SELEKS
PROTEA
D
SI BAKTE
ASE UNT
DAN KINE
SE
INS
ERI PROT
TUK MEN
ERJA PER
TITIN
EKOLAH
STITUT P
TEOLITIK
NINGKAT
RTUMBU
N KURNIA
H PASCAS
PERTANIA
2011
K DAN AP
TKAN KUA
UHAN IKA
ASIH
SARJANA
AN BOGO
PLIKASI
ALITAS P
AN NILA
A
OR
xi
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Seleksi Bakteri Proteolitik dan
Aplikasi Enzim Protease untuk Meningkatkan Kualitas Pakan dan Kinerja
Pertumbuhan Ikan Nila adalah karya saya dengan arahan dosen pembimbing dan
belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi manapun.
Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun
tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan
dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Januari 2011
xii
ABSTRACT
TITIN KURNIASIH. Screening of Proteolytic Bacterias and Application of the Bacterial Protease for Increasing Feed Quality and Growth Performance of Nile Tilapia. Under direction of WIDANARNI and NUR BAMBANG PRIYO UTOMO.
Growth performance of fish is highly influenced by the quality of feed, which is determined dominantly by the quality of feedstuff used and also by supplementation of exogenous enzymes. This study aimed to select protease producing bacteria, to optimize their existing potential in producing protease and to evaluate the effect of dietary supplementation of bacterial protease on feed quality and growth performance of nile tilapia. Ten strains were isolated from gastrointestinal tract of Clarias gariepinus and two of them were selected based on their maximum clear zone diameter on casein agar medium and the absence of pathogenic activity against the test fish. The incubation time for maximum protease production of two strains was 72 hours, and the optimum dosage of bacterial protease was 1000 ml /kg feed, which was determined based on the level of hydrolyzed protein. A feeding study was conducted using 7 treatments with 3 replicate groups of nile tilapia (average weight of 4,07 ± 0.25 g), reared under laboratory condition. Result showed that the addition of A1 and L1 bacterial protease gave a significant increase on feed comsumption, growth rate of nile tilapia, apparent digestibility of dry matter and crude protein, protein retention and efficiency in formulated feed. Fish survival was not different among the seven treatments. This result suggested that bacterial protease was effective in improving feed quality and growth performance of nile tilapia when it was added to formulated feed containing poorly digestible feedstuffs.
xiii RINGKASAN
TITIN KURNIASIH. Seleksi Bakteri Proteolitik dan Aplikasi Enzim Protease untuk Meningkatkan Kualitas Pakan dan Kinerja Pertumbuhan Ikan Nila. Dibimbing oleh WIDANARNI dan NUR BAMBANG PRIYO UTOMO.
Kualitas pakan sangat menentukan laju pertumbuhan ikan. Pakan yang dikonsumsi oleh ikan tidak semuanya dapat dicerna namun ada yang dikeluarkan dalam bentuk limbah berupa feses dan sisa metabolisma lain seperti urin dan amoniak. Besarnya pakan yang dikeluarkan dalm bentuk feses tergantung dari kesesuaian komponen pakan dengan kemampuan enzimatik di saluran pencernaan atau daya cerna.
Pakan yang berkualitas selain dihasilkan dari sumber bahan pakan juga dapat dihasilkan dari penambahan enzim dalam pakan. Penelitian mengenai isolasi dan seleksi bakteri dari saluran pencernaan ikan dan aplikasinya pada pakan telah banyak dilaporkan. Informasi bahwa bakteri dalam saluran pencernaan memiliki kemampuan menyumbangkan enzim pencernaan untuk meningkatkan aktivitas pencernaan, menimbulkan pemikiran untuk menyeleksi bakteri proteolitik sebagai penghasil enzim protease, dan mengaplikasikannya pada pakan untuk meningkatkan kinerja pertumbuhan ikan nila.
Penelitian terdiri dari tiga tahap. Tahap pertama adalah isolasi dan seleksi bakteri proteolitik yang terdiri dari uji zona hidrolisis protein dan uji patogenisitas. Tahap kedua adalah optimasi potensi bakteri terpilih sebagai penghasil protease, yang meliputi 1) pengamatan kerapatan optis dan produksi enzim protease bakteri terpilih dan 2) penentuan dosis enzim protease (0, 200, 400, 600, 800, 1000 ml/kg pakan) yang optimal untuk menghidrolisis pakan, berdasarkan parameter derajat hidrolisis protein pakannya. Tahap ketiga adalah uji pertumbuhan dan kecernaan pakan pada ikan nila, menggunakan rancangan acak lengkap (RAL) dengan 7 perlakuan dan 3 ulangan, yaitu A) pakan formulasi tanpa penambahan enzim, B) pakan formulasi yang ditambah enzim protease bakteri A1, C) pakan formulasi yang diberi enzim bakteri L1, D) pakan komersial tanpa enzim (31% P), E) pakan komersial 31% P yang ditambah enzim A1, F) pakan komersial 31% P yang ditambah enzim L1 dan G) pakan komersial 28% P.
Tahap seleksi zona hidrolisis protein mendapatkan 4 isolat dengan zona hidrolisis tertinggi, sedangkan uji patogenisitas hanya meloloskan 2 isolat, yaitu A1 dan L1. Pada tahap optimasi, didapatkan bahwa fase stasioner bakteri A1 dan L1 terjadi pada umur kultur 2 hari, produksi protease tertinggi terjadi pada sekitar umur 3 hari, dan dosis 1000 ml enzim / kg pakan memberikan derajat hidrolisis protein pakan yang tertinggi (91,99%).
xiv komponen pembangun jaringan tubuh menjadi lebih optimal. Penambahan enzim bakteri A1 dan L1 juga secara nyata meningkatkan efisiensi, retensi protein dan palatabilitas pakan formulasi.
Pada pakan komersial 31% P, penambahan enzim protease hanya meningkatkan kecernaan protein dan total pakan (P<0,05), tapi tidak untuk parameter laju pertumbuhan, efisiensi dan retensi protein pakan. Perbedaan efek ini mungkin disebabkan oleh adanya perbedaan kualitas bahan baku penyusun pakannya. Pada pakan komersial 31% P, dimungkinkan bahwa bahan baku yang digunakan sudah memiliki kualitas yang cukup baik, sehingga penambahan enzim A1 dan L1 tidak memberikan efek yang signifikan.
Kelangsungan hidup ikan nila tidak berbeda nyata di antara tujuh perlakuan. Hal ini disebabkan oleh terjaganya kualitas air media pemeliharaan dan tercukupinya kebutuhan nutrisi yang optimal.
xv
© Hak Cipta milik IPB, tahun 2011
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
xvi
SELEKSI BAKTERI PROTEOLITIK DAN APLIKASI ENZIM
PROTEASE UNTUK MENINGKATKAN KUALITAS PAKAN
DAN KINERJA PERTUMBUHAN IKAN NILA
TITIN KURNIASIH
Tesis
sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada
Program Studi Ilmu Akuakultur
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
xviii Judul Tesis : Seleksi Bakteri Proteolitik dan Aplikasi Enzim Protease
untuk Meningkatkan Kualitas Pakan Dan Kinerja Pertumbuhan Ikan Nila
Nama : Titin Kurniasih NRP : C151 080 271
Program Studi : Ilmu Akuakultur
Disetujui,
Komisi Pembimbing
Dr. Ir. Widanarni, M.Si Dr. Ir. Nur Bambang Priyo Utomo, M.Si Ketua Anggota
Diketahui,
Ketua Program Studi Dekan Sekolah Pascasarjana IPB Ilmu Akuakultur
Prof. Dr. Ir. Enang Harris, M.S Prof. Dr. Ir. Khairil Anwar Notodiputro, M.S
xix
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala
karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam
penelitian yang dilaksanakan pada bulan Januari - September 2010 ini adalah
nutrisi akuakultur, dengan judul Seleksi Bakteri Proteolitik dan Aplikasi Enzim
Protease untuk Meningkatkan Kualitas Pakan Dan Kinerja Pertumbuhan Ikan
Nila.
Terima kasih penulis ucapkan kepada Ibu Dr. Ir. Widanarni, M.Si dan
Bapak Dr. Ir. Nur Bambang Priyo Utomo, M.Si selaku dosen pembimbing, atas
arahan, masukan dan dukungan yang diberikan dalam penyusunan tesis ini dan
Ibu Dr. Munti Yuhana, S.Pi, M.Si selaku dosen penguji yang telah banyak
memberikan saran. Di samping itu, penghargaan penulis sampaikan kepada Bapak
Dr. Zafril Imran Azwar, Bapak Dr. Tri Heru Prihadi, Ibu Mulyasari, M.Si, Ibu
Irma Melati, S.Si, Sdr. Teguh, Bapak Yosep Iskandar dan Sdr. M. Rizki dari Balai
Riset Perikanan Budidaya Air Tawar Bogor, yang telah memberikan kesempatan
dan bantuan seluasnya kepada penulis dalam melaksanakan kegiatan penelitian
ini. Ungkapan terima kasih juga disampaikan kepada orang tua (Warsono alm dan
Siti Toipah), suami Waryat, S.Pi, M.Si dan dua putra tersayang Amirul Falah dan
Nasrul Hidayatullah atas doa, cinta kasih, pengertian serta dorongan yang selalu
diberikan. Penulis juga mengucapkan terima kasih kepada teman-teman
Akuakultur 2008 Program Pascasarjana IPB dan teman-teman di lingkup Balai
Riset Perikanan Budidaya Air Tawar Bogor yang tak dapat disebutkan
satu-persatu, untuk kebersamaan dan dukungannya selama ini.
Semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2011
xx
RIWAYAT HIDUP
Penulis dilahirkan di Cilacap pada tanggal 23 Februari 1975 dari ayah
Warsono (alm) dan ibu Siti Toipah. Penulis merupakan anak kedua dari tiga
bersaudara.
Penulis menyelesaikan pendidikan SMA pada tahun 1992 dan diterima di
Universitas Diponegoro Semarang melalui jalur Program Seleksi Siswa
Berprestasi (PSSB) pada tahun yang sama. Penulis memilih Program Sudi
Perikanan pada Fakultas Perikanan dan Ilmu Kelautan (FPIK) Undip dan lulus
pada tahun 1997.
