PERKEMBANGAN DAN MORFOLOGI GAMETOFIT
TUMBUHAN PAKU MARGA
PTERIS
DI PULAU JAWA
DWI SUNARTI PUSPITASARI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Perkembangan dan Morfologi Gametofit Tumbuhan Paku Marga Pteris di Pulau Jawa adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
DWI SUNARTI PUSPITASARI. Perkembangan dan Morfologi Gametofit Tumbuhan Paku Marga Pteris di Pulau Jawa. Dibimbing oleh TATIK CHIKMAWATI dan TITIEN NGATINEM PRAPTOSUWIRYO.
Pteris L. merupakan salah satu marga besar dari suku Pteridaceae yang tersebar di daerah tropis maupun subtropik. Penelitian beberapa jenis Pteris di Indonesia masih sebatas kajian morfologi sporofit, tipe reproduksi, dan sitologi. Penelitian mengenai perkembangan dan morfologi gametofit pada marga ini belum pernah dilakukan sebelumnya. Penelitian ini dilakukan untuk mempelajari perkembangan dan morfologi gametofit tumbuhan paku marga Pteris di pulau Jawa.
Penelitian telah dilaksanakan pada bulan Juli 2014 hingga April 2015. Pengambilan sampel dilakukan di beberapa lokasi di Pulau Jawa, yaitu: Jawa Barat (Kampus IPB, Situgedhe, Ciomas, dan Cianjur), Jakarta (Mampang Prapatan), dan Jawa Tengah (Sragen dan Banyumas). Delapan jenis Pteris (P. biaurita, P. ensiformis, P. exelsa, P. longipinnula, P. mertensioides, P. tripartita, P. venulosa, dan P. vittata) telah diamati tipe reproduksinya dengan menghitung jumlah spora pada setiap sporangium. Spora seberat 0.002 gram disemai secara merata ke dalam kotak plastik yang berisi media campuran vermiculite, spagnum moss, dan perlite. Perkembangan gametofit diamati setiap tujuh hari sekali sampai gametofit membentuk organ seksual.
Delapan jenis Pteris Jawa yang telah diamati memperlihatkan variasi tipe reproduksi. Empat individu bertipe reproduksi seksual (P. ensiformis, P. mertensioides, P. tripartita, dan P. venulosa), tiap sporangiumnya mengandung 64 spora normal. Tiga jenis lainnya memiliki tipe reproduksi apogami dengan 32 spora normal per sporangium (P. biaurita, P. longipinnula, dan P. vittata). Pteris exelsa bertipe reproduksi apogami abnormal, sporangiumnya mengandung 64 spora tidak normal (bentuk dan ukuran berbeda).
Perkembangan gametofit secara umum memiliki enam fase penting yaitu fase rhizoid, fase rhizoid-protokorm, fase filamen, fase spatula, fase hati muda, dan fase hati dewasa (fase laminar yang telah menghasilkan organ reproduksi). Perkecambahan spora jenis Pteris mengikuti tipe Vittaria. Jumlah sel dalam filamen bervariasi antara 2-32 sel. Perkembangan gametofitnya ada tiga tipe, yaitu: tipe Ceratopteris (P. biaurita, P. exelsa, P. longipinnula, P. mertensioides, dan P. vittata), tipe Adiantum (P. tripartita), dan modifikasi antar kedua tipe tersebut (P. ensiformis dan P. venulosa). Empat jenis Pteris yaitu P. biaurita, P. exelsa, P. longipinnula, dan P. vittata menghasilkan 100% gametofit jantan. Pteris ensiformis menghasilkan sekitar 83 % gametofit jantan dan 17 % hermaprodit, sedangkan pada P. tripartita menghasilkan 73% gametofit jantan dan 27% hermaprodit. Perkembangan gametofit antara jenis Pteris berbeda dalam enam ciri yaitu jumlah sel dalam filamen, lama perkecambahan spora, waktu pembentukan notch, waktu pembentukan organ seksual, bentuk gametofit dewasa, dan tipe perkembangan gametofit.
SUMMARY
DWI SUNARTI PUSPITASARI. Gametophyte Morphology and Development of Pteris from Java Island. Supervised by TATIK CHIKMAWATI and TITIEN NGATINEM PRAPTOSUWIRYO.
Pteris L. is a large fern genus belongs to the Pteridaceae family, which is distributed in both tropical and subtropical regions. The morphology of sporophyte, the type of reproduction, and the cytology of Pteris had been reported. The gametophyte morphology of Pteris in Java has not been studied yet. The objective of this study was to describe the gametophyte morphology and development of Pteris in Java Island.
This study was conducted from July 2014 until April 2015. The samples were collected from several localitions in Java, i.e.: West Java (IPB Campus, Situgedhe, Ciomas, and Cianjur), East Jakarta (Mampang Prapatan), and Central Java (Sragen and Banyumas). Eight Pteris species, P. biaurita, P. ensiformis, P. exelsa, P. longipinnula, P. mertensioides, P. tripartita, P. venulosa, and P. vittata had been observed for its reproduction types by counting the number of spores in each sporangium. As much as 0.002 g spores was sown in transparent box contained of sterile medium containing of vermiculite, spaghnum moss, and perlite with the ratio of 2:2:1. The gametophyte development of each species was observed under a microscope every 7 days until it formed sexual organ.
Eight species of Pteris observed show variations of reproduction type. Four species (P. ensiformis, P. mertensioides, P. tripartita, and P. venulosa) have a sexual reproduction type, with 64 normal spores per sporangium. Three species were apogamous with 32 normal spores per sporangium (P. biaurita, P. longipinnula, and P. vittata). Pteris exelsa was apogamous with 64 irregular spores per sporangium (various in shape and size).
Generally, the development of gametophyte consists of six important phases, namely: rhizoid phase, rhizoid-protocorm phase, filamentous phase, spatulate phase, young heart phase, and mature heart phase (laminar phase bearing reproduction organ). Spore germination of Pteris is the Vittaria type. The number of filament cells is varied 2-32 cell. There are three types of gametophyte development, namely: Ceratopteris-type (P. biaurita, P. exelsa, P. longipinnula, P. mertensioides, and P. vittata), Adiantum-type (P. tripartita), and the modifications of the two types (P. ensiformis and P. venulosa). Four species of Pteris (P. exelsa, P. longipinnula, P. vittata, and P biaurita) produce 100 % monosexual/male gametophyte. Pteris ensiformis produces 17% hermaphrodite gametophyte and 83% male gametophyte, while P. tripartita produce 27% hermaphrodite and 73% male gametophyte. There are six gametophyte differences among species, which are, the number of filament cell, germinated time, notch formation time, shape of gametophyte, gametangium formation time, and the type of gametophyte development.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biologi Tumbuhan
PERKEMBANGAN DAN MORFOLOGI GAMETOFIT
TUMBUHAN PAKU MARGA
PTERIS
DI PULAU JAWA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Judul yang dipilih dalam penelitian ini yaitu “Perkembangan dan Morfologi Gametofit Tumbuhan Paku Marga Pteris di Pulau Jawa”. Penelitian yang berlangsung dari bulan Juli 2014 hingga Agustus 2015 ini dilakukan di Laboratorium Taksonomi Tumbuhan, Departemen Biologi FMIPA IPB dan Laboratorium TREUB, Pusat Konservasi Tumbuhan-Kebun Raya Bogor, LIPI. Sebagian dari tesis ini telah dipublikasi dengan judul “Gametophyte Morphology and Development of Six Species of Pteris (Pteridaceae) from Java Island Indonesia” pada jurnal berskala internasional, The Journal of Tropical Life Science (JTROLIS) 5 (2): 108-114.
