POTENSI CENDAWAN ENDOFIT DALAM MENGENDALIKAN Fusarium oxysporum f.sp. cubense DAN NEMATODA Radopholus similis Cobb.
PADA TANAMAN PISANG BARANGAN (Musa paradisiaca) DI RUMAH KACA
SKRIPSI
OLEH : RIZKI T SINAGA
080302045
ILMU HAMA DAN PENYAKIT TUMBUHAN
PROGRAM STUDI AGROEKOTEKNOLOGI FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA MEDAN
POTENSI CENDAWAN ENDOFIT DALAM MENGENDALIKAN Fusarium oxysporum f.sp. cubense DAN NEMATODA Radopholus similis Cobb.
PADA TANAMAN PISANG BARANGAN (Musa paradisiaca) DI RUMAH KACA
SKRIPSI
OLEH :
RIZKI T SINAGA 080302045
ILMU HAMA DAN PENYAKIT TUMBUHAN
Skripsi Sebagai Salah Satu Syarat Untuk Dapat Meraih Gelar Sarjana Pertanian di Program Studi Agroekoteknologi Fakultas Pertanian Universitas Sumatera Utara,
Medan
Disetujui Oleh Komisi Pembimbing :
Ketua Komisi Pembimbing Anggota Komisi Pembimbing
Dr. Lisnawita, SP, M.Si
NIP.19691005 199403 2 001 NIP. 19530129197903100
Ir. Mukhtar Iskandar Pinem, M.Agr
PROGRAM STUDI AGROEKOTEKNOLOGI FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA MEDAN
ABSTRACT
Rizki Trianno Sinaga. 2013. Potential of endophytic fungal to control Fusarium oxysporum f.sp. cubense (Foc) and nematode Radopholus similis (Rs) on
banana cv. Barangan (Musa paradisiaca) crop in greenhouse. Under supervised Lisnawita and Mukhtar Iskandar Pinem. This research aims to intens and find potential endophytic as biocontrol agents to control Foc and Rs on banana cv. Barangan crop. This research used Completely Randomized Design (CRD) nonfactorial consist thirthteen treatments and three replications; control (without inoculated), inoculated 300 Rs, inoculated Foc, inoculated 300 Rs one week later inoculated Foc two weeks later inoculated endophytic 5BSBH’s isolate, inoculated 300 Rs one week later inoculated Foc two weeks later inoculated endophytic 4BSU’s isolate, inoculated 300
Rs one week later inoculated Foc two weeks later inoculated endophytic 1BJH’s isolate, inoculated 300 Rs one week later inoculated Foc two weeks later inoculated endophytic 1BTAH’s isolate, inoculated 300 Rs one week later inoculated Foc two weeks later inoculated endophytic 1BSHT’s isolate, inoculated endophytic 5BSBH’s isolate one week later inoculated 300 Rs two weeks later inoculated Foc, inoculated endophytic 4BSU’s isolate one week later inoculated 300 Rs two weeks later inoculated Foc, inoculated endophytic 1BJH’s isolate one week later inoculated 300 Rs two weeks later inoculated Foc, inoculated endophytic 5BSBH’s isolate one week later inoculated 300
Rs two weeks later inoculated Foc, inoculated endophytic 1BSHT’s isolate one week later inoculated 300 Rs two weeks later inoculated Foc. The results showed that significant difference on treatment with control, where as each of treatment showed 1BJH’s isolate had the potency as biocontrol agent with to pressed percentage of root lesion as 84%.