Penulis bekerja di perusahaan pakan Manggalindo selama 1 tahun di tahun
1999. Pada tahun 2000, penulis diterima sebagai CPNS (peneliti) di Balai Riset
Perikanan Budidaya Air Tawar Bogor. Pada tahun yang sama, penulis menikah
dengan Waryat, M.Si dan telah dikaruniai 2 orang putra bernama Amirul Falah
dan Nasrul Hidayatullah. Pada tahun 2008 penulis berkesempatan melanjutkan
xxi
DAFTAR ISI
Halaman
DAFTAR TABEL ... xi
DAFTAR GAMBAR ... xii
DAFTAR LAMPIRAN ... xiii
PENDAHULUAN Latar Belakang ... 1
Perumusan Masalah ... 2
Tujuan dan Manfaat Penelitian ... 2
Hipotesis ... 3
Ruang Lingkup... ... 3
TINJAUAN PUSTAKA Kebutuhan Nutrisi Ikan Nila ... 4
Bahan Baku Sumber Protein Pakan ... 6
Enzim Protease ... 9
Protease Mikrob ... 11
Pemanfaatan Enzim Protease dalam Pakan ... 12
METODOLOGI Tempat dan Waktu Percobaan ... 16
Prosedur Penelitian ... 16
Isolasi dan Seleksi Bakteri Proteolitik ... 16
Optimasi Potensi Bakteri Terpilih... 18
Uji Pertumbuhan dan Kecernaan pada Ikan Nila ... 20
Analisis Data ... 23
HASIL DAN PEMBAHASAN Isolasi dan Seleksi Bakteri Proteolitik ... 24
Optimasi Potensi Bakteri Terpilih sebagai Sumber Enzim Protease ... 26
Uji Pertumbuhan dan Kecernaan pada Ikan Nila ... 31
KESIMPULAN ... 36
DAFTAR PUSTAKA ... 37
xxii
DAFTAR TABEL
Halaman
1 Komposisi asam amino sumber protein bahan pakan ... 7
2 Komposisi proksimat tepung darah ... 8
3 Profil asam amino dalam protein tepung darah ... 8
4 Contoh protease asam, netral dan alkalis ... 10
5 Mikrob penghasil protease dan jenis enzim yang dihasilkannya ... 11
6 Ringkasan metode treatment protease pada TBK ... 13
xxiii
DAFTAR GAMBAR
Halaman
1 Zona hidrolisis kasein oleh bakteri proteolitik dan non proteolitik ... 24
2 Kerapatan optis bakteri A1 dan L1 dalam media TSB ... 26
3 Aktivitas enzim protease bakteri A1 dan L1 (µg/menit.ml) ... 27
4 Derajat hidrolisis protein pakan : uji dosis enzim A1dan L1 ... 29
xxiv
DAFTAR LAMPIRAN
Halaman
1 Prosedur uji zona hidrolisis kasein ... 44
2 Prosedur analisis aktivitas enzim protease ... 45
3 Prosedur analisis proksimat (metode Takeuchi 1988) ... 46
4 Komposisi pakan buatan untuk ikan nila ... 49
5 Hasil analisis proksimat pakan percobaan ... 49
6 Prosedur pengukuran derajat hidrolisis pakan ... 50
7 Prosedur analisis kadar kromium pakan dan feses (Takeuchi 1988) ... 51
8 Luas zona hidrolisis kasein sepuluh isolat bakteri ... 52
9 Hasil uji patogenisitas isolat bakteri proteolitik ... 52
10 Kerapatan optis isolat bakteri proteolitik A1 dan L1 ... 53
11 Aktivitas enzim protease isolat A1 dan L1(µg/menit.ml) ... 54
12 Derajat hidrolisis protein pakan oleh enzim A1 dan L1 ... 54
13 Perhitungan kecernaan total dan protein ... 55
14 Perhitungan konsumsi pakan, efisiensi pakan, laju pertumbuhan spesifik (LPS) dan kelangsungan hidup ikan nila ... 56
15 Perhitungan retensi protein ... 57
16 Analisis ragam dan uji Duncan LPS ikan nila ... 58
17 Analisis ragam dan uji Duncan jumlah konsumsi pakan ... 58
18 Analisis ragam dan uji Duncan kecernaan total pakan ... 59
19 Analisis ragam dan uji Duncan kecernaan protein pakan ... 59
20 Analisis ragam dan uji Duncan efisiensi pakan ... 60
21 Analisis ragam dan uji Duncan retensi protein pakan ... 60
22 Analisis ragam dan uji Duncan kelangsungan hidup ikan nila ... 61
xxv
PENDAHULUAN
Latar Belakang
Dalam industri budidaya perikanan, pakan adalah komponen biaya terbesar
bagi kegiatan operasional budidaya. Kian meningkatnya harga pakan ikan tanpa
disertai kenaikan harga jual ikan hasil budidaya adalah permasalahan yang harus
dihadapi setiap pembudidaya ikan. Tingginya harga pakan ikan terutama
disebabkan oleh tingginya harga bahan baku sumber protein pakan yang sejauh ini
masih didominasi oleh tepung ikan, dan juga tingginya kandungan protein yang
diperlukan dalam formulasinya (Gatlin III et al. 2007). Oleh karena itu, upaya pencarian bahan baku sumber protein alternatif yang dapat mengurangi tingkat
inklusi tepung ikan harus giat dilakukan.
Berbagai jenis bahan baku sumber protein alternatif sudah dikaji, misalnya
tepung bungkil kedelai (Saidy & Gaber 2002) dan tepung darah (Johnson &
Summerfelt 2000). Bahan baku alternatif ini diharapkan ketersediaannya cukup,
kontinyu, mudah didapat dan harganya murah. Tepung darah misalnya, memiliki
kandungan protein cukup tinggi hingga 92% (Johnson & Summerfelt 2000),
sedangkan tepung bungkil kedelai hingga 48% (Li et al. 2000). Namun kecernaan protein dan total kedua bahan baku ini masih perlu ditingkatkan. Koefisien
kecernaan protein untuk ikan kerapu bebek (Cromileptes altivelis) berturut-turut adalah 82.4%, 67.2% dan 55.2% untuk tepung ikan, tepung bungkil kedelai dan
tepung darah (Laining et al. 2003).
Tingkat kecernaan suatu bahan dapat ditingkatkan dengan penambahan
enzim pada pakan benih ikan bandeng (Aslamyah 2006), ikan gurami (Rosmawati
2005; Hasan 2000) dan ikan gilthead seabream (Kolkovski et al. 1993), salah satunya adalah enzim protease (Rosmawati 2005). Enzim protease disekresikan
oleh mikrob proteolitik yang mampu menghidrolisis protein kompleks menjadi
protein sederhana sehingga lebih mudah diserap dan dimanfaatkan ikan.
Bakteri yang terdapat di dalam saluran pencernaan ikan diketahui dapat
memberikan manfaat untuk meningkatkan aktivitas pencernaan ikan, karena
kemampuannya mensekresikan berbagai jenis enzim. Penelitian bakteri pada
saluran pencernaan ikan telah banyak dilaporkan (Clarke & Bauchop 1977; Das &
xxvi Jankauskiene 2002; Tae 2003). Walaupun demikian, informasi mengenai
peranannya sebagai sumber enzim pencernaan untuk pakan ikan dan aplikasinya
untuk predigestion masih sangat terbatas, khususnya di Indonesia. Berpijak pada informasi tersebut dan beberapa hasil penelitian bakteri saluran pencernaan ikan,
maka dibangun suatu pemikiran untuk mengisolasi secara selektif bakteri yang
mempunyai aktivitas proteolitik dari saluran pencernaan ikan lele,
memperbanyak bakteri yang terpilih dan memanfaatkan enzim yang
disekresikannya untuk meningkatkan kecernaan pakan untuk ikan nila
(Oreochromis niloticus).
Perumusan Masalah
Kian meningkatnya harga pakan ikan mengharuskan pembudidaya untuk
mendapatkan alternatif bahan baku sumber protein yang lebih murah dan mudah
didapatkan. Permasalahan yang muncul adalah tingkat kecernaan protein bahan
baku alternatif secara umum lebih rendah. Oleh karena itu, penambahan ekstrak
kasar enzim protease yang disekresikan oleh bakteri proteolitik hasil isolasi dari
saluran pencernaan ikan lele, diharapkan dapat meningkatkan kecernaan pakan
yang mengandung bahan baku alternatif tersebut, sehingga kualitasnya diharapkan
dapat menyamai pakan ikan komersial.
Tujuan dan Manfaat Penelitian
Penelitian ini bertujuan untuk :
1. Mendapatkan bakteri yang mempunyai aktivitas proteolitik dari saluran
pencernaan ikan lele, dan menyeleksi bakteri dengan aktivitas proteolitik
tertinggi.
2. Mengoptimasi potensi bakteri terpilih sebagai penghasil enzim protease.
3. Menguji pengaruh penambahan enzim protease bakteri terhadap kualitas
pakan dan kinerja pertumbuhan ikan nila.
Penelitian ini bermanfaat untuk :
1. Pengembangan ilmu nutrisi ikan, khususnya peranan bakteri sebagai
penyedia enzim alamiah untuk pakan ikan.
2. Mendukung program pemerintah untuk pengembangan kegiatan usaha
budidaya perikanan yang berkelanjutan, dengan menyediakan alternatif
xxvii Hipotesis Penelitian
Hipotesis penelitian ini adalah:
1. Ada bakteri yang mempunyai aktivitas proteolitik dari saluran pencernaan
ikan lele.
2. Bakteri proteolitik yang terpilih dapat dioptimalkan kemampuannya dalam
menghasilkan enzim protease.
3. Enzim protease bakteri berpengaruh terhadap kualitas pakan dan kinerja
pertumbuhan ikan nila.
Ruang Lingkup
Penelitian ini terdiri dari tiga tahap, yaitu tahap isolasi dan seleksi bakteri
proteolitik dari saluran pencernaan ikan lele, tahap optimasi potensi bakteri
terpilih sebagai penghasil enzim protease dan tahap pengujian pengaruh
pemberian enzim protease bakteri terhadap kualitas pakan dan kinerja
xxviii
TINJAUAN PUSTAKA
Kebutuhan Nutrisi Ikan Nila
Kebutuhan nutrisi ikan akan terpenuhi dengan adanya pakan. Komponen
pakan yang berkontribusi terhadap penyediaan materi dan energi tumbuh adalah
protein, karbohidrat dan lemak. Kebutuhan ikan akan protein dipengaruhi oleh
berbagai faktor di antaranya adalah ukuran ikan, temperatur air, kadar pemberian
pakan, kandungan energi dalam pakan yang dapat dicerna dan kualitas protein
(Furuichi 1988).
Protein merupakan molekul kompleks yang terdiri dari asam amino
esensial dan non esensial. Protein adalah nutrien yang sangat dibutuhkan untuk
perbaikan jaringan tubuh yang rusak, pemeliharaan protein tubuh untuk
pertumbuhan, materi untuk pembentukan enzim dan beberapa jenis hormon, dan
juga sebagai sumber energi (NRC 1993). Sekitar 65-75% dari tubuh ikan dalam
berat kering merupakan protein (Halver 2001). Ikan menggunakan protein secara
efisien sebagai sumber energi (Lovell 1989).
