TOLERANCE DIFFERENCE BETWEEN TWO SPECIES
(
Perna viridis
AND
Vasticardium
cf
. flavum
) UNDER HEAT AND
HYPOXIA STRESS
NURINA AYU
MARINE SCIENCE PROGRAM POST GRADUATE SCHOOL INSTITUT PERTANIAN BOGOR
ISSUES RELATED WITH THIS THESIS AND THE SOURCE
OF INFORMATION
With this I declare that the thesis of “Tolerance difference between two adapted species (Perna viridis and Vasticardium cf. flavum) under heat and hypoxia stress” is my own work under direction by advisory committee and never been submitted at any other universities. Information sources cited from other authors which is published and unpublished have been mentioned in the texts and listed in references at the end of this thesis.
Bogor, August 2012
ABSTRAK
NURINA AYU. PERBEDAAN TOLERANSI ANTARA DUA SPESIES (Perna
viridis DAN Vasticardium cf. flavum) DI BAWAH PAPARAN STRES TERMAL
DAN HIPOKSIA di bawah bimbingan NEVIATY PUTRI ZAMANI dan KAREN VON JUTERZENKA.
Perubahan lingkungan merupakan salah satu faktor yang memberikan tekanan berat bagi organisme sesil seperti Bivalvia. Faktor lingkungan yang vital adalah temperatur dan ketersediaan oksigen di perairan. Temperatur perairan yang tinggi dan kondisi hipoksia akan mempengaruhi performa Perna viridis dan
Vasticardium cf. flavum. Hipoksia (DO 0,5 mg/l) secara signifikan menyebabkan
stres pada kedua spesies (pperna < 0,01; pvasticardium < 0,01), dan P. viridis memiliki
ketahanan yang lebih tinggi dibandingkan V. cf. flavum dalam menghadapi
hipoksia (p < 0,05). Temperatur tinggi (34°C) juga secara signifikan
mengakibatkan stres pada kedua spesies (pperna < 0,01; pvasticardium < 0,05), meski
tidak ada perbedaan signifikan ketika kedua spesies dipaparkan pada temperatur tinggi (p > 0,05). Dapat disimpulkan bahwa meskipun hipoksia dan temperatur tinggi adalah stressor bagi kedua spesies, namun P. viridis memiliki daya
toleransi yang lebih baik terhadap hipoksia dibandingkan V. cf. flavum;
sementara temperatur tinggi di perairan memiliki dampak yang sama terhadap kemampuan kedua spesies dalam bertahan.
Kata kunci: temperatur, hipoksia, ketahanan hidup, respon, toleransi, Perna
ABSTRACT
NURINA AYU. TOLERANCE DIFFERENCE BETWEEN TWO SPECIES
(Perna viridis AND Vasticardium cf. flavum) UNDER HEAT AND HYPOXIA
STRESS under direction of NEVIATY PUTRI ZAMANI and KAREN VON JUTERZENKA
The environmental shifting is one major stress to sessile organisms such Bivalves. Two of most vital environmental parameters are temperature and oxygen availability. High water temperature and low oxygen concentration leading to hypoxia conditions will affecting the performance of Perna viridis and
Vasticardium cf. flavum. Hypoxia (DO 0,5 mg/l) significantly causing stress to
both species (pperna < 0,01; pvasticardium < 0,01), and P. viridis has higher tolerance
to hypoxia compare to V. cf. flavum (p < 0,05). Heat stress (34°C) also
significantly causing stress to both (pperna < 0,01; pvasticardium < 0,05), though the
values of their survivorship was not differ significantly when they were compared (p > 0,05). Based on the result, can be concluded that even both factors are
stressors for each species, P. viridis has better tolerance to hypoxia than V. cf.
flavum; as in heat stress, both species undergo the same impact in attempt to survive.
Keywords: temperature, hypoxia, survival, response, tolerance, Perna viridis,
SUMMARY
NURINA AYU. TOLERANCE DIFFERENCE BETWEEN TWO SPECIES
(Perna viridis AND Vasticardium cf. flavum) UNDER HEAT AND HYPOXIA
STRESS under direction of NEVIATY PUTRI ZAMANI and KAREN VON JUTERZENKA
The main environmental factor that has been fully known to change globally is temperature. On the other hand, oxygen concentration under water is also changing. These changing on the form of shifting value of temperatures and oxygen concentration are led by many factors such as climate change and human activities. To sessile species Perna viridis and Vasticardium cf. flavum, these shifting on environmental factors will effect the ability to survive. Thus, how far these two species will survive, need to be assessed. This research is aimed to compare two different species from different conditions of habitat with regards to extreme thresholds of temperature and oxygen concentration. P. viridis is expected to have better stress tolerance than V. cf. flavum and that population from polluted area will also perform better than the one from benign area when being exposed to stressful conditions.
This research was conducted from May 2010 to February 2011 in Marine Habitat Laboratorium of IPB, Departemen Ilmu dan Teknik Kelautan, Fakultas Perikanan dan Kelautan. Samples were taken from Muara Angke-Jakarta Bay (P.
viridis) and Panjang Island-Banten Bay (V. cf. flavum). With regards to their
natural habitat, high temperature and hypoxia were chosen as the stressors. Levels of stressors were determined through pilot study thus DO 0,5 mg/l was used as stressor’s level in hypoxia and 34°C was for heat stress.
Sampling from the field was held with the same method for both species; they were transported in a coolbox, immersed in sea water. During the transport, every two hours, water in the coolbox was changed to keep the oxygen supply. In the lab, organisms were kept in acclimatization tanks for 10 to 14 days before the main experiment begun. The main experiment itself was conducted in 3 days for hypoxia stress and 10 days for heat stress. In both acclimatization and main experiments time, water exchange and feeding was done once in a day. Fresh sea water used to exchange the water came from 1,500 L water reservoir that was equipped by biofilter system that ran the whole day. Waste water was put back in the big water reservoir after all water exchanged. Food was given after water exchange using Coral Sand (a solution of living phytoplankton).
control group. Statistic test shows this difference significantly (p < 0,01; mediancontrol = 3; medianstressed = 2,2; n = 36). V. cf. flavum also shows a
statistically significant different response between control and stressed group (p < 0,01; mediancontrol = 2,38; medianstressed = 1,63; n = 24). Significantly different
result by statistical test implies that for P. viridis, hypoxia had been a stressor. The boxplots shows lower proportion of survivor in stressed area; this means groups that had been stressed by hypoxia condition had a lower survivorship compare to those who were in control groups. Corrected survivorship after three days of exposure to hypoxia stress was significantly different between two species (p < 0,05; medianperna = 0,74; medianvasticardium = 0,68; nperna = 18; nvasticardium = 12). The result indicates that P. viridis from Jakarta Bay has higher survivorship compare
to V. cf. flavum from Panjang Island. This implied that P. viridis is more
resistance when they are exposed to hypoxia compare to V. cf. flavum The occurrence of hypoxia events in Jakarta Bay may select those genotypes with a tolerance to hypoxia. In this experience, P. viridis might have undergone selection for tolerance to hypoxic conditions compare to V. cf. flavum In Jakarta Bay eutrophication and sedimentation levels increased over the years which has a documented impact on marine communities. Three major rivers discharge fresh water and silt into Jakarta Bay and has caused predatory gastropods and numerous mollusk species vanished from the area (Van der Meij, 2009).
