjang tubuh 0.28 mm dan lebar 0.17
mm, dengan antena sangat pendek dan tungkai yang tereduksi. Pupa berbentuk bulat
panjang, dibagian toraks agak melebar dan cembung dengan abdomen yang tampak
ukurannya lebih panjang dari panjang alur kaudal (caudal furrow). Hampir separuh bagian operkulumnya menutupi bagian vasiform orifice. Imagonya berwarna kuning dengan panjang tubuh 1-1.5 mm dan sayap yang tertutup oleh tepung berwarna putih.
Lama hidup imago bervariasi tergantung faktor lingkungan. Lama hidup imago
betina sekitar enam hari, tetapi pada kondisi tertentu mampu mencapai 60 hari dan
pada umumnya imago jantan umurnya lebih singkat dibandingkan imago betina,
yaitu sekitar 9-17 hari (CABI 2005).
Kutukebul B. tabaci ini dapat beradaptasi pada suhu hangat yakni berkisar
14 sampai 35 0C, dengan suhu optimum sekitar 25 sampai 30 0C. Pada tanaman
tomat, imago mampu hidup selama 10-15 hari pada suhu 28 sampai 30 0C,
sedangkan imago betina mampu memproduksi 195 telur pada suhu 25 0C (Smith
2009). Hal ini menunjukkan bahwa aktivitas terbesar B. tabaci menyerang tanaman terjadi pada musim panas dan untuk daerah tropis populasi serangga ini
banyak dijumpai pada musim kemarau.
Sepanjang abad ke-20,B. tabacitelah menjadi vektor patogen-patogen tertentu baik di rumah kaca maupun di lahan terbuka di wilayah beriklim hangat ( Martinet al. 2000). Sebagai vektor, kutukebul dilaporkan dapat menularkan beberapa kelompok virus, diantaranya: closterovirus, geminivirus, carlavirus, potyvirus,
nepovirus, luteovirus, dan virus DNA yang berbentuk batang (Markham et al. 1994). Di antara kelompok virus tersebut yang paling banyak ditularkan adalah
closterovirus (Famili Closteroviridae, Genus Crinivirus) dan geminivirus (Famili Geminiviridae, Genus Begomovirus) (Muniyappa & Reddy 1983, Wisler et al. 1998).
Tomato chlorosis virus(ToCV)
ToCV pertama kali tersebar di negara bagian Florida sejak tahun 1989.
Virus ini dengan cepat menyebar ke seluruh dunia dan sampai saat ini
keberadaannya telah dilaporkan di banyak Negara seperti di Perancis (Masseet al.
dengan sangat efisien secara semipersisten, sehingga kejadian penyakit kuning
pada tanaman tomat sangat dipengaruhi oleh kepadatan populasi dan aktivitas
serangga ini (Jacquemondet al.2008).
ToCV merupakan kelompok RNA dengan panjang partikel 800-850 nm
(Wintermantel et al. 2005). Virus ini mempunyai dua jenis genom berupa RNA utas tunggal RNA yaitu RNA 1 dan RNA 2 yang masing-masing berukuran 7.8
dan 8.2 kb. Menurut Martelli et al. (2000), RNA 1 mengkode dua jenis protein yang terlibat dalam replikasi virus, sedangkan RNA 2 mengandung beberapa
gen yaitu sebuah protein kecil yang hidrofobik, sebuah protein berukuran sekitar
60 kDa, dan dua jenis protein mantel yaitu CP dan CPm. Selubung protein minor
(CPm) pada ToCV, yang membentuk bagian ekor/ujung virion memiliki peranan
dalam penularan dengan kutukebul. CPm dari ToCV memiliki kespesifikan
dengan reseptor T. vaporariorum dan B. tabaci. Menurut Wintermantel (2006),
kespesifikan virus dan vektornya sangat ditentukan oleh reseptor yang ada pada
stilet serangga dengan CP dari virus bersangkutan.
Infeksi ToCV pada tanaman tomat menyebabkan klorosis yang pada awalnya
terjadi pada daun-daun bagian bawah kemudian berkebang ke bagian pucuk, pada
lamina daun di antara tulang daun mengalami klorosis (interveinal yellowing).
Setelah munculnya vektor kutukebul, gejala ini berkembang dalam beberapa
minggu. Daun menjadi tebal dan keriting,dan mudah rapuh jika dipatahkan. Virus
ini dapat menyebar dengan cepat ke pertanaman di sekitar sumber virus sesuai
dengan aktivitas kutukebul sebagai vektornya sehingga kejadian penyakit dalam
satu kebun petani sering ditemukan mencapai lebih dari 90% (Navas-Castilloet al.
2000). Kehilangan hasil akibat infeksi virus ini di lapangan menyebabkan ukuran
buah mengecil, jumlah buah berkurang saat panen, dan menurunnya umur tanaman
(Wintermantel 2004).
Hubungan Virus dengan Serangga Vektornya
Sebagian besar virus tanaman ditularkan oleh serangga vektor dan tergantung
pada perilaku serta kapasitas penyebaran vektor tersebut untuk menyebarkan virus
dari tanaman ke tanaman. Ordo Hemiptera merupakan kelompok serangga yang
vektor virus tanaman merupakan anggota subordo Sternorrhyncha (kutukebul,
kutuputih) (Borroret al.1992). Dari 1200 spesies kutukebul yang telah diteliti,hanya empat spesies (B. tabaci, T. vaporariorum, T. abutilonea, dan T. ricini) ditemukan
menularkan virus tanaman (Jones 2003).
Virus ditularkan oleh kutukebul diklasifikasikan berdasarkan lamanya vektor
mempertahankan virus. Klasifikasi ini dapat dibedakan antara non-persisten yaitu
jika kemampuan vektor menularkan virus hilang dalam beberapa menit atau
beberapa jam, semipersisten yaitu jika kemampuan vektor menularkan virus hilang
setelah beberapa hari, dan persisten jika kemampuan vektor untuk menularkan virus
tersimpan untuk beberapa hari atau selama vektor tersebut hidup (Sylvester 1956).
Klasifikasi lain digunakan dalam menunjuk tempat retensi virus dalam vektor.
Dalam hal ini virus dipertahankan pada ujung stilet yang disebut stylet-borne (Kennedy et al. 1962), sedangkan virus dipertahankan pada saluran pencernaan
disebut foregut-borne (Nault & Ammar 1989). Hal ini menjelaskan bahwa virus non-persisten merupakan stylet-borne dan semipersisten adalah foregut-borne (Fereres & Moreno 2009).
Interaksi antara protein selubung virus dengan kutukebul terjadi saat
penempelan partikel virus dengan reseptor sehingga virus dapat tertular. Virus akan
berada dalam tubuh serangga vektor saat diakuisisi. Virus menuju sel epitel saluran
pencernaan dan berasosiasi dengan kelenjar saliva serangga. Virus bersirkulasi
dalam usus serangga sampai akhirnya virus mencapai ke stilet dan masuk ke dalam
tanaman sehat saat vektor makan cairan floem. Virus tersebut memerlukan waktu
akuisisi dan inokulasi satu hari hingga beberapa minggu dalam tubuh serangga
(Gray & Banerjee 1999).