Pertumbuhan maksimum pada ikan nila didapat dengan level protein
35-50%, tetapi level optimum dalam pakan komersil untuk ukuran juvenil sampai
dengan dewasa biasanya 25-35% (Popma & Lovshin 1996). Pada kolam atau
tambak yang memiliki pakan alami yang dapat menyumbangkan protein bagi
ikan, kadar protein yang memadai untuk ikan dapat berkisar antara 20-25%
(Webster & Lim 2002).
Jika ikan kekurangan sumber protein, maka pertumbuhan akan terhambat
dikarenakan protein yang dimakan oleh ikan akan digunakan untuk
mempertahankan fungsi jaringan tubuh yang lebih penting. Hal ini bahkan dapat
menyebabkan terjadinya penurunan bobot ikan karena protein yang terkandung
dalam jaringan tubuh ikan dipecah kembali untuk mempertahankan fungsi
jaringan tubuh yang lebih penting tersebut (NRC 1993; Halver 2001).
Lemak merupakan salah satu makronutrien bagi ikan karena selain
berfungsi sebagai sumber energi non protein dan asam lemak esensial, juga
berfungsi memelihara bentuk dan fungsi fosfolipid, membantu dalam absopsi
vitamin yang larut dalam lemak dan mempertahankan daya apung tubuh (NRC
xxix Lemak pakan merupakan sumber asam lemak esensial yang dibutuhkan
ikan untuk pertumbuhan, pemeliharaan dan metabolisme tubuh (NRC 1993). Satu
gram lemak memiliki energi dalam pakan (gross energy) sebesar 9.4 kkal, sedangkan dalam protein dan karbohidrat sebesar 5.6 dan 4.1 kkal (Watanabe
1988).
Jenis asam lemak yang dibutuhkan ikan di antaranya asam lemak omega 3
dan omega 6, berupa asam linolenat, asam linoleat, EPA dan DHA. Akan tetapi
menurut Takeuchi et al. (1983) dalam Watanabe (1988), jenis asam lemak esensial yang dibutuhkan oleh ikan nila adalah asam lemak linoleat. Kadar lemak
sebesar 5% sudah mencukupi untuk kebutuhan ikan nila, tetapi jika kadar lemak
dalam pakan ditingkatkan menjadi 12% akan memberikan pengaruh berupa
perkembangan maksimal pada ikan nila (Webster & Lim 2002).
Menurut Lovell (1989), sumber lemak yang baik untuk ikan nila adalah
berasal dari minyak nabati seperti minyak jagung atau minyak kedelai yang
memiliki kandungan asam linoleat yang ditunjukkan dengan pertumbuhan yang
lebih baik dibandingkan dengan minyak ikan yang memiliki kandungan EPA.
Kekurangan kadar asam lemak omega 3 dan 6 pada pakan dapat menyebabkan
nafsu makan ikan menurun, pertumbuhan lambat, pembengkakan, pucat dan
timbunan lemak di hati.
Karbohidrat merupakan sumber energi yang murah dan dapat
menggantikan atau menghemat penggunaan protein (protein sparing effect) yang lebih mahal sebagai sumber energi (Millamena 2002). Menurut NRC (1993),
karbohidrat dalam pakan dapat berupa serat kasar atau bahan ekstrak tanpa
nitrogen (BETN). BETN mengandung banyak gula dan pati yang bersifat mudah
dicerna sedangkan serat kasar kaya akan lignin dan selulosa yang sukar untuk
dicerna. Lovell (1989) mengemukakan bahwa pemberian tingkat energi yang
optimum dalam pakan sangat penting karena kelebihan dan kekurangan energi
dapat menurunkan pertumbuhan ikan.
Pemanfaatan karbohidrat oleh ikan berbeda-beda bergantung pada
kompleksitas karbohidrat. Kadar optimum karbohidrat dalam pakan sulit untuk
ditentukan karena protein dan lemak mendahului fungsi karbohidrat sebagai
xxx karbohidrat kompleks sebagai sumber energi utama dalam pakannya pada level
yang tinggi. Ikan-ikan omnivora dan herbivora dapat mencerna karbohidrat yang
berasal dari tumbuhan (Yamada 1983). Ikan-ikan karnivora dapat memanfaatkan
karbohidrat optimum pada tingkat 10-20% dalam pakannya sedangkan ikan-ikan
omnivora mampu memanfaatkan karbohidrat optimum sebesar 30-40% dalam
pakan (Furuichi 1988).
Komponen lain yang dibutuhkan dalam pakan ikan yaitu vitamin dan
mineral. Jumlah vitamin dan mineral yang dibutuhkan dalam pakan sangatlah
kecil namun kehadirannya dalam pakan sangat penting karena dibutuhkan untuk
tumbuh dan menjalankan beberapa fungsi tubuh. NRC (1993) menjelaskan bahwa
mineral merupakan senyawa yang digunakan untuk proses respirasi, osmoregulasi
dan pembentukan kerangka tulang. Vitamin merupakan senyawa organik
kompleks yang diperlukan untuk pertumbuhan normal, reproduksi, kesehatan dan
metabolisme secara umum.
Bahan Baku Sumber Protein Pakan
Tepung Ikan
Tepung ikan merupakan komponen penting dalam pakan. Menurut Lovell
(1989) kandungan protein tepung ikan berkisar 60-80% dengan tingkat kecernaan
protein tinggi (80-95%) dan memiliki kadar lisin dan metionin tinggi. Lisin dan
metionin adalah jenis asam amino yang jumlahnya sedikit pada bahan-bahan
nabati. Tepung ikan menjadi sumber protein utama yaitu sekitar 35-50% dalam
formulasi pakan ikan salmon, udang dan spesies ikan laut lainnya (Dong & Hardy
2000). Protein tepung ikan memiliki kualitas yang unggul, baik dari komposisi
asam amino dan kecernaannya serta disukai oleh ikan (Li et al. 2000).
Nilai nutrisi atau kualitas dari tepung ikan tergantung asal spesies ikan,
tingkat kesegaran ikan, suhu pemanasan saat produksi, penambahan antioksidan,
penyimpanan dan kondisi saat pemindahan (Jobling et al. 2001). Kadar protein dan kadar abu tepung ikan menentukan harga bahan. Tepung ikan yang berasal
dari ikan hering, capelin dan sidat pasir memiliki kandungan protein yang tinggi
dan kadar abu rendah sehingga harganya paling mahal. Sedangkan tepung ikan
yang berasal dari ikan menhaden dan tuna kadar proteinnya lebih rendah dan
xxxi Tepung Bungkil Kedelai (TBK)
TBK diperoleh dari kedelai yang telah diperas minyaknya dengan
ekstraksi pelarutan atau proses penekanan. Protein tepung bungkil kedelai berkisar
antara 42-48% dan kandungan lemaknya berkisar antara 0.5-3.5%. Kandungan
protein dan lemak tepung kedelai tergantung dari proses pembuatan tepung (Li et al. 2000).
TBK telah banyak digunakan sebagai bahan suplemen pada pakan, karena
mempunyai komposisi asam amino yang lebih baik dibandingkan dengan bahan
nabati lain untuk memenuhi kebutuhan ikan (Pongmaneerat & Watanabe 1993).
TBK juga mengandung faktor anti nutrisi yang dapat dihilangkan atau
dideaktivasi melalui pemanasan dan pengeringan (Jobling et al. 2001). Komposisi asam amino beberapa bahan baku sumber protein pakan (NRC 1993) disajikan
pada Tabel 1.
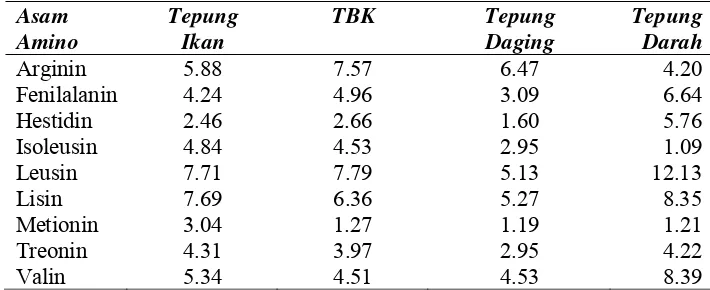
Tabel 1. Komposisi asam amino sumber protein bahan pakan (g/100 g protein)
Asam Amino
Tepung Ikan
TBK Tepung Daging
Tepung Darah Arginin 5.88 7.57 6.47 4.20 Fenilalanin 4.24 4.96 3.09 6.64 Hestidin 2.46 2.66 1.60 5.76 Isoleusin 4.84 4.53 2.95 1.09 Leusin 7.71 7.79 5.13 12.13 Lisin 7.69 6.36 5.27 8.35 Metionin 3.04 1.27 1.19 1.21 Treonin 4.31 3.97 2.95 4.22 Valin 5.34 4.51 4.53 8.39
Tepung Darah
Salah satu alternatif bahan substitusi tepung ikan adalah tepung darah
(Bureau et al. 1999). Di antara bahan makanan dari hewan, tepung darah paling tinggi kadar proteinnya, yaitu mencapai 92%. Tepung darah (blood meal) dihasilkan dari darah hewan yang dikeringkan dan digiling. Darah adalah
kumpulan sel dalam jumlah besar dengan kandungan air tinggi (80%). Protein
globular, albumin dan globulin-D terdapat sebanyak 59, 16 dan 13% dari total
nitrogen yang terkandung dalam tepung darah (Marichal et al. 2000). Tepung darah adalah sumber yang kaya leusin tetapi miskin metionin dan isoleusin
xxxii darah disajikan pada Tabel 2 dan 3. Kombinasi tepung darah dan tepung jagung
akan membantu untuk memperkaya komposisi pakan ikan karena tepung jagung
memiliki kandungan lisin yang rendah dan isoleusin yang tinggi (Harris 1980).
