STABILITAS POTENSI SEDIAAN AKHIR KANDIDAT
FORMULASI VAKSIN ROTAVIRUS PADA
BERBAGAI TEKNIK FREEZING DAN SUHU
PENYIMPANAN
SKRIPSI
Diajukan sebagai salah satu syarat untuk memperoleh gelar Sarjana Farmasi
GIAN PERTELA
NIM. 109102000041
FAKULTAS KEDOKTERAN DAN ILMU KESEHATAN
PROGRAM STUDI FARMASI
Nama : Gian Pertela Program Studi : Strata-1 Farmasi
Judul : Stabilitas Potensi Sediaan Akhir Kandidat Formulasi Vaksin Rotavirus Pada Berbagai Teknik Freezing dan Suhu Penyimpanan
Pada penelitian ini dilakukan pengamatan terhadap pengaruh teknik freezing dan suhu penyimpanan terhadap stabilitas formulasi kandidat vaksin rotavirus. Pengamatan ini ditujukan untuk mendapatkan kombinasi yang paling optimal antara teknik freezing pada suhu -70oC, -152oC, dan nitrogen cair dengan suhu penyimpanan 2 – 8oC, -20oC, dan -70oC. Perlakuan lama penyimpanan adalah 30, 60, 90, dan 120 hari dengan titer virus sebagai parameter kestabilan vaksin. Uji titrasi virus dilakukan menggunakan metode indirect immunofluorescence assay dengan penanda antibodi poliklonal rabbit anti SA 11 sebagai antibodi primer dan antibodi IgG goat anti rabbit sebagai antibodi sekunder untuk mengamati jumlah rotavirus dalam tiap vial kandidat formulasi vaksin.
Data hasil pengamatan menunjukkan bahwa teknik freezing dan suhu penyimpanan berpengaruh terhadap kestabilan kandidat formulasi vaksin. Teknik freezing -152oC dan suhu penyimpanan 2 – 8oC masing-masing memiliki titer paling rendah dibandingkan dengan teknik freezing dan suhu penyimpanan lainnya. Secara keseluruhan, semua teknik freezing dan suhu penyimpanan tidak menunjukkan penurunan titer yang signifikan pada lama penyimpanan 42, 60, 90 dan 120 hari.
Name : Gian Pertela Program Study: Strata-1 Pharmacy
Title : The Stability of Final Preparations Potency of Rotavirus Vaccine Formulation Candidate on Various Freezing Technique and Temperature Storage.
This study is an observation on influence technique of freezing and temperature storage on the stability of the final preparations potency of rotavirus vaccine formulation candidate. This research is aimed at attaining optimal combination between freezing technique at the temperature of -70oC, -152oC, and liquid nitrogen by using the storage temperature of 2-8oC, -20oC, and -70oC. The duration of the storage is 42, 60, 90, and 120 days by the virus titer as vaccine stability parameter. The test on virus titration is committed by using the method of indirect immunofluorescence assay with the antibody signifier of polyclonal rabbit anti SA as primary antibody and IgG goat anti rabbit as secondary antibody to observe the account of rotavirus in each vaccine formulation candidate.
The data of observation result showed that freezing technique and storage temperature have significant influence on the stability of vaccine formulation candidate. The freezing technique of -152oC and storage temperature of 2-8oC have the lowest titer in comparison with the other freezing technique and storage temperature. In conclusion, all freezing techniques and storage temperatures do not show the significant titer decreasing at the storage of 42, 60, 90 and 120 days.
Puji serta syukur saya panjatkan kepada kehadirat Allah SWT yang telah melimpahkan berbagai macam nikmat, karunia, serta kasih sayang-Nya sehingga saya dapat menyelesaikan skripsi ini. Penulisan skripsi dengan judul “Stabilitas Potensi Sediaan Akhir Kandidat Formulasi Vaksin Rotavirus Pada Berbagai Teknik Freezing dan Suhu Penyimpanan” ini dilakukan dalam rangka memenuhi salah satu syarat untuk mencapai gelar Sarjana Farmasi pada Program Studi Farmasi, Fakultas Kedokteran dan Ilmu Kesehatan, Universitas Islam Negeri (UIN) Syarif Hidayatullah Jakarta.
Saya menyadari bahwa tanpa bantuan dan bimbingan dari berbagai pihak, mulai dari masa perkuliahan sampai pada penyusunan skripsi ini, pasti sangatlah sulit bagi saya untuk menyelesaikan skripsi ini. Oleh karena itu, saya mengucapkan terima kasih kepada :
1. Ibu Yuni Anggraeni, M. Farm., Apt. selaku pembimbing pertama dan Ibu drh. Maharani, M. Si selaku Pembimbing kedua, yang memiliki andil besar dalam proses penelitian dan penyelesaian tugas akhir saya ini, semoga segala bantuan dan bimbingan Ibu mendapat imbalan yang lebih baik di sisi-Nya.
2. Bapak Dr. Elvyn Fajrul Jaya Saputra, M. Kes selaku Direktur Perencanaan dan Pengembangan PT Bio Farma (Persero), yang telah bersedia menerima saya untuk melakukan observasi dan penelitian, serta menggunakan segala fasilitas selama penelitian di Divisi Penelitian dan Pengembangan, PT Bio Farma (Persero).
3. Staf Divisi Penelitian dan Pengembangan PT Bio Farma (Persero), Bu Sophia, Pak Nailul, Bu Dini, Bu Niar, Bu Anna, yang telah berkenan menerima saya dengan sangat baik dan memberikan pengalaman yang sangat berharga selama penelitian.
Syarif Hidayatullah Jakarta.
6. Bapak dan Ibu staf pengajar dan karyawan yang telah memberikan bimbingan, bantuan, dan ilmunya selama saya menempuh pendidikan di Program Studi Farmasi, Fakultas Kedokteran dan Ilmu Kesehatan Universitas Islam Negeri (UIN) Syarif Hidayatullah Jakarta.
7. Ayahanda Ir. Burhan Ramdan dan Ibunda Ir. Aswariny Hamid yang sampai saat ini masih mendampingi saya setiap saat dan dengan terus menerus memberikan panutan, kasih sayang, dukungan, motivasi, doa, dan nasehatnya yang tak akan pernah mampu untuk saya membalas semuanya, semoga keduanya mendapatkan balasan yang jauh lebih baik di sisi-Nya. 8. Kakak saya Geibril Kafrawi, S. Kom. satu-satunya saudara kandung yang
selalu memberikan bantuan dan dukungannya.
9. Keluarga kedua saya, seluruh rekan-rekan PSF 2009 yang selalu saya sayangi, Arif, Irsyad, Indah, Nadya.
10.Kakak-kakak senior PSF 2008 dan 2007, adik-adik junior PSF 2010 dan 2011, serta keluarga kecil saya di Badan Eksekutif Mahasiswa (BEM) Farmasi periode 2011 – 2012, Fakultas Kedokteran dan Ilmu Kesehatan Universitas Islam Negeri (UIN) Syarif Hidayatullah Jakarta.
11.Keluarga besar Paduan Suara Mahasiswa (PSM) Universitas Islam Negeri (UIN) Syarif Hidayatullah dan angkatan Maximilian.
12.Serta semua pihak yang telah membantu penulis selama ini yang tidak bisa disebutkan satu persatu.
Akhir kata, saya berharap Allah SWT berkenan membalas segala kebaikan semua pihak yang telah membantu saya hingga saat ini. Semoga skripsi ini dapat membawa manfaat bagi pengembangan ilmu pengetahuan.