Terima kasih penulis ucapkan kepada Dr Ir Tatik Chikmawati, MSi dan Dr Titien Ngatinem Praptosuwiryo, SSi MSi selaku dosen pembimbing yang telah banyak memberikan nasihat, motivasi, saran serta bimbingan. Penulis mengucapkan terima kasih kepada bapak dan ibu dosen Program Studi Biologi Tumbuhan (BOT) atas semua ilmu, pengalaman, bimbingan, dan nasihat yang diberikan. Ungkapan terima kasih juga penulis sampaikan kepada Dirjen Pendidikan Tinggi (DIKTI) atas pemberian Beasiswa Program Pascasarjana Dalam Negeri (BPPDN). Penulis juga mengucapkan terima kasih kepada institusi-institusi dimana penulis dapat menggunakan fasilitasnya (Laboratorium Taksonomi Tumbuhan Departemen Biologi FMIPA IPB dan Laboratorium TREUB, Pusat Konservasi Tumbuhan-Kebun Raya Bogor, LIPI) dan mendapat ijin pengambilan sampel (Kebun Raya Cibodas dan Kebun Raya Baturraden). Ungkapan terima kasih penulis sampaikan kepada orang tua, seluruh keluarga dan sahabat atas segala doa, dukungan, serta kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN ix
1 PENDAHULUAN 1
Tujuan Penelitian 2
2 TINJAUAN PUSTAKA 3
Pteris L. 3
Tipe Reproduksi Tumbuhan Paku 4
Perkembangan Gametofit Marga Pteris 5
3 METODE 10
Waktu dan Tempat Penelitian 10
Bahan Tumbuhan 11
Teknik Pengambilan Bahan Tumbuhan 11
Koleksi Spora 11
Pengamatan Tipe Reproduksi 11
Pengamatan Gametofit 11
Analisis Data 12
4 HASIL DAN PEMBAHASAN 13
Tipe Reproduksi dan Morfologi Spora 13
Perkembangan Gametofit Pteris spp. 15
Deskripsi Morfologi Gametofit Pteris spp. 22
5 SIMPULAN DAN SARAN 33
Simpulan 33
Saran 33
DAFTAR PUSTAKA 34
LAMPIRAN 38
DAFTAR TABEL
1 Urutan fase perkembangan gametofit tumbuhan paku 8
2 Lokasi pengambilan sampel tumbuhan paku Pteris 10
3 Ukuran spora dan tipe reproduksi Pteris spp. 13 4 Perbandingan perkembangan gametofit antara hasil pengamatan 17
dengan hasil Zhang et al. (2008)
5 Inisiasi pembelahan sel awal pada filamen 17 6 Rata-rata jumlah sel pada delapan jenis Pteris spp. yang diamati selama 10
minggu 19
DAFTAR GAMBAR
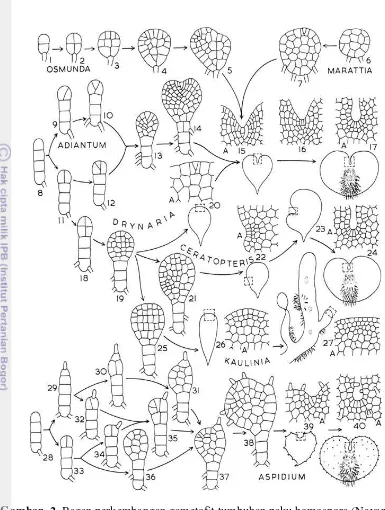
15–17=Gametofit dewasa); (Tipe Adiantum: 8=Filamen; 9–13=Piringan; 14=Gametofit muda; 15–17=Gametofit dewasa); (Tipe Drynaria: 8=Filamen; 11, 18–20=Piringan; 15–17=Gametofit dewasa); ( Tipe Ceratopteris: 8=Filamen; 11, 18, 19, 21=Piringan; 22=Gametofit muda; 23–24=Gametofit dewasa); (Tipe Kaulina: 8=Filamen; 11, 18, 19, 25=Piringan; 26=Gametofit muda; 27=Gametofit dewasa); (Tipe Aspidium: 28–29=Filamen; 30–38=Piringan; 39=Gametofit muda; 40=Gametofit dewasa) (Nayar & Kaur 1971) 7 3 Peta lokasi pengambilan sampel Pteris spp. di Pulau Jawa. 1=GunungSlamet, Baturaden, Banyumas, Jawa Tengah; 2=Sragen, Jawa Tengah; 3= Cianjur, Jawa Barat; 4=Ciomas, Bogor Jawa Barat; 5=Dramaga, Bogor, Jawa Tengah; 6=IPB, Dramaga, Bogor, Jawa Barat; 7=Mampang, Jakarta
Selatan 10
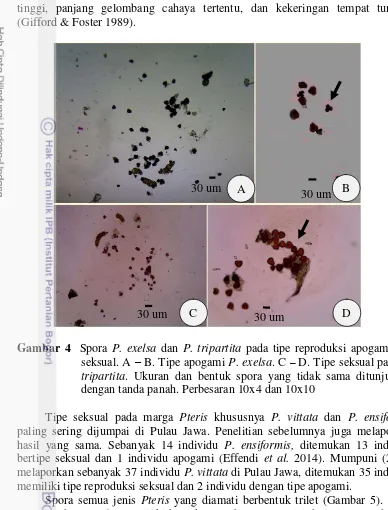
4 Spora P. exelsa dan P. tripartita pada tipe reproduksi apogami dan seksual. A – B. Tipe apogami P. exelsa. C– D. Tipe seksual pada P. tripartita. Ukuran dan bentuk spora yang tidak sama ditunjukkan dengan
tanda panah. Perbesaran 10x4 dan 10x10 14
5 Bentuk Spora Pteris. A. P. biaurita; B. P. ensiformis; C. P. exelsa; D. P. longipinnula; E. P. mertensioides; F. P. tripartita; G. P. venulosa; H. P.
vittata 15
6 Perbandingan kecepatan perkembangan gametofit Pteris spp. Perkecambahan spora ( ), pembentukan laminar ( ), pembentukan
P. longipinnula; E. P. tripartita; F. P. vittata 20
9 Persentase munculnya ekspresi seksual. Gametofit jantan ( ), gametofit hermafrodit ( ) 21 ditunjuk dengan anak panah; D-E-F. Fase spatula; G-H. Fase hati muda; I-J. Hati dewasa; K. Anteridium kosong, sperma telah keluar. m=Daerah
meristem; AN=Anteridium 25
DAFTAR GAMBAR (lanjutan)
dewasa; K. Arkegonium tampak dari atas; L. Anteridium tampak dari atas.
SS=sel sudut; m=Daerah meristem; AR=Arkegonium; AN=Anteridium
26 13 Perkembangan dan morfologi gametofit P. exelsa. A. Fase rhizoid atau
protokorm; B. Fase filamen; C-D. Pembelahan sel di ujung filamen; E-F-G. Fase spatula; H-I. Fase hati muda; J. Fase hati dewasa; K. Anteridium tampak dari samping. m=Daerah meristem; AN=Anteridium; SB=Sel
basal; SC=Sel cincin; ST=Sel tudung 27
14 Perkembangan dan morfologi gametofit P. longipinnula. A. Fase rhizoid atau protokorm; B. Fase filamen; C. Pembelahan sel di sub pangkal dan bagian tengah filamen; D-E. Fase spatula; F-G-H. Fase hati muda; I-J. Fase hewasa; K. Anteridium tampak dari atas. m=Daerah meristem;
AN=Anteridium 28
15 Perkembangan dan morfologi gametofit P. mertensioides. A. Fase rhizoid atau protokorm; B. Fase filamen; C-D. Pembelahan sel di ujung filamen; E-F. Fase spatula; G-H-I. Fase hati muda. m=Daerah meristem 29 16 Perkembangan dan morfologi gametofit P. tripartita. A. Fase rhizoid atau
protokorm; B. Fase filamen; C. Pembelahan sel secara membujur di ujung filamen; D. Pembelahan sel secara miring di ujung filamen; E. Fase spatula; F-G-H. Fase hati muda; I-J. Fase hati dewasa; K. Arkegonium tampak dari atas; L. Anteridium tampak dari atas. m=Daerah meristem;
AR=Arkegonium AN=Anteridium 30
17 Perkembangan dan morfologi gametofit P. venulosa. A-B. Fase rhizoid atau protokorm; C. Filamen; D. Pembelahan sel secara membujur di ujung filamen; E. Pembelahan sel secara miring diujung filamen; F. Fase spatula; G-H-I. Hati muda. m=Daerah meristem; SS=Sel sudut 31 18 Perkembangan dan morfologi gametofit P. vittata. A. Fase rhizoid atau
protokorm; B. Fase filamen; C. Pembelahan sel di ujung filamen; D-E. Fase spatula; F. Pembentukan daerah meristem di bagan tepi piringan; F-G. Fase hati muda; H. Fase hati dewasa; I. Anteridium dewasa tampak dari atas; J-K. Anteridium kosong tampak dari atas. m=Daerah meristem;
DAFTAR LAMPIRAN
1 Pteris biaurita L.: A. Perawakan; B. Lembaran daun; C. Bagian tengah anak daun dengan cuping-cuping subur, memperlihatkan urat daun
berareola pada sinus 39
2 Pteris ensiformis Burm.f.: A. Perawakan; B. Anak daun mandul; C. Anak daun subur; D. Bagian tengah anak daun dengan cuping-cuping subur, memperlihatkan urat daun berareola pada sinus 39 3 Pteris exelsa Gaud.: A. Perawakan; B. Daun; C. Anak daun subur; D.
Bagian tengah anak daun subur, memperlihatkan urat daun bebas pada cuping dan sorus yang indusiumnya masih tertutup 40 4 Pteris longipinnula Wall.: A. Perawakan; B. Daun; C. Bagian tengah
anak daun subur, memperlihatkan urat daun bebas pada cuping dan
sorus masak yang indusiumnya masih tertutup 40
5 Pteris mertensioides Wild.: A. Perawakan; B. Anak daun; C. Bagian tengah anak daun subur dengan cuping-cuping bersorus masak,
indusium sebagian terbuka 41
6 Pteris tripartita Sw.: A. Perawakan; B. Daun; C. Bagian tengah anak daun subur dengan cuping-cuping bersorus masak, indusium terbuka 41 7 Pteris venulosa Blume.: A. Perawakan; B. Daun subur,
memperlihatkan anak daun bersorus masak dengan indusium terbuka 42 8 Pteris vittata L.: A. Perawakan; B. Daun; C. Anak daun subur,
memperlihatkan spora masak dengan indusium terbuka 42
1 PENDAHULUAN
Pteris L. merupakan salah satu marga besar dari suku Pteridaceae yang tersebar di daerah tropis dan subtropis (Holttum 1996). Pteris di dunia diperkirakan mencapai 280 jenis (Copeland 1958) dan 19 jenis diantaranya ditemukan di pulau Jawa (Backer & Posthumus 1939). Pteris umumnya tumbuh di daerah dengan suhu hangat namun juga dapat hidup pada suhu dingin. Marga ini dapat ditemukan di daerah yang ternaungi kanopi maupun terbuka, hidup di tanah, menempel di permukaan batu kapur serta bebatuan lainnya (Holttum 1966; Tryon et al. 1990). Beberapa jenis juga dapat bertahan hidup di daerah yang terkontaminasi oleh arsenik atau logam lainnya (Ma et al. 2001).