Beberapa istilah dalam penularan virus oleh serangga virus yaitu transovarial
merupakan kemampuan serangga dalam menularkan virus yang diturunkan pada
telur serangga, transtadial adalah kemampuan serangga dalam menularkan virus
dari stadia ke stadia selanjutnya. Periode makan akuisisi adalah waktu yang
dibutuhkan serangga untuk mengambil virus dari tanaman terinfeksi sedangkan
periode makan inokulasi adalah waktu yang dibutuhkan serangga untuk
BAHAN DAN METODE
Tempat dan Waktu Penelitian
Survei dan pengambilan sampel kutukebul dan tanaman tomat yang
menunjukkan gejala penyakit klorosis dilakukan di sentra produksi tomat di daerah
Bogor, Cianjur, Batu dan Sukabumi. Identifikasi, pengukuran panjang rostrum dan
sayap kutukebul dilakukan di Laboratorium Biosistematika Serangga sedangkan
pengukuran periode retensi dan identifikasi virus dilakukan di Laboratorium
Virologi Tumbuhan, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut
Pertanian Bogor. Penelitian dilakukan dari bulan April sampai November 2010.
Metode Penelitian
Survei dan Pengambilan Sampel Kutukebul dan Tanaman Tomat Sumber ToCV
Pengambilan sampel kutukebul dilakukan di beberapa daerah sentra produksi
tomat di Jawa Barat dan Jawa Timur yang mempunyai ketinggian tempat yang
berbeda yaitu di Kecamatan Ciawi, Kabupaten Bogor dengan ketinggian 573 m di
atas permukaan laut (mdpl) dengan suhu rata-rata 25.30C; Kecamatan Batu, Kota
Batu 675 mdpl suhu 24.40C; Kecamatan Cikole, Kota Sukabumi 1022 mdpl suhu
22.2 0C; Kecamatan Pacet, Kabupaten Cianjur 1225 mdpl suhu 20.9 0C. Untuk
analisa variasi ukuran tubuh B. tabaci, koleksi imago kutukebul dilakukan dari beberapa pertanaman tomat petani sekitar 5-6 kebun per daerah ketinggian tempat.
Imago kutukebul yang telah dikoleksi dengan aspirator, kemudian dimasukkan ke
dalam tabung gelas yang telah diisi alkohol 70% agar awet sampai diamati di
laboratorium. Untuk analisa periode retensi ToCV dalam tubuh kutukebul, tanaman
tomat sumber ToCV diambil dari tanaman tomat yang menunjukkan gejala khas
penyakit klorosis di daerah Pacet, Cianjur.
Pembuatan Preparat dan IdentifikasiB. tabaci
Identifikasi B. tabaci dilakukan berdasarkan morfologi pupa yang diperoleh dari hasil perbanyakan serangga. Agar morfologi pupa kutukebul dapat diamati dan
asam fuchsin. Preparasi pewarnaan dilakukan sebagai berikut. Pupa kutukebul
direndam ke dalam tabung reaksi yang berisi alkohol 80% dan dipanaskan pada
suhu 80-100 0C selama 10 menit, kemudian dipindahkan ke dalam tabung reaksi
yang berisi KOH 10% dan dipanaskan sampai isi pupa lunak atau terlihat
transparan. Kemudian spesimen dipindahkan ke dalam cawanSyracus.Pada cawan Syracus, spesimen pupa ditekan perlahan pada bagian lingkar dorsal posterior sampai seluruh isi pupa keluar, lalu dicuci dengan aquades sampai bersih dan sisa
KOH hilang. Pewarnaan dilakukan dengan merendam spesimen pupa di dalam
campuran asam fuchsin dan asam asetik glacial dengan perbandingan 1:1 sampai
berwarna merah yaitu sekitar 10-20 menit. Kemudian spesimen pupa direndam
dalam alkohol 80% sampai warna merah yang optimum, kemudian direndam lagi
dalam larutanCarbol xyleneselama satu menit. Spesimen pupa kemudian direndam dalam alkohol absolut beberapa menit, lalu di dalam minyak cengkeh selama 10
menit. Selanjutnya pupa diambil dan diletakkan di tengah kaca objek. Setelah pupa
ditata lurus, diteteskan Canada balsam secara merata dan ditutup dengan kaca penutup, kemudian preparat dikeringkan ke dalam pemanas selama 4-7 hari.
Pengamatan morfologi pupa kutukebul dilakukan di bawah mikroskop compound dan identifikasi dilakukan berdasarkan Martin (2000).
Pengukuran Panjang Rostrum dan SayapB. tabaci
Pengukuran panjang rostrum dan sayap dilakukan terhadap imago betina
B. tabaci yang sudah dikoleksi dari berbagai sentra produksi tomat di Jawa Barat dan Jawa Timur. Pengukuran tersebut dilakukan dengan bantuan mikroskop stereo
yang dilengkapi skala mikrometer. Pengukuran panjang sayap dilakukan cukup
dengan pembesaran 80 kali, sedangkan untuk pengukuran panjang rostrum perlu
pembesaran yang lebih tinggi yaitu 110 kali. Ukuran panjang sebenarnya (dalam
µm) diperoleh dengan membagi ukuran yang teramati di mikroskop dengan nilai
pembesaran mikroskop. Untuk mendapatkan data yang dapat mewakili panjang
Gambar 1 PengukuranB. tabaci. Panjang rostrum (kiri) dan sayap (kanan)
Pengukuran Periode Retensi ToCV pada Tubuh Kutukebul
Periode retensi ToCV pada tubuh kutukebul diukur dengan menginokulasikan
virus dari satu individu serangga vektor ke bibit tomat secara berseri. Satu ekor
imagoB. tabaci yang baru berumur sehari dibiarkan makan akuisisi pada tanaman tomat sakit sumber ToCV atau tanaman tomat sehat, sebagai kontrol selama dua
hari (48 jam), kemudian dipindahkan ke bibit tomat yang baru berumur seminggu
setelah dipindahkan ke pot individu atau dua minggu setelah disemai dan dibiarkan
makan inokulasi selama sehari (24 jam). Setelah makan inokulasi pada satu bibit
tomat, serangga tersebut (individu yang sama) dipindahkan ke bibit tomat baru dan
dibiarkan makan inokulasi juga selama sehari (24 jam). Kegiatan seperti ini terus
dilakukan sampai proses inokulasi berseri dilakukan pada bibit tomat baru yang ke
tujuh. Perlakuan yang sama juga dilakukan terhadap sembilan ekor individu imago
B. tabaci yang lain sehingga perlakuan ini diulangi pada sepuluh ekor serangga
vektor. Setelah proses inokulasi, bibit tomat dipelihara pada kurungan kedap
serangga dan kemunculan gejala khas infeksi ToCV dilakukan setiap hari.
Gambar 2 Tanaman tomat untuk pengujian retensi ToCV
Perbanyakan serangga vektor dilakukan dengan menginvestasikan imago
B. tabaci yang dikumpulkan dari pertanaman tomat di daerah Cisarua, Bogor pada tanaman tomat sehat di dalam kurungan serangga. Telur yang muncul dipelihara
pada kurungan yang sama sampai pupulasinya mencukupi sebagai bahan pengujian.
Tanaman yang digunakan dalam pengujian adalah benih tanaman tomat varietas
Martha yang di tanam di dalam tray yang berisi campuran tanah dan pupuk kandang dengan perbandingan 1:1. Setelah satu minggu bibit tanaman tomat
dipindahkan ke dalam polibag yang berisi campuran tanah dan pupuk. Satu minggu
kemudian tanaman tomat dapat digunakan untuk percobaan.
Untuk memastikan bahwa gejala klorosis yang muncul pada bibit tomat uji
disebabkan oleh infeksi ToCV dan bukan oleh faktor lain, maka dikonfirmasi
melalui reverse transcriptation-polymerase chain reaction (RT-PCR) yang dilakukan sebagai berikut: RNA total diekstraksi dari jaringan daun bibit tomat uji
dengan Rneasy Plant Mini Kits (Qiagen Inc., Chatsworth, CA., USA) mengikuti prosedur yang telah ditetapkan oleh Qiagen. RNA total yang telah diekstraksi
digunakan sebagai template dalam reaksi RT.