Halaman
HALAMAN JUDUL ……….……… ii
HALAMAN PERNYATAAN ORISINALITAS ……… iii
HALAMAN PERSETUJUAN PEMBIMBING ……… iv
HALAMAN PENGESAHAN ……….. v
ABSTRAK ………….……… vi
ABSTRACT ……… vii
KATA PENGHANTAR …..……… viii
DAFTAR ISI ……....……… x
DAFTAR TABEL ……… xii
DAFTAR GAMBAR ……… xiii
DAFTAR LAMPIRAN ……… xiv
BAB 1 PENDAHULUAN ………….………. 1
1.1Latar Belakang ..……….……… 1
1.2Perumusan Masalah ..………. 3
1.3Tujuan Penelitian ..………. 4
1.4Manfaat Penelitian .……… 4
BAB 2 TINJAUAN PUSTAKA ……… 5
2.1 Virus ………..………. 5
2.1.1 Karakteristik Virus .……..……… 6
2.1.2 Klasifikasi Virus ..……… 7
2.1.3 Replikasi Virus …………..……… 8
2.2 Rotavirus ..………. 9
2.2.1 Karakteristik Rotavirus ………. 9
2.2.2 Epidemiologi dan Imunitas ………..………… 12
2.2.3 Respon Imun Tubuh ………. 14
2.3 Vaksin ……… 16
2.3.1 Definisi Vaksin ………. 16
2.4 Freezing (Pembekuan) ..………. 28
2.5 Stabilitas Vaksin dalam Aspek Penyimpanan ……… 30
2.6 Immunofluorescence ...……….……….. 32
2.7 Protein Stabilizer, Sukrosa ..……..……… 34
BAB 3 METODOLOGI PENELITIAN ……….. 38
3.1 Tempat dan Waktu Penelitian ...………. 38
3.2 Alat Penelitian ……… 38
3.3 Bahan Penelitian ……… 38
3.4 Prosedur Penelitian ……… 38
3.4.1 Pembuatan Sediaan Kandidat Vaksin Rotavirus ..… 38
3.4.2 Variasi Perlakuan Teknik Freezing dan Suhu Penyimpanan pada Sediaan Kandidat Vaksin …….. 39
3.4.3 Evaluasi Sediaan ……….. 40
3.4.4 Uji Stabilitas Vaksin Kandidat Vaksin Rotavirus Selama Penyimpanan …....……… 41
3.4.5 Metode Analisis Data …....……… 44
3.5 Alur Penelitian ……….……….……… 45
BAB 4 HASIL DAN PEMBAHASAN ……….…. 46
4.1 Formulasi dan Evaluasi Sediaan ..………. 46
4.1.1 Uji Fisik …..……….…. 46
4.1.2 Uji Sterilitas .………..……….…………. 47
4.1.3 Uji Potensi ..……….. 48
4.2 Uji Stabilitas …...……… 49
BAB 5 KESIMPULAN DAN SARAN ……….. 62
5.1 Kesimpulan ………...……… 62
5.2 Saran …...………...……… 62
DAFTAR PUSTAKA ……..……… 64
Halaman
Tabel 2.1 Perbedaan virus dan sel …...……….. 5
Tabel 3.1 Desain perlakuan pada kandidat vaksin rotavirus ..……… 39
Tabel 4.1 Hasil uji fisik ….……… 47
Tabel 4.2 Hasil uji sterilitas …….……….. 48
Tabel 4.3 Hasil uji potensi ….………...………. 48
Tabel 4.4 Desain plate 96 well pada immunofluorescence assay …..…… 51
Tabel 4.5 Konsentrasi rotavirus dalam kandidat formulasi vaksin dengan teknik freezing -70oC …...………..….. 52
Tabel 4.6 Konsentrasi rotavirus dalam kandidat formulasi vaksin dengan teknik freezing -152oC ….……….…. 52
Tabel 4.7 Konsentrasi rotavirus dalam kandidat formulasi vaksin dengan teknik freezing menggunakan nitrogen cair ………... 52
Halaman Gambar 2.1 Penyandian gen dan struktur tiga dimensi dari partikel
rotavirus ………..… 10 Gambar 2.2 Penggambaran skematis protein struktural dari rotavirus …. 11 Gambar 2.3 Vaksin reassortant rotavirus manusia-sapi (RotaTeq)……… 27 Gambar 2.4 Profil penurunan suhu pada pembekuan cepat dan
pembekuan lambat ..……… 29 Gambar 2.5 Visualisasi mikroskop fluorescence ...……… 32 Gambar 2.6 Skema imunofluoresensi langsung dan tidak langsung .…… 33 Gambar 2.7 Struktur kimia sukrosa ...……… 35 Gambar 2.8 SEM (Scanning Electron Microscope) film tipis lapisan
monolayer .………. 36
Gambar 4.1 Hubungan teknik freezing suhu penyimpanan pada sediaan
kandidat formulasi vaksin rotavirus terhadap potensinya ….. 56 Gambar 4.2 Hubungan teknik freezing dan waktu penyimpanan pada sediaan
kandidat formulasi vaksin rotavirus terhadap potensinya ... 56 Gambar 4.3 Hubungan suhu penyimpanan dan waktu penyimpanan pada
sediaan kandidat formulasi vaksin rotavirus terhadap
potensinya …..………..………….. 57 Gambar 4.4 Karakteristik intracellular ice dan extracellular ice pada
Halaman Lampiran 1. Data jumlah sel MA104 yang ber-fluorescence pada tiap
perlakuan teknik freezing dan suhu penyimpanan selama
30, 60, 90, dan 120 hari ………. 69 Lampiran 2. Contoh perhitungan yang digunakan pada penelitian ... 73 Lampiran 3. Data perbandingan hasil pengolahan statistik repeated
measures yang dipisahkan sesuai dengan kelompok teknik
freezing-nya pada sediaan kandidat vaksin rotavirus .……. 75 Lampiran 4. Data perbandingan hasil pengolahan seluruh data
(tanpa pemisahan kelompok) menggunakan analisis statistik repeated measures pada sediaan kandidat vaksin rotavirus .. 80 Lampiran 5. Grafik konsentrasi rotavirus terhadap teknik freezing dan
BAB 1
PENDAHULUAN
1.1 Latar Belakang
Rotavirus merupakan penyebab yang paling sering menimbulkan penyakit gastroenteritis pada anak-anak khususnya pada bayi usia hingga 3 tahun di seluruh dunia. Rotavirus menyebabkan diare dan muntah-muntah yang dapat mengakibatkan kematian pada bayi akibat dehidrasi. Hasil penelitian dunia memaparkan bahwa setiap tahunnya penyakit gastroenteritis yang disebabkan oleh rotavirus di Asia, Afrika, dan Amerika Latin mengakibatkan kurang lebih 500.000 kematian (Cortese et al., 2009). Menurut hasil penelitian, di Indonesia, prevalensi terjadinya diare yang disebabkan oleh rotavirus telah terdeteksi tersebar di beberapa wilayah dengan rentang mulai dari 38% hingga 69% pada bayi dan anak usia di bawah umur lima tahun. Hasil data prevalensi rotavirus di Indonesia selama Januari hingga April 2007 mengalami peningkatan dari hasil investigasi sebelumnya (Radji, et al., 2010). Pada tahun 1990 hingga awal 2000 saat belum dikenalnya vaksin rotavirus, di Amerika Serikat terjadi 2,7 juta kasus gastroenteritis pada bayi umur di bawah 5 tahun, di mana sekitar 410.000 adalah penderita rawat jalan, 205.000-272.000 penderita memasuki Unit Gawat Darurat (UGD), 55.000-70.000 penderita rawat inap, dan 20-60 penderita mengalami kematian (Glass et al., 2009). Biaya tahunan yang dikeluarkan oleh Amerika Serikat dalam menangani permasalahan rotavirus ini bahkan melebihi $ 1 milyar (Kang, 2006).
banyak mengalami mutasi genetik karena dapat mengalami reassortant, yakni proses swapping (penukaran) materi genetik antara dua strain untuk menghasilkan strain virus baru. Selain G1, G2, G3, dan G4, saat ini muncul rotavirus strain G9 yang dapat menginfeksi manusia di dunia, dan muncul pula strain G12 di India dengan frekuensi kemunculan yang meningkat. Hal ini merepresentasikan kemungkinan kemunculan genotip rotavirus yang lain dan menjadi tantangan saat ini dan masa depan, terkait vaksin rotavirus (Angel, et al., 2007).
Dalam proses pengembangan vaksin, aspek penting selain menentukan formulasi yang ideal adalah stabilitas dalam penyimpanannya. Hal ini dikarenakan persoalan cold-chain (upaya menjaga stabilitas suhu dingin yang diperlukan produk untuk tetap berada di rentang tertentu selama proses produksi hingga distribusi) menjadi penting diperhatikan pada sediaan biologis, dalam hal ini vaksin (Huynh-Ba dan Zahn, 2009). Jika cold-chain tidak dapat dipertahankan, vaksin yang poten sekalipun tingkat efikasinya akan hilang (Parthsarthy et al., 2001). Uji stabilitas dengan metode freeze-thaw maupun heat-stress yang dilakukan terhadap produk pun tidak dapat memperkirakan secara pasti kualitas produk khususnya waktu simpan dan suhu penyimpanan karena dapat mengakibatkan kemungkinan lainnya, seperti terjadinya degradasi dalam formulasi zat biologis, sehingga perlu dilakukan penelitian lebih lanjut (Huynh-Ba dan Zahn, 2009).
Dalam hal freezing, tidak hanya suhu pada proses pembekuan saja yang berpengaruh pada komponen aktif vaksin, laju pembekuan (slow freeze dan quick freeze) juga berpengaruh terhadap induksi denaturasi dengan mekanisme yang tidak terkontrol (Kramer et al., 2002). Ketika vaksin mengalami kerusakan akibat pembekuan, potensi dan efikasi yang dimiliki vaksin tersebut tidak dapat dipulihkan, artinya kerusakan tersebut permanen (irreversible) (Dietz et al., 1997). Vaksin yang mengalami kerusakan akibat pembekuan memiliki imunogenisitas yang lebih rendah dan cenderung mengalami reaksi lokal, seperti abses steril (Mansoor, 1997). Selain studi optimasi teknik freezing, studi optimasi suhu penyimpanan vaksin dalam kemasan ampul atau vial juga perlu dilakukan dalam upaya memenuhi stabilitas vaksin pada saat proses distribusi dan proses penyimpanan agar vaksin tersebut dapat mempertahankan viabilitas antigen. Suhu penyimpanan vaksin bermacam-macam dan secara umum dibutuhkan suhu yang rendah untuk kondisi penyimpanannya, seperti pada kulkas (2 hingga 8oC), freezer (-10 hingga -20oC), atau bahkan pada suhu ultra rendah (40 hingga -80oC).
Studi optimasi teknik freezing maupun suhu penyimpanan akan dilakukan pada penelitian ini, mengingat stabilitas sediaan vaksin terkait pengaruhnya terhadap suhu tidak dapat diperkirakan secara pasti dengan metode yang lain. Penelitian ini dilakukan dengan menerapkan prinsip-prinsip quality by design melalui analisis secara statistika agar diketahui kombinasi teknik freezing dan suhu penyimpanan kandidat vaksin rotavirus yang tepat dari desain-desain yang telah diusulkan, dengan mempertimbangkan stabilitas vaksin tersebut agar viabilitas dan imunogenisitasnya dapat dipertahankan.
1.2 Perumusan Masalah
1. Bagaimanakah pengaruh berbagai kombinasi antara teknik freezing dan suhu penyimpanan yang dilakukan pada sediaan kandidat formulasi vaksin rotavirus terhadap kestabilan sediaan vaksin yang diperoleh?
1.3 Tujuan Penelitian
Penelitian ini bertujuan untuk mengamati dan mengevaluasi pengaruh berbagai kombinasi teknik freezing dan suhu penyimpanan pada sediaan akhir kandidat formulasi vaksin rotavirus terhadap kestabilan sediaan, serta dapat menentukan kombinasi teknik freezing dan suhu penyimpanan yang paling optimum untuk diterapkan dalam proses produksi vaksin rotavirus nantinya.
1.4 Manfaat Penelitian
BAB 2
TINJAUAN PUSTAKA
2.1 Virus
Virus adalah makhluk hidup aseluler yang merupakan penyebab infeksi pada manusia, hewan, dan tanaman atau bahkan bakteri. Mekanisme virus dalam menyebabkan penyakit sama seperti sel eukariot dan prokariot lainnya yakni dengan cara menyerang sel yang rentan. Virus tidak memiliki ciri hidup yang sama dengan sel. Perbedaan sifat antara virus dan sel dapat dilihat pada tabel 2.1 (Bauman, 2012).