Tabel 2. Komposisi proksimat tepung darah (Halimatusadiah 2009)
Komposisi (% bobot kering) Protein 84.52 Lemak 7.05 Abu 3.23 Serat Kasar 0.75 BETN 4.44
Tabel 3. Profil asam amino dalam protein tepung darah (Johnson & Summerfelt 2000)
Profil Asam Amino (%) Metionin 0.76 Treonin 3.12 Lysin 8.75 Triptofan 1.54 Isoleusin 0.63 Hestidin 7.32 Valin 9.12 Leusin 13.61 Arginin 4.12 Phenylalanin 6.28
Tepung darah komersial diproduksi dalam jumlah besar dengan cara
spray-dried, dan disebut spray-dried blood cells (SBC). Tepung darah SBC dibuat dengan cara menyemprotkan darah segar ke aliran udara panas bersuhu 316oC kemudian dimasukkan ke dalam vakum bersuhu rendah yaitu 49oC. Produk SBC mengandung protein sangat tinggi (92% berat kering), kandungan lisin dan leusin
tinggi (9% dan 13.61% dari total protein) dan hanya mengandung sedikit mineral
fosfor (0.33%). SBC mengandung Fe sangat tinggi sampai pada level 2700 mg/kg
(Johnson & Summerfelt 2000), dibandingkan dengan tepung ikan yang
mengandung Fe sekitar 400-800 mg/kg, dan tepung kedelai 140 mg/kg. Kadar zat
besi yang tinggi ini memungkinkan untuk pemakaian tepung darah dalam pakan
xxxiii Kendala dalam pemakaian tepung darah adalah tingkat kecernaan yang
rendah dibandingkan tepung ikan, sebagaimana dilaporkan Laining et al. (2003) yaitu sebesar 55.2%.
Enzim Protease
Protease adalah enzim yang mampu menghidrolisis ikatan peptida pada
protein. Molekul yang berfungsi sebagai unit penyusun polimer protein adalah
asam amino yang terangkai melalui ikatan peptida. Jumlah asam amino penyusun
protein berkisar dari puluhan sampai ribuan. Biasanya protein yang tersusun oleh
lebih dari 10 asam amino dikenal sebagai polipeptida. Istilah protein ditujukan
bagi polimer asam amino dengan jumlah di atas 100 (Suhartono 1989a).
Kerentanan struktur protein terhadap hidrolisis oleh protease berkaitan
dengan strukturnya. Keseluruhan struktur protein dan fungsi hayatinya ditentukan
oleh struktur primer protein, yaitu deret asam amino pada protein. Jumlah ikatan
peptida yang dapat diuraikan oleh suatu protease bergantung pada jenis asam
amino penyusun protein dan jenis asam amino yang letaknya saling berdekatan.
Selain itu, struktur sekunder, acak, tersier dan kuarterner menentukan efektivitas
kerja protease terhadap protein tersebut. Hal lain yang penting diketahui adalah,
protease memecah ikatan peptida dengan bantuan molekul air (Suhartono 1989a).
Kelompok protease atau proteinase diproduksi secara ekstraselular dan
intraselular, serta memainkan peranan penting dalam proses metabolisme sel dan
regulasinya. Peran utama protease ekstraselular di alam, sebagaimana enzim
ekstraselular lainnya, adalah menghidrolisis substrat polimer (polipeptida)
berukuran besar menjadi molekul kecil sehingga dapat diserap oleh sel. Protease
intraselular membantu keseimbangan antara sintesis dan degradasi protein dan
memegang peranan penting dalam berbagai proses seperti pembentukan dan
germinasi spora, pematangan protein, koagulasi darah fibrinolisis, pengontrolan
tekanan darah, diferensiasi, modifikasi dan sekresi berbagai enzim (Rao et al. 1998).
Pembentukan spora bakteri dan bagian tubuh kapang melibatkan
proteolisis dan mungkin juga melibatkan protease ekstraselular. Pada beberapa
xxxiv karbon dan nitrogen. Banyak laporan menyatakan bahwa sintesis protease netral
dan jenis subtilisin mengalami hambatan selama pertumbuhan pada media miskin
karbon (Suhartono 1989a).
Ditinjau dari lingkungan daya kerjanya, protease digolongkan menjadi
protease asam yang bekerja pada pH asam, protease netral yang bekerja pada pH
netral dan protease alkalis yang bekerja pada pH basa (Tabel 4).
Tabel 4. Contoh protease asam, netral dan alkalis (Suhartono 1989a) Jenis Protease Keterangan
Protease asam
Renin Renin digunakan di dalam pembuatan keju
Renin mikrob Dihasilkan oleh Mucor miehei, Mucor pusillus dan Endothia parasitica
Pepsin Biasanya diperoleh dari lambung sapi atau babi Protease asam kapang Biasanya dihasilkan oleh jenis Aspergillus dan
Rhizopus
Protease netral
Tripsin pankreas Endoprotease dari pankreas sapi atau babi Papain Endoprotease dari getah pepaya
Bromelin Endoprotease dari tanaman nenas Protease bakteri Endoprotease dari Bacillus subtilis
Protease alkali
Protease bakteri Spesifikasi luas dihasilkan oleh sejumlah Bacillus
Protease termasuk golongan enzim yang relatif ’kuat’, karena tahan
kondisi pH dan suhu lingkungan yang ekstrim. Dengan demikian, enzim ini lebih
mudah ditangani. Salah satu yang agak menyimpang dari sifat ini adalah pepsin,
yang stabil di dalam larutan enzim encer, tetapi cepat terdenaturasi pada pH
netral. Tampaknya hal ini merupakan bagian dari mekanisme protektif makhluk
hidup yang dirancang untuk meniadakan aktivitas pepsin di luar lambung
(Suhartono 1989a).
Protease Mikrob
Protease dapat dihasilkan oleh hewan, tumbuhan dan mikrob. Meskipun
demikian yang paling banyak digunakan saat ini sebagai penghasil protease
komersial adalah mikrob karena produktivitasnya yang tinggi sangat efisien
dipandang dari sudut waktu dan tempat produksi, kemudahan pengaturan produksi
dan tingginya peluang perbaikan produksi melalui teknik optimasi fermentasi,
xxxv mencantumkan nama species mikrob penghasil protease dan jenis yang
dihasilkannya.
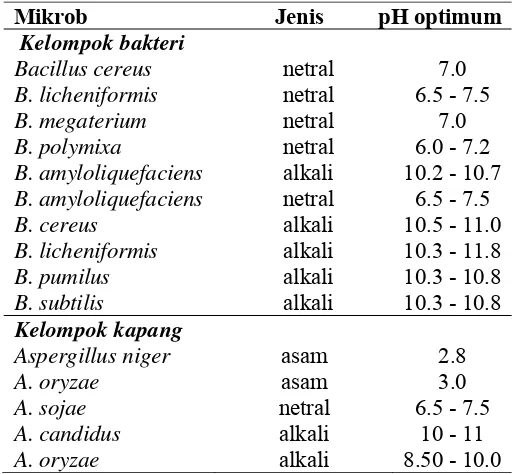
Tabel 5. Mikrob penghasil protease dan jenis enzim yang dihasilkannya (Suhartono 1989b)
Mikrob Jenis pH optimum
Kelompok bakteri
Bacillus cereus netral 7.0
B. licheniformis netral 6.5 - 7.5
B. megaterium netral 7.0
B. polymixa netral 6.0 - 7.2
B. amyloliquefaciens alkali 10.2 - 10.7 B. amyloliquefaciens netral 6.5 - 7.5
B. cereus alkali 10.5 - 11.0
B. licheniformis alkali 10.3 - 11.8
B. pumilus alkali 10.3 - 10.8
B. subtilis alkali 10.3 - 10.8 Kelompok kapang
Aspergillus niger asam 2.8
A. oryzae asam 3.0
A. sojae netral 6.5 - 7.5
A. candidus alkali 10 - 11
A. oryzae alkali 8.50 - 10.0
Berbagai jenis bakteri seperti Bacillus, Lactobacillus, Pseudomonas, Clostridium, Proteus dan Serratia serta kapang Aspergillus, Penicillium, Rhizopus, Endothia dan Mucor merupakan penghasil enzim protease yang berpotensi. (Rao et al. 1998). Spesies Bacillus banyak menghasilkan protease serin alkali dan protease logam, di antaranya protease serin alkali yang dihasilkan
oleh Bacillus licheniformis yang lebih dikenal dengan nama subtilisin. Jenis enzim ini juga diproduksi oleh Bacilus pumilus. Dari golongan kapang, protease yang banyak dihasilkan umumnya juga termasuk golongan serin alkali. Kapang
yang memproduksi enzim protease antara lain Aspergillus orizae, Aspergillus sydowi dan Aspergillus flavus. Kapang penghasil enzim termostabil dengan suhu optimum sekitar 65-70oC adalah Tritirachum album dan Malbranchea ounchella. Protease yang dihasilkan oleh Staphylococcus aureus dinamakan protease staphylococcus, sedangkan clostripain yang termasuk dalam golongan protease
xxxvi Penggunaan protease dalam berbagai industri menyebabkan nilai
penjualannya meningkat. Industri pangan menggunakan protease untuk
pengempukan daging, penjernihan bir, pembuatan protein, hidrolisat dan kecap,
serta pembuatan roti dan kue dengan tekstur khusus. Industri detergen
memanfaatkan protease untuk komponen pembersih yang bersifat biodegradable (ramah lingkungan), sedangkan industri kulit memanfaatkan protease untuk
proses perontokan bulu (dehairing) dan pra pewarnaan. Selain itu protease dipakai untuk mengolah skleroprotein ulat sutera sebelum proses pemintalan
benang dan untuk campuran salep penghalus bekas luka dan obat bantu
pencernaan. Dunia bioteknologi modern memerlukan protease di dalam
prosedur-prosedur ekstraksi DNA dan pengolahan protein terapis (Rao et al. 1998). Pemanfaatan Enzim Protease dalam Pakan
Pengujian in Vitro
Pretreatment dengan enzim yang dilakukan sebelum prosesing pakan
merupakan alternatif cara yang lebih aman, dibandingkan memberikan enzim
secara langsung ke dalam saluran pencernaan hewan target melalui pakan. Cara
yang pertama lebih praktis; enzim cukup bekerja aktif selama masa inkubasinya
saja terhadap substrat, biasanya 24 jam (Thorpe & Beal 2001; Aslamyah 2006;
Fitriliyani 2010), dan setelah produk hidrolisis didapatkan, maka enzim tersebut
tidak perlu dipertahankan aktivitasnya lagi.
Bahan baku yang banyak dikaji efeknya setelah ditreatment dengan enzim
protease adalah kedelai. Target utama pemberian protease pada kedelai adalah
kandungan anti-nutritional factors (ANFs), seperti inhibitor tripsin, lektin dan protein antigenik. Tabel 6 menggambarkan detail treatment protease pada TBK
dalam penelitian-penelitian terhadap babi dan unggas.
Tabel 6. Ringkasan metode treatment protease pada TBK.
Bahan Enzim Treatment Referensi
TBK Protease asam
0.1% protease ditambahkan pada TBK (800 g kg-1) pH 4.5. Inkubasi 3 jam 50oC, pH netral, keringkan 65oC.
Rooke et al.