Pteris memiliki karakter daun majemuk menyirip, mulai dari menyirip tunggal hingga menyirip ganda. Daun steril dari anggota Pteris umumnya berukuran lebih pendek dan terletak di bagian bawah, sedangkan daun fertil lebih sempit dan panjang (Holttum 1966). Jenis-jenis Pteris mudah dikenali dari karakter sorus. Sorus tersusun linier dan terletak di sepanjang tepi daun, tetapi biasanya tidak mencapai ujung anak daun. Struktur ini dilindungi oleh indusium palsu yang terbentuk dari tepi daun yang menggulung (Copeland 1958; Holttum 1966).
Secara taksonomi, Pteris sangat menarik karena ditemukan banyak jenis kompleks yang disebabkan oleh poliploidi, hibridisasi (Walker 1962), dan apogami (Chang 1991) yang dapat mengakibatkan variasi morfologi (Walker 1958). Jenis kompleks yang telah dilaporkan sebelumnya yaitu P. cadieri di Taiwan (Chao et al. 2010) dan P. cretica di Amerika (Martinez & Morbelli 2009). Apogami mempunyai kaitan erat dengan poliploidi dan mengambil peran yang sangat penting untuk terjadinya jenis kompleks (Lovis 1977).
Gametofit merupakan generasi tumbuhan paku penghasil gamet yang berkembang dari spora (Rhagavan 2005). Fase perkembangan gametofit meliputi perkecambahan spora, pembentukan laminar, bentuk dewasa, dan gametangia. Fase tersebut merupakan informasi penting untuk mempelajari evolusi, filogeni, biologi reproduksi, dan penciri tumbuhan paku (Nayar & Kaur 1971; Chiou & Farrar 1997). Tipe perkecambahan Vittaria merupakan karakter paling maju, sedangkan tipe yang paling primitif adalah Amorphous. Pada tumbuhan paku homospora tipe perkembangan gametofit yang lebih primitif yaitu tipe Adiantum, sedangkan tipe Kaulinia merupakan tipe yang paling maju (Nayar & Kaur 1971).
2
Perkembangan dan morfologi gametofit jenis-jenis tumbuhan paku di Indonesia masih sedikit diinformasikan karena baru mencakup beberapa jenis yaitu Cyathea contaminans (Handayani & Hartini 2003), Dicksonia blumei (Hartini 2005), Polypodiaceae (Nurchayati 2010), dan Asplenium nidus (Praptosuwiryo 2010). Penelitian terdahulu melaporkan bahwa suku Polypodiaceae memiliki bentuk gametofit yang sangat bervariasi. Jenis Dryopteris concolor dan Pityrogramma calomelanos memiliki protalium berbentuk hati yang memanjang, sedangkan protalium berbentuk hati melebar ditemukan pada Adiantum caudatum, Asplenium nidus, dan Nephrolepis falcate (Nurchayati 2010). Asplenium nidus memiliki tipe perkecambahan spora Vittaria dan perkembangan gametofit Aspidium (Praptosuwiryo 2010).
Informasi beberapa jenis Pteris di Indonesia masih terbatas pada kajian morfologi sporofit, tipe reproduksi, dan sitologi seperti P. biaurita, P. ensiformis, P. multifida, P. tripartita, dan P. vittata (Zubaidah 1998; Praptosuwiryo & Darnaedi 2008; Hastuti et al. 2011; Effendi et al. 2014; Mumpuni 2014). Penelitian tersebut melaporkan bahwa P. biaurita, P. ensiformis, P. multifida, dan P. vittata memiliki tingkat ploidi dan tipe reproduksi yang berbeda. Tingkat ploidi diploid dan tetraploid apogami dijumpai pada P. biaurita (Zubaidah 1998). Ploidi diploid dan triploid seksual dijumpai pada P. ensiformis var. victoriae, sedangkan tetraploid seksual ditemukan pada P. ensiformis var. ensiformis (Effendi et al. 2014). Pteris multifida memiliki dua tipe ploidi yaitu triploid apogami dan tetraploid seksual (Hastuti et al. 2011). Pteris vittata juga memiliki dua tingkat ploidi yaitu tingkat ploidi tetraploid seksual (Praptosuwiryo & Darnaedi 2008; Mumpuni 2014) dan pentaploid apogami (Mumpuni 2014). Sejauh ini penelitian mengenai perkembangan dan morfologi gametofit Pteris di Pulau Jawa belum pernah dilakukan. Tulisan ini melaporkan hasil penelitian mengenai perkembangan dan morfologi gametofit tumbuhan paku khususnya dari marga Pteris di Pulau Jawa.
Tujuan Penelitian
3
2
TINJAUAN PUSTAKA
Pteris L.
Pteris merupakan marga besar tumbuhan paku dari suku Pteridaceae yang memiliki distribusi luas. Marga ini tersebar di daerah tropis hingga subtropis (Holttum 1966). Beberapa ahli taksonomi memberikan dugaan jumlah Pteris di dunia berbeda, sekitar 250 jenis (Holttum 1966; Tryon et al. 1990) dan 280 jenis (Copeland 1958). Sementara itu, 19 jenis tersebar di Pulau Jawa yaitu P. biaurita L., P. cretica L., P. dalhousiae Hook., P. ensiformis Burm.f., P. exelsa Gaud., P. geminata Wall., P. heteromorpha Fee., P. longipinnula Wall., P. mertensioides Wild., P. multifida Poir., P. pellucida C. Perls., P. pellucens J. Agardh., P. quadriaurita Retz., P. reducta Backer., P. semipinnata L., P. tripartita Sw., P. venulosa Blume., P. vittata L., dan P. wallichiana J. Agardh. (Backer & Phosthumus 1939).
Jenis-jenis Pteris dapat tumbuh di berbagai habitat seperti di daerah terbuka maupun ternaungi kanopi, hidup di tanah, menempel di bebatuan maupun tembok, di tepi aliran sungai, dan hutan (Holttum 1966; Tryon et al. 1990; Peris 2012). Pteris di Afrika dapat hidup pada ketinggian 15 – 2750 m dpl (Peris 2012), sedangkan di Indonesia jenis-jenis dari marga ini dapat tumbuh pada ketinggian 0
– 2161 m dpl (Zubaidah 1998; Saputra & Qotrunnada 2011; Effendi et al 2014; Mumpuni 2014). Pteris vittata dapat tumbuh pada ketinggian 0 m dpl di Pulau Karimunjawa Jawa Tengah, tetapi juga mampu tumbuh pada ketinggian 2161 m dpl, dengan suhu 18.8 oC - 36.7 oC, kelembaban 35.1% - 89.4% dan intensitas
cahaya 130 - 1772 Lux (Saputra & Qotrunnada 2011; Mumpuni 2014). Effendi et al. (2014) melaporkan bahwa P. ensiformis di Jawa dapat ditemukan pada ketinggian 11 - 650 m dpl. Jenis P. biaurita tersebar pada ketinggian 400 - 600 m dpl di Jawa timur (Zubaidah 1998). Chang et al. (1992) melaporkan P. biaurita merupakan jenis yang predominan pada daerah tropis sampai subtropis dan dapat ditemukan di dataran rendah sampai dataran tinggi.
Secara etimologi bahasa Yunani, Pteris berasal dari kata “pteron” yang artinya sayap atau bulu. Anak daun yang menyerupai bulu merupakan ciri khas dari marga Pteris (Olsen 2007). Selain ciri tersebut, marga Pteris memiliki karakteristik lain yaitu daun majemuk bervariasi mulai dari menyirip tunggal hingga menyirip ganda. Daun steril terletak di bagian bawah berukuran lebih pendek dan lebar jika dibandingkan dengan daun fertil. Anak daun berhadapan dengan suatu anak daun pada bagian ujung dan anak daun paling bawah bercabang atau tidak bercabang. Sorus linier tersusun sepanjang tepi daun tetapi tidak sampai pada ujung anak daun dan dilindungi oleh indusium palsu (Holttum 1966). Marga ini mempunyai kromosom dasar x=29 (Walker 1962).
4
ujung anak daun dan tulang tengah daun bersayap, sedangkan P. ensiformis memiliki daun steril memanjang dengan bagian tepi bergigi. Pteris scabripes memiliki tepi anak daun steril yang bergerigi. Anak daun bercuping dengan tulang daun bebas dimiliki oleh P. asperula, P. dalhousiae, P. exelsa, P. grevilleana, P. longipinnula, P. mertensioides, dan P. semipinnata, sedangkan tulang daun menjala dimiliki oleh P. biaurita dan P. tripartita (Holttum 1966; Andrews 1990). Pteris mempunyai banyak manfaat untuk kehidupan manusia. Selain dimanfaatkan sebagai tanaman hias (de Winter & Amoroso 2003), jenis-jenis Pteris telah banyak digunakan sebagai obat tradisional (Chen et al. 2007). Daun P. ensiformis di Bougainvelle, digunakan untuk mengontrol menstruasi (Srivastava 2007). Daun P. ensiformis juga telah dimanfaatkan di Malaysia sebagai jus untuk obat disentri, mengobati pembengkakan di leher, dan menjaga kesehatan mulut; sedangkan di New Guinea dimanfaatkan sebagai obat bisul, borok, dan luka (de Winter & Amoroso 2003). Di Bougainvelle daun P. tripartita dimanfaatkan sebagai ramuan untuk membantu proses melahirkan (Srivastava 2007).