Tidak membelah diri atau tumbuh Membelah diri dan tumbuh
Aseluler Seluler
Merupakan parasit intraseluler Kebanyakan hidup bebas Mengandung DNA atau RNA dengan
Berukuran ultramikroskopik, antara 10 nm hingga 400 nm
Diameternya 200 nm hingga 12 cm
Memiliki kapsid yang mengandung protein di sekitar genom, dan beberapa memiliki amplop
Dikelilingi oleh membran fosfolipid dan juga dinding sel
Bereplikasi dengan memanfaatkan enzim dan organel sel inang
2.1.1 Karakteristik Virus
Virus merupakan makhluk hidup yang berukuran sangat kecil, aseluler dan bersifat infeksius yang biasanya memiliki satu atau lebih bagian asam nukleat, dapat berupa RNA maupun DNA. Virus tidak memiliki membran sitoplasma, sitosol, dan organ fungsional lainnya. Ukuran virus paling kecil memiliki diameter 10 nm, sedangkan yang terbesar memiliki diameter 400 nm, di mana 400 nm merupakan ukuran terkecil dari sel bakteri. Genom virus dapat berupa DNA untai tunggal, DNA untai ganda, RNA untai tunggal, atau RNA untai ganda (Bauman, 2012).
Virus memiliki status ekstraseluler dan interseluler. Di luar sel, dalam keadaan ekstraseluler, virus disebut dengan virion. Pada dasarnya, virion terdiri dari selubung protein yang disebut dengan kapsid. Kapsid dari virus tersusun atas sub-unit protein yang disebut dengan kapsomer, beberapa kapsomer tersusun dari protein jenis tunggal saja, dan beberapa lainnya tersusun atas beberapa jenis protein yang berbeda. Asam nukleat virus yang dikelilingi oleh kapsidnya disebut dengan nukleokapsid. Lapisan paling luar dari virion berfungsi sebagai pelindung sekaligus pengenal yang akan berikatan dengan senyawa kimia komplemen dari permukaan sel inang spesifiknya. Setelah virus masuk dan berada di dalam, status interseluler pun dimulai, dan kapsid akan dilepaskan. Virus tanpa kapsid akan berwujud sebagai asam nukleat saja, tetapi masih disebut dengan virus (Bauman, 2012).
Semua virus tidak memiliki membran sel, tetapi sebagian virus memiliki envelope yang komposisinya sama dengan membran sel dan terletak mengelilingi nukleokapsid. Virus yang memiliki membran disebut dengan virion bersampul (enveloped virion), sedangkan virion tanpa envelope disebut dengan virion telanjang (non-enveloped virion). Sama seperti membran sitoplasma, envelope virus tersusun atas fosfolipid lapis rangkap dan protein. Protein pada envelope dan glikoprotein seringkali memegang peranan penting dalam mengenali sel inang (Bauman, 2012).
dari virus polihedral membentuk bola dengan struktur yang kasar dan kaku, dengan bentuk yang mirip dengan kubah. Jenis dari kapsid polihedral yang paling umum adalah ikosahedron, dengan 20 sisi yang dimiliki. Sedangkan pada virus komplek, kapsidnya memiliki bentuk yang berbeda yang tidak dapat dimasukkan ke dalam dua kategori lainnya (heliks dan polihedral). Contoh dari virus komplek ini adalah virus cacar, yang memiliki beberapa lapisan penutup (termasuk lemak) dan tidak memiliki kapsid yang mudah diidentifikasi (Bauman, 2012).
Virus dapat bersifat sangat spesifik. Ia menyerang tidak hanya inang tertentu, tetapi bahkan hanya sel tertentu pada inang. Sebagai contoh, HIV (Human Immunodeficiency Virus) secara spesifik menyerang situs pembantu Limfosit-T (helper T-limphocytes) pada manusia, dan tidak menghasilkan efek pada sel otot maupun tulang manusia. Spesifikasinya bergantung pada ketepatan afinitas dari protein atau glikoprotein yang berada di permukaan dari sel inang. Sebaliknya, beberapa virus ada yang bersifat sangat umum, menginfeksi berbagai jenis sel dari berbagai inang yang berbeda. Contoh dari virus general ini adalah virus west nile yang dapat menginfeksi hampir semua spesies dari burung, beberapa mamalia, dan beberapa reptil (Bauman, 2012).
2.1.2 Klasifikasi Virus
ditulis miring, misalnya human immunodeficiency virus dan rabies virus (Bauman, 2012).
2.1.3 Replikasi Virus
Virus tidak dapat bereproduksi sendiri karena virus tidak memiliki gen dari semua enzim yang dibutuhkan untuk replikasi, dan juga tidak memiliki ribosom yang mana sangat berperan dalam sintesis protein. Virus sangat bergantung pada enzim dan organel dari inangnya untuk membentuk virion-virion baru. Begitu suatu inang berada di bawah kontrol dari genom virus, maka inang tersebut akan dipaksa untuk mereplikasi materi genetik dari virus dan menerjemahkan protein virus, termasuk kapsomer dan enzim virus (Bauman, 2012).
1) Siklus replikasi litik
Siklus replikasi dari virus biasanya berakhir pada kematian dan lisis dari sel inang. Karena dalam siklus ini lisis yang dialami oleh sel merupakan penanda dari sudah dekatnya akhir dari siklus, maka tipe replikasi ini disebut dengan siklus replikasi litik. Siklus ini terjadi pada kebanyakan bakteriofag, contohnya adalah bakteriofag T4. Secara umum, siklus ini terdiri atas penempelan dari virion kepada sel inang, pemasukan virion atau genom virus ke dalam sel inang, sintesis dari asam nukleat baru dan protein viral melalui enzim dan ribosom dari sel inang, pembentukan virion baru dalam sel inang, dan terakhir yaitu pelepasan virion baru dari sel inang (Bauman, 2012).
2) Siklus replikasi lisogenik
Beberapa bakteriofag mengalami modifikasi siklus replikasi dimana sel inang yang telah diinfeksi masih dapat tumbuh dan melakukan reproduksi normal sampai beberapa generasi sebelum akhirnya mengalami lisis. Siklus ini disebut dengan siklus replikasi lisogenik, dan fage yang mengalami siklus ini disebut dengan fage lisogenik (Bauman, 2012).
sel inang tidak mengalami kerusakan dan genom dari fage tidak serta merta mengambil alih kontrol dari sel melainkan bertahan sementara dalam bentuk inaktifnya yang disebut dengan profage. Profage bertahan dalam bentuk inaktif dengan mengkode protein yang menekan gen dari profage. DNA virus berfusi dengan DNA dari kromosom sel. Setiap sel mereplikasikan kromosom yang terinfeksi, maka profage akan ikut terreplikasi. Profage akan dikeluarkan dari kromosom melalui proses rekombinasi atau peristiwa genetik lainnya, yang kemudian memasuki fase litik kembali (induksi). Setelah terjadi induksi, akan terjadi tahapan yang sama dengan siklus litik, yakni diawali dengan sintesis, dilanjutkan dengan pembentukan, dan diakhiri dengan pelepasan (Bauman, 2012).
2.2 Rotavirus
2.2.1 Karakteristik Rotavirus
Rotavirus pertama kali ditemukan sekitar tahun 1960an di dalam hewan dan pertama kali diungkapkan terdapat pada manusia ketika ditemukannya virus tersebut pada usus anak-anak yang mengalami gastroenteritis akut dengan
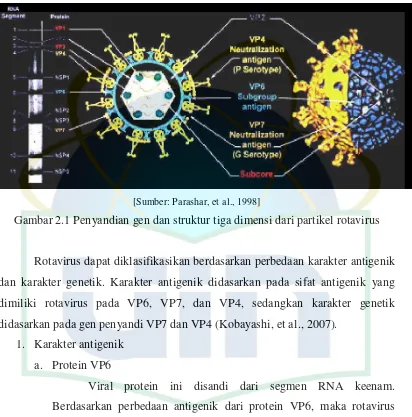
menggunakan mikroskop elektron. Kata “rota” berasal dari bentuknya yang menyerupai roda. Rotavirus merupakan virus dengan karakteristik tiga lapis ikosahedron protein kapsid, yakni lapisan luar (kapsid luar), lapisan tengah (kapsid dalam), dan lapisan inti, dengan diameter 70-nm dan termasuk dalam family Reoviridae (Dennehy, 2008). Lapisan inti dari rotavirus merupakan genom virus yang terdiri dari sebelas segmen RNA untai ganda yang menyandikan enam viral protein struktural (VP1, VP2, VP3, VP4, VP6, VP7) dan lima viral nonstruktural (NSP1, NSP2, NSP3, NSP4, NSP5) (Parashar, et al., 1998). Perkembangan terakhir menyatakan terdapat enam viral protein nonstruktural, dengan tambahannya berupa NSP6 (Ward, et al., 2008).
lapisan inti disusun oleh VP2. Di dalam lapisan inti terdapat VP1 dan VP3 serta sebelas segmen RNA (Clemens, et al., 1999).
[Sumber: Parashar, et al., 1998]
Gambar 2.1 Penyandian gen dan struktur tiga dimensi dari partikel rotavirus
Rotavirus dapat diklasifikasikan berdasarkan perbedaan karakter antigenik dan karakter genetik. Karakter antigenik didasarkan pada sifat antigenik yang dimiliki rotavirus pada VP6, VP7, dan VP4, sedangkan karakter genetik didasarkan pada gen penyandi VP7 dan VP4 (Kobayashi, et al., 2007).