The proportion of survivor in the group of stressed P. viridis was lower than in the control group. The statistical test showed this difference to be significant (p < 0,01; mediancontrol = 10,5; medianstressed = 6,25; n = 20). V. cf. flavum also
showed a statistically significant different response between control and stressed group (p < 0,05; mediancontrol = 9,25; medianstressed = 4; n = 14). The result
indicates that after being exposed to 34°C water temperature for ten days, V. cf.
flavum had reached lower survivorship compare to those that was kept under
ambient water temperature (27°C, control). The high temperature caused both species to decrease its performance up to the point of causing mortality. The survivorship is lower in the stress group and this can be considered as stress effect. Corrected survivorship comparison showed no significant difference between two species after ten days of exposure to heat stress (p > 0,05; xperna =
0,57; xvasticardium = 0,55). The result shows no significant difference, the species are
not responding differently to heat. This probably due to the fact that P. viridis from Jakarta Bay and V. cf. flavum from Banten Bay share a similar habitat background in term of water temperature. Temperature in Jakarta and Banten Bay vary between 32-33°C and 31-33°C respectively (Thoha et al., 2007; Badria, 2007).
Copyright © 2012 Bogor Agricultural University Copyright is Protected by Law
It is a prohibited to cite all or part of this thesis without referring to and mentioning the source. Citation only permitted for the sake of education, research, scientific writing, report writing, critical writing, or reviewing scientific
problem. Citation doesn’t inflict the name and honor of Bogor Agricultural University.
It is prohibited to republish and reproduce all or part of this thesis without
thewriter permission from Bogor Agricultural University.
TOLERANCE DIFFERENCE BETWEEN TWO SPECIES
(
Perna viridis
AND
Vasticardium
cf.
flavum
) UNDER HEAT AND
HYPOXIA STRESS
NURINA AYU
Thesis
as one of the requirements to achieve
Master of Science in
Marine Science Program
GRADUATE SCHOOL
BOGOR AGRICULTURE UNIVERSITY BOGOR
Evaluator in thesis defence:
Dr. Hawis Madduppa, S.Pi., M.Si.
Title : Tolerance difference between two adapted species (Perna viridis and
Vasticardium cf. flavum) under heat and hypoxia stress
Name : Rr. Nurina Ayu NIM : C551080201
Approved
Advisory Committee
Dr. Ir. Neviaty P. Zamani, M.Sc Dr. Karen von Juterzenka
(Head) (Member)
Known by
Head of Marine Science Program Dean of Graduate School
Dr. Ir. Neviaty P. Zamani, M.Sc Dr. Ir. Dahrul Syah M.Sc.Agr.. Ketua
Date of examination:
Date of graduation:
PREFACE
I praised Allah SWT whose blessing has had me finished this thesis. I would like to express my gratitude to all the parties who supports and help, so that this research and thesis can be done:
1. Advisory committee: Dr. Ir. Neviaty Putri Zamani, M.Sc. Dr. Karen Von Juterzenka for the patient, supports, advices, helps, and time during the making of this thesis until it is completed.
2. Mareike Huhn as my research counterparts, for the great ideas, lots of inputs, hard works, and most importantly: to be a very great friend.
3. Institut Pertanian Bogor from where everything was started.
4. Kiel University Foundation for financially supporting the project GAME. 5. Dr. Martin Wahl from IFM-GEOMAR in Kiel as the initiator of GAME
Project that allowed students to be involved in international network of scientists.
6. Dr. Mark Lenz for sharing excellent knowledge in the whole process of planning, conducting, analyzing and communicating science.
7. Dr. Ir. Etty Riani, M.Si. as seminar moderator and Dr. Hawis Madduppa, S.Pi., M.Si. as examiner who gave supporting and positive inputs for the better content of this thesis.
8. Team GAME VIII for every experience and motivation; and they are: Mareike Huhn, Charles Ma, Haruka Kagiwada, Florian Bechtholsheim, David J. Madariaga, Andreas Pansch, Anne Phillip, Sophia Schubert.
9. IKL and TEK 2008 friends who kept asking so I always have reasons to continue.
10. Yuliana F. Syamsuni, Dian R. Widianari, and Veronica Louhenapessy, Armin Fabritzek and Yasser Ahmed, M. Reza Cordova, Denti Risawati, Oktavika Mayasari, Syamsul Hidayat, Muhammad Taufik, Sabam P. Situmorang, Afdal Djalius, Jali, Dondy, Citra, Ketuk, Pak Danu, Bu Yanti, Kohar, Eri. loads of thanks.
Last and very special thanks to:
11. My dearest family: Amin Kuspomo, Titi H. Milihani and Nurani Luthfi for every tears and pray; support and believe; for being the wonderful parents you are. None of these will happened if there were not you.
12. My (other) parents: Abdul Manan and Rubiati, for all the courage, guidance, and acceptance. Nothing better.
13. Hesti Wahyuningsih for being in the same level of rush yet still giving so many help in which I couldn’t return (yet). Thank you very much.