(1996) TBK Protease
alkali
0.1% protease ditambahkan pada TBK (800 g kg-1) pH 8.5. Inkubasi 16 jam 50oC, tidak dikeringkan.
Rooke et al. (1998) TBK Subtilisin 0.1% protease ditambahkan pada TBK (berikan air 1:2)
pH 4.5. Inkubasi 3 jam 50oC, pH 7, keringkan 65oC.
Caine et al. (1997) TBKTL Protease
(P4)
0.25% protease ditambahkan pada TBK (berikan air 1:3) Inkubasi 24 jam 20oC, tak dikeringkan.
Beal et al. (1998a) TBKTL Protease
(P3)
0.25% protease ditambahkan pada TBK (berikan air 1:3) Inkubasi 24 jam, 20oC, tak dikeringkan.
xxxvii Note : TBK = tepung bungkil kedelai, TBKTL = TBK tinggi lemak
Huo et al. (1993) melaporkan bahwa empat jenis protease bakterinya mampu menginaktivasi inhibitor tripsin dan lektin pada kedelai mentah.
Penurunan level inhibitor tripsin yang paling efektif (96%) terjadi pada dosis
protease 1% dan lama inkubasi 12 jam. Menurutnya, protease bakteri lebih efektif
dibandingkan protease kapang dalam menurunkan inhibitor tripsin. Rooke et al. (1996) mendapatkan bahwa TBK yang diberi protease memiliki kandungan
protein antigenik yang lebih rendah dibandingkan dengan yang tidak. Caine et al. (1997) mengobservasi kondisi optimal untuk treatment TBK dengan subtilisin Bacillus subtilis dan mendapatkan bahwa inkubasi pada suhu 50oC dan pH 4.5 merupakan kondisi optimal. Mereka juga melaporkan adanya peningkatan
kelarutan protein yang signifikan. Beal et al. (1998a) menguji potensi 3 protease secara in vitro dan mendapatkan adanya peningkatan kecernaan nitrogen yang signifikan (P < 0.05) sebesar 5–12% dibandingkan kontrolnya. Beal et al (1998c) juga menguji efek salah satu dari 3 protease tersebut (P4) menggunakan
SDS-PAGE, dan menemukan adanya penurunan jumlah dan densitas pita protein
dengan bobot molekul lebih besar dari 66 kDA, mengindikasikan adanya reaksi
hidrolisis protein. Rooke et al. (1998) juga menggunakan SDS-PAGE untuk menguji efek protease P1 dan P2 terhadap TBK, dan ternyata konsentrasi nitrogen
ά-amino terlarut meningkat setelah perlakuan. P1 menurunkan konsentrasi protein antigenik lebih baik daripada P2.
Pengujian in Vivo
Penggunaan enzim protease yang bertujuan untuk meningkatkan
kecernaan protein dan efisiensi pakan telah diteliti secara intensif pada unggas dan
babi. Penelitian pada babi melaporkan adanya peningkatan signifikan untuk
efisiensi pakan, tetapi tidak untuk kecernaan protein dan energi (O’-Doherty &
xxxviii harian pada babi yang mengkonsumsi pakan berbasis TBK yang ditreatment
dengan protease. Beal et al. (1998b ; 1999) juga melaporkan adanya peningkatan signifikan untuk parameter pertambahan bobot harian dan efisiensi pakan pada
babi yang diberi pakan berbasis TBK setelah ditreatment enzim protease.
Sedangkan Szczurek et al. (2001) mendapatkan bahwa penambahan protease tidak memberikan efek yang berbeda untuk kecernaan protein dan energi. Perbedaan
efek ini mungkin disebabkan oleh adanya perbedaan jenis protease dan formulasi
pakan yang digunakan.
Pemberian enzim protease eksogen ke dalam pakan dengan tujuan
meningkatkan pertumbuhan pada ikan telah dilakukan oleh beberapa peneliti.
Hasan (2000) memberikan enzim papain untuk pakan benih ikan gurami, dan
melaporkan bahwa laju pertumbuhan harian, efisiensi pakan, retensi protein dan
kelangsungan hidup tertinggi terdapat pada pemberian pakan yang ditambah
enzim sebanyak 1.3 – 1.7% dari berat pakan. Penelitian Kolkovski et al. (1993) yang memberikan enzim berasal dari pankreas dalam pakan larva gilthead seabream (Sparus aurata, Sparidae, Linnaeus), mendapatkan bahwa pakan buatan yang diperkaya dengan enzim dari pankreas memberikan pertumbuhan yang lebih
tinggi dibandingkan dengan yang diberi pakan alami. Laju penyerapan dari pakan
yang diperkaya enzim didapat sebesar 30%, dan kelangsungan hidup dari
ketiganya tidak memperlihatkan perbedaan. Rosmawati (2005) melaporkan
adanya peningkatan signifikan untuk kecernaan protein pakan ikan gurami setelah
diberi enzim pepsin dan pankreatin, tetapi tidak mendapatkan hasil
menggembirakan untuk parameter pertumbuhan. Ng et al. (2002) menguji pengaruh pemberian enzim komersial Allzyme Vegpro yang mengandung
protease terhadap pakan berbasis PKM (palm kernel meal), dan melaporkan bahwa penambahan enzim meningkatkan laju pertumbuhan dan efisiensi pakan
dibandingkan pakan kontrolnya. Davis et al. (1998) memberikan enzim protease pada pakan udang dan melaporkan adanya peningkatan kecernaan protein pakan
dari 65.3% menjadi 74.3% pada dosis 0.4 gram protease / 100 g pakan, namun
pada uji pertumbuhan, penambahan enzim protease pada dosis tersebut justru
xxxix (FP) dan canola:pea (CP) untuk ikan rainbow trout (Oncorhynchus mykiss). Hasilnya sangat menarik bahwa produk CP yang diberi protease meningkat
kecernaan total, protein, lemak dan energinya secara signifikan dibandingkan
kontrolnya, sedangkan produk FP tidak ada perubahan. Demikian juga pada uji
pertumbuhan, penambahan protease pada pakan berbasis CP meningkatkan
efisiensi pakannya, tetapi tidak untuk pakan berbasis FP. Penulisnya
merekomendasikan perlunya dilakukan penelitian mendetail mengenai efek
xl BAHAN DAN METODE
Tempat dan Waktu Percobaan
Kegiatan isolasi dan seleksi bakteri proteolitik dilakukan di Laboratorium
Mikrobiologi Nutrisi, Balai Riset Perikanan Budidaya Air Tawar (BRPBAT)
Bogor, kegiatan ekstraksi enzim protease dilakukan di Laboratorium Institute de Recherche pour le Developpement (IRD-Perancis) di Balai Riset Budidaya Ikan Hias Depok, kegiatan pengujian aktivitas enzim protease dilakukan di
Laboratorium Nutrisi Fakultas Peternakan IPB, pengujian hidrolisis pakan secara
in vitro dan analisis proksimat bahan baku dan pakan percobaan dilakukan di Laboratorium Kimia Nutrisi, BRPBAT Bogor, dan percobaan in vivo dilakukan di Laboratorium Basah Nutrisi, BRPBAT Bogor. Penelitian ini dilaksanakan dari
bulan Januari-September 2010.
Prosedur Penelitian
Isolasi dan Seleksi Bakteri Proteolitik
Isolasi Bakteri Proteolitik
Prosedur isolasi bakteri yang mempunyai aktivitas proteolitik dilakukan
dengan metode selektif, yang mengacu pada metode yang dilakukan pada hewan
terrestrial seperti petunjuk Hungate (1966), serta mengkombinasikannya dengan
prosedur isolasi bakteri dari saluran pencernaan ikan seperti yang dilakukan oleh
Nakayama et al. (1994); Hoshino et al. (1997); Jankauskiene (2002) dan Tae (2003).
Sumber inokulum berasal dari saluran pencernaan ikan lele dan air kolam
pemeliharaan ikan lele sebagai pembanding. Dasar pemikirannya adalah bahwa
lele memiliki kelebihan dalam hal efisiensi pakan, dengan nilai konversi
mendekati 1. Diduga kelebihannya ini disebabkan karena lele berasosiasi dengan
bakteri yang mampu mensekresikan enzim-enzim pencernaan, terutama protease,
pada saluran pencernaannya. Isolasi bakteri juga dilakukan terhadap sampel air
kolam pemeliharaan lele, dengan anggapan bahwa komposisi bakteri di saluran
pencernaan suatu organisme, dimungkinkan tidak akan berbeda jauh dengan
komposisi bakteri di air media hidupnya.
Pengambilan isi saluran pencernaan ikan lele sebagai sumber inokulum
xli ikan lele dewasa yang telah dimatikan. Lambung dan usus digerus, kemudian
sebanyak 1 gram ditambah dengan 1 ml cairan fisiologis (NaCl 0.85%) steril
(Aslamyah 2006) dan dihomogenkan dengan vortex.
Sumber inokulum diambil sebanyak 1 ml dan diencerkan dalam tabung
pengenceran serial yang berisi 9 ml larutan fisiologis, sebanyak 10 tabung
pengencer (hingga pengenceran 1010 kali). Setelah dihomogenkan, maka dari setiap tabung pengencer diambil larutan sebanyak 0.1 ml, dan disebarkan dalam
cawan petri berisi TSA yang dibuat secara duplo. Kultur ini kemudian diinkubasi
pada suhu 29oC selama 24-48 jam sampai koloni bakteri dapat tumbuh, dalam suasana aerob.
Koloni bakteri yang tumbuh diidentifikasi berdasarkan perbedaan warna,
bentuk dan ukurannya. Setiap jenis koloni yang didapat dimurnikan dengan
metode penggoresan kuadran, sampai didapatkan koloni bakteri yang tunggal dan
seragam.
Kultur murni selanjutnya diperbanyak atau diperkaya untuk mendapatkan
isolat. Sebagian isolat bakteri digunakan sebagai kultur stok dan sebagian lagi
dipakai sebagai inokulum pada percobaan berikutnya. Pengayaan dilakukan
dengan cara menumbuhkan masing-masing isolat ke dalam media TSB kemudian
diinkubasi pada suhu 29 oC selama 24 jam. Kultur yang didapat siap untuk diawetkan. Pengawetan dilakukan dengan menyimpan isolat-isolat yang telah
diperoleh ke dalam media gliserol 15-20% yang selanjutnya disebut kultur stok.