Reaksi RT dilakukan dengan total volume 10 µl yang mengandung 2 µl RNA
total, 1 µl buffer RT 10X, 0.35 µl 50 mM DTT (dithiothreitol), 2 µl 10 mM dNTP
(deoksiribonukleotida triphosphat), 0.35 µl M-MuLV Rev, 0.35 µlRNase inhibitor, 0.75 µl oligo (dT), dan 3.2 µl H2O. Reaksi RT dilakukan dalam sebuahAutomated
Thermal cycler(Gene Amp PCR System 9700; PE Applied Biosystem, USA) yang diprogram untuk satu siklus pada suhu 25 0C selama 5 menit, 42 0C selama 60
menit, dan 70 0C selama 15 menit. Complementary DNA hasil RT digunakan
sebagai template dalam reaksi PCR menggunakan pasangan primer yang telah
didesain khusus untuk mengamplifikasi ToCV yaitu ToCV-CF (5’-GTGTCAGGC
CATTGTAAACCAAG-3’) dan ToCV-CR (5’-CACAAAGCGTTTCTTTTCATA
AGCAGG-3’) dengan prediksi ukuran produk 360 bp.
Reaksi PCR dilakukan dengan total volume 25 µl, terdiri atas 1 µl primer
ToCV-CF, 1 µl primer ToCV-CR, 2.5 µl buffer PCR 10X + Mg2+, 0.5 µl 10 mM
dNTP, 0.3 µl Taq DNA polymerase, 18.7 µl H2O, dan 1 µl DNA template.
awal pada 94 0C selama 4 menit. Kemudian dilanjutkan dengan 30 siklus yang
terdiri dari denaturasi pada 94 0C selama 1 menit, penempelan primer (annealing) pada 620C selama 1 menit, dan pemanjangan pada 720C selama 2 menit. Khusus
untuk siklus terakhir, ditambahkan 10 menit pada 720C untuk tahapan sintesis, dan
siklus berakhir pada suhu 40C.
Produk PCR kemudian dielektroforesis pada 1.0% gel agarose. Elektroforesis
dilakukan dengan tegangan 50 volt selama 45 menit. Hasil elektroforesis
divisualisasikan dengantransluminator ultraviolet. Pita DNA yang terbentuk pada
hasil elektroforesis tersebut dipotret dengan menggunakan kamera digital.
Analisis Data
Data pengukuran panjang rostrum dan sayap B. tabaci diolah menggunakan Analisis Sidik Ragam (ANOVA) dengan program The Statistical Analysis System
(SAS) 9.0 for Windows. Pengaruh yang berbeda nyata akan dilakukan uji lanjut
HASIL DAN PEMBAHASAN
Identitas Kutukebul Pengkoloni Pertanaman Tomat
Kutukebul yang dikumpulkan dari pertanaman tomat di daerah Cisarua,
Bogor diperbanyak di tanaman tomat dalam kurungan kedap serangga dan
digunakan dalam penelitian ini telah berhasil diidentifikasi. Tubuh imago kutukebul
ini berwarna kuning dengan sayap yang ditutupi oleh sekresi berupa tepung
berwarna putih, dengan panjang tubuh 1.0-1.5 mm. Sayap terdiri dari dua pasang
dan transparan seperti tenda dengan posisi saat istirahat terlihat menyempit ke
depan (Gambar 3, kiri). Ciri-ciri tersebut sesuai yang disebutkan oleh Kalshoven
(1981) tentang ciri-ciri imagoB. tabaci.
Gambar 3 Morfologi B. tabaci. Imago (kiri) dan puparium (kanan): (1) basal tungkai tengah dan belakang, (2) ruas abdomen VII, (3) operculum, (4) vasiform orifice, (5) lingula, (6) caudal furrow, dan (7) caudal setae.
Identifikasi lebih lanjut yang dilakukan menggunakan kunci identifikasi
Martin (2000) berdasarkan morfologi puparium memastikan bahwa kutukebul ini
adalahB. tabaci. Ciri-ciri morfologi puparium yang ditemukan bersesuaian dengan B. tabaci adalah sebagai berikut: Puparium berbentuk bulat panjang, dengan bakal
mata terpisah. Mempunyai tujuh pasang rambut dorsal memanjang, trakea dengan
pinggiran seperti sisir terdiri dari gigi-gigi yang jelas, lingula memanjang membentuk lidah, tetapi bagian submargin tidak mempunyai barisan papila, serta
basal tungkai tengah dan belakang tidak berseta. Terdapat satu pasangcaudal setae pada ujung anal yang sama panjangnya.Vasiform orificeterdapat di daerah sebelum
0.3 mm 0.2 mm
ujung ujung posterior puparium, berbentuk segitiga, dan ukurannya lebih panjang
dari panjang caudal furrow. Operculum hampir seluruh bagian menutupi bagian vasiform orifice(Gambar 3, kanan).
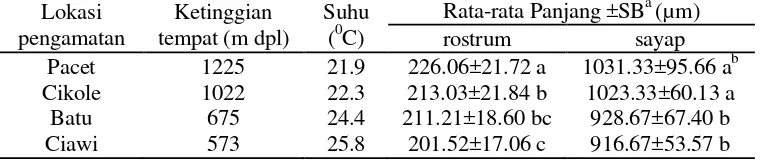
Variasi Panjang Rostrum dan Panjang SayapB. tabaci
Kutukebul B. tabaci yang diamati pada pertanaman tomat di daerah dengan ketinggian tempat yang berbeda memperlihatkan variasi panjang rostrum dan sayap
(Tabel 1). ImagoB. tabaciyang hidup di daerah dataran yang lebih tinggi memiliki
rostrum berukuran nyata lebih panjang dari imago B. tabaciyang hidup di daerah yang lebih rendah. Demikian juga ukuran sayap imago B. tabaci yang hidup di daerah dataran yang lebih tinggi nyata lebih panjang dibandingkan dengan imago
B. tabaciyang hidup di daerah yang lebih rendah.
Pengukuran yang dilakukan oleh Oliveira et al.(2004) juga memperlihatkan
hasil yang sama dengan penelitian ini yaitu pada Dalbulus maidis (Hemiptera: Cicadellidae). Wereng yang berada di dataran tinggi memiliki ukuran lebih besar,
bobot tubuh lebih berat, dan warna lebih gelap dibandingkan dengan spesies wereng
yang sama yang berada di dataran rendah. Variasi ukuran anggota tubuh wereng
tampaknya lebih dipengaruhi oleh perbedaan suhu lingkungan hidupnya. Menurut
Ayoade (1983) tinggi-rendahnya suatu daerah, mempengaruhi suhu pada daerah
tersebut. Semakin tinggi suatu tempat, maka suhu akan semakin rendah dan
intensitas cahaya semakin tinggi.
Seperti data yang disajikan dalam Tabel 1, terlihat bahwa B. tabaci yang ditemukan di daerah Pacet, daerah pengamatan dengan ketinggian tertinggi (1225 m
dpl) dan dengan kondisi suhu terendah (20.9 0C), mempunyai ukuran rostrum
sebesar 226.06±21.72 µm dan sayap sebesar 1031.33±95.66 µm. Kedua parameter
ini menunjukkan ukuran rostrum dan sayap terpanjang dibandingkan tempat
pengamatan lainnya. Murai & Toda (2002) juga menemukan bahwa imagoThrips tabaci yang pada stadia nimfanya berada pada suhu rendah memiliki bobot tubuh
yang lebih berat dibandingkan imago serangga yang pada stadia nimfanya berada
pada suhu tinggi.