Beberapa penelitian menunjukkan bahwa marga Pteris memiliki berbagai macam senyawa fenol dan antioksidan sehingga dapat digunakan sebagai obat (Chen et al. 2007). Senyawa antioksidan dan fenolik pada daun Pteris memiliki peran penting yang berkaitan dengan gizi manusia sebagai agen pencegahan terhadap beberapa penyakit dan melindungi jaringan tubuh dari cekaman oksidatif (Scalbert et al. 2005). Senyawa fenolik pada P. ensiformis yang paling banyak yaitu senyawa 7-O-caffeoylhydroxymaltol 3-O-b-D-glucopyranoside, senyawa 3.5-di-O-caffeoylquinic acid, dan hispidin 4-O-b-D-glucopyranoside (Chen et al. 2007). Kandungan senyawa diterpen yang diisolasi dari Pteris telah terbukti menjadi agen antitumor dengan menginduksi apoptosis (Chen et al. 2004).
Pemanfaatan Pteris sudah diterapkan dalam aspek lingkungan. Pteris vittata di Florida telah digunakan sebagai hiperakumulasi logam berat Arsenik (As) (Ma et al. 2001). Pteris vittata tidak hanya efisien dalam mengambil jumlah besar As (2.3% berat kering tanaman) dari tanah, tetapi juga di translokasi biomassa As di atas tanah sampai 90% dari total serapan As. Oleh karena itu, memungkinkan P. vittata dimanfaatkan untuk remediasi tanah yang terkontaminasi logam (Tu & Ma 2002).
Tipe Reproduksi Tumbuhan Paku
Tumbuhan paku mengalami dua fase pergiliran keturunan yaitu fase gametofit dan sporofit yang keduanya hidup bebas. Fase gametofit merupakan fase singkat, sedangkan sporofit merupakan fase dominan. Dalam masa hidupnya, siklus hidup dan pembentukan spora pada tumbuhan paku homospora dapat terjadi secara seksual ataupun apogami (Klekowski 1973).
5 Pada siklus hidup tumbuhan paku apogami, sporofit yang terbentuk merupakan proliferasi gametofit tanpa adanya fertilisasi (Evans 1964). Tumbuhan paku tipe apogami umumnya memiliki 8 sel induk spora yang akan membentuk 32 spora yang masing-masing spora memiliki jumlah kromosom yang sama dengan sporofit induk. Satu sel induk spora mengalami tiga kali pembelahan mitosis, tetapi pada fase anafase awal mengalami gagal berpisah sehingga menghasilkan delapan sel induk spora yang masing-masing mengalami penggandaan jumlah kromosom. Sel induk spora tersebut mengalami meiosis secara normal dengan kromosom bivalen menghasilkan 32 spora (Manton 1950; Evans 1964; Klekowski 1973).
Gas etilen, asam suksinat, asam naftalena asetat (NAA), asam gibberelat, konsentrasi fosfor yang tinggi, panjang gelombang cahaya tertentu dan kekeringan tempat tumbuh dapat menginduksi terjadinya apogami (Gifford & Foster 1989). Selain itu, faktor lingkungan seperti cahaya, ketinggian tempat dan suhu daerah memiliki peran dalam pengaturan tipe reproduksi (Kato & Iwatsuki 1986). Respon setiap jenis tumbuhan terhadap ketinggian tempat tidak sama. Pteris biaurita dengan tipe reproduksi apogami paling sering dijumpai di dataran rendah, sedangkan tipe campuran semakin banyak ditemukan pada daerah yang lebih tinggi (Zubaidah 1998). Akan tetapi, pada jenis Dryopteris tipe reproduksi apogami paling sering ditemukan di dataran tinggi dan suhu dingin, dan tipe seksual lebih sering ditemukan di daerah bersuhu panas (Hirabayashi 1974).
Tingkat ploidi, jumlah spora per sporangium, ukuran spora, perkembangan gametofit, dan pembentukan serta morfologisporofit muda mempunyai kaitan erat dengan tipe reproduksi (Huang et al. 2011). Sekitar 75% tumbuhan paku apogami adalah polipoid (Walker 1962). Hal ini disebabkan oleh fenomena apogami dapat memungkinkan penggandaan jumlah kromosom (Evans 1964). Pada umumnya
ukuran spora tipe apogami lebih besar dibandingkan dengan tipe seksual, misalnya pada Diplazium (Praptosuwiryo & Darnaedi 1994). Morfologi daun dapat berkaitan dengan tipe reproduksi, seperti pada Dryopteris sparsa. Tipe seksual memiliki helaian daun lebih kecil dengan pasangan anak daun lebih jarang, basiskopik anak daun terbawah sederhana sedangkan tipe apogami helaian daun lebih lebar, pasangan anak daun lebih rapat dengan ujung meruncing, dan basiskopik anak daun terbawah lebih besar dan menyirip (Darnaedi 1987; Zubaidah 2006). Akan tetapi, pada jenis tertentu seperti pada P. multifida
morfologi daun tidak berkaitan dengan tipe reproduksi karena keragaman dan bentuknya hampir tidak berbeda (Hastuti et al. 2011).
Perkembangan Gametofit Marga Pteris
6
pembelahan sel berada di daerah ekuator merupakan karakter tipe Gleicheina, Cyathea, Hymenophyllum, Mecodium, dan Trichomanes (Nayar & Kaur 1971).
Gambar 1 Bagan tipe-tipe perkecambahan spora tumbuhan paku homospora (Nayar & Kaur 1971)
Tipe perkecambahan spora Pteris mengikuti tipe Vittaria. Pada tipe ini inisial sel membelah pada ujung bagian polar. Pembelahan selanjutnya menghasilkan filamen yang terdiri dari beberapa sel. Bagian proksimal memanjang membentuk filamen yang akhirnya akan menjadi rhizoid. Rhizoid pada umumnya tidak mengandung klorofil, cenderung tumbuh ke arah bawah, berfungsi untuk menghisap air dan nutrisi dari dalam media tumbuh (Nayar & Kaur 1971). Penelitian Zhang et al. (2008) melaporkan rhizoid pertama pada Pteris fauriei memiliki klorofil.
7
8
Tabel 1 Urutan fase perkembangan gametofit tumbuhan paku
Tipe perkembangan gametofit Nomor fase perkembangan gametofit
Osmunda 1, 2, 3, 4, 5, 15, 16, 17
Sekitar 41% dari jenis tumbuhan paku marga Pteris di Taiwan memiliki tipe reproduksi apogami (Chang 1991). Gametofit tumbuhan paku apogami umumnya tidak menghasilkan arkegonium (Knobloch 1966). Hal tersebut dikarenakan arkegonium mengalami aberasi sehingga tidak berfungsi. Akan tetapi, beberapa jenis tumbuhan paku apogami dalam masa perkembangan gametofitnya dapat menghasilkan anteridium dan arkegonium, misalnya pada jenis Diplazium megaphyllum dan Cheilanthes hirsuta. Keberadaan rambut dan sisik pada area embrionik, tracheid tanpa gumpalan, dan ada tidaknya midrib pada daun pertama juga menjadi ciri individu tersebut bertipe apogami (Chiou et al. 2006).
Keberhasilan perkecambahan spora dan perkembangan morfologi gametofit dipengaruhi oleh media tumbuh, kematangan spora, air, kelembaban, aerasi, dan pH (Toogood 1999). Perkembangan gametofit Cyatheaceae memberikan hasil yang berbeda pada media tanah, air destilata dan media agar. Spora pada media agar lebih cepat tumbuh jika dibandingkan dengan media tanah dan air destilata. Fase protallium pada media tanah dapat menghasilkan lebih banyak organ seksual. Hasil penelitian sebelumnya menunjukkan bahwa gametofit berbentuk hati serta arkegonium ditemukan pada media tanah dan agar, sedangkan pada media distilasi tidak ada arkegonium. Namun, media agar memiliki kelemahan yaitu lebih mudah terkontaminasi oleh bakteri dan jamur. Keberadaan bakteri dan jamur dapat menghambat perkembangan spora (Chen et al. 2008)
Kerapatan gametofit dapat mempengaruhi ukuran gametofit dan ekspresi seksual. Fenomena yang terjadi pada jenis Osmunda cinnamomea semakin rapat gametofit menyebabkan ukuran gametofit semakin kecil. Gametofit betina banyak ditemukan pada populasi dengan kerapatan rendah atau tinggi. Adapun gametofit jantan dan hermafrodit lebih banyak ditemukan pada populasi dengan kerapatan intermediet atau sedang. Terbentuknya gametofit jantan dan hermafrodit disebabkan adanya perkawinan antar individu (Huang et al. 2004). Gametofit betina, gametofit jantan dan monoesis pada P. ensiformis ditemukan pada kerapatan rendah sedangkan gametofit hermafrodit ditemukan pada kerapatan tinggi (Chiou 1992). Ekspresi seksual sebenarnya tidak hanya dipengaruhi oleh kerapatan gametofit tetapi juga diinduksi oleh antheridiogen yang merupakan hormon pertumbuhan dan penentu ekspresi seksual (Schneller et al. 1990).
9 jumlah sel pada filamen yang lebih sedikit. Selain itu, aktivitas pembelahan sel lebih lambat jika dibandingkan pada kondisi cahaya yang cukup (Rhagavan 2005). Penelitian Kamachi et al. (2007) menunjukkan bahwa cahaya merah mampu menekan laju perkembangan gametofit betina pada jenis tipe mutan Ceratopteris richardii.