1. Karakter antigenik a. Protein VP6
Viral protein ini disandi dari segmen RNA keenam. Berdasarkan perbedaan antigenik dari protein VP6, maka rotavirus diklasifikasikan menjadi tujuh grup, yaitu rotavirus A, B, C, D, E, F, dan G. Grup A-C merupakan grup yang ditemukan pada manusia dan hewan, sedangkan grup D-G hanya ditemukan terdeteksi pada hewan. Karena rotavirus A merupakan satu-satunya dalam grup ini yang menjadi sumber utama dari etiologi penyakit diare pada anak, maka dalam pembuatan anti-rotavirus vaksin tersebut difokuskan pada grup ini (Kobayashi, et al., 2007).
Klasifikasi rotavirus dari lapisan luar yang terdiri dari dua protein struktural ini didasarkan pada kemampuannya yang dapat menginduksi netralisasi virus (virus-neutralizing) serum antibodi dan mampu menginduksi proteksi sistem imun untuk melawan rotavirus di dalam hewan (Clemens, et al., 1999).
[Sumber: Angel, et al., 2007]
Gambar 2.2 Penggambaran skematis protein struktural dari rotavirus
Protein struktural pertama, VP7, merupakan viral protein yang disandi oleh segmen RNA ke tujuh. Variasi antigenik dari protein ini disebut tipe glikoprotein (tipe G), sesuai dengan susunan glikoprotein dari antigen ini (Clemens, et al., 1999). Saat ini, rotavirus pada manusia dapat memperlihatkan 12 tipe G yang berbeda, dimana lima tipe, yaitu G1, G2, G3, G4, dan G9 merupakan rotavirus tipe G yang paling sering muncul di dalam hasil isolasi dari kasus-kasus penyakit diare (Dennehy, 2008).
penggabungan dari 12 tipe G dengan 15 tipe P dapat menghasilkan 160 kombinasi P-G yang berbeda, namun hanya lima kombinasi yang sering terisolasi dari kasus diare pada manusia, di antaranya P[8]G1, P[4]G2, P[8]G3, P[8]G4, P[8]G9 (Dennehy, 2008).
2. Karakter genetik
Klasifikasi rotavirus menggunakan karakter genetik dilakukan berdasarkan urutan nukleotida gen pengkode VP7 dan VP4. Hal tersebut disebabkan gen pengkode VP7 dan VP4 mempunyai beberapa daerah spesifik genotipe, yaitu daerah yang bervariasi antara satu genotipe dengan genotipe lain, tetapi relatif sama pada satu genotipe yang sama (Gentsch, et al., 1992). Dua galur (strain) memiliki genotipe yang sama bila memiliki kesamaan urutan asam amino pada protein VP7 dan VP4 sebesar 89% atau lebih. Hal tersebut ditetapkan berdasarkan hasil analisis terhadap galur dari serotipe yang sama (Kobayashi, et al., 2007).
Klasifikasi genotipe dan serotipe memiliki korelasi yang erat. Galur dengan suatu genotipe tertentu akan menunjukkan reaksi yang bersesuaian pada reaksi serologinya. Sebanyak 16 tipe gen penyandi VP7 (genotipe G) dan 28 tipe gen penyandi VP4 (genotipe P) rotavirus A telah ditemukan menginfeksi manusia atau hewan (Kobayashi, et al., 2007).
2.2.2 Epidemiologi dan Imunitas
prevalensi rotavirus di Indonesia selama Januari hingga April 2007 mengalami peningkatan dari hasil investigasi sebelumnya (Radji, et al., 2010).
Penyebaran dari penyakit ini tidak sama rata terdistribusi antara negara maju dan negara berkembang. Hal ini mungkin disebabkan faktor sosioekonomi dan epidemiologi. Negara-negara maju mempunyai angka kesakitan yang tinggi namun angka kematiannya rendah, sedangkan kasus kematian mayoritas terjadi pada negara berkembang. Sebagai contoh, di Amerika Serikat terdapat 3 juta kasus per tahun dengan angka kematian kurang dari 100, diperkirakan bahwa rotavirus dihubungkan dengan 4 – 5% dari seluruh pasien anak rawat inap, dan diantara 1 dari 67 pasien anak dan 1 dari 85 pasien anak dengan usia lima tahun menjalani rawat inap di rumah sakit dengan penanganan gastroenteritis yang disebabkan rotavirus. Hasil rata-rata ini tidak dapat disangkal pada tahun 1993 hingga 2002. Dari konteks tersebut, bukan hal yang mengherankan bahwa intervensi cost-effective dari vaksin rotavirus menjadi bahan pertimbangan di Amerika Serikat (Angel, et al., 2007).
Epidemiologi rotavirus sangat kompleks karena fenoma ini terus berubah. Adanya distribusi varietas galur rotavirus manusia yang berbeda berpengaruh pada faktor geografis, misalnya virus P[8]G1 lebih sering dijumpai terdapat di Amerika Utara, Eropa, dan Australia daripada di Amerika Selatan, Asia, dan Afrika, dan merupakan daerah yang paling sering ditemuinya rotavirus selama 30 tahun terakhir. Di beberapa daerah di India, Brazil, dan Afrika, rotavirus P[6]G9, P[6]G5, dan P[6]G8, berturut-turut lebih sering ditemukan daripada di tempat manapun (Cortese, 2009). Satu kemungkinan penjelasan dari hal yang dijabarkan ini adalah adanya reassortment (penyusunan kembali materi genetik) antara rotavirus hewan dan manusia. Adanya peningkatan terkait fakta-fakta transmisi zoonotik dari rotavirus hewan menjadi rotavirus manusia, menyebabkan rotavirus hewan dapat menyebabkan infeksi langsung atau terjadinya penyusunan kembali satu atau lebih segmen genom dari rotavirus hewan yang masuk ke dalam sirkulasi rotavirus manusia (Angel, et al., 2007).
rotavirus juga sangatlah penting. Dari hasil studi-studi epidemiologi diawal tahun 1990 menunjukkan dominasi dari rotavirus strain G1-G4, tetapi setelah itu strain P[8]G9 atau P[6]G9 muncul di seluruh dunia dan rotavirus strain G9 terdeteksi sebanyak 4,1% dari seluruh isolat dalam studi baru-baru ini. Seperti yang telah dilaporkan oleh dunia, di Indonesia distribusi penyebaran genotip rotavirus tipe G dan tipe P pun sama, yakni tipe G1, G2, G3, G4, G9 dan tipe P[4], P[6], P[8], dengan rotavirus tipe G1 dan tipe P[8] adalah strain yang paling sering muncul (Radji, et al., 2010). Strain baru G12, terdeteksi pula di India baru-baru ini dan frekuensi kemunculannya meningkat, dimana hal ini merepresentasikan kemungkinan kemunculan genotip rotavirus yang lain dan menjadi tantangan saat ini dan masa depan, terkait vaksin rotavirus (Angel, et al., 2007).
Infeksi simtomatik merupakan infeksi yang paling sering terjadi pada anak-anak antara umur 6 bulan sampai 2 tahun, dan penularan tampaknya terjadi melalui jalur fekal-oral. Infeksi nosokomial juga sering terjadi. Menjelang umur 3 tahun, 90% anak-anak mempunyai antibodi serum terhadap satu tipe rotavirus atau lebih. Manusia dan hewan dapat terinfeksi sekalipun memiliki antibodi. Reinfeksi rotavirus umum terjadi, di mana bayi dapat mengalami reinfeksi sampai 5x sampai umur 2 tahun. Faktor-faktor imun lokal, misalnya IgA sekretorik atau interferon, penting dalam perlindungan terhadap infeksi rotavirus. Selain itu reinfeksi bila ada antibodi yang beredar dapat menggambarkan adanya serotipe ganda virus. Infeksi asimtomatik sering terjadi pada bayi sebelum berumur 6 bulan, di saat antibodi protektif ibu didapat secara pasif oleh bayi yang baru lahir. Infeksi neonatal semacam itu tidak mencegah reinfeksi. Antibodi rotavirus telah dideteksi dalam susu ibu sampai 9 bulan setelah melahirkan. Imunitas yang didapatkan sewaktu anak-anak ini menyebabkan infeksi rotavirus relatif jarang terjadi pada orang dewasa (Brooks, 2007).
2.2.3 Respon Imun Tubuh
mengemukakan bahwa infeksi akut rotavirus di dalam tubuh dapat menimbulkan respon imun humoral yang terdiri dari antibodi imunoglobulin (Ig)M, IgG, dan IgA, dan respon imun rotavirus-spesifik sel T sitotoksik pada lamina propria jaringan usus (Desselberger, 2000). Mekanisme respon imun pada tubuh manusia secara umum diasumsikan bahwa efektor imunologi tersebut aktif di mukosa usus. Hal ini dikarenakan rotavirus bereplikasi di dalam enterosit usus, sehingga respon imun paling nyata pada tubuh manusia terhadap rotavirus adalah sekresi IgA pada usus. Pada tubuh manusia, titer IgA rotavirus serta IgG rotavirus dilaporkan terdeteksi pada serum dan berkolerasi terhadap perlindungan rotavirus setelah terjadi infeksi alami rotavirus (Ward, et al., 2008).
Protein rotavirus yang memiliki sifat imunogenik terbesar adalah VP6, namun protein ini tidak menstimulasi netralisasi serum antibodi tubuh. Meskipun telah dilaporkan demikian, antibodi IgA dengan protein VP6 secara langsung dapat memberikan perlindungan dengan mekanisme yang belum dapat dipahami secara jelas yang diduga melibatkan inhibisi replikasi intraseluler rotavirus dalam enterosit yang terinfeksi selama antibodi berjalan menuju lumen usus. Sedangkan antibodi yang ditujukan langsung pada protein VP4 atau VP7 dapat merangsang netralisasi virus serum antibodi dan diyakini memberikan perlindungan dengan mekanisme netralisasi klasik. Kedua protein tersebut juga dapat menginduksi respon serotipe, baik serotipe spesifik maupun serotipe silang (Ward, et al., 2008). Studi pada bayi dengan infeksi alami rotavirus sangat penting untuk memahami kekebalan tubuh terhadap rotavirus. Tingkat serum spesifik IgA rotavirus terdeteksi setelah infeksi alami pada anak-anak yang berkorelasi dengan tingkat IgA usus, sehingga beberapa penelitian mengemukakan bahwa tingkat serum IgA memberikan korelasi terhadap respon perlindungan. Setelah terjadi respon antibodi IgA dalam tubuh, perkembangan sistem proteksi antibodi tubuh terhadap rotavirus dilanjutkan dengan respon sel T terhadap rotavirus (Angel, et al., 2007).
yang mensekresikan interferon (IFN)- , bukan interleukin (IL)-13 atau IL-4. Sedangkan pada anak-anak dengan gastroenteritis akibat rotavirus, jumlah sel-sel tersebut sangat rendah atau tidak terdeteksi. Akibatnya, pola sitokin yang disekresikan oleh CD4+ sel T spesifik rotavirus pada anak-anak tidak dapat jelas digambarkan, bisa hanya berupa campuran T-helper1 (Th1) dan Th2 (Angel, et al.,
2007).