Keywords: Endophytic fungal, Fusarium oxysporum f.sp. cubense, nematode
ABSTRAK
Rizki Trianno Sinaga. 2013. Potensi cendawan endofit dalam mengendalikan Fusarium oxysporum f.sp. cubense (Foc) dan Nematoda Radopholus similis (Rs) pada tanaman pisang Barangan (Musa paradisiaca) di rumah kaca. Dibawah bimbingan Lisnawita dan Mukhtar Iskandar Pinem. Penelitian ini bertujuan untuk mendapatkan cendawan endofit yang berpotensi sebagai agens biokontrol untuk mengendalikan populasi Rs dan Foc pada tanaman pisang Barangan. Penelitian menggunakan rancangan acak lengkap nonfaktorial dengan 13 perlakuan yang diulang sebanyak 3 kali, yaitu kontrol (tanpa inokulasi), inokulasi 300 Rs,
inokulasi Foc, inokulasi 300 Rs seminggu kemudian inokulasi Foc dua minggu kemudian inokulasi endofit isolat 5BSBH, inokulasi 300 Rs seminggu kemudian inokulasi Foc dua minggu kemudian inokulasi endofit isolat 4BSU, inokulasi 300 Rs
seminggu kemudian inokulasi Foc dua minggu kemudian inokulasi endofit isolat 1BJH, inokulasi 300 Rs seminggu kemudian inokulasi Foc dua minggu kemudian inokulasi endofit isolat 1BTAH, inokulasi 300 Rs seminggu kemudian inokulasi Foc dua minggu kemudian inokulasi endofit isolat 1BSHT, inokulasi endofit isolat 5BSBH seminggu kemudian inokulasi 300 Rs dua minggu kemudian inokulasi Foc, inokulasi endofit isolat 4BSU seminggu kemudian inokulasi 300 Rs dua minggu kemudian inokulasi Foc,
Kata kunci: cendawan endofit, Fusarium oxysporum f.sp. cubense, nematoda
Radopholus similis.
KATA PENGANTAR
Puji dan syukur penulis panjatkan kepada Tuhan Yang Maha Esa atas rahmat-Nya sehingga penulis dapat menyelesaikan skripsi ini dengan benar.
Skripsi yang berjudul ”Potensi Cendawan Endofit dalam Mengendalikan Fusarium Oxysporum f.sp. cubense dan Nematoda Radopholus Similis Cobb. pada Tanaman Pisang Barangan (Musa paradisiaca) Di Rumah Kaca” merupakan salah satu syarat untuk dapat meraih gelar Sarjana Pertanian di Program Studi Agroekoteknologi Fakultas Pertanian Universitas Sumatera Utara, Medan
Pada kesempatan ini penulis mengucapkan terima kasih
kepada Komisi Pembimbing Dr. Lisnawita, SP, M.Si selaku Ketua dan Ir. Mukhtar Iskandar Pinem, M.Agr selaku Anggota, yang telah memberikan bimbingan
kepada penulis dalam menyelesaikan skripsi ini.
Penulis menyadari masih banyak kesalahan dalam penyusunan skripsi ini. Oleh karena itu, penulis mengharapkan saran dan kritik yang bersifat membangun demi penyempurnaan skripsi ini. Akhir kata penulis mengucapkan terima kasih.
Penulis
DAFTAR ISI
Halaman
ABSTRACT ... i
ABSTRAK ... ii
RIWAYAT HIDUP ... iii
KATA PENGANTAR ... v
DAFTAR ISI ... vi
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... ix
DAFTAR LAMPIRAN ... x
PENDAHULUAN Latar belakang ... 1
Tujuan penelitian ... 3
Hipotesis penelitian ... 3
TINJAUAN PUSTAKA
Fusarium oxysporum f.sp. cubense ... 4
Biologi cendawan ... 4
Infeksi, epidemiologi penyakit dan siklus hidup ... 6
Gejala serangan ... 7
Faktor yang mempengaruhi perkembangan penyakit ... 9
Radopholus similis Cobb. Klasifikasi nematoda Radopholus similis Cobb. ... 10
Morfologi ... 10
Betina ... 10
Jantan ... 11
Biologi dan siklus hidup ... 11
Gejala serangan ... 13
Pengelolaan penyakit ... 15
Cendawan endofit ... 16
Potensi cendawan endofit dalam mengendalikan penyakit layu Fusarium dan nematoda R. similis pada tanaman pisang.. ... 17
BAHAN DAN METODE Tempat dan waktu penelitian ... 19
Bahan dan alat penelitian ... 19
Metode penelitian ... 19
Pelaksanaan penelitian ... 21
Persiapan tanaman inang ... 21
Perbanyakan cendawan endofit pada media jagung . ... 22
Pembuatan media wortel steril ... 22
Perbanyakan Radopholus similis ... 22
Perbanyakan cendawan Fusarium oxysporum f.sp.cubense ... 23
Aplikasi perlakuan ... 23
Aplikasi Radopholus similis ... 23
Aplikasi cendawan endofit ... 24
Aplikasi Fusarium oxysporum f.sp.cubense ... 24
Pemeliharaan tanaman ... 24
Peubah amatan ... 24
Periode inkubasi (hsi) ... 24
Kejadian penyakit (%) ... 24
Keparahan penyakit (%) ... 24
Persentase nekrosis jaringan akar (%) ... 25
Persentase akar mati (%) ... 26
Berat basah akar (g) ... 26
Populasi akhir nematoda R. similis (ji) ... 26
HASIL DAN PEMBAHASAN Periode inkubasi (hsi) dan Kejadian penyakit (%) ... 27
Keparahan penyakit (%) ... 30
Nekrosis jaringan akar (%) ... 32
Akar mati (%) ... 34
Berat basah akar (g) ... 36
Populasi akhir nematoda R. similis (ji) ... 38
Kesimpulan ... 41
1 Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap periode inkubasi (hsi) dan kejadian penyakit pada 60 hsi (%) ...