Seleksi Bakteri Proteolitik
Untuk mendapatkan bakteri yang berpotensi tinggi sebagai penghasil
enzim protease, maka seleksi bakteri dilakukan melalui tahapan pengujian
aktivitas proteolitik dan pengujian patogenisitas. Pengujian aktivitas proteolitik
bertujuan untuk mengukur besarnya aktivitas proteolitik masing-masing isolat
dengan uji hidrolisis kasein (Lampiran 1). Aktivitas proteolitik ditandai dengan
adanya zona bening di sekeliling isolat yang ditumbuhkan pada media agar yang
mengandung kasein 2%. Pengujian patogenisitas bertujuan untuk mengetahui
apakah isolat bakteri yang diperoleh merupakan bakteri patogen terhadap ikan uji
atau tidak. Masing-masing isolat disuntikkan pada kelompok ikan nila sehat
xlii dengan kandungan bakteri sebanyak 107 cfu/ml. Sebagai kontrol digunakan larutan fisiologis yang juga disuntikkan pada satu kelompok ikan nila.
Pemantauan dilakukan setiap hari selama dua minggu setelahnya, apakah
didapatkan perbedaan mortalitas apabila dibandingkan dengan kontrol, atau
adanya tanda klinis penyakit pada ikan uji.
Setelah melakukan kedua tahap seleksi bakteri proteolitik, selanjutnya
ditentukan isolat bakteri terpilih yang akan menjalani pengujian selanjutnya.
Seleksi ditentukan berdasarkan peringkat luasnya zona hidrolisis kasein, dan yang
terbukti tidak bersifat patogen terhadap ikan nila.
Optimasi Potensi Bakteri Terpilih sebagai Sumber Enzim Protease
Pemantauan Produksi Enzim Protease Bakteri
Percobaan ini dilakukan dengan mengukur produksi enzim protease yang
disekresikan oleh bakteri proteolitik terpilih dari waktu ke waktu, yang diukur dari
aktivitasnya (Lampiran 2). Tujuannya untuk mendapatkan titik waktu yang
optimal untuk pemanenan enzim protease. Pengamatan dilakukan dilakukan setiap
4 jam sekali selama 4 hari (25 titik pemantauan) dari waktu kulturnya.
Pemantauan produksi enzim protease ini disertai dengan pengamatan optical density (OD) atau kerapatan optis dengan spektrofotometer pada panjang gelombang 620 nm (Hadioetomo 1990). Nilai prosentase transmitan dikonversi
menjadi nilai kerapatan optik mengikuti tabel menurut Hadieutomo (1993).
Mula-mula bakteri proteolitik terpilih diperbanyak dalam media TSB di
dalam tabung-tabung reaksi berkapasitas 15 ml (masing-masing 25 tabung).
Produksi enzim kasar mengacu pada metode yang dilakukan Wang et al. (2008), dengan modifikasi pada suhu dan pengenceran media yang digunakan. Sebanyak
0.1 ml sumber inokulum bakteri proteolitik yang terpilih diinokulasikan dalam
setiap 9.9 ml media di dalam tabung reaksi, kemudian diinkubasi pada suhu 37oC. Setelah itu kultur dihentikan dengan cara sentrifus pada titik waktu yang
diinginkan (waktu ke-0, 4, 8, 12, 16 jam, dan seterusnya) dengan kecepatan
12.000 g selama 20 menit pada suhu 4 oC. Supernatan yang diperoleh (filtrat ekstrak enzim kasar) ditambah ammonium sulfat (608 g/L) dan disimpan semalam
xliii mM pH 7, dan diuji aktivitas enzim proteasenya di laboratorium dengan metode
Bergmeyer dan Graß1 (1986).
Penentuan Dosis Enzim Protease Bakteri
Percobaan ini bertujuan menguji secara in vitro dosis enzim protease yang optimal untuk mengkatalisis reaksi hidrolisis protein. Perlakuan yang diujikan
adalah ekstrak enzim protease yang berasal dari kultur bakteri A1 dan L1
sebanyak 0, 200, 400, 600, 800 dan 1000 ml /kg pakan, dengan 2 ulangan. Stok
kultur disiapkan dengan cara menumbuhkan 40 ml sumber inokulum cair bakteri
dalam 3960 ml TSB baru dan diinkubasi selama 72 jam pada suhu 37oC. Homogenisasi dengan vortex dilakukan sesering mungkin selama inkubasi ini.
Perlakuan dosis 1000 ml misalnya, disiapkan dengan cara mengambil stok kultur
yang telah siap sebanyak 1000 ml. Kultur cair ini selanjutnya disentrifus dengan
kecepatan 12.000 g pada suhu 4oC selama 20 menit. Supernatan diambil dan ditambah dengan ammonium sulfat (608 g/L), disimpan semalam pada suhu 4oC, dan disentrifus kembali pada kecepatan 12.000, suhu 4 oC selama 20 menit. Endapan yang terbentuk diambil dan dilarutkan kembali dalam 50 ml buffer fosfat
50 mM pH 7. Sediaan ini dapat disimpan lama dalam freezer, dan dapat
digunakan kembali apabila diperlukan. Perlakuan dosis yang lain disiapkan
dengan cara yang sama, dengan menyesuaikan volume stok kultur yang diambil
sesuai perlakuannya (0, 200, 400, 600 dan 800 ml stok kultur).
Ekstrak enzim kasar yang telah dikeluarkan dari freezer dibiarkan dalam
suhu kamar beberapa saat supaya suhunya menyesuaikan. Supaya tercampur rata
pada campuran bahan baku pakan, maka sediaan ekstrak enzim kasar diencerkan
dengan air hangat hingga volumenya 1000 ml, dihomogenkan, kemudian
dicampurkan pada kombinasi bahan baku untuk 1 kg pakan yang telah dihaluskan
dan diaduk merata. Bahan baku penyusun pakan formulasi dalam percobaan ini
sebelumnya telah dianalisis komposisi proksimatnya dengan metode yang
tercantum pada Lampiran 3, dan komposisi bahan dan proksimat pakan tercantum
pada Lampiran 4 dan 5. Campuran pakan dan enzim kasar ini kemudian
diinkubasi selama 24 jam pada suhu 37oC. Parameter yang diamati adalah derajat hidrolisis protein (Lampiran 6).
xliv Pakan yang digunakan dalam penelitian ini adalah pakan buatan
(formulasi) dan pakan komersial sebagai kontrol. Komposisi bahan pakan
formulasi dan komposisi proksimat pakan percobaan tersaji di Lampiran 4 dan 5.
Enzim yang diberikan pada pakan percobaan adalah enzim dari bakteri A1
dan L1, dengan dosis 1000 ml / kg pakan. Cairan enzim dituangkan ke dalam
campuran bahan baku pakan formulasi atau pakan komersial yang sudah
dihaluskan, sesuai dengan perlakuan yang ditentukan, dan diaduk merata.
Selanjutnya inkubasi dilakukan pada suhu 37oC selama 24 jam.
Rancangan dalam percobaan ini adalah Rancangan Acak Lengkap (RAL)
dengan perlakuan sebagai berikut :
A. Pakan formulasi kontrol (kadar protein 28%)
B. Pakan formulasi yang ditambah enzim dari bakteri A1
C. Pakan formulasi yang ditambah enzim dari bakteri L1
D. Pakan komersial kontrol berkadar protein 31%
E. Pakan komersial yang ditambah enzim dari bakteri A1
F. Pakan komersial yang ditambah enzim dari bakteri L1
G. Pakan komersial berkadar protein 28%
Ikan nila dengan berat rata-rata 4.07 ± 0.25 gram ditebar dengan
kepadatan 10 ekor per akuarium. Wadah yang digunakan adalah akuarium
berukuran 60 x 60 x 50 cm sebanyak 21 buah, yang masing-masing diisi air
bervolume 90 liter dan dilengkapi sistem resirkulasi. Pengaturan dan penempatan
wadah perlakuan dilakukan secara acak dengan menggunakan bilangan acak
(Steel & Torrie 1995).
Pemberian pakan dilakukan 3 kali sehari sebanyak 5% dari bobot
biomassa ikan nila per akuarium per hari. Penyesuaian bobot biomassa ikan uji
dilakukan dengan sampling setiap 15 hari sekali. Jumlah pakan yang diberikan
dicatat untuk mendapatkan data konsumsi pakan, efisiensi pakan dan retensi
protein. Percobaan pertumbuhan ini dilakukan selama 60 hari.
Penggantian air di tandon filter dilakukan setiap 3 hari sekali, dan
penyiponan kotoran dilakukan setiap hari. Pengukuran kualitas air dilakukan pada
awal, tengah dan akhir masa pemeliharaan meliputi suhu, pH, oksigen terlarut dan
xlv Parameter kinerja pertumbuhan yang diamati adalah:
Laju Pertumbuhan Spesifik (LPS)
Laju pertumbuhan spesifik ikan uji dihitung mengikuti rumus yang
digunakan oleh Mundheim et al. (2004) yaitu: LPS = (ln Wt – ln Wo) x 100
T Keterangan:
LPS = laju pertumbuhan spesifik (%)
Wt = rata-rata bobot individu pada akhir penelitian (g) Wo = rata-rata bobot individu pada awal penelitian (g) T = lama waktu pemeliharaan (hari)
Jumlah Konsumsi Pakan
Jumlah pakan yang dikonsumsi ikan uji dihitung dengan cara menimbang
pakan yang diberikan setiap hari, dan juga pakan yang tersisa setiap hari
sebagai pengurangnya. Jumlah keseluruhan pakan yang dikonsumsi pada
setiap unit percobaan selama 60 hari dicatat sebagai data jumlah konsumsi
pakan.
Retensi Protein
Nilai retensi protein dihitung berdasarkan persamaan Takeuchi (1988)
sebagai berikut :
RP (%) = [(F-I)/P] x 100%
Keterangan :
RP = retensi protein (%)
F = jumlah protein tubuh ikan pada akhir penelitian (g) I = jumlah protein tubuh ikan pada awal penelitian (g) P = jumlah protein yang dikonsumsi ikan (g)
Efisiensi Pakan
Perhitungan efisiensi pakan didasarkan pada NRC (1977), yaitu besarnya
rasio perbandingan antara pertambahan bobot ikan yang didapatkan
dengan jumlah pakan yang dikonsumsi ikan. Semakin besar nilai
pertambahan bobot maka efisiensi pakan semakin besar.
EP (%) = (Wt + D) – Wo x 100 JKP
Keterangan :
EP = efisiensi pakan (%)
xlvi JKP = Jumlah pakan yang diberikan selama penelitian (g)
Tingkat Kelangsungan Hidup
Tingkat kelangsungan hidup dihitung berdasarkan persamaan yang
dikemukakan oleh Huisman (1987) yaitu:
SR (%) = Nt x 100% No
Keterangan;
Nt = jumlah ikan pada akhir penelitian (ekor) No = jumlah ikan pada awal penelitian (ekor)
Uji Kecernaan Protein dan Total Pakan
Pengujian daya cerna pakan oleh ikan nila dilakukan secara terpisah dari
uji pertumbuhan. Hal ini dimaksudkan agar kegiatan pengumpulan feses tidak
mengganggu pertumbuhan ikan uji. Akuarium yang digunakan untuk uji
kecernaan berukuran lebih besar, yaitu 100 x 60 x 50 cm.