Bogor, August 2012
BIOGRAPHY
Author was born in Purbalingga on May 8th 1984. She completed her elementary school in 1996, middle school in 1999 and high school in 2002. She took Water Resources Management as her major in undergraduate years in Jendral Soedirman University in Purwokerto and was graduated in the year of 2008 with thesis entitled “Effect of restricted feeding on immunity system of Red-bally Pacu
(Colossoma sp.)”. She was then registered as Master Degree Student in Bogor
Agricultural University (IPB) on August 2008. She joined an international research project GAME that was based in Kiel, Germany and issued her thesis based on the work of the project. Her thesis entitled “Tolerance difference
between two adapted species (Perna viridis and Vasticardium cf. flavum) under
heat and hypoxia stress” was her last work as one of the requirements to achieve
xxi
2.3 Environmental Factors and Stress ... 10
2.4 Effects of High Temperature and Low Dissolved Oxygen ... 11
3. MATERIALS AND METHODS ... 15
a. Hypoxia Experimental Set-up... 21
b. Heat-stress Experimental Set-up ... 23
3.3.7 Feeding ... 25
3.4 Data Analysis ... 25
4. RESULTS AND DISCUSSION ... 27
4.1 Pilot Study ... 27
4.2.1 Effects of hypoxia to Perna viridis and
Vasticardium cf. flavum ... 28
4.2.2 Tolerance differences to hypoxia between two
species ... 31
4.3 Heat Stress ... 34
4.3.1 Effects of heat stress to Perna viridis and
Vasticardium cf. flavum ... 34
4.3.2 Tolerance differences to heat stress between two
species ... 37
5. CONCLUSION AND RECOMMENDATION ... 39
5.1 Conclusion ... 39
5.2 Recommendation ... 39
REFERENCES ... 41
xxiii
LIST OF FIGURES
Page
1. Perna viridis ... 6
2. Vasticardium cf. flavum ... 8
3a. Shell appearance of Vasticardiumburchardi ... 9
3b. Gross anatomy of Vasticardiumburchardi ... 9
3c. Cross section of the Vasticardiumburchardi ... 9
4a. Sampling location, Muara Kamal-Jakarta Bay ... 15
4b. Sampling location, Panjang Island-Banten Bay ... 15
5. Map of sampling locations from where the samples were taken ... 16
6. Set-up of hypoxia experiments ... 23
7. Set-up of heat experiments ... 24
8. Example of Kaplan-Meier Survival Curve of one replicates of 10
individuals in which survival was observed every 12 hours ... 26
9. Survivals of pilot study under hypoxia stress ... 27
10. Survivals of pilot study under heat stress ... 27
11a. Proportion of survivor of Perna viridis after three days of exposure to
low oxygen concentration ... 29
11b. Proportion of survivor of Vasticardium cf. flavum after three days of
exposure to low oxygen concentration ... 29
12. Corrected survivorship of Perna viridis and Vasticardium cf. flavum
during the exposure of hypoxia stress ... 32
13a. Proportion of survivor of Perna viridis after ten days of exposure to
heat stress ... 34
13b. Proportion of survivor of Vasticardium cf. flavum after ten days of
exposure to heat stress ... 35
14. Corrected survivorship of Perna viridis and Vasticardium cf. flavum
LIST OF APPENDIX
Page
1. Pictures of some materials used during the experiment ... 51
2. Proportion of survivor data of Perna viridis under hypoxia stress ... 54
3. Proportion of survivor data of Vasticardium cf. flavum under hypoxia stress 55
4. Proportion of survivor data of Perna viridis under heat stress ... 56
5. Proportion of survivor data of Vasticardium cf. flavum under heat stress ... 57
6. Corrected survivorship data of Perna viridis under hypoxia stress
experiment ... 58
7. Corrected survivorship data of Vasticardium cf. flavum under
hypoxia stress experiment ... 59
8. Corrected survivorship data of Perna viridis under heat stress
experiment ... 60
9. Corrected survivorship data of Vasticardium cf. flavum under heat
1. INTRODUCTION
1.1 Background
Global change is occuring. It leads to ecosystem shifting, affecting lots of
species, not only terrestrial, but also marine species. In case of marine ecosystems,
the whole ecosystem will be affected, from open seas to shallow waters. This stirs
many species in critical zone of living eventually. One major aspect that is
changing is the sea water temperature. Sea water is heating and causing impacts to
marine organisms (Barnes et al., 2010; Karl and Trenberth in Lovejoy and Hannah, 2005; Tewksbury et al., 2008).
The Ocean is divided into several ecosystems, and it is in coastal
ecosystems where the most complex ocean dynamics are happening. The
biodiversity in shallow waters can be very high since they live in it were
supported by full penetration of sunlight and also nutrient inputs from lands that
leads to high productivity. On the other side, environmental factors (such as
temperature and oxygen availability) are affecting the life of many shallow water
species (Livingston, 2001; English et al., 1997; Levinton, 1982). When the temperature rises, it causes problems to marine organisms because the ability of
each marine species or organisms to resist environmental change is different and
geographically uneven (Barnes et al., 2010).
Raising temperature of sea water will cause the loss of dissolved oxygen in
the water (Neumann and Pierson, 1966). Whilst oxygen is necessary to sustain the
life of water organisms, when its supply is cut beyond to the point that sustains the
life of organisms, it will affect the ecosystem’s balance. The decline of dissolved
oxygen in the water is affected not only by temperature raising. Oxygen
consumption by organisms and nitrogen cycle also consuming dissolved oxygen
and hence declining the level of dissolved oxygen. When the level of dissolved
oxygen is so low, leading suffocation to organisms, this condition is known as
hypoxia (Diaz, 2001; Levinton, 1982). Hypoxia is today’s world-wide problem.
This problem has been accelerated with an increased input of nutrients during the
last decades. For benthic community, this can affect its structure in the bottom
Benthic and sessile organisms were the kind that will be affected the most
by environmental changes. Some mussels will absorb inputs from mainland to be
accumulated inside their tissues, like what Perna viridis did. Known as bioindicator due to its ability to survive in high-stressed environmental condition,
this species can accumulate high amount of metal in its tissues (Yap et al., 2003).
However, there are also other species that is eliminated from their original habitat
since they could not tolerate the changes; for example some species of the genus
Vasticardium was disappeared and lately distributed differently in Kepulauan
Seribu area (Van der Meij et al., 2009).
Any organism is subjected to a range of environmental variation during its
lifetime. The ability to survive environmental changes is ultimately determined by
its genome or genetic composition. When an environmental change such as
temperature rising occurs, organisms will react as showing their adaptive
response. Some environmental changes will bring the organism into a zone of
lethality. The tolerance towards environmental changes varies among species,
some species will be able rather extreme condition while the other will not be able
to survive (Levinton, 1982).
Sessile marine organisms like P. viridis and Vasticardium cf. flavum will be forced to different level of environmental parameters. Some might recover and
adapt successfully while the other will remain suffering. It is possible to find a
massive death of population in the environment that is changing because the stress
level originating in from the occurring changes might be too high so that one
specific species has no opportunity to catch up.
1.2 Problem Definition
Eventually, a shifting environment causes different effects on different
species. The main environmental factor that has been fully known to change
globally is temperature. On the other hand, oxygen concentration under water is
also changing. These changing on the form of shifting value of temperatures and
oxygen concentration are led by many factors such as climate change and human
3
environmental factors will affect the ability to survive. Thus, how far these two
species will survive, need to be assessed.
1.3 Objective
The objective of this research is to compare two different species of
bivalves from different conditions of shallow water habitat with regards to
extreme thresholds of temperature and oxygen concentration.
1.4 Hypothesis
The hypotheses for this research are:
− Hypoxia stress tolerance of Perna viridis and Vasticardium cf. flavum will be different.
5
2. LITERATURE REVIEW
2.1 Perna viridis
Perna viridis, known as Kerang Hijau in local area, or green mussel
(internationally), is one of the most popular organisms that are consumed by
human. This mussel belongs to the class of Bivalve and family of Mytilidae. They
are natural filter feeders which concentrate microorganisms present in their
surrounding waters. They live in rocky littoral and shallow sublitoral ecosystems.
They attach to hard substrate using byssus threads (Rajagopal, 2006). It is
informed that they formed extensive beds on the rocky shores which receive
effluents from fertilizer complex (Reddy and Menon, 1979).
Classification of P. viridis based on Linnaeus (1758): Kingdom : Animalia
This mussel has sagittally elongated and transversally flattened with a
blue-green coloured periostracum shell (Siddall, 1980). The mantle tissue is dark species has a long mobile foot that it uses for small-scale horizontal and vertical
movements (Seed, 1999). P. viridis filtering its food by pumping water through its siphon to extract microalgae, zooplankton, bacteria and particulate organic matter
Figure 1. Perna viridis; pictures were taken during the experiment (Huhn, 2011).