27
2 Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap tingkat keparahan penyakit (%) ...
30
3 Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap nekrosis jaringan akar (%) ...
33
4 Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap persentase akar mati (%) ...
35
5 Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap berat basah akar (g) ...
37
6 Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap populasi akhir nematoda R. similis (ji) ...
38
PENDAHULUAN
ABSTRACT
Rizki Trianno Sinaga. 2013. Potential of endophytic fungal to control Fusarium oxysporum f.sp. cubense (Foc) and nematode Radopholus similis (Rs) on
banana cv. Barangan (Musa paradisiaca) crop in greenhouse. Under supervised Lisnawita and Mukhtar Iskandar Pinem. This research aims to intens and find potential endophytic as biocontrol agents to control Foc and Rs on banana cv. Barangan crop. This research used Completely Randomized Design (CRD) nonfactorial consist thirthteen treatments and three replications; control (without inoculated), inoculated 300 Rs, inoculated Foc, inoculated 300 Rs one week later inoculated Foc two weeks later inoculated endophytic 5BSBH’s isolate, inoculated 300 Rs one week later inoculated Foc two weeks later inoculated endophytic 4BSU’s isolate, inoculated 300
Rs one week later inoculated Foc two weeks later inoculated endophytic 1BJH’s isolate, inoculated 300 Rs one week later inoculated Foc two weeks later inoculated endophytic 1BTAH’s isolate, inoculated 300 Rs one week later inoculated Foc two weeks later inoculated endophytic 1BSHT’s isolate, inoculated endophytic 5BSBH’s isolate one week later inoculated 300 Rs two weeks later inoculated Foc, inoculated endophytic 4BSU’s isolate one week later inoculated 300 Rs two weeks later inoculated Foc, inoculated endophytic 1BJH’s isolate one week later inoculated 300 Rs two weeks later inoculated Foc, inoculated endophytic 5BSBH’s isolate one week later inoculated 300
Rs two weeks later inoculated Foc, inoculated endophytic 1BSHT’s isolate one week later inoculated 300 Rs two weeks later inoculated Foc. The results showed that significant difference on treatment with control, where as each of treatment showed 1BJH’s isolate had the potency as biocontrol agent with to pressed percentage of root lesion as 84%.
Keywords: Endophytic fungal, Fusarium oxysporum f.sp. cubense, nematode
ABSTRAK
Rizki Trianno Sinaga. 2013. Potensi cendawan endofit dalam mengendalikan Fusarium oxysporum f.sp. cubense (Foc) dan Nematoda Radopholus similis (Rs) pada tanaman pisang Barangan (Musa paradisiaca) di rumah kaca. Dibawah bimbingan Lisnawita dan Mukhtar Iskandar Pinem. Penelitian ini bertujuan untuk mendapatkan cendawan endofit yang berpotensi sebagai agens biokontrol untuk mengendalikan populasi Rs dan Foc pada tanaman pisang Barangan. Penelitian menggunakan rancangan acak lengkap nonfaktorial dengan 13 perlakuan yang diulang sebanyak 3 kali, yaitu kontrol (tanpa inokulasi), inokulasi 300 Rs,
inokulasi Foc, inokulasi 300 Rs seminggu kemudian inokulasi Foc dua minggu kemudian inokulasi endofit isolat 5BSBH, inokulasi 300 Rs seminggu kemudian inokulasi Foc dua minggu kemudian inokulasi endofit isolat 4BSU, inokulasi 300 Rs
seminggu kemudian inokulasi Foc dua minggu kemudian inokulasi endofit isolat 1BJH, inokulasi 300 Rs seminggu kemudian inokulasi Foc dua minggu kemudian inokulasi endofit isolat 1BTAH, inokulasi 300 Rs seminggu kemudian inokulasi Foc dua minggu kemudian inokulasi endofit isolat 1BSHT, inokulasi endofit isolat 5BSBH seminggu kemudian inokulasi 300 Rs dua minggu kemudian inokulasi Foc, inokulasi endofit isolat 4BSU seminggu kemudian inokulasi 300 Rs dua minggu kemudian inokulasi Foc,
Kawasan Asia Tenggara dikenal sebagai pusat asal mula Genus Musa. Lebih dari 60 spesies Musa dilaporkan berada di kawasan ini. Sebagai salah satu daerah di Asia Tenggara, Indonesia diperkirakan memiliki 200 kultivar pisang. Tumbuh di berbagai tipe tanah serta dimanfaatkan untuk berbagai macam kepentingan. Dari areal pertanaman pisang di dunia, Indonesia memiliki hampir semua jenis organisme pengganggu tumbuhan (OPT) pisang (Nazir dan Jumjunidang, 2003).