Studi terbaru yang membandingkan pola ekspresi gen dalam sel mononuklear darah perifer pada anak dengan penyakit diare akibat infeksi rotavirus dan anak yang sehat menunjukkan bahwa pada anak dengan infeksi rotavirus mengalami peningkatan ekspresi gen yang terlibat dalam diferensiasi, pematangan, aktivasi, dan ketahanan sel B, tetapi terjadi penurunan mRNA untuk gen yang terlibat dalam berbagai tahap perkembangan sel T. Selain itu, studi ini juga menunjukkan bahwa terjadi penurunan jumlah limfosit total dan penurunan proporsi jumlah sel T CD4+ dan CD8+ pada sel mononuklear darah perifer, sehingga diketahui bahwa rotavirus dapat mengubah homeostasis sel T. Dengan demikian dapat diketahui bahwa respon sel T terhadap rotavirus pada anak yang terinfeksi rotavirus memiliki intensitas yang rendah, namun respon sel T terhadap rotavirus lebih kuat pada orang dewasa yang sehat (Angel, et al., 2007).
2.3 Vaksin
2.3.1 Definisi Vaksin
Vaksin adalah bibit penyakit yang sudah dilemahkan atau dimatikan dengan prosedur tertentu, digunakan untuk merangsang pembentukan zat kekebalan tubuh, sehingga tubuh dapat menahan serangan penyakit yang bersangkutan. Sebuah vaksin pada dasarnya terdiri atas organisme atau bagian dari organisme (antigen) penyebab penyakit yang relevan dalam memproduksi antibodi. Antigen tersebut dipresentasikan sedemikian rupa dalam bentuk yang tidak berbahaya bagi manusia atau hewan, namun dapat merangsang respon sistem imun untuk menghasilkan antibodi (Kayne dan Jepson, 2004).
yang akan dapat melindungi seseorang yang telah divaksinasi tersebut terhadap penyakit ketika ia terkena agen infeksi di kemudian hari. Kemampuan vaksin untuk merespon sistem imun tanpa memicu terjadinya penyakit sering kali dikombinasikan dengan ajuvan, yakni zat yang dapat memancing atau menarik sel-sel inflamasi tambahan ke tempat bekerjanya vaksin dan merangsang mereka untuk melepaskan berbagai macam sitokin dalam jumlah yang lebih besar. Selanjutnya sinyal-sinyal kimia tersebut akan merangsang dan mengaktifkan sel makrofag dan limfosit untuk memperoleh tambahan fungsi perlindungan (NPI Guide, 2002).
Karena sifat unik dari virus dan sel-sel patogen lainnya, vaksin idealnya dapat membentuk antibodi yang kuat dan meningkatkan respon sel terhadap agen asing penyebab infeksi tersebut di dalam tubuh ketika dipaparkan. Vaksin yang efektif harus memiliki sifat imunogenisitas yang baik sehingga dapat menginduksi respon imun dan merangsang produksi antibodi yang dapat menghancurkan agen patogen sebelum masuk ke dalam sel dan dapat mendatangkan sel-sel T sitotoksik yang dapat menghancurkan sel-sel yang patogen di dalam tubuh. Respon bersama inilah yang akan melindungi tubuh seseorang dari serangan penyakit (NPI Guide, 2002).
Respon imun yang baik pada vaksin virus harus mencakup efek antibodi pada permukaan epitel. Efek ini dapat diperoleh dari IgA lokal atau IgG dan IgM ekstravaskular setempat. Antibodi pada permukaan epitel akan mampu melindungi badan yang dapat mencegah virus masuk ke sirkulasi tubuh. Respon antibodi terhadap virus dapat ditemukan in vitro dengan tiga mekanisme, yakni menetralkan inefektivitas virus dan melindungi pejamu yang rentan, mengikat komplemen, dan mencegah adherens dan aglutinasi eritrosit oleh beberapa jenis virus (haemaglutination inhibition) (Baratawidjaja dan Rengganis, 2010).
membuktikan bahwa sIga dapat mencegah virus masuk secara lokal di tempat. Pada IgG dan IgM, keduanya dapat ditemukan pada sekresi setempat. IgG dan IgM telah ditemukan pula dalam eksudat. Hal ini menandakan bahwa Ig dapat bekerja secara eksta-vaskuler (Baratawidjaja dan Rengganis, 2010).
2.3.2 Jenis-jenis Vaksin
Tiap vaksin memiliki keunikan dalam hal komposisi dan formulasi. Perbedaan ini mencerminkan bahwa tidak hanya agen infeksi dalam vaksin yang berbeda, melainkan juga bagaimana vaksin tersebut digunakan dan melalui mekanisme apa vaksin tersebut dapat bekerja menghasilkan suatu efek (NPI Guide, 2002).
Berdasarkan Farmakope Indonesia edisi IV, jenis-jenis vaksin dibagi menjadi 3 kategori, di antaranya:
1. Vaksin bakteri, yakni vaksin yang dibuat dari biakan galur bakteri yang sesuai dalam media cair atau padat yang sesuai dan mengandung bakteri hidup atau inaktif atau komponen imunogeniknya.
2. Vaksin toksoid bakteri, yakni vaksin yang diperoleh dari toksin yang telah dikurangi atau dihilangkan sifat toksisitasnya hingga mencapai tingkat tidak terdeteksi, tanpa mengurangi sifat imunogenisitasnya.
3. Vaksin virus dan riketsia, yakni vaksin yang berasal dari suspensi virus atau riketsia yang ditumbuhkan dalam telur berembrio, dalam biakan sel atau dalam jaringan yang sesuai, atau dapat berupa virus atau riketsia hidup atau inaktif atau komponen imunogeniknya, di mana dalam vaksin virus hidup umumnya dibuat dari virus galur khas yang virulensinya telah dilemahkan.
Jenis-jenis vaksin yang digunakan manusia saat ini juga dapat dibagi menjadi tiga kelompok utama, yaitu:
1. Vaksin Live-Attenuated
infeksi tersebut telah kehilangan salah satu atau lebih sifat yang berkontribusi terhadap patogenisitasnya yang menyebabkan mereka tidak mampu menimbulkan penyakit pada orang yang sehat. Proses pelemahan ini dapat diperoleh dengan menumbuhkan suatu zat patogen di bawah kondisi tumbuh yang abnormal. Mutan yang dihasilkan melalui proses ini kemudian diuji kehilangan daya virulensinya namun harus tetap dapat mempertahankan sifat imunogenisitasnya. Vaksin tuberkulosis (TBC), campak, gondok, rubella, dan polio adalah beberapa contoh dari vaksin live-attenuated ini (McMullan, 2009).
2. Vaksin Inactivated
Vaksin inactivated memanfaatkan proses inaktivasi kimia zat patogen
dengan menggunakan formalin atau -propiolakton atau ekstraksi menggunakan detergen nonionik seperti Triton X-100. Jenis vaksin ini relatif tidak memerlukan proses pembuatan yang rumit dan biaya yang dikeluarkan lebih murah. Vaksin dengan menggunakan pendekatan ini harus dapat memastikan sifat imunogenik dari agen infeksi tersebut dapat dipertahankan. Komponen agen infeksi dalam vaksin ini harus tidak dapat bereproduksi, tidak dapat menimbulkan penyakit, dan biasanya diberikan dalam dosis ganda untuk memperoleh kekebalan sistem imun. Vaksin ini perlu diperhatikan setelah proses inaktivasi selesai, karena resiko terjadinya pembalikkan virulensi masih memungkinkan terjadi meskipun kecil (McMullan, 2009). Contoh vaksin inactivated di antaranya adalah vaksin influenza, rabies, hepatitis A dan B, pertusis, serta tetanus (NPI Guide, 2002).
3. Vaksin Subunit
muncul karena adanya beberapa faktor yang mempengaruhi bahkan mengharuskan digunakannya vaksin subunit dibandingkan dengan vaksin organisme utuh, misalnya pada virus hepatitis B di mana sampai saat ini masih merupakan kemustahilan dan ketidakmampuan untuk melemahkan virus tersebut. Selain itu vaksin subunit juga dapat dibuat menggunakan racun yang diinaktivasi, sebagai contoh yaitu tetanus toksin yang dihasilkan dari Clostridium tetani, di mana eksotoksin bakteri inilah yang merupakan faktor virulensi, bukan bakterinya (McMullan, 2009).
2.3.3 Komponen dan Bahan Eksipien Vaksin
Di dalam formulasi dan proses pembuatan vaksin, terdapat beberapa komponen dan eksipien yang sering digunakan dan terkandung di dalam vaksin, di antaranya yaitu (NCIRS, 2009):
1. Komponen aktif
Komponen aktif vaksin sering dikenal sebagai ‘antigen’ vaksin yang
dapat menginduksi terjadinya respon imun. Komponen ini merupakan bentuk modifikasi atau bentuk sebagian dari virus, bakteri, atau toksin yang dapat menyebabkan penyakit, sesuai dengan spesifikasi vaksin tersebut. Antigen vaksin ini diubah dari bentuk aslinya sehingga tidak lagi dapat menyebabkan penyakit namun harus tetap dapat menghasilkan respon imun yang sesuai. Ada sejumlah cara untuk mendapatkan komponen aktif ini, yakni menggunakan virus hidup yang dilemahkan (live-attenuated), virus inaktif (inactivated), menggunakan suatu bagian dari virus atau bakteri (misalnya lapisan terluar polisakarida dari suatu virus atau bakteri), atau dengan menggunakan toksin yang dihasilkan oleh suatu bakteri.