Perna viridis is known as native species in the Indo-Pacific region of Asia,
primarily distributed along the Indian and the Southeast Asian coast, then
stretches across the Persian Gulf, India, Malaysia, Indonesia, Papua New Guinea,
the South Pacific islands, and also north to Japan (Siddall, 1980; Rajagopal,
2006). They inhabit marine intertidal, subtidal, and estuarine environments with
high salinity. It is a characteristic species of mid and sublittoral faunal zones,
which often found with high density of populations. Naturally, the mussels form a
thick carpet-like growth on rocky surfaces and submerged structures (Huang et al., 1983; Rao, 1990).
In P. viridis, the sexes are separated, but the difference between males and
females are not distinguishable by external morphology (Rajagopal, 2006). The
interaction between endogenous factors and environment is believed to control the
life cycle of this mussel. The ability of this mussel to concentrate pollutants from
surrounding waters has been used by many researches to employ these mussels as
7
Monirith et al., 2003). It has a high ability to reach very high biomass levels, to withstand environmental fluctuations, to concentrate a variety of inorganic
environmental pollutants, to colonize artificial marine habitats and to invade new
geographic territories (Rajagopal et al., 2006).
Many studies reports that food availability, temperature, oxygen
concentration, salinity, pollutants, flow of water, substratum, and seasons are
factors that have influenced the life aspects of P. viridis included its reproduction, larval development and growth rate (Widdows, 1991; Seed, 1976, Rajagopal,
1991). With these many factors, they are still characterized by fast growth and
relatively high tolerance to many environmental variables. They have the ability
to survive under extreme environmental conditions such as in temperature range
of 6-37,5°C (Morton, 1987) and from saturated oxygen concentration to hypoxic
conditions (Wang et al., 2011; Huhn, 2011). For these reasons, commercial
Vasticardium cf. flavum lives from the intertidal to the subtidal in flat sandy
shore with dense seagrass beds. This species is digging vertically into the soft
substrate. It is distributed widely in Indo-Pacific waters and eastern to central part
of Indian Oceans based on the Indo-Pacific Molluscan Database it is recorded in
the GenBank under the taxname 381334. This species can grow relatively big (5 -
8 cm), thus it is also consumed by human like Perna viridis, although there are no record of this species being cultured. One study reports that Vasticardium
burchardi (Dunker, 1877) has been considered as an aquaculture candidate due to
Classification of V. cf. flavum based on Lamarck (1819) Kingdom : Animalia
Phylum : Mollusca
Class : Bivalvia
Subclass : Heterodonta
Order : Veneroida
Family : Cardiidae
Subfamily : Trachycardiinae
Genus : Vasticardium
Species : Vasticardium cf. flavum
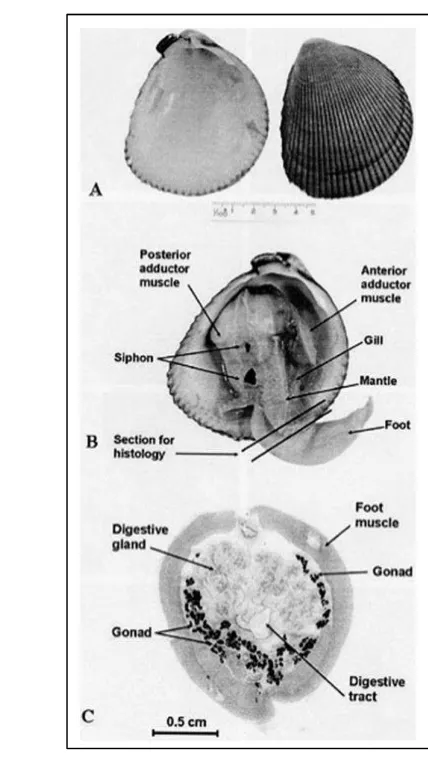
Figure 2. a) Vasticardium cf. flavum during acclimatization time; b) V. cf.
flavum in steady phase (the siphon protruded); c) V. cf. flavum under
heat (between 27-34°C) before the stressor’s level was reached, the cockle has already showed a valve opening reaction.
One research about the genus Vasticardium reported that Vasticardium
burchardi in Jeju Island is a hermaphrodite, but the reproductive stages of the
cockles were evaluated separately as oogenesis and spermatogenensis. During
summer on early August, this species in the resting stage with water temperature
ranged from 22-28°C. It was then considered mature in April, followed by May
9
Figure 3. A) shell appearance; B) gross anatomy; C) cross section of the cockle,
Vasticardiumburchardi (Limpanont et al., 2010).
Presence of this species in Indonesian waters has never been deeply
investigated. It is one of the most common cockles in almost all part of Indonesia
area, but there were no report mentioned this species specifically. It is considered
abundant since it is easy to find, yet not sufficiently taken into account as
potential research subject. Appearance of Vasticardium species in Jakarta and
Banten Bay once reported by van der Meij et al. (2009). This report informed about how anthropogenic factors can affect the presence of mollusks in
surrounding waters, and the genus Vasticardium was one of those eliminated due to high pollution. Hylleberg (2009) also mentioned the occurrence of this species
in Bali and Sunda Strait.
those who were collected around Jeju Island, South Korea, were spread at depths
between 10-30 m only (Limpanont et al., 2010; Vidal, 2007). Apparently, V. cf.
flavum was spread throughout the world, even though so far researches reporting
about this species were limited to description of the species’ taxa and reproductive
cycle only.
2.3 Environmental Factors and Stress
Marine organisms’ life will always be influenced by their environment.
There are some factors that will affect many aspects in their life, such as their
physiological aspects and ecological aspects. Environmental factors such as
temperature, salinity, oxygen, resource availability could cause effect on growth,
fecundity and even survival of the organisms when they are shifting or
fluctuating.
One factor that plays the most important role in organisms’ life is
temperature. Active animal life is limited to a narrow range of temperatures. In
global range, the latitudinal thermal gradient is accompanied by major
biogeographic changes in pelagic and bottom assemblages of organisms. At the
lower extreme the freezing of seawater results in the formation of ice crystals that
disrupt cells and terminate metabolic activity. At lethally high temperature
physiological integration is impaired and enzymes are inactivated (Levinton,
1982).
Sea surface temperature has increased worldwide with an average of
0.1-0.2°C since 1976 (Parmesan 2006). On a local scale increases in sea surface
temperature of even >1°C in the last 50 years have been reported (Sorte et al. 2010). In Jakarta Bay itself, rising temperature of seawater had degraded marine
biodiversity; such as in Pari Island where 50-60 % coral reefs were found bleach
(DFID & World Bank, 2007).
High temperatures makes oxygen is less soluble. At the same time,
dissolved oxygen-dependent respiration rates of most microbes, macrofauna and
algae increase markedly. Increasing temperatures exacerbate low oxygen stress
and promote the expansion of anoxic dead zones, especially in marine areas with
11
and agricultural waste releases, and in coastal waters and deep basins with poor
circulation. Diaz (2001) mentioned that the condition where oxygen concentration
decline beyond the point that sustains most animal life is called hypoxia. Hypoxia
has occurred throughout the world, especially to areas without intensive
regulation of nutrient inputs. Once it occurred, hypoxia quickly became an annual
event.
changes are happened all of sudden, organisms’ performance will be disturbed at
at some lethal point, they won’t be able to survive (Riani, 2012).