10 Pengamatan perkembangan morfologi gametofit tumbuhan paku marga Pteris dilakukan di Laboratorium Treub Pusat Konservasi Tumbuhan, Kebun Raya Bogor, LIPI dan Laboratorium Taksonomi Tumbuhan, Departemen Biologi, Institut Pertanian Bogor (IPB).
Gambar 3 Peta lokasi pengambilan sampel Pteris spp. di Pulau Jawa. 1=Gunung Slamet, Baturaden, Banyumas, Jawa Tengah; 2=Sragen, Jawa Tengah; 3=Cianjur, Jawa Barat; 4=Ciomas, Bogor Jawa Barat; 5=Dramaga, Bogor, Jawa Tengah; 6=IPB, Dramaga, Bogor, Jawa Barat; 7= Mampang, Jakarta Selatan
Bahan Tumbuhan
Delapan jenis tumbuhan paku Pteris dengan organ lengkap dan berspora dikoleksi dari tujuh lokasi di Pulau Jawa (Tabel 2). Spesimen herbarium disimpan di Laboratorium Taksonomi Tumbuhan, Departemen Biologi, IPB.
Tabel 2 Lokasi pengambilan sampel tumbuhan paku Pteris
No Nama Jenis No
Koleksi Lokasi
1 Pteris biaurita L. DSP 050 Ds. Krikilan, Sangiran, Kab. Sragen Jawa Tengah
2 Pteris ensiformis Burm.f. DSP 035 Jl. Agatis Kampus IPB, Dramaga, Kab. Bogor 3 Pteris exelsa Gaud. DSP 045 Kebun Raya Cibodas, Cianjur, Jawa Barat
Ds. Situgedhe, Dramaga, Kab. Bogor Jawa Barat Kebun Raya Cibodas, Cianjur, Jawa Barat 6
Ds. Tanglebun, Ciomas, Kab. Bogor, Jawa Barat G. Slamet, Kemutuk Lor, Baturaden, Kab. Banyumas Jawa Tengah
11 Metode
Teknik Pengambilan Bahan Tumbuhan
Lokasi pengambilan sampel ditentukan dengan pengamatan spesimen jenis-jenis Pteris Jawa di Herbarium Bogoriense, LIPI Cibinong untuk mengetahui persebaran Pteris di Pulau Jawa. Selanjutnya dilakukan pengambilan sampel dengan menggunakan metode jelajah ke lokasi untuk mencari jenis-jenis tumbuhan paku marga Pteris pada setiap lokasi yang telah ditentukan (Tabel 2). Tumbuhan paku yang diambil berupa tumbuhan yang mempunyai organ lengkap dan telah menghasilkan spora (Rugayah et al. 2004). Sebelum dikoleksi, tumbuhan paku dengan organ lengkap tersebut didokumentasi. Tumbuhan hasil koleksi dibuat spesimen herbarium sesuai dengan metode standar (Vogel 1987). Daun subur dengan spora masak yang telah dicuci bersih dimasukkan ke dalam lipatan koran untuk pengamatan perkembangan gametofit. Material daun fertil dikumpulkan, diberi label kemudian diatur di dalam lipatan kertas koran dan diawetkan, selanjutnya untuk pengamatan tipe reproduksi.
Koleksi Spora
Spora yang telah masak ditandai dengan berubahnya warna indusium dari hijau menjadi coklat. Anak daun tempat menempelnya spora dicuci dalam air mengalir untuk menghindari terjadinya kontaminasi dengan spora dari jenis paku lain. Anak daun dikering anginkan sampai kering, kemudian dimasukkan ke dalam kertas amplop samson yang telah diberi label. Spora dipisahkan dari kotoran (bulu-bulu atau sisik-sisik daun) dengan memiringkan amplop pelan-pelan sehingga kotoran akan jatuh lebih dulu dan spora yang lebih lembut cenderung tetap melekat di dalam amplop (Chiou & Farrar 2001).
Pengamatan Tipe Reproduksi
Satu sporangium diletakkan di atas kaca benda, kemudian ditutup dengan kaca penutup. Sporangium dipecah dengan bantuan jarum dengan cara mengetukkan jarum pada kaca penutup. Spora yang tersebar dihitung dengan bantuan hand-counter. Setiap individu paling sedikit diamati 10 sporangium (Knobloch 1966). Individu yang mempunyai 32 spora pada setiap sporangiumnya digolongkan sebagai individu bertipe apogami, sedangkan yang berisi 64 spora per sporangium digolongkan sebagai individu bertipe seksual (Walker 1962).
Pengamatan Gametofit
12
yang diamati yaitu, spora berkecambah, filamen, laminar, warna rhizoid, bentuk gametofit, dan ekspresi seksual, sedangkan parameter pertumbuhan yang diamati adalah jumlah sel dan ukuran gametofit. Pengamatan perkembangan dan morfologi gametofit menggunakan mikroskop stereo, kemudian difoto dengan mikroskop cahaya setiap tujuh hari sekali sampai gametofit membentuk organ seksual (Huang et al. 2009).
Analisis Data
13
4
HASIL DAN PEMBAHASAN
Tipe Reproduksi dan Morfologi spora
Sebanyak delapan jenis Pteris telah diamati tipe reproduksinya, 4 jenis diketahui mempunyai tipe reproduksi seksual, dan 4 jenis lainnya bertipe reproduksi apogami (Tabel 3). Tipe reproduksi seksual yang memiliki 64 spora per sporangium ditemukan pada jenis P. ensiformis, P. mertensioides, P. tripartita, dan P. venulosa, sedangkan tipe reproduksi apogami ditemukan pada jenis P. biaurita, P. exelsa, P. longipinnula, dan P. vittata.
Tabel 3 Ukuran spora dan tipe reproduksi Pteris spp.
Jenis Diameter ekuatorial (μm)
Panjang Polar
(μm) Jumlah per sporangium spora
Tipe dalam tipe reproduksi apogami karena memiliki bentuk yang berbeda (Gambar 4 A – B). Mumpuni (2014) melaporkan bahwa tipe apogami pada P. vittata juga memiliki 64 spora dengan bentuk dan ukuran berbeda. Bentuk dan ukuran spora berbeda dapat dihasilkan dalam satu sporangium yang sama karena pada saat meiosis beberapa kromosom saling berpasangan dan membentuk kromosom univalen, bivalen, dan multivalen yang mengakibatkan kromosom tidak tersebar secara merata, sehingga hasil akhir meiosis menunjukkan keadaan jumlah kromosom yang tidak seimbang dan menghasilkan bentuk spora tidak normal
(Manton 1950; Klekowski 1973).
Pteris biaurita memiliki tipe reproduksi apogami. Studi sebelumnya melaporkan bahwa tipe apogami pada P. biaurita paling sering ditemukan daripada tipe seksual (Zubaidah 1998; Darnaedi 1992; Chang et al. 1992).
14
tinggi, panjang gelombang cahaya tertentu, dan kekeringan tempat tumbuh (Gifford & Foster 1989).
Gambar 4 Spora P. exelsa dan P. tripartita pada tipe reproduksi apogami dan seksual. A – B. Tipe apogami P. exelsa. C –D. Tipe seksual pada P. tripartita. Ukuran dan bentuk spora yang tidak sama ditunjukkan dengan tanda panah. Perbesaran 10x4 dan 10x10
Tipe seksual pada marga Pteris khususnya P. vittata dan P. ensiformis paling sering dijumpai di Pulau Jawa. Penelitian sebelumnya juga melaporkan hasil yang sama. Sebanyak 14 individu P. ensiformis, ditemukan 13 individu bertipe seksual dan 1 individu apogami (Effendi et al. 2014). Mumpuni (2014) melaporkan sebanyak 37 individu P. vittata di Pulau Jawa, ditemukan 35 individu memiliki tipe reproduksi seksual dan 2 individu dengan tipe apogami.
Spora semua jenis Pteris yang diamati berbentuk trilet (Gambar 5). Akan tetapi, pada P. exelsa memiliki bentuk spora beragam mulai dari trilet, tetrahedral, dan elips (Gambar 4B). Semua jenis Pteris memiliki apertur triradiat. Ukuran spora baik diameter ekuatorial dan panjang polar bervariasi antar jenis (Tabel 3). Pteris longipinnula memiliki ukuran spora paling besar dari pada jenis lain, sedangkan ukuran yang paling bervariasi terdapat pada P. exelsa. Warna spora bervariasi, spora dari P. biaurita, P. longipinnula, dan P. tripartita berwarna coklat, spora dari P. ensiformis berwarna coklat tua, spora dari P. vittata memiliki warna coklat keputihan, sedangkan spora P. venulosa berwarna kekuningan.
30 um
30 um 30 um
30 um B
C D
15
Gambar 5 Bentuk Spora Pteris. A. P. biaurita; B. P. ensiformis; C. P. exelsa; D. P. longipinnula; E. P. mertensioides; F. P. tripartita; G. P. venulosa; H. P. vittata
Perkembangan Gametofit Pteris spp.