2. Adjuvan
Penggunaan aluminium adjuvan dalam vaksin umumnya berarti hanya sedikit kandungan antigen per dosis vaksin, di mana dalam kasus tertentu, dosis antigen lebih sedikit diperlukan. Keberadaan adjuvan dalam vaksin sering dikaitkan dengan reaksi lokal yang terjadi di tempat injeksi setelah dilakukan vaksinasi.
3. Pelarut (Diluent)
Pelarut merupakan cairan yang diberikan atau disiapkan secara terpisah dan digunakan untuk melarutkan vaksin dengan konsentrasi yang tepat sebelum pemberian. Pelarut yang biasanya digunakan adalah saline steril atau air steril.
4. Stabilisator
Stabilisator merupakan komponen tambahan yang digunakan untuk membantu menjaga efektivitas suatu vaksin dengan menjaga antigen dan komponen vaksin lainnya tetap stabil selama proses pembuatan dan penyimpanan. Stabilisator mampu mencegah komponen vaksin menempel pada sisi vial vaksin. Contoh stabilisator adalah laktosa dan sukrosa, glisin dan monosodium glutamat (keduanya merupakan asam amino atau garam-garam dari asam amino), dan serum albumin manusia atau sapi. Gelatin, di mana merupakan hasil hidrolisis sebagian dari kolagen, biasanya berasal dari sapi atau babi, dapat ditambahkan ke dalam vaksin sebagai stabilisator.
5. Preservatif
Preservatif atau pengawet digunakan untuk mencegah terjadinya kontaminasi jamur dan / atau bakteri dalam vaksin. Pengawet terdapat pada beberapa vaksin, tapi tidak semua vaksin. Awalnya, pengawet digunakan untuk mencegah kontaminasi bakteri dari kemasan dosis ganda, namun saat ini kemasan dosis ganda sudah jarang bahkan tidak lagi digunakan. Pengawet yang digunakan di antaranya tiomersal, fenoksietanol, dan fenol.
6. Komponen Jejak
sel, protein telur, ragi, antibiotik, atau agen penginaktif. Biasanya, hanya sedikit sekali zat ini terdeteksi dalam produk akhir sediaan vaksin.
2.3.4 Proses Produksi Live-Virus Vaksin
Dalam memproduksi live-virus vaksin, terdapat beberapa unsur signifikan yang harus dilakukan. Pada umumnya, proses manufaktur vaksin ini dibagi menjadi dua proses utama yaitu proses produksi bulk vaksin dari pemanenan virus tunggal (single virus harvest) dan proses produksi produk obat, dalam hal ini adalah vaksin. Pemanenan virus tunggal dimaksudkan untuk memisahkan virus yang akan digunakan dalam pembuatan vaksin dan virus strain lain yang mungkin hidup bersama dalam suatu kultur induk. Setiap pemanenan virus tunggal, perlu dilakukan uji untuk kontrol setiap batch sel virus yang dihasilkan. Masing-masing virus tunggal diuji secara individu dan disimpan beku pada temperatur yang rendah sampai terformulasi menjadi produk akhir. Stabilitas data dari bulk vaksin biasanya memungkinkan dapat disimpan 18 hingga 36 bulan sehingga memudahkan proses produksi menjadi produk akhir, uji klinis, dan distribusi komersial (PATH, 2006).
Dalam proses kultur virus tunggal, setidaknya terdapat beberapa tahap utama sebelum akhirnya diperoleh bulk vaksin, yaitu (PATH, 2006):
1. Resusitasi sel
Dalam proses resusitasi biasanya dibutuhkan medium (contoh DMEM atau Dulbecco's modified Eagle's medium), serum (contoh BCS atau Bovine Calf Serum), NaHCO3, dan antibiotik. Semua bahan tersebut dicampurkan
dengan takaran tertentu dan didistribusikan ke dalam T-flasks. Kemudian sel beku yang digunakan untuk kultur dicairkan, dan dimasukkan ke dalam T-flasks yang berisi medium. Kemudian dilakukan observasi sel dengan menggunakan mikroskop untuk memastikan keadaan sel dan dilakukan inkubasi pada temperatur 37oC (Freshney, 2005).
2. Pasase sel
Pasase adalah proses subkultur sel. Pasase dilakukan ketika sel sudah mencapai kondisi konfluen (kondisi meratanya sel sebagai sel monolayer sampai menutupi wadah kultur). Pada proses pasase, medium pertumbuhan dikeluarkan dari T-flasks, kemudian sel dicuci dengan menggunakan PBS (phosphate buffer saline) tanpa Ca2+ dan Mg2+. PBS akan menghilangkan serum yang terkandung di dalam medium. Serum harus dihilangkan agar tidak menganggu kerja tripsin. Setelah sel dicuci dengan PBS, ditambahkan tripsin untuk melepas sel dari permukaan T-flasks. Tripsin memiliki aktivitas proteolitik yang dapat menyebabkan kerusakan protein pada membran sel. Inkubasi suspensi tripsinisasi dilakukan selama 5-8 menit. Waktu inkubasi yang terlalu lama dapat menyebabkan kerusakan membran sel akibat degradasi membran protein oleh tripsin (Hsiang et al, 2010).
mencegah pertumbuhan mikroba. Antibiotik yang diberikan ada dalam jumlah sedikit karena bersifat sitotoksik. Setelah dilakukan penambahan medium baru, sel tersebut diinkubasi kembali pada suhu 37oC (Freshney, 2005).
3. Inokulasi dan kultivasi virus
Kultivasi virus dilakukan setelah dilakukan pergantian medium kultivasi. Sebelumnya sel dicuci kemudian virus dimasukkan ke dalam kultur agar dapat menginfeksi sel. Infeksi sel ini akan mengakibatkan CPE (cytopathic effect). CPE ditandai dengan adanya perubahan bentuk sel, pelepasan sel dari subtrat, lisis, dan apoptosis. Khusus untuk sel vero yang diinfeksi oleh virus, sel akan terlepas dari substrat (Freshney, 2005).
4. Pemanenan virus yang menginfeksi kultur sel
Setelah proses inokulasi dan penginfeksian virus pada sel kultur, dapat dilakukan pemanenan virus dengan cara menghilangkan medium, mencuci sel terinfeksi tersebut, kemudian dilakukan tripsinasi dengan tripsin atau EDTA. Didiamkan beberapa saat untuk memberikan waktu kerja dari proses tripsinasi. Terakhir dilakukan pengendapan dan pencucian sel dengan resuspensi dan sentrifugasi (Freshney, 2005).
5. Proses downstream (purifikasi dan karakterisasi)
Ada banyak faktor yang harus dipertimbangkan dalam mengembangan strategi pengolahan hilir (downstream), dengan pemahaman bahwa setiap langkah pengolahan yang termasuk di dalamnya mengakibatkan hilangnya viabilitas virus. Tujuan pengolahan hilir di antaranya harus dapat menghilangkan puing-puing sel pada proses pemanenan, mengurangi atau menghilangkan impurities, menghasilkan konsentrat virus, menambahkan virus stabilizer, mensterilkan virus, dan meningkatkan potensi agen adventif (PATH, 2006).
titer virus dalam pemanenan terlalu rendah untuk diformulasikan menjadi produk akhir atau untuk menghilangkan air yang dapat mengurangi volume penyimpanan. Konsentrat tersebut dapat diperoleh dengan ultrafiltrasi atau ultrasentrifugasi. Larutan virus stabilizer yang biasanya mengandung sukrosa ditambahkan untuk membantu mempertahankan kestabilan virus selama proses penyimpanan -60oC sebagai konsentrat vaksin monovalen. Terakhir, preparasi vaksin harus dilakukan sterilisasi menggunakan filtrasi (PATH, 2006).
Dalam hal karakterisasi, ada enam persyaratan utama yang perlu dilakukan dan diperhatikan, yaitu demonstrasi dalam hal tidak adanya kontaminasi silang, konfirmasi spesies atau strain, korelasi dengan jaringan asal, penentuan apakah virus mengalami perubahan atau transformasi, indikasi apakah virus rentan terhadap ketidakstabilan genetik dan variasi fenotipik, terakhir adalah mengidentifikasi virus tertentu dalam kelompok yang sama (strain yang bebeda) (Freshney, 2005).
Setelah serangkaian proses produksi virus tunggal tersebut selesai, dilakukan proses pembekuan sehingga diperoleh bulk vaksin monovalen beku yang stabil agar dapat disimpan sebelum diformulasikan menjadi produk akhir. Proses manufaktur vaksin dilanjutkan dengan proses produksi produk akhir dimulai dengan mencairkan sejumlah bulk vaksin beku. Kemudian dicampurkan konsentrat vaksin tersebut ke dalam mixing vessel. Konsentrat vaksin kemudian dilarutkan dengan pelarut yang sesuai dan ditambahkan dengan virus stabilizer dan eksipien lain sesuai dengan kekuatan dan potensi yang diinginkan. Formula vaksin tersebut kemudian diisi sesuai dengan dosis akhir kemasan. Dilakukan lypophilize vial apabila proses ini dibutuhkan. Setelah itu sediaan vaksin diberi label dan dievaluasi. Sediaan akhir vaksin disimpan pada temperatur yang sesuai untuk menjaga viabilitas virus (PATH, 2006).