Fluctuations in environmental factors that reducing the performance of
organisms is defined by stress (after GAME, 2011). At certain range of point, the
fluctuations in environmental factors can be a positive trigger for organisms. It
can prepare the organisms against the same kind of disturbance that might occur
in the future. When this is happened, then the fluctuations are considered still in
the range of organisms’ tolerance range. After recovery, organisms can be more
resistance under unlikely conditions than before. If the fluctuations keep going up
to the point of no chance of recovery, their physiological process can decrease and
if it is continued mortality can happen (Riani, 2012).
2.4 Effects of High Temperature and Low Dissolved Oxygen
Sea temperature affected the life of organisms inside, especially the sessile
bivalves. In tropical region, annual sea temperature fluctuated narrowly compare
to those in subtropic and high latitude region. The reason to this is mainly because
the equal quantity of sunlight penetration during the whole year. Sea surface
temperature in Indonesia is ranged from 27 to 30°C throughout the year (Qu et al., 2005). In shallow waters, this temperature could be higher since the sunlight may
Most studies have investigated the environmental parameters that stand
behind the successful stories of Perna viridis invasion (Cheung, 1993; Wong, 1999; Yap et al., 2002; Wong and Cheung, 2003; Gao, 2008). The ability of organisms, including Perna viridis, to survive during the temperature and oxygen change plays an important role in the species dispersal. High stress tolerance is
known as one of the key characteristic of marine invasive species. In addition to
fast growth and reproductive rate and high plasticity on the living requirement, it
is thought to be one of the traits that make the invasion so successful (Stachowicz
and Byrnes, 2006).
Temperature is one role factor of mollusks life. Rajagopal (2006) stated that
temperature has a great effect on larval stage of mollusks and determine their
growth. On average, 20-25°C is a point when mortality of mollusks will occur.
Meanwhile, Adnan (2009) mentioned that temperature also affecting the
concentration of dissolved oxygen in water. This is happened indirectly; the
higher the water temperature, the less water will hold the oxygen in solution. The
concentration of dissolved oxygen is high when the water quality is good (Adnan,
2009).
Dissolved oxygen is one main chemical factor that is inevitably needed for
respirations by most of water organisms. Dissolved oxygen came from diffusion
and also appeared as a result of photosynthesis by chlorophyll-ed organisms. The
interaction between dissolved oxygen and water temperature as mentioned before
is holding an important role for water organisms. The lack of dissolved oxygen
will suffers lots of them, beside the fact that it’s also disturbing the environmental
balance; but apparently, mussels have special ability to survive the unlikely
environment. The decrease of dissolved oxygen for several days, supposed to be
not very significant for mussels, because they can close their shells and stay
relatively still to reduce the reduce oxygen consumption for respiration (Quayle in
Setiobudiandi, 2000).
As popular cultured species in Indonesia, P. viridis that is cultured can reach its maximum growth between the range of temperature from 15 to 32°C (Vitner,
2001). In South Korea, one species of the family Cardiidae, Vasticardium
13
temperature during the winter is 14°C, then developed its reproduction function in
around 22 – 28°C (Limpanont et al., 2010). In fact, the ability of a species to grow and reproduce is a trait of suitable habitat condition for it.
Cultured P. viridis reach its optimum growth under the range of dissolved oxygen from 3 to 8 mg/L (Kantor Mentri KLH & LON LIPI in Porsepwandi,
1998). For V.buchardi (Dunker, 1877), which survived particularly on Jeju Island of South Korea and was able to preserve its population in the location (33°10’ –
33°40’N, 126°60’E) (Limpanont et al., 2010), the concentration of dissolved oxygen in the surrounding water was reported to be in range from 3,30 to 8,43
15
3. MATERIALS AND METHODS
3. 1. Time and Location
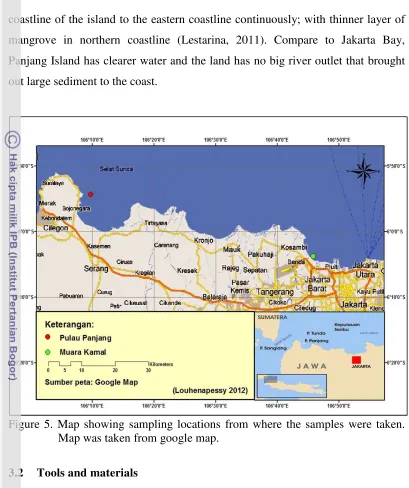
This research had been held from May 2010 to Februari 2011. This research
was held in di Habitat Laboratory of IPB, Fakultas Ilmu dan Teknik Kelautan.
Samples were taken from Muara Kamal-Jakarta Bay and Panjang Island-Banten
Bay. Muara Kamal is a fishing harbor with a lot of green mussel farms spread not
far from the harbor; while Panjang Island is located in northern part of Banten
Bay (Figure 3-1). In the lab, all treatments were conducted in the laboratorium to
control the consistency of the experiments.
Figure 4. a) Muara Kamal-Jakarta Bay; b) Panjang Island-Banten Bay. Pictures were taken during sampling.
Jakarta Bay is a popular area for green mussel cultures; mussels that
cultured there are known to have thick meat and were preferred for human
consumption. However, apart from its suitable environmental conditions for
mussels, Jakarta Bay itself is suffered by large input from the mainland which
houses over 12 million inhabitants in the area. As an impact, from 1937 to 2005,
predatory gastropods and numerous mollusk species have vanished. Vasticardium
cf. flavum is mentioned as one of them (van der Meij, 2009).
Panjang Island, on the other side, is an island in Banten Bay with area ±820
Ha inhabited by 2.699 inhabitants (in the year 2000). This island is surrounded by
smaller islands that are uninhabited. Mangrove ecosystem in this island can be
coastline of the island to the eastern coastline continuously; with thinner layer of
mangrove in northern coastline (Lestarina, 2011). Compare to Jakarta Bay,
Panjang Island has clearer water and the land has no big river outlet that brought
out large sediment to the coast.
Figure 5. Map showing sampling locations from where the samples were taken. Map was taken from google map.
3.2 Tools and materials
The tools used in these experiments were cool-boxes, basic diving
equipments, latex gloves and aquarium filter, thermometer, water canister, plastic
containers of 4 and 7 l, plastic tanks for acclimatization, air stones, water heaters,
fiber mesocosms, water pump, oxymeter, biofilter, and water reservoir of 1.000 l.
The main material used in this research is sea water which was provided in
17
3.3 Research Methods 3.3.1 Species
Associated with the term environmental shifting, sea organisms most likely
will be affected. When environment is shifting, organisms will also change its
behavior in attempt to fit to the new condition. At some point, when the change is
unlikely, this will lead to stress. This effect could cause a bigger impact to sessile
organisms instead of mobile ones. Sessile organisms such as bivalves will not be
able to move to avoid unlikely condition of environmental and thus will suffer
from stress.