Perkembangan gametofit tumbuhan paku secara umum terdiri dari enam fase penting yaitu fase rhizoid, fase rhizoid-protokorm, fase filamen, fase spatula, fase hati muda, dan fase hati dewasa. Fase rhizoid ditandai dengan munculnya rhizoid di bagian polar. Munculnya rhizoid dan sel pertama yang berklorofil menandai dimulainya fase rhizoid-protokorm. Pada fase filamen, sel protokorm membelah menjadi beberapa sel sehingga menghasilkan protalus berbentuk benang uniseriate. Sel-sel ujung atau sub ujung pada filamen membelah secara membujur, kemudian mengalami pembelahan sel secara berulang sehingga protalus menyerupai bentuk spatula, inilah fase spatula. Fase hati muda dimulai ketika sel-sel poligonal yang terletak di sub ujung spatula mulai membesar dan protalus mulai menjantung tetapi belum menghasilkan organ reproduksi. Pada fase hati dewasa, protalus menjatung dan telah menghasilkan organ reproduksi.
A
A B C
D E F
16
Perkecambahan Spora dan Perkembangan Gametofit
Kecepatan perkecambahan spora jenis Pteris yang diamati bervariasi (Gambar 6). Perkecambahan spora umumnya terjadi pada 10-20 hari setelah semai (HSS). Pteris ensiformis mengalami perkecambahan yang paling cepat yaitu 10 HSS. Pteris mertensioides merupakan jenis yang paling lambat berkecambah sekitar 20 HSS.
Perkecambahan spora jenis Pteris mengikuti tipe Vittaria. Karakteristik tipe Vittaria ditandai dengan sel pertama yang terbentuk tegak lurus dengan rhizoid (Nayar & Kaur 1971). Tipe Vittaria nampaknya umum terjadi dalam marga Pteris seperti yang dideskripsikan oleh Mendoza et al. (1997), Prada et al. (2008), Zhang et al. (2008), Martínez (2010), dan Martinez et al. (2013).
Gambar 6 Perbandingan kecepatan perkembangan gametofit Pteris spp. Perkecambahan spora ( ), pembentukan laminar ( ), pembentukan organ seksual ( )
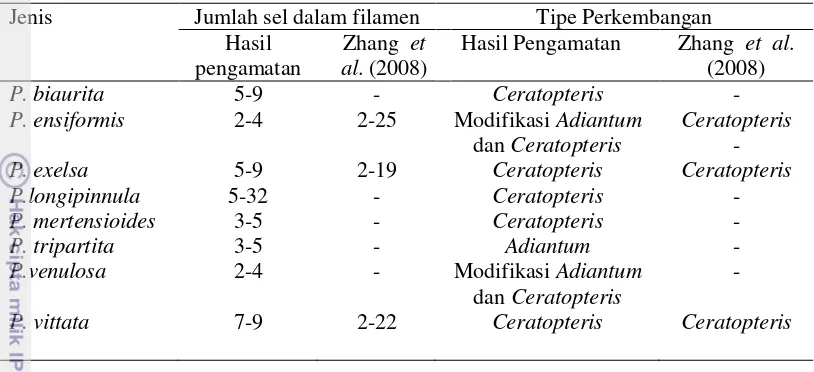
17 Tabel 4 Perbandingan perkembangan gametofit antara hasil pengamatan dengan
hasil Zhang et al. (2008)
Jenis Jumlah sel dalam filamen Tipe Perkembangan Hasil
P. ensiformis 2-4 2-25 Modifikasi Adiantum
dan Ceratopteris
Ceratopteris
-
P. exelsa 5-9 2-19 Ceratopteris Ceratopteris P.longipinnula
P. vittata 7-9 2-22 Ceratopteris Ceratopteris
Tipe perkembangan gametofit jenis Pteris yang diamati bervariasi yaitu tipe Ceratopteris, Adiantum, dan modifikasi antara kedua tipe tersebut. Akan tetapi, penelitian sebelumnya melaporkan bahwa perkembangan gametofit yang terjadi di dalam marga Pteris umumnya memiliki tipe Ceratopteris (Nayar & Kaur 1971).
Pteris biaurita, P. exelsa, P. longipinnula, P. mertensioides, dan P. vittata memiliki tipe Ceratopteris (Gambar 11,13,14,15). Tipe perkembangan tersebut mirip dengan P. deflexa, P. tristicula, P. cretica, P. berteroana, P. fauriei, P. incompleta, P. multifida, P. vittata, dan P. wallichiana (Martínez 2010; Martinez et al. 2013; Zhang et al. 2008; Prada et al. 2008). Pada tipe Ceratopteris, pembelahan sel pertama terjadi secara longitudinal di bagian ujung kadang-kadang di sub pangkal, selanjutnya diikuti pembelahan secara transversal. Pembelahan sel pertama secara longitudinal terjadi pada posisi sel berbeda tergantung jenis Pteris, pada sel ke-2 pada P. venulosa dan P. ensiformis. Pada jenis P. vittata mengalami pembelahan sel pertama secara longitudinal pada sel ke-8 (Tabel 5). Seiring dengan berjalannya waktu, sel-sel membelah sehingga terbentuk piringan. Letak daerah meristem berada di tepi sehingga protalium muda yang terbentuk tidak simetri. Sel-sel meristem membelah secara terus-menerus menghasilkan bentuk gametofit menjatung.
Tabel 5 Inisiasi pembelahan sel awal pada filamen
Jenis Inisiasi pembelahan pada sel ke-
P. biaurita 7 dan 9
18
Pteris ensiformis dan P. venulosa memiliki tipe perkembangan gametofit yang unik yaitu merupakan modifikasi dari tipe Adiantum dan tipe Ceratopteris. Pada fase perkembangan awal tersebut, sel ujung mengalami pembelahan sel secara miring. Selanjutnya diikuti pembelahan sel ke-2 yang mengakibatkan sel bagian ujung menyudut. Fase perkembangan selanjutnya mengikuti tipe Ceratopteris karena sel-sel meristem berada di bagian tepi sehingga gametofit yang terbentuk tidak simetris. Sel-sel meristem terus membelah seiring berjalannya waktu mengakibatkan gametofit berbentuk menjantung. Penelitian sebelumnya melaporkan bahwa jenis P. ensiformis memiliki tipe perkembangan gametofit yaitu tipe Ceratopteris (Chiou 1992; Zhang et al. 2008).
Berdasarkan deskripsi Nayar & Kaur (1971), perkembangan gametofit P. tripartita mengikuti tipe Adiantum. Pembelahan sel berawal secara longitudinal, diikuti pembelahan di sisi bagian kiri secara miring sehingga sel yang terbentuk menyudut. Pembelahan sel secara longitudinal dan transversal terjadi secara berulang, sehingga membentuk piringan. Letak sel-sel meristematik terbentuk di bagian ujung gametofit. Selanjutnya gametofit berbentuk menjantung karena adanya pembelahan berulang pada daerah notch (sel-sel meristem). Jenis P. tripartita membentuk notch paling awal dibandingkan jenis lain. Namun hasil penelitian sebelumnya melaporkan bahwa perkembangan gametofit P. tripartita dari India termasuk tipe Ceratopteris (Ravi et al. 2014).
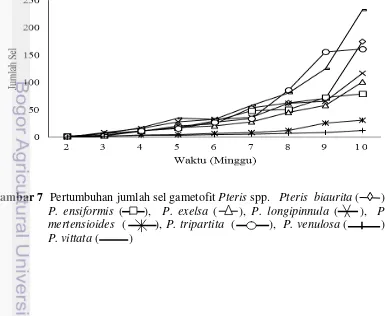
Secara umum, jumlah sel bertambah seiring dengan bertambahnya umur gametofit, tetapi jumlah sel antar jenis mengalami penambahan sel yang bervariasi (Gambar 7). Tingkat pertumbuhan gametofit antar jenis Pteris mulai berbeda pada minggu ke-2, dan pertumbuhan gametofit terus berbeda seiring bertambahnya usia gametofit. Pteris biaurita, P. ensiformis, P. exelsa, P. tripartita, dan P. vittata mulai mengalami perkecambahan pada minggu ke-2 dan menghasilkan sekitar 1 sampai 6 sel. Jenis P. longipinnula, P. mertensioides, dan P. venulosa mulai menghasilkan sel pada minggu ke-3.
19 Pteris vittata menunjukkan tingkat pertumbuhan paling cepat pada minggu ke-4 hingga minggu ke-7. Pada minggu ke-8 hingga minggu ke-9, P. vittata mengalami pertumbuhan sel gametofit yang lebih lambat daripada P. tripartita, kemudian mengalami pertumbuhan sel gametofit paling cepat pada minggu ke-10. Pteris venulosa merupakan jenis yang mengalami pertumbuhan sel paling lambat pada setiap minggunya dibandingkan dengan jenis lain. Pertumbuhan sel tiap minggunya sangat sedikit sekitar 1 sampai 6 sel.
Analisis data pertumbuhan sel dengan ANOVA dan uji lanjut Duncan menunjukan terdapat perbedaan pertumbuhan sel antar jenis secara nyata (Tabel 6; Lampiran 1 dan 2). berkembang dengan baik. Kedua jenis tersebut mengalami pertumbuhan gametofit yang sangat lambat jika dibandingkan dengan jenis lain. Penambahan ukuran gametofit dan jumlah sel juga sangat lambat. Fase pembentukan laminar P. mertensiodes terjadi pada 111 HSS dan 180 HSS pada P. venulosa, sedangkan jenis lainnya pada umur yang lebih muda yaitu 40–92 HSS telah menghasilkan organ reproduksi. Pertumbuhan dan perkembangan yang lambat ini kemungkinan disebabkan oleh faktor lingkungan yang kurang mendukung untuk pertumbuhan spora seperti tempat tumbuh, kelembaban, dan suhu. Habitat alami P. venulosa sangat menyukai daerah yang sangat lembab dan teduh, tumbuh di dekat aliran sungai. Pteris mertensioides merupakan koleksi di Kebun Raya Cibodas dengan distribusi yang sempit dan jarang ditemukan. Kedua jenis tersebut tumbuh di habitat asli pada suhu sekitar 17–270C, sedangkan suhu ruang di laboratorium percobaan sekitar 27–310C. Keberhasilan perkecambahan spora dan perkembangan gametofit dipengaruhi oleh media tumbuh, kematangan spora, air, kelembaban, aerasi, dan pH (Toogood 1999).