2.3.5 Vaksin Rotavirus
vaksin rotavirus ini difokuskan terutama pada pengiriman strain rotavirus hidup yang dilemahkan melalui rute oral. Uji coba vaksin rotavirus pertama menggunakan rotavirus strain sapi RIT4237 (P[1]G6), menghasilkan suatu keberhasilan sebesar 55-62% terhadap diare rotavirus dan tingkat efikasi sebesar 80-88% terhadap penyakit diare yang lebih parah. Meskipun ini merupakan awal yang menggembirakan, baik RIT4237 dan jenis vaksin sapi lain, WC3 (P[5]G6), memiliki hasil yang tidak konsisten. Ketika diuji di negara berkembang, khasiat yang rendah diamati dalam uji coba yang dilakukan di negara-negara berkembang termasuk Gambia, Peru, Rwanda, dan Republik Afrika Tengah. Sebuah vaksin rhesus strain (RRV), P[3]G3, diuji secara luas, termasuk di negara yang relatif miskin, Venezuela, di mana didapatkan hasil bahwa vaksin ini memiliki tingkat efikasi sebesar 85-90% terhadap penyakit diare yang paling parah (Kang, 2006).
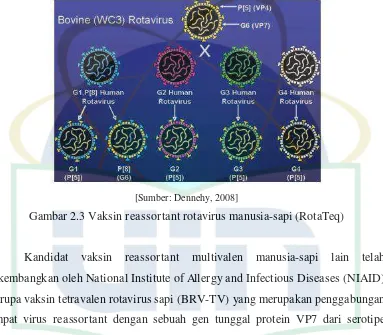
Secara umum, jenis vaksin rotavirus dapat dibagi menjadi dua kelompok, yaitu vaksin berbasis rotavirus hewan dan vaksin berbasis rotavirus manusia. Saat ini, pada vaksin berbasis rotavirus hewan tidak hanya berbentuk vaksin monovalen rotavirus hewan saja, tetapi juga sudah berkembang vaksin multivalen. Vaksin ini berkembang setelah diketahuinya kemampuan rotavirus untuk reassort selama infeksi campuran secara in vitro yang memungkinkan diproduksinya vaksin reassortant yang diistilahkan sebagai pendekatan “Jennerian”. Virus reassortant mengandung beberapa gen dari induk rotavirus hewan dan beberapa gen dari induk rotavirus manusia. VP7 dianggap penting dalam memberikan perlindungan, oleh karena itu reassortant rotavirus yang digunakan sebagai vaksin menyisipkan gen VP7 manusia untuk memberikan respon imun protektif (Dennehy, 2008).
strain induk rotavirus sapi WC3 (P[5]G6) dan satu reassortant rotavirus yang mengekspresikan protein tambahan VP4 (P[8]) dari strain induk rotavirus manusia dan kapsid terluar protein VP7 (G6) dari strain induk rotavirus sapi (Dennehy, 2008).
[Sumber: Dennehy, 2008]
Gambar 2.3 Vaksin reassortant rotavirus manusia-sapi (RotaTeq)
Kandidat vaksin reassortant multivalen manusia-sapi lain telah dikembangkan oleh National Institute of Allergy and Infectious Diseases (NIAID) berupa vaksin tetravalen rotavirus sapi (BRV-TV) yang merupakan penggabungan empat virus reassortant dengan sebuah gen tunggal protein VP7 dari serotipe manusia G1, G2, G3, atau G4 dan gen dari strain rotavirus sapi Inggris (P[7]G6). Dengan munculnya serotipe G9 dan G8 sebagai serotipe epidemiologis penting di berbagai daerah, pengembang vaksin di NIAID berencana untuk menambah reassortant manusia-sapi dengan spesifikasi G8 dan G9 pada vaksin tetravalen tersebut, sehingga menghasilkan formula vaksin heksavalen (Dennehy, 2008).
rekannya di Australia, telah menjadi kandidat vaksin berbasis rotavirus manusia sebagai vaksin oral pada bayi usia 3 bulan yang aman dan dapat ditoleransi dengan baik. Saat ini, kandidat vaksin RV3 juga dikembangkan oleh Biofarma, Indonesia (Dennehy, 2008). Kandidat vaksin berbasis rotavirus manusia lainnya juga dikembangkan oleh Bharat Biotech dengan menggunakan dua strain Indian neonatal di antaranya, 116E (P[11]G9) yang merupakan rotavirus reassortant alami berupa strain manusia dengan gen tunggal VP4 (P[11]) sapi dan I321 yang merupakan rotavirus strain reassortant manusia-sapi yang terdiri dari gen utama yang berasal dari sapi dan hanya memiliki dua segmen gen saja yang berasal dari manusia (Kang, 2006).
2.4 Freezing (Pembekuan)
Pembekuan adalah proses pemindahan panas dari bahan yang disertai dengan perubahan fase dari cair ke padat (Tambunan, 1999). Syarief dan Kumendong (1992) menyatakan bahwa pembekuan adalah kegiatan menurunkan suhu bahan pangan atau produk di bawah suhu titik bekunya dan sejumlah air berubah menjadi kristal es. Tahap pembekuan merupakan tahap penting dalam proses lyopilisasi karena efek potensial yang dihasilkannya terhadap terjadinya agregasi protein (Willemer, 1992; Searles, 2001).
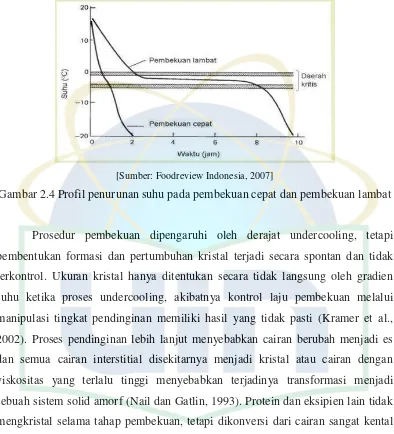
pembekuan merupakan salah satu faktor kritis yang menentukan mutu produk beku yang dihasilkan (Foodreview Indonesia, 2007).
[Sumber: Foodreview Indonesia, 2007]
Gambar 2.4 Profil penurunan suhu pada pembekuan cepat dan pembekuan lambat
Prosedur pembekuan dipengaruhi oleh derajat undercooling, tetapi pembentukan formasi dan pertumbuhan kristal terjadi secara spontan dan tidak terkontrol. Ukuran kristal hanya ditentukan secara tidak langsung oleh gradien suhu ketika proses undercooling, akibatnya kontrol laju pembekuan melalui manipulasi tingkat pendinginan memiliki hasil yang tidak pasti (Kramer et al., 2002). Proses pendinginan lebih lanjut menyebabkan cairan berubah menjadi es dan semua cairan interstitial disekitarnya menjadi kristal atau cairan dengan viskositas yang terlalu tinggi menyebabkan terjadinya transformasi menjadi sebuah sistem solid amorf (Nail dan Gatlin, 1993). Protein dan eksipien lain tidak mengkristal selama tahap pembekuan, tetapi dikonversi dari cairan sangat kental menjadi amorf solid yang kaku (seperti kaca) yang berisi sekitar 10-30% air (Pikal, 1990; Hatley et al., 1996).
parameter yaitu laju pendinginan (berdampak pada ukuran kristal yang dihasilkan) dan suhu akhir pembekuan berpengaruh dalam induksi denaturasi protein secara kompleks (Franks, 1990; Wang, 2000). Dampak dari tingkat pendinginan pada stabilitas protein bervariasi secara substansial. Laju pembekuan juga mempengaruhi stabilitas protein ditinjau dari mekanisme denaturasi protein (Wang, 2000). Pembekuan lambat (misal diproduksi dengan shelf freezing) mengarah kepada pertumbuhan kristal es dengan ukuran yang besar dan memiliki kecenderungan terhadap zat terlarut yang terkonsentrasi secara maksimal. Dalam keadaan ini, inkompatibilitas antara zat terlarut dapat menyebabkan pemisahan fase (Heller, et al., 1999).
Pembekuan cepat dapat menyebabkan peningkatan agregat tidak larut dari rGH (recombinant Growth Hormone) (Eckhardt et al., 1991) dan pembentukan formasi berbagai macam agregat dari IgG sapi dan manusia (Sarciaux et al., 1998). Pada teknologi pangan, proses pembekuan lambat yang menghasilkan kristal es dengan ukuran besar, berpeluang untuk menusuk dan merusak sel jaringan pangan, sehingga menyebabkan sel kehilangan air dan kehilangan keteguhan tekstur (Foodreview Indonesia, 2007). Pada pembekuan cepat menunjukkan terjadinya sedikit kehilangan aktivitas LDH (Lactat Dehydrogenase Hormone) (Nema dan Avis, 1993) dan terjadi sedikit perubahan dalam struktur sekunder hemoglobin dalam larutan dekstran/PEG (Heller et al, 1999). Alasannya mungkin disebabkan karena pembekuan cepat mencegah pertumbuhan kristal secara luas, sehingga dapat mengurangi konsentrasi larutan yang menginduksi denaturasi protein (Wang, 2000).
2.5 Stabilitas Vaksin dalam Aspek Penyimpanan
hingga - 20⁰C), atau bahkan pada suhu penyimpanan ultra rendah (-40⁰ hingga -80⁰C). Hal ini menjadi ketentuan standar ruangan stabilitas suhu rendah. Aspek lainnya yang perlu diperhatikan adalah produk biologi seperti vaksin umumnya sangat mahal dan seringkali memakan waktu yang cukup banyak untuk diproduksi dan beberapa hanya dapat diproduksi dalam batch dengan jumlah yang sedikit bergantung pada sifat dasar dari teknologi yang digunakan (Huynh-Ba dan Zahn, 2009). Dalam proses produksi skala besar, bahan aktif vaksin yang diperoleh setelah proses produksi bahan aktif (API atau Production of Active Ingredients) biasanya disimpan sebelum diformulasikan menjadi vaksin (PATH, 2006).
Vaksin, seperti pula produk biologi lainnya, membutuhkan penanganan ekstra ketika dikaitkan dengan stabilitas farmasetikanya. Aktivitas biologi dari protein misalnya, yang mana merupakan penyusun utama dari vaksin, berasal tidak hanya dari struktrur ikatan kovalen primernya tetapi juga dari konformasi lipatan yang menghasilkan struktur sekunder dan tersier. Konformasi ini dapat dengan mudah berubah tanpa harus memecah ikatan kovalennya, dan ketika hal ini menyebabkan denaturasi maka seluruh atau sebagian aktivitas biologis yang membuat protein atau vaksin memiliki aktivitas dapat hilang. Untuk itulah, perlu ditangani dengan sangat hati-hati (Huynh-Ba dan Zahn, 2009).