This experiment was using P. viridis and Vasticardium cf. flavum as studied organisms. They were from the same class, Bivalvia that is recorded spread
throughout the world. P. viridis was chosen because it was abundant in Jakarta Bay and easy to find; while V. cf. flavum was taken because it was relatively easy to find in Panjang Island water. Even though these species have different niches,
they share the same lifestyle, as filter feeder in the ecosystem. It will be
interesting to see the response of these species under the given stressors.
3.3.2 Sampling
Sampling was conducted in several stages. The first stage was sampling for
pilot study to determine the stressor levels, and the second stage was sampling for
the main experiments. Sampling for both stages were taken from Muara Kamal,
Jakarta Bay and Panjang Island, Banten Bay. The sample collection methods were
the same for pilot and main experiment.
P. viridis samples from Jakarta Bay were cultured and harvested off ropes.
Mussels that were chosen were those around 4 – 6 cm in length. They were placed
directly inside a coolbox, still attached to their ropes, submerged under fairly
amount of water. The water that was used was the water from the sampling spot,
which has been filtered before being poured into the coolbox.
Vasticardium cf. flavum samples were collected manually by snorkeling and
were picked up from the sand directly by hand in the eastern side of Panjang
Island. Coolboxes that were half-filled by sea water were placed on small boats
substrate, around seagrasses. Since they were rather hidden by the sand, every
cockle found was taken as sample without considering their size. They were
placed in the coolboxes without sand, and compare to P. viridis, the amount of the found V. cf. flavum, was much less. Only after they were kept in the lab, the size of these cockles can be measured. They were ranging from 3 – 6 cm in length.
Some very young (much smaller in size) cockles then were kept separately and
not used during the experiments.
Both species were transported inside the cool-boxes and brought to the lab
using car. At every time of carriage, the water temperature inside the cool-boxes
was reduced by adding covered ice blocks to lower the organisms stress. Every
two of eight hours transport, the water was replaced with fresh sea water patched
inside 20L volume of water jar that was directly collected from the sampling site,
due to increase the oxygen supply to avoid high number of mortality of the
mussels.
During transport, P. viridis was not revoked from the fouling structures and during acclimatization period in the lab as well. Each individual was revoked after
they were about to be put inside the small plastic containers as the treatment
begun. Their lengths were measured right before they were included into the
containers.
During acclimatization period, V. cf. flavum was first kept in a glass tank filled with sand. After a while they were moved to the same plastic tank as Perna
viridis. This was purposely done to reduce the shock of different environment
condition between their natural habitat and the artificial habitat in the lab. The
same procedure of length measurement as in case of P. viridis was conducted to
V. cf. flavum before their treatments begun.
3.3.3 Acclimatization
Acclimatization time for each species from each sampling site was around
10 days to 2 week period. This period was determined by considering some
factors like the health condition of the organisms during transport, the recovery
ability of the organisms after transport, and the adaptation ability of the organisms
19
stressful condition due to the bumping and shaking, after that they need to
recovered and get used to the lab condition before the experiment started; for this
reason each species from each sampling site was acclimatized in different periods.
Vasticardium cf. flavum needed longest period of acclimatization since they were
transported without any other media but water that caused biggest bumping effect
during transport, moreover they needed a longer recovery time and also longer
adaptation phase due to completely different kind of habitat they got after they
were put in the lab where there were no natural sandy substrate and no seagrasses.
Different story happened to P. viridis which transported still with their attaching media, they were transported attached to the rope from Jakarta Bay and in those,
and they were put in the cool-boxes.
During acclimatization phase, both P. viridis and V. cf. flavum were put in acclimatization tanks which included 3 big plastic tanks, and 4 glass tanks, only 2
of the glass tanks were filled by sand, while the rest were left without substrate. In
every plastic tank there was one filtering pump and an airstone that bubbling
oxygen into the water, the glass tank was not equipped with filtering pump but
there was an airstone.
Water exchange in acclimatization phase was done every day. The fresh
seawater came from a big water reservoir attached to the biofiltering system that
ran the whole day. Waste water was put back in the big water reservoir after all
acclimatization tanks refilled. After water exchange, mussels were fed with 1
pipette of Coral Sand per one tank. Coral Sand is a solution of living
phytoplankton. This food was also given to the mussels during the experiment.
3.3.4 Stressors
Stressors chosen were heat stress and hypoxia stress. The event of rising
temperature of seawater brought by climate change would cause stress to
organisms in their natural habitats. Furthermore, increasing temperatures
exacerbate low oxygen stress and promote the expansion of anoxic dead zones.
This is a scenario that happened in natural habitat and causing stress to bivalves.
the point of upper limit compare to non-tropical species; therefore it’s not possible
they will experienced the mentioned scenario in their natural habitat.
Taking this as consideration, heat and hypoxia were chosen as the stressors
of the experiment. These two stressors were also feasible to mimic in laboratory
experiments. The levels of the stressor were then decided through pilot studies.
3.3.5 Pilot Studies
Pilot studies were conducted to determine the stress level used in the main
experiments. Stress is described as fluctuations in environmental parameters
reducing the performance of an organism (Lenz et al. in prep.). Oxygen depletion and high temperature are two abiotic factors that were selected as stressors in this
experiment. The stress levels where minimum oxygen and maximum temperature
levels were determined each in a two-week experiment.
During this pilot study, three levels of temperatures were divided to
determine the main experiment’s temperature level; they were 32, 34, and 36°C.
While 32°C is known as the upper thermal limit that support optimum growth to
P. viridis, 34 and 36°C were chosen to mimic worst scenario that could occur
naturally in the mussels’ habitats. A control group was also provided in the pilot
study with ventilated water condition in 27°C water temperature, known as the
average temperature of the ecosystem. Meanwhile, oxygen level at 1,5 and 0,5
mg/l were used, the last level is known as the critical level of oxygen depletion
towards hypoxia in ecosystems (Diaz and Rosenberg, 2008).
Each treatment had five replicates; one replicate was one 7 l plastic
container filled with 4 l of seawater and 10 individuals. Consequently, both
species were exposed to >5 mg/l DO (ambient oxygen), 1,5 mg/l DO (low
oxygen) and 0,5 mg/l (hypoxia). Temperature levels were 27°C (ambient
temperature), 33°C (temperature in habitat + 4°C), 34°C (temperature in habitat +
5°C) and 35°C (temperature in habitat + 6°C). The respective other abiotic factor
was kept constant.
During the pilot study, data was collected every 12 hours. This was decided
to avoid the decreasing of water quality after one organism is dead. When one of
21
problem of water quality in the treatment container, that is why 12-hours checking
will prevent the problem because then the dead individual can be taken and/or
period of treatment. Proportion of survivors was noted as the response, and then
the result was used in between-population comparisons. Between-population
comparisons meant to compare the stress tolerance between population P. viridis
and Vasticardium cf. flavum from Panjang Island waters. Based on the results on
pilot study, the stress level used for hypoxia treatment was 0,5 mg/l while for heat
stress experiment the stress level was 34°C.
a. Hypoxia Experimental Set-up
In this experiment, there were treatment and control groups each with
10 replicates. Both groups were put randomly in one big bench so that every
container will receive even condition. Control containers were 7 l volume
plastic container filled with only 4 l of seawater equipped with airstones,
while treatment containers were 4 l volume plastic container full filled with
water and then closed tightly in purpose to avoid oxygen go in or out of the
water. Low oxygen water that filled the treatment containers came from one
canister of water supply where seawater was flushed by nitrogen bubbles. The
nitrogen bubbling process took some time until the oxygen level in the water
reach 0,5 mg/l. After that the water was discharged into plastic container
through water outlet in the bottom of the canister. Using hose as help, water
was drained smoothly into the plastic containers then the lid was covered
direct yet tightly.