Pembuatan pakan percobaan untuk uji kecernaan dilakukan sama seperti
pakan untuk uji pertumbuhan, namun ditambahkan 0.6% Cr2O3 sebagai indikator
kecernaan. Pakan diberikan pada ikan selama 3 minggu dan pengumpulan feses
mulai dilakukan pada hari ketujuh dengan cara menyedot feses di dasar akuarium
dengan selang kecil dan ditampung di ember. Selanjutnya feses yang mengendap
di dasar ember disaring dan dikumpulkan dalam botol film. Feses yang terkumpul
dikeringkan dalam oven bersuhu 110 selama 4-6 jam, dan dianalisis kandungan
Cr2O3 (Lampiran 7) dan kadar proteinnya.
Penghitungan nilai kecernaan berdasarkan Takeuchi (1988):
KT = 100 x (1 – b/b’)
KP = 100 x [1 – (a’/a x b/b’)]
Keterangan :
KT = kecernaan total (%) KP = kecernaan protein (%)
a = kadar nutrien (protein) dalam pakan (bobot kering) a’ = kadar nutrien (protein) dalam feses (bobot kering) b = kadar indikator Cr2O3 dalam pakan (% bobot kering) b’ = kadar indikator Cr2O3 dalam feses (% bobot kering)
xlvii Data hasil isolasi dan seleksi bakteri proteolitik, data aktivitas enzim
protease, kerapatan optis dan uji derajat hidrolisis pakan dianalisis secara
deskriptif. Data hasil uji pertumbuhan dan kecernaan pakan dianalisis secara
statistika dengan Anova dan uji Duncan dengan selang kepercayaan 95%.
HASIL DAN PEMBAHASAN
Isolasi dan Seleksi Bakteri Proteolitik
Isolasi Bakteri Proteolitik
Ikan lele yang digunakan untuk isolasi bakteri berasal dari petani lele di
daerah Cibalagung Bogor, sebanyak 4 ekor. Saluran pencernaan yang didapatkan
dari masing-masing ikan digerus menjadi satu sebelum dilakukan kegiatan isolasi.
Isolasi juga dilakukan terhadap sampel air kolam pemeliharaan lele dari tempat
yang sama sebagai pembanding.
Kegiatan isolasi bakteri dari saluran pencernaan ikan lele dan kolam
pemeliharaan ikan berturut-turut mendapatkan 7 dan 3 isolat yang dipilih
berdasarkan perbedaan morfologi koloninya. Koloni-koloni bakteri yang
didapatkan memiliki karakter morfologis antara lain bulat kecil, bulat sedang,
bulat besar, dan warna yang didapatkan adalah putih kusam, putih susu, krem,
kuning, dan bening.
Seleksi Bakteri Proteolitik Aktivitas Proteolitik
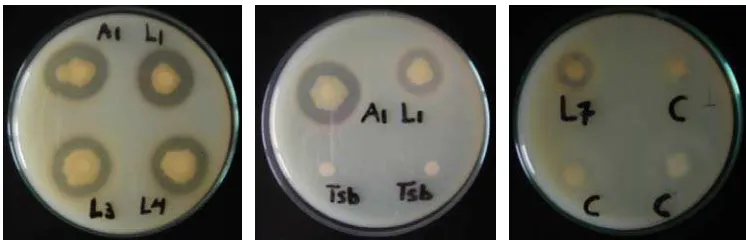
Hasil uji aktivitas proteolitik bakteri hasil isolasi disajikan pada Gambar 1.
Aktivitas proteolitik positif ditandai dengan adanya zona bening di sekeliling
koloni isolat, yang merupakan luas areal hidrolisis substrat kasein oleh isolat
bakteri.
[image:38.612.133.506.558.678.2]
xlviii Hasil pengukuran luas diameter hasil aktivitas proteolitik disajikan pada
Lampiran 8. Empat bakteri dengan diameter hidrolisis kasein tertinggi adalah A1,
L1, L4 dan L3 yang menghasilkan zona bening dengan diameter berturut-turut
sebesar 30, 28, 28 dan 25 mm. Enam isolat lainnya tidak terpilih untuk mengikuti
pengujian patogenisitas karena zona hidrolisis kaseinnya terlalu sempit (11, 10, 8,
8, 6 dan 5 mm). Keempat bakteri tersebut selanjutnya mengikuti pengujian
patogenisitas.
Luas zona hidrolisis kasein dijadikan sebagai dasar acuan pertama dalam
seleksi bakteri proteolitik, yang mengindikasikan kemampuannya dalam
memanfaatkan protein untuk kelangsungan hidupnya, dengan terlebih dahulu
merombak protein menjadi asam-asam amino. Meskipun menurut Suhartono
(1989b), dalam beberapa kasus, luas zona hidrolisis kasein tidak secara otomatis
langsung berkorelasi dengan produktivitasnya dalam menghasilkan enzim
protease, namun metode ini masih cukup efektif untuk seleksi awal. Gupta dan
Khare (2006) dan Tang (2008) juga melakukan metode zona bening ini untuk
menyeleksi bakteri proteolitik, dan pada tahap terakhirnya memperoleh
Pseudomonas aeruginosa yang mampu memproduksi protease dalam jumlah besar.
Uji Patogenisitas
Uji patogenisitas bertujuan untuk mengetahui apakah isolat yang
didapatkan bersifat patogen terhadap ikan uji atau tidak. Meskipun isolat tidak
diberikan secara langsung dalam keadaan hidup pada pakan uji (hanya ekstrak
enzimnya saja), namun dikhawatirkan ekstrak protease dari bakteri patogen dapat
berefek negatif terhadap ikan uji. Pengujian dilakukan terhadap 4 bakteri yang
memiliki zona hidrolisis kasein tertinggi (A1, L1, L3 dan L4) dan larutan
fisiologis sebagai kontrol menggunakan ikan nila sebagai ikan uji.
Hasil uji patogenisitas yang dilakukan selama dua minggu di akuarium
disajikan pada Lampiran 9. Tingkat kelulusan hidup ikan nila yang diinjeksi isolat
A1 dan L1 sebesar 100% hingga akhir masa pengamatan, sama dengan kontrol,
yang mengindikasikan bahwa isolat A1 dan L1 tidak bersifat patogen terhadap
ikan nila. Tingkat kelangsungan hidup ikan nila pada perlakuan injeksi isolat L3
xlix Berdasarkan analisis secara deskriptif, isolat A1 dan L1 terpilih sebagai
bakteri penghasil enzim protease pada percobaan selanjutnya. Pertimbangannya
adalah berdasarkan peringkatnya yang tertinggi pada uji zona hidrolisis kasein,
dan terbukti tidak bersifat patogen pada ikan nila.
Optimasi Potensi Bakteri Terpilih sebagai Sumber Enzim Protease Kerapatan Optis Bakteri A1 dan L1
Nilai kerapatan optis mencerminkan kepadatan populasi bakteri di dalam
cairan kultur. Meskipun nilai kerapatan optis ini tidak bisa membedakan antara
populasi bakteri yang hidup dengan yang mati, namun metode ini cukup efektif
menggambarkan dinamika pertumbuhan bakteri. Hasil pengamatan nilai kerapatan
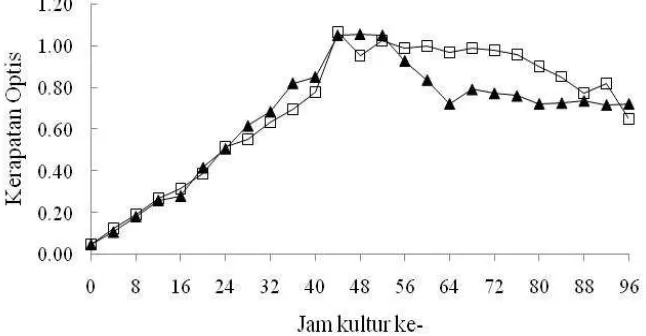
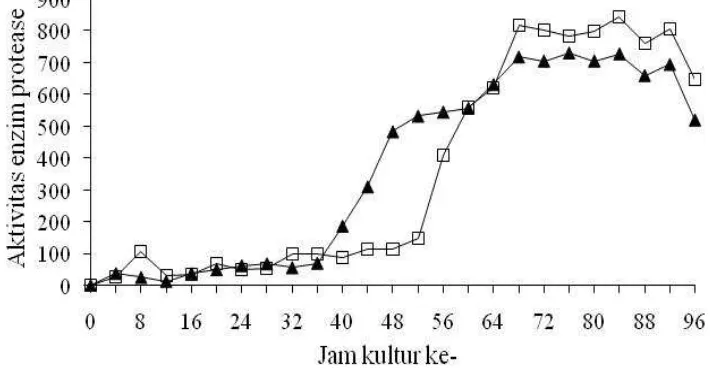
optis bakteri A1 dan L1 setiap 4 jam selama empat hari tersaji di Gambar 2 dan
Lampiran 10. Kurva pertumbuhan pada Gambar 2 memperlihatkan bahwa kedua
bakteri mengalami fase-fase pertumbuhannya pada waktu yang hampir
[image:40.612.159.482.368.536.2]bersamaan.
Gambar 2. Kerapatan optis cairan kultur bakteri A1 (▲) dan L1 (□) dalam pengamatan selama 4 hari pada waktu kulturnya.
Fase pertumbuhan awal (lag phase) dan fase eksponensial bakteri A1 dan L1 dimulai dari awal kultur hingga jam ke-44 (mendekati 2 hari). Fase stasioner
bakteri A1 terjadi antara jam ke-44 sampai jam ke-52, sedangkan bakteri L1
antara jam ke-44 sampai jam ke-68. Fase kematian terjadi setelah stasioner, di
mana populasi bakteri cenderung terus mengalami penurunan.
Menurut Pelczar dan Chan (1986), fase pertumbuhan bakteri terdiri dari
l pertumbuhan yang cepat (fase logaritma atau eksponensial), kemudian mendatar
(fase statis atau stasioner) dan akhirnya fase penurunan populasi sel-sel hidup
(fase kematian atau penurunan). Penentuan fase pertumbuhan bermanfaat untuk
mengetahui kapan waktu panen sel yang tepat untuk memproduksi suatu produk
atau senyawa metabolit, antara lain enzim (Suhartono 1989b).