Data yang ditabulasikan dalam Tabel 1 juga menunjukkan bahwa ukuran
pengamatan. Perbedaan ketinggian masing-masing tempat sudah dapat memberikan
pengaruh nyata terhadap panjang rostrum B. tabaci.Panjang sayapB. tabaciyang hidup di daerah Pacet tidak nyata berbeda dengan yang hidup di daerah Cikole.
Perbedaan ketinggian tempat daerah-daerah ini tampaknya belum cukup untuk
memberikan perbedaan pengaruh nyata terhadap panjang sayapB. tabaci. Hal yang sama juga terlihat pada sayapB. tabaciyang hidup di daerah Batu dan Ciawi.
Tabel 1 Panjang rostrum dan sayapB. tabaci
Lokasi pengamatan
Ketinggian tempat (m dpl)
Suhu (0C)
Rata-rata Panjang ±SBa(µm)
rostrum sayap
Pacet 1225 21.9 226.06±21.72 a 1031.33±95.66 ab
Cikole 1022 22.3 213.03±21.84 b 1023.33±60.13 a
Batu 675 24.4 211.21±18.60 bc 928.67±67.40 b
Ciawi 573 25.8 201.52±17.06 c 916.67±53.57 b
a
SB = Simpangan baku
b
angka yang diikuti huruf yang berbeda pada kolom yang sama menunjukkan hasil berbeda nyata (uji selang ganda Duncan α 0.05)
Periode Retensi ToCV dalam TubuhB. tabaci
Dalam penelitian ini dan juga penelitian yang dilakukan oleh Fitriasari (2010),
telah berhasil dilakukan penularan ToCV penyebab penyakit klorosis pada tanaman
tomat melalui satu individu imago B. tabaci. Membiarkan B. tabaci melakukan makan akuisisi pada tanaman tomat bergejala klorosis sebagai sumber ToCV sudah
cukup untuk menjadikan serangga tersebut menjadi infektif dan dapat menularkan
virus ke tanaman tomat baru. Lamanya periode infektifB. tabacidalam menularkan ToCV telah berhasil diukur dalam penelitian ini (Tabel 2). Periode retensi diukur
mulai saat serangga vektor menjadi infektif sampai tidak mampu lagi menularkan
virus. Pengukuran dilakukan dengan penularan berseri yaitu serangga vektor (dalam
hal ini B. tabaci) segera setelah menjadi infektif (setelah 48 jam periode makan
akuisisi) dipindahkan setiap 24 jam untuk makan inokulasi pada bibit tomat baru.
Kemampuan penularan dilihat dari muncul tidaknya gejala klorosis pada bibit tomat
yang diinokulasi.
Pada penelitian ini, gejala khas penyakit klorosis muncul pada bibit tomat uji
berkisar antara 2 sampai 3 minggu setelah inokulasi. Seperti disajikan pada Gambar
klorosis berwarna kuning terutama pada jaringan di antara tulang daun. Gejala yang
sama juga telah dilaporkan oleh Fitriasari (2010) yang menularkan ToCV pada
tomat varietas Martha. Menurut Accottoet. al(2001) gejala lain yang timbul pada
tanaman tomat di lapangan akibat infeksi ToCV dapat berupa daun nekrosis, daun
menggulung ke bawah, beberapa daun pucuk dapat berubah warna menjadi ungu,
diikuti dengan penurunan produksi buah. Kehilangan hasil terjadi karena area
fotosintesis pada daun berkurang.
Gambar 4 Tanaman tomat uji yang memperlihatkan gejala klorosis setelah diinokulasi ToCV melalui B. tabaci (kiri) dan yang tidak memperlihatkan gejala (kanan).
Dalam masa infektif, beberapa B. tabacimampu menularkan ToCV ke bibit tomat pada pemindahan ke-4, atau dengan kata lain periode retensinya mencapai 4
hari (Tabel 2). Namun demikian, kebanyakanB. tabacimampu menularkan ToCV hanya sampai hari ke-3. Sampai saat ini belum ada laporan tentang periode retensi
B. tabaciterhadap virus ToCV.
Tabel 2 Masa infektif Bemisia tabaci dalam penularan berseri Tomato chlorosis virusa
Pengamatan hari ke
Tanaman yang diinokulasi dan reaksinya
A B C D E F G H I J
1 + + + + + + + + + +
2 + + + + + + + + + +
3 + + + + + - + + +
-4 - - + - + - - - +
-5 - - -
-6 - - mt - - -
-7 - mt mt - - mt - mt -
-a
Periode retensi B. tabaci terhadap ToCV hasil penelitian ini lebih singkat dibandingkan hubungan geminivirus dengan serangga vektornya, misalnyaTomato yellow leaf curl virus(TYLCV) dengan vektorB. tabaci.Hasil penelitian Sulandari
(2004), menyatakan periode retensi B. tabaci terhadap TYLCV mencapai 6 hari. Penelitian periode retensi B. tabaci terhadap Squash leaf curl virus(SLCV), hasil penelitian Cohenet al. (1983) menunjukkan hasil periode retensi yang cukup lama yakni mencapai 26 hari. Stenger et al.(1990) menyatakan bahwaB. tabaci hanya mampu menahanPepper leaf curl virus(PepLCV) dalam tubuhnya selama 10 hari,
sedangkan Idris & Brown (1998) menemukan periode retensi B. tabaci terhadap Sinaloa tomato leaf curl virus (STLCV) lebih dari 9 hari dan terputus-putus. Adanya perbedaan periode retensi yang cukup jauh dari hasil penelitian ini dengan
penelitian lainnya di atas karena adanya perbedaan dari sifat virus. Kelompok
ToCV yang digunakan dalam penelitian ini bersifat semipersisten, sedangkan
penelitian lain menggunakan virus yang persisten dalam tubuh B. tabaci. Perbandingan ini dilakukan karena belum adanya laporan periode retensi virus
golongan crinivirus terhadap vektornya.
Imago B. tabaciyang digunakan dalam penelitian ini mempunyai lama hidup sekitar 8 hari. Menurut Kurniawan (2007) imago B. tabaci biotipe-B yang
diperbanyak di rumah kaca dapat hidup sampai hari ke-20. Hal ini mungkin
disebabkan adanya zat antiviral di dalam tubuhnya yang berasal dari tanaman yang
berpengaruh negatif atau pengaruh langsung dari virus pada serangga vektornya
(Cohenet al.1983). Antiviral yang berasal dari tanaman selain berpengaruh negatif pada serangga vektornya, juga dapat menurunkan konsentrasi virus yang terdapat di
dalam tubuh serangga. Menurut Cohenet al.(1983), konsentrasi virus menurun 1-2% per hari sampai hari ke-20. Keberadaan virus di dalam tubuh kutukebul juga
menyebabkan lama hidupnya turun sekitar 25% (Sulandari 2004). Selain itu, ada
faktor abiotik atau faktor lingkungan yang mempengaruhi lama hidup kutukebul,
salah satunya adalah suhu lingkungan. Menurut Subagyo (2010), peningkatan suhu
40C (dari 250C ke 290C) akan memperpendek siklus hidupB. tabacipada tanaman tomat.
genom berupa RNA utas tunggal RNA yaitu RNA 1 dan RNA 2 yang
masing-masing berukuran 7.8 dan 8.2 kb. Crinivirus merupakan kelompok virus yang penyebarannya terbatas pada jaringan floem dan terakumulasi pada tingkat rendah
pada tanaman yang terinfeksi. Oleh karena itu, pembuatan antiserum sulit dilakukan
dan sampai saat ini belum tersedia antiserum untuk deteksi ToCV. Pada penelitian
ini, deteksi virus ini dilakukan melalui pendekatan molekuler yaitu melalui
RT-PCR. Deteksi dengan RT-PCR memerlukan sepasang primer yang didesain khusus
untuk mendeteksi virus tersebut. Pasangan primer yang digunakan dalam penelitian
ini telah didesain khusus berdasarkan analisa sekuen ToCV yang diunduh dari
GenBank.