The measurement of oxygen was conducted by inserting oxymeter into
small hole on the lid of the canister. Oxygen concentration was checked
occasionally and once the required level was reached, I stopped the bubbling
back inside. After the plastic containers were finally filled with low-oxygen
water, I measured the oxygen concentration inside to make sure the level of
oxygen was fit.
Individuals were inserted into containers by opening the lid slightly
after they were put on the bench. The number of individual that was put
inside the plastic containers was different for P. viridis and V. cf. flavum There were 10 individuals of P. viridis per replicate while there were only 4 individuals of V. cf. flavum This difference was done because of the different biomass of two species.
During the experiment, water exchange was done after mortality
checking. Dead individual was taken out and being replaced by a new one
that has been marked. Replacement individuals won’t be included into the
count of mortality if they were dead, because they exposed to different time
of stress. They were only being put there to keep the respiration ratio inside
the plastic containers.
Daily water exchange was done by the same procedure as the way it
was filled in the first time. The different was it only half water removed from
the plastic container was refilled by the low-oxygen water from the canister.
Removing the waste water was conducted using hand pump. However if the
water quality looked really bad due to contamination of dead individual, the
23
Figure 6. Set-up of hypoxia experiments. Upper left: 60 l- barrel with deoxygenized water (white barrel), tank of filtered water (orange) and nitrogen tank with PVC-hose; upper right: nitrogen inlet and sealed cover of 60 l- barrel; lower left: replicate with 10 mussels and hypoxic water; lower right: replicates of ambient oxygen and hypoxia stress treatments (Huhn, 2011)
b. Heat Stress Experimental Set-up
Heat stress experimental set-up required one long bench to put plastic
containers as control, and two fiber mesocosms to put plastic containers as
treatment. The mesocosms were filled with heated fresh water surrounding
the containers that were filled with the already heated seawater. However, the
heated water was not reached in one day, but it was adjusted by increasing the
temperature 2 degrees per day before finally reached 34°C; the heating started
from ambient temperature (27°C). Data was also collected in 12-hours period,
started after 34°C water temperatures was reached. In this set-up, all
containers were equipped by airstones and the containers used were only 7 l
volume fiber containers filled with 4 l seawater. Containers inside fiber
mesocosms were supported by bricks to keep it stable and in the meantime
also covered by glass covers to made it heavier so it did not flowing. In
addition, to keep the temperature level stable, pieces of styrofoam framed and
There were 5 individuals of P. viridis and 3 individuals of V. cf. flavum in each of replicate. The number of individuals was different from oxygen
depletion experiment due to different stock of organisms left. The heat stress
was held after the oxygen depletion, and to avoid different condition of
organisms, the same pool of sample units that was left after oxygen depletion
experiment were used. Due to the same reason, replacement dead mussel by
new ones also was not done.
Daily water exchange were conducted the same as in hypoxia treatment
by replacing half water in the containers with the fresh one. The exchange
water was prepared before in a bucket with particular amount of fresh-filtered
sea water from water reservoir of 1.000 l. The bucket was submersed by fresh
water that was heated using immersion heater.
25
3.3.7 Feeding
During the time mussels and cockles were kept in the lab, both species were
fed using Coralsands (DT’s Premium Blend Live Marine Phytoplankton), a liquid
phytoplankton food that had a concentration of 2,5 x 106 cells/ml. In acclimatization phase, Coralsands was given daily by the ammount of1 ml food
solution per 80 l water. In the pilot study and the main experiments, mussels were
fed with 0,1 ml food solution per containers once every two days. This amount of
food was chosen based on recommendations by Coralsands.
3.4 Data Analysis
The data extracted from the experiment for analysis were proportion of
survivorship and corrected value of survivorship. Proportion of survivorship was
calculated as the sum of proportional survival of each observation point. As the
observation point was conducted every twelve hours, in term of time unit as day,
survival rates were multiplied by 0,5. This proportion was used to detect the effect
of the stressors applied to P. viridis and Vasticardium cf. flavum The example of how this proportion was obtained is illustrated by Figure 3.4. Corrected values of
survivorship were counted to get the values that can be compared to see tolerance
difference between two species. These values were obtained by calculating the
mean value for controls and divide the value of each stressed replicate by the
Figure 8. Example of d every 12 hours. The curve is known as Kaplan
analyzed using R 2.10.1 for Windows (R Deve
tect whether the stressors applied had an effec
st was used; in other case, when the data were n
nsformable to normality, Mann-Whitney U-Tes
hod was used to see tolerance difference betwe
normality was Shapiro-test while to test the hom
test was conducted. Boxplot was produced to il
27
IV. RESULTS AND DISCUSSION
4.1 Pilot Study
The experiments were conducted after determining stress level through a
pilot study. The Pilot study was conducted using Perna viridis from Jakarta Bay.
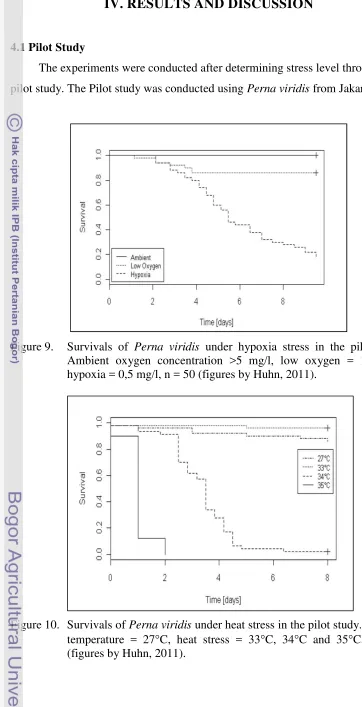
Figure 9. Survivals of Perna viridis under hypoxia stress in the pilot study. Ambient oxygen concentration >5 mg/l, low oxygen = 1,5 mg/l, hypoxia = 0,5 mg/l, n = 50 (figures by Huhn, 2011).
The pilot study for hypoxia showed that after 10 days of observation,
proportion of survivor was 1,0 in ambient oxygen, 0,86 in low oxygen, and 0,18
in hypoxia condition. Meanwhile in heat stress treatment, proportion of survivor
of 33°C of water was the highest to the point of 0,96, it was 0,86 in 27°C, and
0,02 in 34°C; as for the 35°C water temperature, proportion of survivor has
reached zero point in two days.
Pilot study was conducted to determine the level of stress that would lead to
a high number of mortality but still in the range of organisms’ tolerance. As P.
viridis is known to have relatively wide range of tolerance to environment
variability, the result of the pilot study implied that the mussel suffered in the
level DO of 0,5 mg/l and 34°C. However, those levels didn’t give an impact such
as sudden mass mortality so it was not lethal to the entire population. Those levels
were considered as environmental stress (Levinton, 1982) to the mussels and
therefore were being used as stressors’ levels in the main experiments.