Gametofit Dewasa dan Ekspresi seksual
Bentuk gametofit dewasa pada jenis-jenis Pteris yang diamati bervariasi. Gametofit P. tripartita berbentuk menjantung simetri, sedangkan jenis lainnya berbentuk menjantung asimetri. Pteris biaurita memiliki ujung gametofit bergelombang, sedangkan jenis lainnya memiliki ujung rata (Gambar 8).
20
Pteris yang diamati yaitu P. biaurita, P. exelsa, P. longipinnula, dan P. vittata menghasilkan 100% gametofit jantan, sedangkan dua jenis lainnya yaitu P. ensiformis dan P. tripartita menghasilkan gametofit dewasa jantan dan hermafrodit. Spora P. ensiformis menghasilkan sekitar 83 % gametofit jantan dan 17 % gametofit hermafrodit, sedangkan pada P. tripartita menghasilkan 73% gametofit jantan dan 27% gametofit hermafrodit (Gambar 9).
Gambar 8 Bentuk ujung gametofit. A. P. biaurita; B. P. ensiformis; C. P. exelsa; D. P. longipinnula; E. P. tripartita; F. P. vittata
Hasil penelitian ini berbeda dengan penelitian sebelumnya yang melaporkan bahwa P. exelsa, P. ensiformis, dan P. vittata menghasilkan 100% gametofit bertipe hermafrodit (Zhang et al. 2008; Martinez 2010). Perbedaan hasil penelitian tersebut kemungkinan diakibatkan faktor dalam yaitu tipe reproduksi dan faktor luar seperti nutrisi, kerapatan gametofit, pengaruh cahaya, serta interaksi antara gametofit (Knobloch 1966; Huang et al. 2004; De Soto et al. 2008).
C
D F F
D E F
C B
21
Gambar 9 Persentase munculnya ekspresi seksual. Gametofit jantan ( ), gametofit hermafrodit ( )
Pembentukkan organ seksual antara jenis tumbuhan paku yang diamati membutuhkan waktu yang berbeda (Gambar 7). Semua gametofit jantan maupun hermafrodit pada jenis Pteris menghasilkan anteridium terlebih dahulu. Hal ini berbeda dengan P. vittata yang menghasilkan arkegonium terlebih dahulu (Khare & Kaur 1983). Dari semua jenis yang telah menghasilkan organ reproduksi, gametofit Pteris mulai menghasilkan anteridium pada 42 – 92 HSS (Tabel 7). Pembentukan organ seksual yang paling awal terjadi pada P. vittata, sekitar 40 HSS, sedangkan P. longipinnula menghasilkan anteridium paling akhir pada hari ke-92 setelah semai. Organ anteridium berbentuk bulat telur, tersusun atas sel tudung, sel cincin dan sel basal (Gambar 10A). Pada umumnya anteridia terbentuk pada permukaan sayap, bagian belakang dekat takik, dan tepi sayap. Arkegonium terbentuk di bagian dekat takik. Sel kanal pada arkegonium akan terbuka ketika terjadi fertilisasi (Gambar 10B).
Gambar 10 Organ seksual. A. Anteridium tampak dari samping; B. Arkegonium tampak dari atas. ST= Sel tudung; SC=Sel cincin; SB=Sel basal; AN=Anteridium; AR= Arkegonium
Ukuran gametofit dewasa berbeda antara jenis Pteris (Tabel 7). Pteris vittata memiliki ukuran gametofit dewasa paling sempit (0.53 – 0.77 mm x 0.18 – 0.40 mm). Ukuran gametofit paling besar dimiliki oleh P. ensiformis, 1.23 mm – 1.74 mm x 0.80 – 2.10 mm.
A B
R SB
SC ST
A B
AN
22
Tabel 7 Ukuran gametofit dewasa tumbuhan paku Pteris
Jenis Gametofit Dewasa Umur gametofit
(hari setelah semai )
Jenis-jenis yang bertipe reproduksi apogami yaitu P. exelsa, P. biaurita, P. longipinnula, dan P. vittata menghasilkan anteridium yang fungsional dan tidak menghasilkan arkegonium. Tipe reproduksi mempengaruhi ekspresi seksual gametofit. Gametofit tumbuhan paku apogami biasanya menghasilkan anteridium yang fungsional dan menghasilkan arkegonium (Knobloch 1966). Anteridium fungsional ditandai dengan berubahnya bentuk anteridium setelah sperma keluar (Nayar & Kaur 1971). Kejadian yang sama juga ditemukan pada P. cretica, P. pellucidifolia, dan P. wulaensis (Huang et al. 2011). Akan tetapi, P. wulaensis menghasilkan arkegonium dan terdapat embrio yang mengindikasi individu tersebut apogami. Apogami P. cretica, Diplazium megaphyllym, dan Cheilanthes hirsuta menghasilkan gametofit hermafrodit, tetapi arkegonium kehilangan fungsi karena saluran sel leher tidak terbuka (Laird & Sheffield 1986; Chiou et al. 2006; Huang et al. 2009).
Fase perkembangan gametofit meliputi fase rhizoid, fase rhizoid-protokorm, fase filamen, fase spatula, fase hati muda, dan fase hati dewasa. Fase-fase tersebut mempunyai pola yang konstan dan stabil pada tingkatan takson yang lebih tinggi. Selain itu, fase perkembangan gametofit memiliki sedikit karakter dan sifatnya tumpang tindih sehingga fase ini tidak dapat digunakan sebagai pembeda antar jenis (Nayar & Kaur 1971).
Deskripsi Morfologi Gametofit Pteris spp.
1. Pteris biaurita L. (Gambar 11)
23 tampak dari samping tersusun atas sel basal, sel cincin dan sel tudung, tersebar di seluruh permukaan.
2. Pteris ensiformis Burm.f. (Gambar 12)
Spora trilet, diameter 31.3-36.8 μm, panjang polar 29.8-34.5 μm. Tipe perkecambahan Vittaria, rhizoid uniseriate tegak lurus dengan sel pertama. Filamen terdiri atas 2-4 sel, pembelahan sel secara miring, sel ujung filamen menyudut. Gametofit muda asimetri; sel-sel meristem terbentuk di bagian lateral. Gametofit dewasa asimetri, agak menjantung, 1.23-1.70 x 0.80-2.10 mm, ujung membundar; cuping terbuka atau tertutup di atas dasar sinus; pangkal membaji dan menyempit; rhizoid terletak di bagian pangkal sampai bagian tengah. Sayap asimetri, tepi rata; sel memanjang atau poligonal, dinding sel bagian tepi hampir lurus, sel tersusun rapat; takik cekung. Tulang tengah seperti saluran, bagian distal memiliki bantalan. Anteridium bulat tampak dari atas; tampak dari samping tersusun atas sel basal, sel cincin dan sel tudung, tersebar di seluruh permukaan. Arkegonium terdiri atas 4 sel leher, terletak di dekat takik.
3.Pteris exelsa Gaud. (Gambar 13)
Spora trilet, diameter 21.4-50.9 μm, panjang polar 17.9-46.7 μm. Tipe perkecambahan Vittaria, rhizoid uniseriate tegak lurus dengan sel pertama. Filamen terdiri atas 5-9 sel; pembelahan sel longitudinal diikuti transversal di ujung filamen. Gametofit muda asimetri; sel-sel meristem terbentuk di bagian lateral. Gametofit dewasa asimetri, agak menjantung, 0.71-1.78 x 0.79-1.78 mm, ujung membundar; cuping terbuka atau tertutup di atas dasar sinus; pangkal membaji dan menyempit; rhizoid terletak di bagian pangkal sampai bagian tengah. Sayap asimetri, tepi rata; sel memanjang atau poligonal, dinding sel bagian tepi hampir lurus, sel tersusun rapat; takik cekung. Tulang tengah seperti saluran, bagian distal memiliki bantalan. Anteridium bulat tampak dari atas; tampak dari samping tersusun atas sel basal, sel cincin dan sel tudung, tersebar di seluruh permukaan.
4. Pteris longipinnula Wall. (Gambar 14)
Spora trilet, diameter 50.8-57.6 μm, panjang polar 44.5-50.4 μm. Tipe perkecambahan Vittaria, rhizoid uniseriate tegak lurus dengan sel pertama. Filamen terdiri atas 5-32 sel; pembelahan sel longitudinal diikuti transversal di ujung filamen. Gametofit muda asimetri; sel-sel meristem terbentuk di bagian lateral. Gametofit dewasa asimetri, agak menjantung, 1.18-1.76 x 0.56-1.52 mm, ujung membundar; cuping terbuka atau tertutup di atas dasar sinus; pangkal membaji dan menyempit; rhizoid terletak di bagian pangkal sampai bagian tengah. Sayap asimetri, tepi rata; sel memanjang atau poligonal, dinding sel bagian tepi hampir lurus, sel tersusun rapat; takik cekung. Tulang tengah seperti saluran, bagian distal memiliki bantalan. Anteridium bulat tampak dari atas; tampak dari samping tersusun atas sel basal, sel cincin dan sel tudung, tersebar di seluruh permukaan.