M K+ D3 D4
K-Gambar 5 Hasil elektroforesis menggunakan pasangan primer spesifik ToCV-CF dan ToCV-CR. RNA diekstraksi dari sampel tanaman tomat yang positif terinfeksi oleh ToCV (K+), sampel bibit tomat uji yang memperlihatkan gejala klorosis (D3) dan yang tidak memperlihatkan gejala (D4) setelah diinokulasi, dan sampel tanaman tomat sehat (K-). M adalah marker 100 bp DNA ladder.
Pada penelitian ini, keberhasilan penularan ToCV melalui imago B. tabaci dilihat dari kemunculan gejala klorosis pada bibit tomat uji. Untuk memastikan
bahwa gejala klorosis tersebut disebabkan oleh karena keberadaan ToCV dalam
jaringan tanaman maka dilakukan verifikasi melalui RT-PCR yang hasilnya
disajikan pada Gambar 5.
RT-PCR yang telah dilakukan dengan menggunakan pasangan primer
CGTTTCTTTTCATAAGCAGG-3’] berhasil mengamplifikasi DNA berukuran 360
bp. Produk PCR ini sesuai dengan prediksi berdasarkan sikuen ToCV isolat
NC007341 yang berasal dari Florida, USA (Wintermantel et al. (2005). Seperti
terlihat pada Gambar 4, hasil RT-PCR dari sampel bibit tomat uji yang bergejala
klorosis memperlihatkan pita DNA berukuran 360 bp, sama dengan hasil RT-PCR
dari sampel tanaman tomat yang sudah diketahui terinfeksi ToCV (kontrol positif).
Hasil penelitian ini memverifikasi bahwa bibit tomat yang menunjukkan
gejala klorosis setelah diinokulsi adalah benar disebabkan oleh keberadaan ToCV di
dalam jaringannya. RT-PCR dari sampel tanaman tomat sehat yang tidak
KESIMPULAN DAN SARAN
Kesimpulan
Ukuran panjang rostrum dan sayap B. tabaci bervariasi pada ketinggian tempat dan suhu lingkungan yang berbeda. Ukuran rostrum B. tabaci yang ditemukan di daerah Pacet adalah 226.06±21.72 µm, Cikole 213.03±21.84 µm,
Batu 211.21±18.60 µm, Ciawi 201.52±17.06 µm. Ukuran sayap B. tabaci yang
ditemukan di daerah Pacet 1031.33±95.66 µm, Cikole 1023.33±60.13 µm, Batu
928.67±67.40 µm, dan di Ciawi 916.67±53.57 µm.
Kutukebul B. tabacimasih infektif menularkan ToCV pada hari ke-4 setelah periode makan akuisisi, atau periode retensi ToCV pada tubuhB. tabaci adalah 4 hari.
Saran
Perlu dilakukan penelitian lanjutan tentang pengukuran tubuhB. tabacibagian lain, pada inang tanaman selain tomat dan kisaran tinggi tempat yang lebih luas,
yaitu <500 m dpl dan >1200 m dpl. Selain itu perlu dilakukan penelitian tentang
periode retensi ToCV dengan menggunakan kutukebul vektor ToCV dan varietas
DAFTAR PUSTAKA
[BMKG] Badan Meteorologi Klimatologi dan Geofisika. 2011. Data Temperatur dan Kelembaban. Bogor: BMKG.
[CABI] Centre for Agriculture and Bioscience International. 2005. Corp protection compendium 2005 [CD-ROM]. Wallingford, UK: CAB International.
Accotto GP, Vaira AM, Vecchiati M, Finetti Sialer MM, Gallitelli D, Davino M. 2001. First report of tomato chlorosis virus in Italy. Plant Disease85:1208.
Aidawati N, Hidayat SH, Suseno R, Sosromarsono S. 2002. Transmission of an Indonesian isolate ofTobacco leaf curl virus(Geminivirus) byBemisia tabaci Genn. (Hemiptera: Aleyrodidae). Plant Pathology18:231-236.
Ayoade JO. 1983. Introduction to Climatology for the Topics. John Wiley & Sons, Ney York.
Borror DJ, Triplehorn, Johnson. 1992. An Introduction to the Study of Insects. Edisi ke-6. New York: Saunders College Publishing.
Cohen S, Duffus JE, Larsen RC, Liu HY, Flock RA. 1983. Purification, serology, and vector relationships of Squash leaf curl virus a whitefly transmitted geminivirus. Phytopathology3:1669-1673.
Duffus JE, Liu H-Y, Wisler GC. 1996. Tomat infectious chlorosis virus-a new clostero-like virus transmitted by Trialeurodes vaporariorum. European Journal of Plant Pathology102:219-226.
Fereres A, Moreno A. 2009. Behavioural aspects influencing plant virus transmission by homopteran insects.Virus Research141:158-168.
Fitriasari ED. 2010. Keefektifan kutukebul dalam menularkan virus penyebab penyakit kuning pada tanaman tomat. [tesis]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Gray SM, Banerjee N. 1999. Mechanism of arthropod transmission of plant and animal viruses. Microbiol. Mol. Biol. Rev.3:128-148.
Hartono S, Wijonarko A. 2007. Karakterisasi Biologi Molekuler Tomato Infectious Chlorosis Virus Penyebab Penyakit Kuning pada Tanaman Tomat di Indonesia. Jurnal Akta Agrosia Edisi Khusus2:139-146.
Hirota T, Natsuaki T, Murai T, Nishigawa H, Niibori K, Goto K, Hartono S, Suastika G, Okuda S. 2010. Yellowing disease of tomato caused by Tomato chlorosis virus newly recognized in Japan. J Gen Plant Pathology 76:168-171.
Idris AM, Brown JK. 1998. Sinaloa tomato leaf curl geminivirus: biological and and moleculer evidence for a new subgroup III virus. Phytopathol 88:648-657.
Jacquemond M, Verdin E, Dalmon A, Guilbaud L, Gognalons P. 2008. Serological and molecular detection of Tomato chlorosis virus and Tomato infectious chlorosis virusin tomato. Plant Pathology58:1365:3059.
Jones DR. 2003. Plant viruses transmitted by whitefly. European Journal of plant pathology109:195-219.
Kalshoven LGE. 1981. The Pests of Crops in Indonesia. Laan PA van der, penerjemah. Jakarta: Ichtiar Baru-van Hoeve. Terjemahan dari: De Plagen van de Cultuurgewassen in Indonesie.
Katanya ARA, Stavridou E, Farhan K. Livieratos IC. 2008. Nucleotide sequence analysis and detection of a Greek isolate of tomato chlorosis virus. Plant Pathology57:819-824.
Kennedy JS, Day MF, Eastop VF. 1962. A Conspectus of Aphids as Vectors of Plant Viruses. Commonwealth Institute of Entomology, London.
Kingsolver JG, Huey RB. 2008. Size, temperature, and fitness: three rules. Evolutionary Ecology Research10:251-268.