4.2 Hypoxia Stress
4.2.1 Effects of hypoxia to Perna viridis and Vasticardium cf. flavum
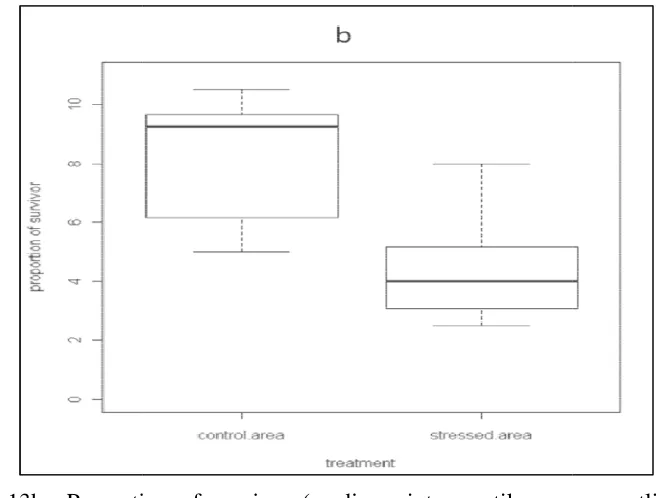
The Proportion of survivors in the group of stressed Perna viridis is lower than the control group. Statistic test shows this difference significantly (p < 0,01;
mediancontrol = 3; medianstressed = 2,2; n = 36) (Figure 11a).
Significantly different result by statistical test implies that for P. viridis, hypoxia had been a stressor. The boxplots shows lower proportion of survivor in
stressed area; this means groups that had been stressed by hypoxia condition had a
lower survivorship compare to those who were in control groups. In other words,
hypoxia in DO amount of 0,5 mg/l is decreasing the performance of P. viridis, and it lead to mortality.
Meanwhile, Vasticardium cf. flavum also shows a statistically significant different response between control and stressed group (p < 0,01; mediancontrol =
2,38; medianstressed = 1,63; n = 24) (Figure 11b), which means that hypoxia also a
stress to the species during the experiment. The lower survivorship of V. cf.
flavum after being exposed to hypoxia condition indicates the decreasing of
several days, the cont
shows better performa
Figure 11a. Proportion of non-outl >5 mg/l (
Figure 11b. Proport outlier (control
ontrol group that had been treated by more prefe
mance than those that were exposed to hypoxia
oportion of survivors (medians, interquartile ranges outlier ranges) of P. viridis after three days of ex
/l (control) and 0,5 mg/l (stressed); n = 36; p < 0,01.
oportion of survivors (medians, interquartile ra er ranges) of after three days of exposure t ontrol) and 0,5 mg/l (stressed); n = 24; p < 0,01.
29
referable condition
ia stress.
nges, outliers and
exposure to DO p < 0,01.
One factor that may decrease the performance of mussels is environmental
stress. Many researches had observed the impact of hypoxia on organisms
whereas the thresholds of hypoxia proposed in the literature mostly refer to a
value of 2 mg/l. This threshold refers to the oxygen level for fisheries collapse,
but the diversity of behavioral and physiologic adaptations to hypoxia suggests
that different taxa are likely to exhibit different vulnerability to hypoxia and
therefore may have different oxygen thresholds (Vaquer-Sunyer and Duarte,
2008).
Bivalves are the taxa that considered as the most tolerant to hypoxia stress
Vaquer-Sunyer and Duarte, 2008; Stickle et al., 1989). They have the ability to close their valves and slowing down their heart rates when their circumstances are
unlikely. This behavior was seen in both Mytilus edulis and Perna viridis (Theede
et al., 1969; Huhn, 2011) as also happened during the experiment. P. viridis had
closed its shell during three days exposure of hypoxia.
Meanwhile, Vasticardium cf. flavum that was not provided with substrate to burrow during the experiment, showed the same idleness as P. viridis. Despite its siphon that slightly protruded, Vasticardium cf. flavum remained inactive. Even so, after several hours of exposure to hypoxia, they closed down their shells
totally and only opened when they were dead and no longer had the strength to
keep the shells closed. This was a confirmation of what was stated by Shick et al. (1986) that sessile and infaunal bivalves generally show a strong resistance to low
oxygen condition due to a reduction in activity and hence energy used. Compare
to crustaceans, which tolerance to hypoxia were correlated to activity level and
metabolic rate, bivalves such as Crassostrea virginica died because they remained closed and could not feed and maintain an aerobic metabolic rate; presumably
because of anoxic conditions produced by dredging which resulted in an oxygen
demand of spoil bank sediments and modification of the local hydrographic
regime (Andrews, 1982; Hoese and Ancelet, 1987; Stickle et al., 1989).
Many studies reported that bivalves, in fact, have the ability to survive in
extreme hypoxia condition. The reason is because in many instances a part or all
of the blood supply from the general surface returns to the auricle without passing
31
chitons goes direct to the auricle by passing the gills. This mechanism is most
conspicuously on bivalves, even though it also occurs in some pulmonates and
prosobranch. There is also some evidence that to a varying degree bivalves can
respire anaerobically such as in case of Mya arenaria that can survive without oxygen for 8 days during which period the glygocen of its tissue decreased; but of
course this level of ability is again, species-specific (Newell in Wilbur and Yonge,
1964). This might come in term with the distribution of molluscs on the shore, the
latitude difference, and also the variation of lifestyles. Oysters and mussels
usually simply close down completely under condition of low oxygen. During low
tide, mussel Mya (genus) builds up an oxygen debt to be repaid by an increased rate of pumping when the tide returns. Meanwhile, many other species are not
closing their shell valves completely so the free edges of the mantle can protrude
through the shell gape.
This experiment also shows that under certain circumstances where both
species had experienced several stages of unfavorable conditions (transport,
acclimatization phase, and handling during the preparation of the experiments),
they might decrease their ability to recover. However, the lab condition was not
their natural condition, this may put more stress to their conditions and decrease
their performance faster compared to when hypoxia event occurred in their natural
habitat.
4.2.2 Tolerance differences to hypoxia between two species
Corrected survivorship after three days of exposure to hypoxia stress was
significantly different between two species (p < 0,05; medianperna = 0,74;
medianvasticardium = 0,68; nperna = 18; nvasticardium = 12) (Figure 12). The result
indicates that P. viridis from Jakarta Bay has higher survivorship compare to
Vasticardium cf. flavum from Panjang Island. This implied that P. viridis is more
Figure 12. Corrected ure of hypoxia stress. P < 0,05.
a species to stress is determined by abiotic and
ection through stressful conditions (Huhn, 2011)
luding predators, pathogens, and competitors; w
onmental parameters including temperature, oxyge
the water (Catford et al., 2008). Considering they were taken from, the difference in resul
avum when they were exposed to hypoxia stress
idis might be already adapted to the unfavorabl
flavum that lives in a less polluted area. As re
hose genotypes with a tolerance to hypoxia. In