Pemantauan Aktivitas Enzim Protease Bakteri A1 dan L1
Kegiatan ekstraksi enzim yang dilanjutkan dengan pengujian aktivitas
enzim protease bakteri A1 dan L1, dilakukan selama 4 hari setiap 4 jam sekali
pada waktu kulturnya, sebanyak 25 titik pengamatan (Gambar 3 dan Lampiran
11). Kegiatan ini dimaksudkan untuk mengetahui adanya fluktuasi produksi enzim
protease oleh kedua bakteri, karena daya sintesis enzim oleh bakteri dari satu fase
[image:41.612.141.496.338.524.2]ke fase lain tidak sama.
Gambar 3. Aktivitas enzim protease (µg/menit.ml) bakteri A1 (▲) dan L1 (□) dalam pengamatan selama 4 hari pada waktu kulturnya.
Dari Gambar 3 terlihat bahwa aktivitas enzim protease bakteri A1 mulai
mengalami peningkatan pada titik waktu ke-11 (atau jam ke-40) dan semakin
meningkat pada titik waktu berikutnya, sedangkan pada bakteri L1, peningkatan
yang signifikan dimulai pada titik waktu ke-14 (atau jam ke-52). Bakteri A1 mulai
meningkatkan sekresi enzim proteasenya menjelang memasuki fase stasioner,
sedangkan bakteri L1 di dalam fase stasioner. Enzim dari kedua bakteri
li bersamaan, yaitu pada titik waktu 18 hingga 24 (atau jam 68 hingga
ke-92). Tingkat produksi tertinggi ini justru dihasilkan ketika kedua bakteri berada di
dalam fase penurunan.
Menurut Suhartono (1989a), sintesis enzim ekstraselular oleh bakteri
dalam jumlah terbesar secara normal terjadi pada saat sebelum sporulasi, yaitu
pada akhir fase eksponensial dan awal stasioner. Keadaan ini membawa pada
suatu dugaan bahwa kemungkinan ada hubungan sebab akibat antara eksoenzim
dan sporulasi. Bacilus sp misalnya, memproduksi protease serin pada tahap akhir pertumbuhan, yaitu saat sel memasuki fase sporulasi, dan Bacillus subtilis meningkatkan produksi protease jenis subtilisin pada saat menjalani proses
sporulasi.
Berdasarkan hasil pengamatan pada tahap ini, maka diputuskan bahwa
pemanenan enzim protease untuk bakteri A1 dan L1 dilakukan pada umur kultur 3
hari (72 jam), yaitu pada posisi aman di dalam rentangan masa produksi enzim
protease yang tertinggi. Hasil ini serupa dengan Wang et al. (2008), yang melaporkan bahwa kondisi produksi protease yang optimal oleh bakteri
Chryseobacterium taeanense TKU001 adalah dengan inkubasi kultur bakteri tersebut pada suhu 37oC selama 3 hari. Tang (2008) yang mengisolasi bakteri penghasil protease dan mengoptimasi produksi protease oleh Pseudomonas aeruginosa juga melaporkan hal yang sama. Mabrouk et al. (1999) yang mengoptimasi produksi protease Bacillus licheniformis melaporkan hal yang berbeda, bahwa waktu inkubasi selama 5 hari pada suhu 37oC menghasilkan protease dengan aktivitas yang maksimal.
Penentuan Dosis Enzim Protease Bakteri
Setelah mendapatkan waktu kultur bakteri yang optimal untuk memanen
enzim protease, tahap selanjutnya adalah menentukan dosis enzim protease yang
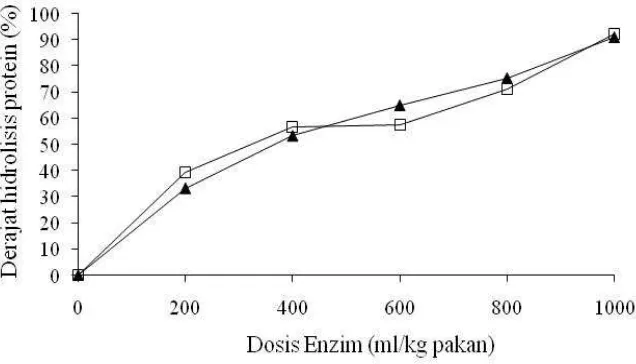
tepat untuk menghidrolisis protein pakan percobaan. Hasil uji derajat hidrolisis
protein pakan formulasi yang diberi ekstrak enzim protease bakteri A1 dan L1
lii Gambar 4. Nilai derajat hidrolisis protein pakan (%) pada beberapa level dosis
enzim protease bakteri A1 (▲) dan L1 (□).
Dari Gambar 4 terlihat bahwa derajat hidrolisis protein pakan percobaan
meningkat bersamaan dengan meningkatnya dosis enzim protease yang diberikan.
Pada dosis 1000 ml enzim / kg pakan, derajat hidrolisis protein mencapai rataan
91.99% (±0.65). Ini mengandung pengertian bahwa dari 100 gram protein yang
terkandung di dalam pakan, sebanyak 91.99 gram telah diubah menjadi bentuk
protein terlarut, dan sisanya (8.01 gram) masih berupa protein tidak larut. Dosis
1000 ml ekstrak enzim protease / kg pakan terpilih untuk diaplikasikan dalam
percobaan selanjutnya.
Dosis 1000 ml enzim protease perkilogram pakan dalam penelitian ini
setara dengan memberikan enzim protease dengan total aktivitas 8 x 105 µg perkilogram pakan. Hou et al. (2010) memberikan kombinasi enzim-enzim protease untuk menghidrolisis tulang rangka limbah ikan Pollock sebanyak 1.2
g/kg substrat dengan aktivitas 1.2 x 105 µg/g atau setara dengan 1.44 x 105 µg/kg substrat, dan berhasil memecahkan ikatan peptida pada substrat sebanyak 25%.
Beal et al. (1998c) menghidrolisis tepung bungkil kedelai (TBK) dengan enzim protease sebanyak 2.5 g/ kg TBK dengan aktivitas 4 x 105 µg/g atau setara dengan 10 x 105 µg/kg TBK, dan melaporkan adanya penurunan jumlah dan densitas ikatan protein yang mengindikasikan adanya hidrolisis protein TBK.
Tingginya derajat hidrolisis protein pada pakan percobaan yang
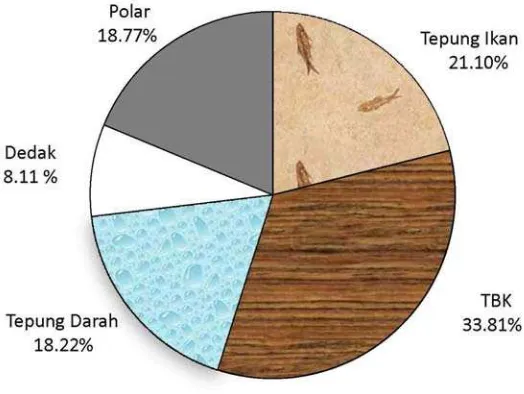
liii kontribusi beberapa bahan baku pakannya. Protein yang terdapat di dalam pakan
formulasi sebesar 31.12 % bobot kering pakan berasal dari sumbangan TBK
sebesar 10.52%, tepung ikan 6.56%, polar 5.84%, tepung darah 5.67% dan dedak
2.52%. Prosentase sumbangan protein dari masing-masing bahan baku
[image:44.612.189.451.213.410.2]dibandingkan dengan total protein pakan ditampilkan pada Gambar 5.
Gambar 5. Prosentase sumbangan protein masing-masing bahan baku terhadap total protein pakan formulasi (%).
TBK, tepung ikan dan tepung darah sebagai tiga komponen terbesar
penyumbang protein dalam pakan formulasi akan mengalami perombakan protein
yang signifikan bila dihidrolisis oleh enzim protease. Hou et al. (2010) yang menggunakan kompleks enzim protease untuk menghidrolisis limbah ikan Pollock
Alaska yang terdiri dari daging dan tulang buangan melaporkan adanya penurunan
signifikan jumlah ikatan peptida. Beal et al. (1998a) menggunakan teknik in vitro untuk mengevaluasi kemampuan beberapa jenis enzim protease dalam
meningkatkan kecernaan nitrogen TBK, dan menemukan peningkatan sebesar
12% daripada kontrolnya. Rooke et al. (1998) melaporkan adanya peningkatan konsentrasi asam amino TBK setelah ditreatment dengan enzim protease. TBK
dalam pakan percobaan yang merupakan penyumbang terbesar dalam komposisi
protein pakan akan secara signifikan menambah jumlah protein yang terhidrolisis.
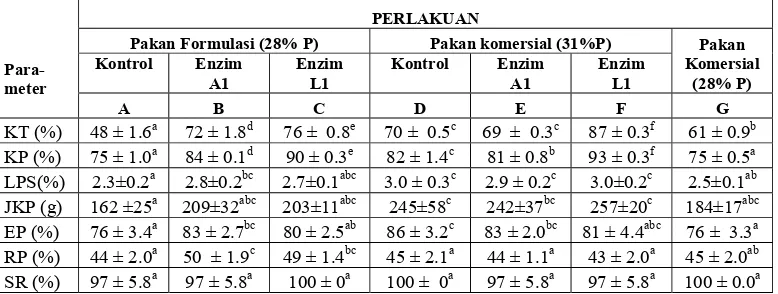
liv Hasil uji kecernaan total dan protein pakan untuk ikan nila disajikan di
Tabel 7, sedangkan perhitungan nilai kecernaan pakan oleh ikan nila, analisis
ragam dan uji Duncannya berturut-turut tersaji di Lampiran 13, 18 dan 19. Pakan
formulasi yang ditambah enzim A1 (pakan B) dan L1 (pakan C) meningkat nilai
kecernaan protein dan totalnya secara signifikan dibandingkan kontrolnya (pakan
A), dengan nilai kecernaan total 48, 72 dan 76%, dan kecernaan protein pakan 75,
84 dan 90%, berturut-turut untuk pakan A, B dan C. Pakan B dan C telah
mengalami proses pencernaan awal dengan cukup baik, sehingga jumlah nutrien
terhidrolisisnya lebih banyak dibandingkan pakan A, dan kecernaannya
[image:45.612.119.508.341.488.2]meningkat.
Tabel 7. Laju pertumbuhan spesifik (LPS), jumlah konsumsi pakan (JKP), efisiensi pakan (EP), retensi protein (RP), kecernaan protein pakan (KP), kecernaan total pakan (KT) dan kelangsungan hidup (SR) ikan nila.
Para-meter
PERLAKUAN
Pakan Formulasi (28% P) Pakan komersial (31%P) Pakan Komersial
(28% P) Kontrol Enzim
A1
Enzim L1
Kontrol Enzim A1
Enzim L1
A B C D E F G