Kurniawan, HA. 2007. Neraca kehidupan kutukebul, Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) biotipe-B dan non-B pada tanaman mentimun (Curcumis sativus L) dan cabai (Capsicum annuum L.). [Tesis]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Lozano GE, Moriones E, Navas-Castillo J. 2006. Complete nucleotide sequence of the RNA2 of the crinivirus tomato chlorosis virus. Archives of Virology 151:581-587.
Markham PG, Bedford ID, Liu S, Pinner MS. 1994. The transmission of geminiviruses byBemisia tabaci. Pesticide Science42:123-128.
Martin JH, Misfud D, Rapisarda C. 2000. The whiteflies (Hemiptera: Aleyrodidae) of Europe and Mediteranian basin. Buletin of Entomological Research 90:407-448.
Masse D, lefeuvre P, Delatte H, Karime ALA, Hostachy B, Reynaud B, Lett JM. 2008. Tomato chlorosis virus: first report in Mayotte Island. Plant Pathlogy 57:388.
Mock. 1969. Land Capability Appraisal in Indonesia. Soil Research Institute, Bogor.
Morales FJ. 2001. Conventional breeding of resistence to Bemisia tabaci-transmitted geminiviruses.Crop Prot20:825-843.
Muniyapa V, Reddy DVR. 1983. Transmission of Cowpea mild mottle virus by Bemisia tabaciin non persistent manner. Plant Dis67:391-393.
Murai T, Toda S. 2002. Variation of Thrips tabaci in colour and size: Thrips and Tospoviruses: Proceedings of the 7th International Symposium on Thysanoptera Calabria. Australian National Insect Collection Canberra, 377– 378.
Nault LR, Ammar ED. 1989. Leafhopper and planthopper transmission of plant viruses. AnnRev Entomol34:503-529.
Navas-Castillo J, Camero R, Bueno M, Moriones E. 2000. Severe yellowing outbreaks in tomato in Spain associated with Infections of Tomato chlorosis virus. Plant Disease84:835-837.
Oliveira CM, Lopes JRS, Dias CTDS, Nault LR. 2004. Influence of latitude and elevation on polymorphism among populations of the corn leafhopper, Dalbulus maydis(DeLong and Wolcott) (Hemiptera: Cicadellidae), in Brazil. Enviromental Entomology33(5):1192-1199.
Prado SS, Hung KY, Daugherty MP, Almeida RPP. 2010. Indirect effect of temperature on stink bug fitness, via maintenance of gut-associated simbionts. Applied and Environmental Microbiology:1261-1266.
Segev L, Wintermantel WM, Polston JE, and Lapidot M. 2004. First report of tomato chlorosis virus in Israel. Plant Disease88:1160.
Smith PE. 2009. Whitefly: identification and biologyin New Zealand greenhouse tomato crops. Factsheet 1. Horticulture New Zealand. Fresh tomato product group.
Subagyo VNO. 2010. Neraca kehidupan kutukebul, Bemisia tabaci(Gennadius) (Hemiptera: Aleyrodidae) pada tanaman tomat (Lycopersicon esculentum Mill.), tanaman cabai (Capsicum annuum L.), dan gulma babadotan (Ageratum conyzoides L.) pada suhu 25 °C dan 29 °C. [skripsi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Sulandari S. 2004. Karakterisasi biologi, serologi, dan analisis sidik jari DNA virus penyebab penyakit daun keriting kuning cabai. [disertasi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Sylvester ES. 1956. Beet yellows virus transmission by the Green peach aphid. J Econ Entomol49:789-800.
Tsai WS, Shih SL, Dreen SK, Hanson P. 2004. First report of the occurrence of tomato chlorosis virus and tomato infectious chlorosis virus in Taiwan. Plant Disease88:311.
Usman K. 2003. Kemampuan hidup Bemisia tabaci Genn. (Hemiptera: Aleyrodidae) pada tanaman tomat (Lycopersicon esculentumMill.) dan cabai (Capsicum annuum Linn.). [skripsi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Wintermantel WM. 2004. Emergence of greenhouse whitefly (Trialeurodes vaporariorum) transmitted crinivirus as threats to vegetable and fruit production in North America. ASP: San Diego.
Wintermantel WM, Wisler GC, Anchieta1 AG, et al. 2005. The complete nucleotide sequence and genome organization of tomato chlorosis virus. Arch Virol150:2287–2298.
Wintermantel WM, Wisler GC. 2006. Vector specificity, host range, and genetic diversity of tomato chlorosis virus. Plant Disease90:814-9.
Wisler GC, Duffus JE, Liu H-Y, Li RH. 1998a. Ecology and epidemiology of whitefly-transmitted closteroviruses. Plant Disease82:270–80.
Wisler GC, Duffus JE,Li RH, Liu H-Y, Lowry DS. 1998b. Tomato chlorosis virus: a new whitefly-transmitted, phloem-limited, bipartite closterovirus of tomato. Phytopathology88:402–9.
Tabel Lampiran1 Panjang rostrum dan panjang sayap B. tabaci dari Kecamatan Pacet, Kabupaten Cianjur
Individu Panjang sayap (µm) Panjang rostrum (µm)
1 1160 236.36
2 1120 236.36
3 1160 254.55
4 980 218.18
5 980 209.09
6 1140 227.27
7 1140 254.55
8 1000 227.27
9 1020 236.36
10 920 200.00
11 1120 227.27
12 1140 263.64
13 1040 236.36
14 1140 227.27
15 1100 236.36
16 1000 200.00
17 900 245.45
18 1120 227.27
19 840 181.82
20 960 227.27
21 900 254.55
22 1000 218.18
23 980 181.82
24 840 227.27
25 1000 236.36
26 1120 236.36
27 1100 245.45
28 1060 200.00
29 960 181.82
30 1000 227.27
Total 30940 6781.82
[image:36.611.126.505.110.594.2]Tabel Lampiran 2 Panjang rostrum dan panjang sayap B. tabaci dari Kecamatan Cikole, Kota Sukabumi
Individu Panjang sayap (µm) Panjang rostrum (µm)
1 1180 209.09
2 1060 218.18
3 1160 227.27
4 980 181.82
5 1000 190.91
6 1020 209.09
7 1000 245.45
8 960 200.00
9 980 227.27
10 1040 236.36
11 1020 227.27
12 1000 218.18
13 1060 227.27
14 1080 227.27