Pisang (Musa sp.) merupakan salah satu komoditas buah-buahan penting di Indonesia yang diusahakan secara meluas dari dataran rendah sampai dataran tinggi (Lisnawita et al. 1998). Di Sumatera Utara produksi pisang sejak tahun 2004-2009 cenderung mengalami peningkatan dengan rata-rata 7,5% per tahun. Namun pada tahun 2010, produksi pisang mengalami penurunan sebesar 9,7% dari tahun 2009. Salah satu penyebabnya adalah gangguan OPT. Salah satu OPT yang menjadi perhatian pada tanaman pisang saat ini adalah penyakit layu Fusarium yang disebabkan oleh Fusarium oxysporum f.sp. cubense. Pada musim tanam 2011 luas serangan pada tanaman pisang Barangan di Propinsi Sumatera Utara adalah seluas 6,63 ha. (Dinas Pertanian Sumatera Utara, 2012).
Pada rizosfer tanaman pisang yang terserang layu fusarium, ditemukan beberapa jenis nematoda, antara lain Radopholus similis, Meloidogyne spp., Rotylechus reniformis, Helicotylenchus spp., dan Pratylenchus coffeae. Keberadaan nematoda tersebut menyebabkan tanaman pisang lebih rentan terhadap fusarium (Lisnawita et al., 1998).
Radopholus similis merupakan spesies nematoda utama pada tanaman pisang yang penyebarannya sangat luas. Nematoda ini juga dapat menginfeksi bonggol pisang. Kerusakan yang ditimbulkan lebih parah dibanding kerusakan yang disebabkan spesies nematoda lain. Di Indonesia, spesies ini juga ditemukan di beberapa sentra produksi pisang. Serangan nematoda ini berpotensi sebagai salah satu faktor pembatas produksi pisang (Jumjunidang, 2009).
Serangan bersama oleh patogen yang berbeda pada saat yang sama pada suatu tanaman dapat menimbulkan kerusakan yang lebih tinggi, lebih rendah atau sama bila dibandingkan dengan patogen yang menyerang secara tunggal. Hasil penelitian menunjukkan bahwa interaksi antara nematoda parasit dengan patogen tanah lainnya bersifat sinergistik. Hal tersebut karena luka yang ditimbulkan oleh penetrasi nematoda parasit pada akar dapat menjadi jalan masuk bagi patogen tanah lainnya (Blake, 1966). Infeksi dapat terjadi pada akar yang luka dan akar yang tidak luka. Infeksi nematoda R. similis dapat memicu serangan layu Fusarium (Jumjunidang et al., 2009).
menimbulkan gejala pada tanaman inang (Carrol, 1988). Sebagian besar cendawan endofit berasal dari kelas Ascomycetes, kelas Basidiomycetes dan
Deuteromycetes. Beberapa genus cendawan yang tergolong endofit adalah:
Acremonium, Aspergillus, Fusarium, Trichoderma, Gliocladium, Alternaria, Beuveria,
Penicilium, Mucor, Phyllosticta, Pestalotia dan Phoma (Carrol, 1988 dalam IPB, 2008). Tujuan penelitian
Penelitian ini bertujuan untuk mendapatkan cendawan endofit yang berpotensi sebagai agens biokontrol untuk mengendalikan populasi R. similis dan F. oxysporum f.sp. cubense pada tanaman pisang Barangan di Sumatera Utara.