5. Pteris mertensioides Wild. (Gambar 15)
24
Filamen terdiri atas 3-5 sel; pembelahan sel longitudinal diikuti transversal di ujung filamen. Gametofit muda asimetri; sel-sel meristem terbentuk di bagian lateral.
6. Pteris triparita Sw. (Gambar 16)
Spora trilet, diameter 50.2-51.6 μm, panjang polar 40.7-48.7 μm. Tipe perkecambahan Vittaria, rhizoid uniseriate tegak lurus dengan sel pertama. Filamen terdiri atas 3-5 sel; pembelahan sel secara miring, sel ujung filamen menyudut. Gametofit muda asimetri; sel-sel meristem terbentuk di bagian lateral. Gametofit dewasa asimetri, agak menjantung, 0.53 - 0.77 x 0.18- 0.40 mm, ujung membundar; cuping terbuka atau tertutup di atas dasar sinus; pangkal membaji dan menyempit; rhizoid terletak di bagian pangkal sampai bagian tengah. Sayap asimetri, tepi rata; sel memanjang atau poligonal, dinding sel bagian tepi hampir lurus, sel tersusun rapat; takik cekung. Tulang tengah seperti saluran, bagian distal memiliki bantalan. Anteridium bulat tampak dari atas; tampak dari samping tersusun atas sel basal, sel cincin dan sel tudung, tersebar di seluruh permukaan. Arkegonium terdiri atas 4 sel leher, terletak di dekat takik.
7. Pteris venulosa Blume. (Gambar 17)
Spora trilet, diameter 32.5-37.4 μm, panjang polar 33.8-41.1 μm. Tipe perkecambahan Vittaria, rhizoid uniseriate tegak lurus dengan sel pertama. Filamen terdiri atas 3-5 sel; pembelahan sel secara miring, sel ujung filamen menyudut. Gametofit muda asimetri; sel-sel meristem terbentuk di bagian lateral.
8. Pteris vittata L. (Gambar 18)
25
Gambar 11 Perkembangan dan morfologi gametofit P. biaurita. A. Fase rhizoid atau protokorm; B. Fase filamen; C.
Pembelahan sel secara membujur di ujung filamen; D-E-F. Fase spatula; G-H. Fase hati muda; I-J. Fase hati dewasa; K. Anteridium kosong, sperma telah keluar. m=Daerah meristem; AN=Anteridium
C
K J
I
D E F
G H I
J K
m
m
m
AN
26
Gambar 12 Perkembangan dan morfologi gametofit P. ensiformis. A. Fase rhizoid atau protokorm; B. Fase filamen; C. Pembelahan sel secara miring di ujung filamen; D-E. Fase spatula; F-G-H-I. Fase hati muda; J. Fase hati dewasa; K. Arkegonium tampak dari atas; L. Anteridium tampak dari atas. SS=sel sudut; m=Daerah meristem; AR=Arkegonium; AN=Anteridium
D
C B
J I
G
A B C
D
K m
m
m
AR
AN m
SS
E F
L H
G
J
27
Gambar 13 Perkembangan dan morfologi gametofit P. exelsa. A. Fase rhizoid atau protokorm; B. Fase filamen; C-D. Pembelahan sel di sub ujung filamen; E-F-G. Fase spatula; H-I. Fase hati; J. Fase hati dewasa; K. Anteridium tampak dari samping. m=Daerah meristem; AN=Anteridium; SB=Sel basal; SC=Sel cincin; ST=Sel tudung
D
H G
F E
I
m
m
SC
m
E F
H G
A B C
D
I
K J
SB
28
Gambar 14 Perkembangan dan morfologi gametofit P. longipinnula. A. Fase rhizoid atau protokorm; B. Fase filamen; C.
Pembelahan sel di sub ujung dan bagian tengah filamen; D-E. Fase spatula; F-G-H. Fase hati muda; I-J. Fase hati dewasa; K. Anteridium tampak dari atas. m=Daerah meristem; AN=Anteridium
m
m
AN
m
m
A B C
D E F
G H I
29
Gambar 15 Perkembangan dan morfologi gametofit P. mertensioides. A. Fase rhizoid atau protokorm; B. Fase filamen; C-D. Pembelahan sel secara membujur di ujung filamen; E-F. Fase spatula; G-H-I. Fase hati muda. m=Daerah meristem
A
A
A A
A B C
D E F
G H I
m m
30
Gambar 16 Perkembangan dan morfologi gametofit P. tripartita. A. Fase rhizoid atau protokorm; B. Fase filamen; C. Pembelahan sel secara membujur di ujung filamen; D. Pembelahan sel secara miring di ujung filamen; E. Fase spatula; F-G-H. Fase hati muda; I-J. Fase hati dewasa; K. Arkegonium tampak dari atas; L. Anteridium tampak dari atas. m=Daerah meristem; AR=Arkegonium AN=Anteridium
D
C B
A
E
K I H
G
A B C
D E F
G H I
J K L
m
m
AR
31
Gambar 17 Perkembangan dan morfologi gametofit P. venulosa. A-B. Fase rhizoid atau protokorm; C. Filamen; D. Pembelahan sel secara membujur di ujung filamen; E. Pembelahan sel secara miring diujung filamen; F. Fase spatula; G-H-I. Fase hati muda. m=Daerah meristem; SS=sel sudut
A
D
B C
E
G H I
F SS
32
Gambar 18 Perkembangan dan morfologi gametofit P. vittata. A. Fase rhizoid atau protokorm; B. Fase filamen; C. Pembelahan sel di ujung filamen; D-E. Fase spatula; F. Pembentukan daerah meristem di bagan tepi piringan; F-G. Fase hati muda; H. Fase hati dewasa; I. Anteridium dewasa tampak dari atas; J-K. Anteridium kosong tampak dari atas. m=Daerah meristem; AN=Anteridium
A
J
B C
D E F
G H I
K m
AN
33
5
SIMPULAN DAN SARAN
Simpulan
Spora jenis-jenis Pteris memiliki bentuk trilet dan memiliki warna yang bervariasi. Ukuran spora bervariasi tergantung jenis dan tipe reproduksinya. Jenis spora yang mempunyai ukuran paling besar yaitu P. longipinnula (diameter ekuatorial 50.8-57.6 μm dan panjang polar 44.5-50.5 μm), sedangkan P. exelsa mempunyai ukuran spora paling bervariasi (diameter ekuatorial 21.4- 50.9μm dan panjang polar 17.9-46.7 μm).
Tipe reproduksi seksual ditemukan pada P. ensiformis, P. mertensioides, P. tripartita, dan P. venulosa, sedangkan tipe reproduksi apogami ditemukan pada jenis P. biaurita, P. exelsa, P. longipinnula, dan P. vittata. Jenis Pteris apogami tidak selalu memiliki 32 spora per sporangium. Jenis P. biaurita, P. exelsa, P. longipinnula, dan P. vittata mempunyai tipe reproduksi apogami dengan jumlah spora 32 per sporangium, sedangkan P. exelsa apogami dengan 64 spora per sporangiumnya.
Secara umum, perkembangan gametofit jenis Pteris yang diamati memiliki fase sebagai berikut: fase rhizoid, fase rhizoid-protokorm, fase filamen, fase spatula, fase hati muda, dan fase hati dewasa. Perkecambahan spora jenis Pteris mengikuti tipe Vittaria dicirikan oleh sel-sel yang terbentuk tegak lurus dengan rhizoid. Perkecambahan spora umumnya terjadi antara 10-20 HSS. Jumlah sel dalam filamen bervariasi antara 2-32 sel.
Perkembangan gametofit jenis Pteris bervariasi yaitu tipe Ceratopteris, Adiantum, dan modifikasi antar kedua tipe tersebut. Lima jenis Pteris yaitu P. biaurita, P. exelsa, P. longipinnula, P. mertensioides, dan P. vittata mengikuti tipe Ceratopteris. Perkembangan gametofit P. tripartita mengikuti tipe Adiantum. Pteris ensiformis dan P. venulosa mengikuti tipe Adiantum pada awal perkembangannya dan fase selanjutnya mengikuti Ceratopteris.
Bentuk gametofit dewasa bervariasi. Pteris tripartita mempunyai bentuk menjantung simetri, sedangkan bentuk gametofit jenis lainnya menjantung asimetri. Gametofit mulai menghasilkan anteridium pada 40-92 HSS. Berdasarkan ANOVA dan uji lanjut Duncan, kecepatan pertumbuhan sel berbeda antara jenis Pteris. Pteris vittata mengalami pertumbuhan sel paling cepat, pada minggu ke-10 memiliki jumlah sel sekitar 232.27. Pertumbuhan sel paling lambat dialami oleh P. venulosa dengan jumlah sel pada minggu ke-10 sekitar 12.43. Perbedaan perkembangan gametofit antara jenis Pteris diantaranya jumlah sel dalam filamen, lama perkecambahan spora, waktu pembentukan notch, waktu pembentukan organ seksual, bentuk gametofit dewasa, dan tipe perkembangan gametofit. Dengan demikian, data perkembangan gametofit tidak dapat digunakan sebagai pembeda antar jenis Pteris.
Saran