15 1000 190.91
16 920 172.73
17 1020 190.91
18 980 190.91
19 960 181.82
20 920 172.73
21 960 227.27
22 1040 190.91
23 1040 236.36
24 1000 218.18
25 1080 227.27
26 1020 227.27
27 1040 245.45
28 1100 245.45
29 1080 209.09
30 1000 218.18
Total 30700 6390.91
[image:37.611.116.505.111.576.2]Tabel Lampiran 3 Panjang rostrum dan panjang sayap B. tabaci dari Kecamatan Batu, Kota Batu
Individu Panjang sayap (µm) Panjang rostrum (µm)
1 900 200.00
2 900 218.18
3 900 218.18
4 960 200.00
5 1020 209.09
6 1040 209.09
7 900 218.18
8 1020 200.00
9 900 218.18
10 860 200.00
11 1000 218.18
12 900 218.18
13 840 209.09
14 900 190.91
15 960 236.36
16 1000 227.27
17 880 200.00
18 920 200.00
19 900 227.27
20 980 227.27
21 960 227.27
22 880 227.27
23 800 181.82
24 980 245.45
25 840 181.82
26 1000 236.36
27 1000 236.36
28 1020 200.00
29 800 181.82
30 900 172.73
Total 27860 6336.36
[image:38.611.116.505.111.593.2]Tabel Lampiran 4 Panjang rostrum dan panjang sayap B. tabaci dari Kecamatan Ciawi, Kabupaten Bogor
Individu Panjang sayap (µm) Panjang rostrum (µm)
1 975 218.18
2 1012.5 236.36
3 987.5 190.91
4 937.5 218.18
5 962.5 218.18
6 900 200.00
7 837.5 181.82
8 950 200.00
9 787.5 163.64
10 887.5 218.18
11 862.5 190.91
12 837.5 209.09
13 812.5 190.91
14 950 190.91
15 850 181.82
16 925 209.09
17 887.5 200.00
18 900 200.00
19 912.5 209.09
20 937.5 200.00
21 937.5 218.18
22 962.5 218.18
23 950 200.00
24 937.5 209.09
25 925 200.00
26 975 236.36
27 900 172.73
28 962.5 190.91
29 937.5 190.91
30 900 181.82
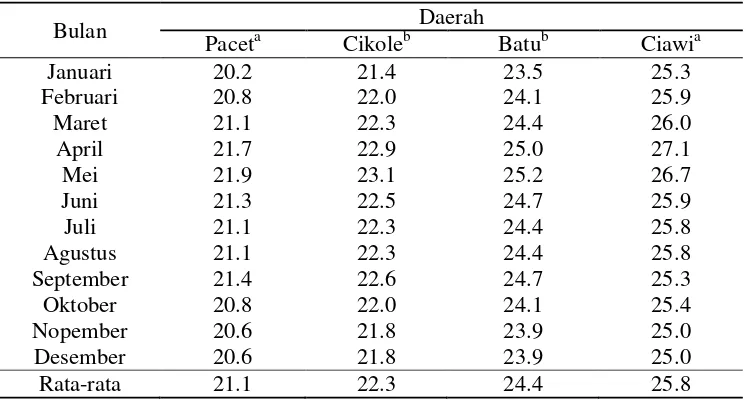
Total 27500 6045.45
[image:39.611.106.503.109.589.2]Tabel Lampiran 5 Data temperatur (0C) bulanan tahun 2010
Bulan Daerah
Paceta Cikoleb Batub Ciawia
Januari 20.2 21.4 23.5 25.3
Februari 20.8 22.0 24.1 25.9
Maret 21.1 22.3 24.4 26.0
April 21.7 22.9 25.0 27.1
Mei 21.9 23.1 25.2 26.7
Juni 21.3 22.5 24.7 25.9
Juli 21.1 22.3 24.4 25.8
Agustus 21.1 22.3 24.4 25.8
September 21.4 22.6 24.7 25.3
Oktober 20.8 22.0 24.1 25.4
Nopember 20.6 21.8 23.9 25.0
Desember 20.6 21.8 23.9 25.0
Rata-rata 21.1 22.3 24.4 25.8
a
Sumber: Badan Meteorologi Klimatologi dan Geofisika
b
Bemisia tabaci
GENNADIUS (HEMIPTERA: ALEYRODIDAE)
PADA TANAMAN TOMAT (Lycopersicum esculentum
MILL.):
PANJANG ROSTRUM DAN SAYAP PADA BEBERAPA
KETINGGIAN TEMPAT SERTA PERIODE RETENSI
Tomato chlorosis virus
(ToCV)
HERLIE ARIFEBRIAWAN
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
ABSTRAK
HERLIE ARIFEBRIAWAN,Bemisia tabaciGennadius (Hemiptera: Aleyrodidae) pada Tanaman Tomat (Lycopersicum esculentumMill.): Panjang Rostrum dan Sayap pada Beberapa Ketinggian Tempat serta Periode Retensi Tomato chlorosis virus (ToCV). Dibimbing olehDEWI SARTIAMIdanGEDE SUASTIKA.
Bemisia tabacidigolongkan ke dalam ordo Hemiptera, subordo Sternorrhyncha, superfamili Aleyrodoidea, dan termasuk ke dalam famili Aleyrodidae. Faktor lingkungan dapat mempengaruhi ukuran panjang tubuh kutukebul ini. Serangga yang berada di dataran tinggi memiliki ukuran lebih besar, bobot tubuh lebih berat, dan warna lebih gelap dibandingkan dengan spesies serangga yang sama yang berada di dataran rendah. Sepanjang abad ke-20, B. tabaci telah menjadi vektor patogen-patogen tertentu baik di rumah kaca maupun di lahan terbuka di wilayah beriklim hangat. Penyakit Tomato chlorosis virus (ToCV) adalah salah satu penyakit yang ditularkan olehB. tabaci.Penyakit ToCV ini termasuk baru yang dikategorikan “new emerging disease” akibat pengaruh pemanasan global. Penelitian ini bertujuan untuk (1) mengetahui variasi panjang rostrum dan sayap B. tabaciberdasarkan ketinggian tempat hidupnya dan (2) mengukur periode retensi ToCV yang ditularkanB. tabaci. Penelitian ini dilakukan dengan pengukuran panjang rostrum dan panjang sayap B. tabacidari beberapa tempat dengan ketinggian berbeda. SampelB. tabaciberasal dari Ciawi (Bogor) ketinggian 573 m dpl (di atas permukaan laut), Pacet (Cianjur) ketinggian 1225 m dpl, Batu (Batu) ketinggian 675 m dpl dan Cikole (Sukabumi) ketinggian 1022 m dpl. Hasil penelitian menunjukkan bahwaB. tabaciyang memiki panjang rostrum dan panjang sayap yang berbeda-beda di setiap tempat. Panjang rostrum dari daerah Pacet 226.06±21.72 µm, Cikole 213.03±21.84 µm, Batu 211.21±18.60 µm, Ciawi 201.52±17.06 µm. Panjang sayap dari daerah Pacet 1031.33±95.66 µm, Cikole 1023.33±60.13 µm, Batu 928.67±67.40 µm, Ciawi 916.67±53.57 µm. Penelitian yang lain adalah mengakuisisi B. tabaci ke dalam kurungan serangga yang berisi tanaman tomat yang positif virus ToCV dan sebagian lagi dimasukkan ke dalam kurungan serangga yang berisi tanaman tomat yang sehat. Periode makan akuisisi selama 48 jam. Seekor imagoB. tabaciyang telah di akuisisi dipindahkan ke tanaman uji. Setiap 24 jamB. tabacitersebut dipindahkan ke tamanan uji berikutnya secara berseri selama tujuh hari. Percobaan dilakukan dengan 10 ulangan dari B. tabaci yang di-akuisisi ke tanaman tomat bervirus ToCV dan 10 ulangan dari B. tabaci yang di-akuisisi ke tanaman tomat sehat. Hasil penelitian menunjukkan bahwa B. tabaci viruliferus dalam seri inokulasi ke tanaman tomat mampu menularkan virus hingga hari ke-4, hal ini dilihat dari timbulnya gejala ToCV pada tanaman tomat tersebut. Untuk konfirmasi bahwa gejala klorosis yang muncul pada tanaman tomat uji disebabkan oleh infeksi ToCV dilakukan dengan RT-PCR.