Hipotesis penelitian
Terdapat beberapa cendawan endofit yang berpotensi sebagai agens biokontrol untuk mengendalikan populasi R. similis dan F. oxysporum f.sp.cubense pada tanaman pisang Barangan di Sumatera Utara.
Kegunaan penelitian
1. Sebagai Salah Satu Syarat Untuk Dapat Meraih Gelar Sarjana Pertanian di Program Studi Agroekoteknologi Fakultas Pertanian Universitas Sumatera Utara, Medan.
Kawasan Asia Tenggara dikenal sebagai pusat asal mula Genus Musa. Lebih dari 60 spesies Musa dilaporkan berada di kawasan ini. Sebagai salah satu daerah di Asia Tenggara, Indonesia diperkirakan memiliki 200 kultivar pisang. Tumbuh di berbagai tipe tanah serta dimanfaatkan untuk berbagai macam kepentingan. Dari areal pertanaman pisang di dunia, Indonesia memiliki hampir semua jenis organisme pengganggu tumbuhan (OPT) pisang (Nazir dan Jumjunidang, 2003).
Pisang (Musa sp.) merupakan salah satu komoditas buah-buahan penting di Indonesia yang diusahakan secara meluas dari dataran rendah sampai dataran tinggi (Lisnawita et al. 1998). Di Sumatera Utara produksi pisang sejak tahun 2004-2009 cenderung mengalami peningkatan dengan rata-rata 7,5% per tahun. Namun pada tahun 2010, produksi pisang mengalami penurunan sebesar 9,7% dari tahun 2009. Salah satu penyebabnya adalah gangguan OPT. Salah satu OPT yang menjadi perhatian pada tanaman pisang saat ini adalah penyakit layu Fusarium yang disebabkan oleh Fusarium oxysporum f.sp. cubense. Pada musim tanam 2011 luas serangan pada tanaman pisang Barangan di Propinsi Sumatera Utara adalah seluas 6,63 ha. (Dinas Pertanian Sumatera Utara, 2012).
Pada rizosfer tanaman pisang yang terserang layu fusarium, ditemukan beberapa jenis nematoda, antara lain Radopholus similis, Meloidogyne spp., Rotylechus reniformis, Helicotylenchus spp., dan Pratylenchus coffeae. Keberadaan nematoda tersebut menyebabkan tanaman pisang lebih rentan terhadap fusarium (Lisnawita et al., 1998).
Radopholus similis merupakan spesies nematoda utama pada tanaman pisang yang penyebarannya sangat luas. Nematoda ini juga dapat menginfeksi bonggol pisang. Kerusakan yang ditimbulkan lebih parah dibanding kerusakan yang disebabkan spesies nematoda lain. Di Indonesia, spesies ini juga ditemukan di beberapa sentra produksi pisang. Serangan nematoda ini berpotensi sebagai salah satu faktor pembatas produksi pisang (Jumjunidang, 2009).
Serangan bersama oleh patogen yang berbeda pada saat yang sama pada suatu tanaman dapat menimbulkan kerusakan yang lebih tinggi, lebih rendah atau sama bila dibandingkan dengan patogen yang menyerang secara tunggal. Hasil penelitian menunjukkan bahwa interaksi antara nematoda parasit dengan patogen tanah lainnya bersifat sinergistik. Hal tersebut karena luka yang ditimbulkan oleh penetrasi nematoda parasit pada akar dapat menjadi jalan masuk bagi patogen tanah lainnya (Blake, 1966). Infeksi dapat terjadi pada akar yang luka dan akar yang tidak luka. Infeksi nematoda R. similis dapat memicu serangan layu Fusarium (Jumjunidang et al., 2009).
menimbulkan gejala pada tanaman inang (Carrol, 1988). Sebagian besar cendawan endofit berasal dari kelas Ascomycetes, kelas Basidiomycetes dan
Deuteromycetes. Beberapa genus cendawan yang tergolong endofit adalah:
Acremonium, Aspergillus, Fusarium, Trichoderma, Gliocladium, Alternaria, Beuveria,
Penicilium, Mucor, Phyllosticta, Pestalotia dan Phoma (Carrol, 1988 dalam IPB, 2008). Tujuan penelitian
Penelitian ini bertujuan untuk mendapatkan cendawan endofit yang berpotensi sebagai agens biokontrol untuk mengendalikan populasi R. similis dan F. oxysporum f.sp. cubense pada tanaman pisang Barangan di Sumatera Utara.