Metode freeze-thaw atau heat-stress biasanya diterapkan untuk membantu memperkirakan proses penyimpanan dan mencegah terjadinya penyimpangan pengiriman. Akan tetapi, meskipun sudah diterapkan, kualitas produk khususnya waktu simpan produk tetap tidak dapat dipastikan begitu saja. Karena baik metode freeze-thaw maupun heat-stress yang dilakukan terhadap produk pun dapat mengakibatkan kemungkinan lainnya, seperti terjadinya degradasi dalam formulasi zat biologis (Huynh-Ba dan Zahn, 2009).
2.6 Immunofluorescence
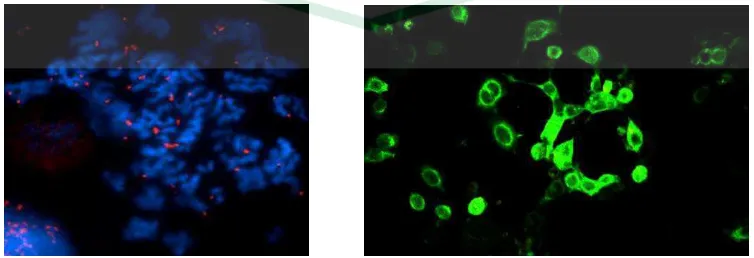
Imunofluoresensi (Immunofluorescence atau IF) adalah teknik laboratorium yang umum digunakan pada hampir seluruh aspek biologi. Teknik ini telah banyak digunakan baik dalam penelitian dan diagnostik klinis, misalnya evaluasi suspensi sel, kultur sel, jaringan, dan microarray untuk mendeteksi protein tertentu (Coons, 1941; Coons dan Kaplan, 1950). Dalam teknik IF, antibodi secara kimiawi terkonjugasi oleh pewarna fluorescent seperti fluorescein isothiocyanate (FItC) atau tetramethyl rhodamine isothiocyanate (tRItC). Antibodi yang terikat (baik secara langsung maupun tidak langsung) dengan antigen yang sesuai dapat ditandai dengan pewarna fluorescent sehingga deteksi antigen dapat dilakukan dan dapat diukur fluoresensi tersebut dengan menggunakan flow cytometer, array scanner (automated imaging instrument), atau divisualisasikan menggunakan mikroskop fluorescence (Robinson, et al., 2009).
Dua metode utama pelabelan immunofluorescent adalah langsung (direct) dan tidak langsung (indirect). Pada imunofluoresensi langsung, antibodi yang terikat pada molekul yang cocok (antigen) secara kimiawi terkonjugasi dan ditandai oleh pewarna fluorescent. Sedangkan pada imunofluoresensi tidak langsung, bukan antibodi spesifik yang terikat pada antigen (antibodi primer) yang ditandai pewarna fluorescent, melainkan antibodi anti-immunoglobulin (antibodi sekunder) yang menempel pada antibodi primerlah yang ditandai dengan pewarna fluorescent (Gambar 2.6) (Robinson, et al., 2009).
[Sumber: Robinson, et al., 2009]
Gambar 2.6 Skema imunofluoresensi langsung dan tidak langsung
baik dalam penempelan dengan antigen maupun dengan antibodi sekunder (Robinson, et al., 2009).
Secara luas, metode fluoresensi tidak langsung lebih banyak digunakan dibandingkan imunofluoresensi langsung. Aplikasi imunofluoresensi langsung kebanyakan digunakan pada kasus-kasus tertentu, misalnya digunakan untuk lokalisasi IgG dalam kompleks imun pada biopsi kulit bagian dermal-epidermal junction dari pasien yang menderita lupus eritematosus sistemik (Jennet dan Wick, 1988).
2.7 Protein Stabilizer
Osmolit memiliki kemampuan untuk melindungi komponen seluler terhadap terjadinya denaturasi akibat pengaruh lingkungan. Kemampuan dari suatu bahan osmolit tersebut untuk melindungi protein merupakan pengetahuan yang berharga untuk mengatasi permasalahan stabilitas protein, yang salah satunya berkaitan dengan bioteknologi dan biofarmasetika. Salah satu golongan osmolit alami yang dapat berfungsi sebagai pelindung protein adalah golongan gula atau poliol, seperti sukrosa, trehalosa, sorbitol, dll (Bolen, D.W, 2001).
Gula atau poliol banyak digunakan sebagai protein stabilizer di dalam larutan atau selama proses freeze-thawing dan freeze-drying. Keduanya baik digunakan sebagai cryoprotektan dan lyoprotektan. Tingkat dari stabilisasi yang dimiliki oleh gula dan poliol secara umum bergantung pada konsentrasinya (Street, 2006). Dari hasil penelitian Tanaka et al (1991), sakarida dapat memproteksi protein dengan langsung berinteraksi dengan protein dan konsentrasi dari sakarida yang cukup dapat membentuk lapisan monomolekular pada permukaan protein untuk mencapai stabilitas dari protein. Namun, peningkatan konsentrasi gula atau poliol sampai melewati batas atau limit stabilisasi akan menyebabkan terjadinya destabilisasi protein selama freeze-drying.
dan trehalosa merupakan eksipien yang sering digunakan. (Ward et al., 1999; Change et al., 1996).
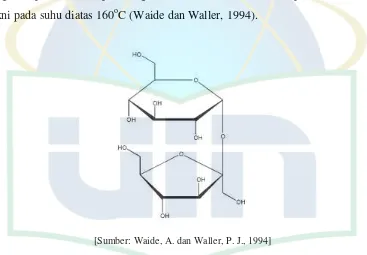
Sukrosa ( -D-fructofuranosyl-α-D-glucopyranoside) berbentuk kristal tidak berwarna, serbuk putih kristalin, tidak berbau, memiliki rasa yang manis, dan biasa digunakan dalam membuat gula-gula (manisan); coating agent; bahan granulasi; suspending agent; pemanis; pengikat pada tablet; pengisi pada tablet dan kapsul; therapeutic agent; viscosity-increasing agent. Sukrosa larut dalam air, sukar larut dalam ethanol, dan praktis tidak larut dalam kloroform. Sukrosa memiliki kestabilan yang baik pada suhu ruang dan memiliki tingkat kelembaban yang cukup. Sukrosa dapat mengalami karamelisasi bila mencapai titik didihnya, yakni pada suhu diatas 160oC (Waide dan Waller, 1994).
[Sumber: Waide, A. dan Waller, P. J., 1994] Gambar 2.7 Struktur kimia sukrosa
konformasi relaksasi dari protein (Hagen et al., 1995, 1996). Bahan amorf disini secara struktural lebih mirip bentuk liquid dibandingkan dengan bentuk kristal. Kaca dalam mekanisme ini dapat dibagi menjadi dua tipe, yaitu mudah pecah (fragile) dan kuat (strong). Sukrosa memiliki bentuk kaca tipe mudah pecah, dimana pada tipe ini, viskositas akan meningkat lebih mendalam dari pada kaca tipe kuat bila diberikan temperatur dibawah temperatur transisi kaca. Dengan alasan demikian, maka eksipien yang membentuk kaca tipe mudah pecah ini memiliki kemampuan sebagai penstabil protein yang lebih baik (Angell, 1995).
[Sumber: Predoi, 2010]
Gambar 2.8 Scanning Electron Microscope film tipis lapisan monolayer sukrosa
Selain itu, mekanisme sukrosa lainnya dikenal dengan hipotesis penggantian air, dimana mekanisme ini melibatkan formasi dari ikatan hidrogen antara protein dengan sukrosa pada saat akhir proses pengeringan untuk memenuhi ikatan hidrogen gugus polar pada permukaan protein sebagai pengganti dari air yang hilang. Mekanisme ini terjadi pada sediaan solid (Crowe et al., 1993; Allison et al., 1998). Untuk menjaga kestabilan protein dalam bentuk solid agar dapat disimpan dalam jangka waktu yang lama, sukrosa dapat menghambat terjadinya agregasi secara signifikan dan menghambat terjadinya degradasi kimia karena dapat mengurangi deamidasi yang terjadi (Tanaka et al., 1991).
folding, unfolded (U) ↔ native (N), osmolit yang memiliki sifat proteksi
BAB 3
METODOLOGI PENELITIAN
3.1 Tempat dan Waktu Penelitian
Penelitian dilakukan pada bulan Januari 2013 di Laboratorium Research and Development PT Biofarma, Bandung, Indonesia. Penelitian ini dilakukan kurang lebih selama 5 bulan terhitung dari bulan Januari 2013.
3.2 Alat Penelitian
Vial, Laminar Air Flow (LAF) cabinet, inkubator, inkubator CO2, kulkas,
freezer, ultra-low freezer, syringe filter (Filter Acrodisc), plate 96 well, mikroskop, mikroskop inverted, mikroskop fluorescence, pipet mikro, pipet mikro multi-channel, pipet tip, magnetic stirrer, tanki nitrogen cair, hemocytometer, T-flasks, ice bath, dan peralatan-peralatan gelas yang umum digunakan di laboratorium.
3.3 Bahan Penelitian
Bulk vaksin rotavirus beku, sukrosa steril, nitrogen cair, FTM (Fluid Thioglycolate Medium) steril dan SCDM (Soybean Casein Digest Medium) steril yang diperoleh dari bagian media PT Biofarma, sel MA 104 (ginjal monyet), media pertumbuhan sel (DMEM (Dulbecco’s Modified Eagle’s Medium), FBS (Fetal Bovine Serum), antibiotik, dan NaHCO3), aseton 80%, alkohol 70%,
tripsin, trypan blue, antibodi poliklonal rabbit anti SA 11, antibodi IgG goat anti rabbit, dan PBS (Phosphate Buffered Saline).
3.4 Prosedur Penelitian
3.4.1 Pembuatan Sediaan Kandidat Vaksin Rotavirus