PENDAHULUAN
Latar Belakang
Bemisia tabaci digolongkan ke dalam ordo Hemiptera, subordo Sternorrhyncha, superfamili Aleyrodoidea, dan termasuk ke dalam famili
Aleyrodidae (Borror et al. 1992). Spesies B. tabaci merupakan kutukebul yang memiliki kisaran inang luas. Kalshoven (1981), mengelompokkan tanaman inang
dari serangga ini meliputi beberapa famili, yaitu famili Compositae, Cucurbitaceae,
Cruciferae, dan Solanaceae. Menurut Hill (1987), tanaman inang utama B. tabaci
adalah kapas, tembakau, tomat, ubi jalar, ubi kayu, beberapa jenis gulma, serta
tanaman lain yang dapat menjadi inang alternatif. Hal ini menyebabkan banyaknya
nama umum B. tabaci yang dikenal luas, diantaranya adalah kutukebul kapas (cotton whitefly), kutukebul tembakau (tobacco whitefly), dan kutukebul ubi jalar (sweetpotato whitefly) (Kalshoven 1981).
Imago B. tabaciberwarna kuning dengan panjang tubuh 1-1.5 mm dan sayap yang tertutup oleh tepung berwarna putih. Lama hidup imago bervariasi tergantung
faktor lingkungan. Lama hidup imago betina hingga 16 hari, sedangkan imago
jantan umurnya lebih singkat dibandingkan imago betina, yaitu sekitar 9-15 hari
(CABI 2005).
Suhu tubuh serangga sangat dipengaruhi oleh suhu lingkungan tempat
serangga tersebut hidup. Serangga beradaptasi dengan suhu lingkungan untuk
menjaga kebugaran tubuhnya. Salah satu bentuk adaptasi serangga adalah dengan
adanya perbedaan ukuran tubuh (Kingsolver 2008). Serangga yang hidup pada suhu
rendah memerlukan nutrisi yang lebih banyak dan memiliki metabolisme yang lebih
lambat dibandingkan serangga yang hidup pada suhu tinggi. Serangga yang
memiliki metabolisme rendah mempunyai ukuran tubuh lebih panjang (Prado
2010). Menurut Oliveira et al. (2004) serangga yang berada di dataran tinggi memiliki ukuran lebih besar, bobot tubuh lebih berat, dan warna lebih gelap
dibandingkan dengan spesies serangga yang sama yang berada di dataran rendah.
Data mengenai variasi morfologi B. tabaci berdasarkan ketinggian tempat belum ada, sehingga diperlukan penelitian mengenai pengetahuan variasi morfologi
Beberapa tahun belakangan,B. tabacitelah menjadi masalah utama bagi para petani di seluruh dunia. WalaupunB. tabacidianggap sebagai grup serangga tropis, spesies berbahaya ini banyak sekali ditemukan di seluruh belahan lain dunia,
terutama di daerah beriklim subtropis. Sepanjang abad ke-20, B. tabaci telah menjadi vektor patogen-patogen tertentu baik di rumah kaca maupun di lahan
terbuka di wilayah beriklim hangat (Martin et al. 2000). Stadia nimfa dan imago kutukebul merupakan stadia yang menyebabkan kerusakan tanaman (Morales
2001).
Tomato chlorosis virus (ToCV) adalah salah satu virus tanaman yang ditularkan oleh B. tabaci. ToCV diketahui tidak dapat ditularkan melalui cairan perasan tanaman sakit ataupun melalui benih. ToCV ditularkan ke dalam jaringan
tanaman oleh kutukebul Bemisia tabaci biotipe A dan B, Trialeurodes abutilonea (Wisler et al. 1998b), dan T. vaporariorum (Wintermantel & Wisler 2006).
Menurut Wisler & Duffus (2001) B. tabaci merupakan kutukebul yang memiliki efisiensi menularkan ToCV lebih baik dibanding kutukebul yang lainnya.
ToCV ini termasuk baru yang dikategorikan “new emerging disease” akibat
pengaruh pemanasan global (Segev et al. 2004). ToCV pertama kali tersebar di negara bagian Florida, USA sejak tahun 1989. Virus ini dengan cepat menyebar ke
seluruh dunia dan sampai saat ini keberadaannya telah dilaporkan di banyak negara
seperti di Perancis (Masseet al. 2008), Spanyol (Navas-Castilloet al. 2000; Lozano et al.2006), Taiwan (Tsaiet al.2004), dan Yunani (Katanyaet al.2008). Menurut Hartono & Wijonarko (2007), penyakit ini pada tanaman tomat telah menyebar di
sentra pertanaman tomat di Magelang, Jawa Tengah dan Purwakarta, Jawa Barat.
Penyakit ToCV ini mempunyai gejala menguning pada bagian interval daun
(Duffuset al. 1996, Hirotaet al. 2010), bintik-bintik nekrotik kecil (Wintermantel & Wisler 2006), mengeriting (Hirota et al. 2010), dan gejala lanjutan akan menyebabkan daun tampak berwarna merah kecoklatan (Wisler et al. 1998a). Menurut Wintermantel et al. (2005), ToCV adalah virus yang berbentuk panjang
lentur (flexuous filamentous) dengan ukuran diameter 12 nm dan panjang rata-rata 800-850 nm.
perlu dilakukan untuk mengetahui lama waktu retensi virus ToCV dalam tubuh
serangga vektorB. tabaci.
Tujuan Penelitian
Penelitian ini bertujuan untuk (1) mengetahui variasi panjang rostrum dan
sayapB. tabaciberdasarkan ketinggian tempat hidupnya dan (2) mengukur periode retensi ToCV yang ditularkannya.
Manfaat Penelitian
Hasil penelitian diharapkan dapat memberikan informasi dasar mengenai
perbandingan morfologi yang mencakup panjang rostrum dan sayap kutukebul
B. tabaci pada beberapa ketinggian tempat yang berbeda dan lama periode retensi kutukebul B. tabaci yang berperan sebagai vektor ToCV dalam tanaman
TINJAUAN PUSTAKA
Bemisi tabaci
Kutukebul B. tabaci digolongkan ke dalam ordo Hemiptera, subordo Sternorrhyncha, superfamili Aleyrodoidea, dan termasuk kedalam famili Aleyrodidae
(Bororet al. 1992). SeranggaB. tabacimerupakan spesies kutukebul yang memiliki kisaran inang luas. Kalshoven (1981), mengelompokkan tanaman inang dari serangga
ini meliputi beberapa famili, yaitu famili Compositae, Cucurbitaceae, Cruciferae, dan
Solanaceae. Menurut Hill (1987), tanaman inang utama B. tabaci adalah kapas,
tembakau, tomat, ubi jalar, ubi kayu, beberapa jenis gulma, serta tanaman lain yang
dapat menjadi inang alternatif. Hal ini menyebabkan banyaknya nama umum
B. tabaci yang dikenal luas, diantaranya adalah kutukebul kapas (cotton whitefly), kutukebul tembakau (tobacco whitefly), dan kutukebul ubi jalar (sweetpotato whitefly) (Kalshoven 1981).
Imago B. tabacimenurut Usman (2003) lebih menyukai daun tanaman tomat dibandingkan dengan daun tanaman cabai. Permukaan daun tanaman tomat secara
genetik memiliki rambut-rambut daun yang banyak sehingga dapat meningkatkan
suhu dan kelembaban mikro dan mengakibatkan pertumbuhan dan perkembangan
serta kehidupan serangga ini lebih kondusif. Selain itu, kondisi permukaan daun
tanaman tomat tersebut lebih memberi kenyamanan serangga ini untuk melakukan
aktivitas makan dan bertelur karena serangga ini lebih mudah menjangkarkan
tungkai-tungkainya dibandingkan pada tanaman cabai yang memiliki permukaan
daun yang lebih licin.
Menurut Martin (2000), ciri morfologi B. tabaci adalah sebagai berikut: Telur
yang baru diletakkan berwarna kekuningan dan biasanya tertutup lilin, warna telur akan
berubah setelah 24 jam menjadi berwarna coklat. Nimfa instar satu berbentuk bulat
panjang, berwarna