Hipotesis penelitian
Terdapat beberapa cendawan endofit yang berpotensi sebagai agens biokontrol untuk mengendalikan populasi R. similis dan F. oxysporum f.sp.cubense pada tanaman pisang Barangan di Sumatera Utara.
Kegunaan penelitian
1. Sebagai Salah Satu Syarat Untuk Dapat Meraih Gelar Sarjana Pertanian di Program Studi Agroekoteknologi Fakultas Pertanian Universitas Sumatera Utara, Medan.
TINJAUAN PUSTAKA
Fusarium oxysporum f.sp. cubense Biologi cendawan
Menurut Semangun (1996) cendawan Fusarium diklasifikasikan sebagai berikut: Kingdom : Fungi
Divisio : Eumycota
Sub divisio : Deuteromycotina Class : Deuteromycetes Ordo : Moniliales Family : Tuberculariaceae Genus : Fusarium
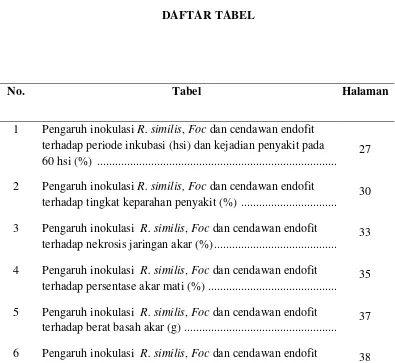
Gambar 1. Fusarium oxysporum f.sp. cubense, klamidospora (A) (http://www. Cultures of Fusarium oxysporum f.sp. cubense diakses tanggal 26 Februari 2012). Mikrokonidia (B) dan makrokonidia Fusarium oxysporum
f.sp. cubense (C) (Perbesaran objek 400X).
Makrokonidium berbentuk sabit, bertangkai kecil, kebanyakan bersel 4, berukuran 22-36 x 4-5 μm (Gambar 1C). Klamidospora bersel satu, jorong atau bulat, berukuran 7-13 x 7-8 μm, terbentuk di tengah hifa atau pada makrokonidium, seringkali berpasangan (Gambar 1A) (Wardlaw, 1972).
Di alam cendawan membentuk konidium. Konidiofor bercabang-cabang dan makrokonidium berbentuk sabit, bertangkai kecil, seringkali berpasangan. Miselium terutama terdapat di dalam sel khususnya di dalam pembuluh kayu, juga membentuk miselium yang terdapat di antara sel-sel, yaitu di dalam kulit dan jaringan parenkim di dekat terjadinya infeksi. Inokulum Foc terdiri atas makrokonidia, mikrokonidia, klamidospora dan miselia. Cendawan dapat bertahan lama di dalam tanah selama beberapa tahun. Populasi patogen dapat bertahan secara alami di dalam tanah dan pada akar-akar tanaman sakit. Apabila terdapat tanaman peka, melalui akar yang luka dapat segera menimbulkan infeksi (Departemen Pertanian, 2012).
Infeksi, epidemiologi penyakit dan siklus hidup
B
Fusarium oxysporum f.sp. cubense yang terdapat disuatu daerah dapat disebabkan oleh penyaluran tanaman yang terinfeksi saat manusia beraktifitas. Patogen dapat berpindah dari jaringan tanaman yang sakit sebagai klamidospora yang dirangsang berkecambah oleh inang atau kontak dengan jaringan sehat tanaman rentan. Miselia dan konidia hasil perkecambahan klamidospora yang diproduksi setelah 6-8 jam, 2-3 hari kemudian akan menginfeksi akar sekunder atau tersier. Patogen masuk ke zona vaskular dari rimpang akar tanaman yang sakit, kemudian bergerak keluar dari sistem vaskular masuk ke sel parenkim. Selanjutnya konidia terbentuk dan klamidospora terbentuk di dalam tanah ketika tanaman mati, berlangsung aktif sampai beberapa tahun (Perez dan Vicente, 2004).
Fusarium oxysporum f.sp. cubense termasuk cendawan imperfektif, awalnya bernama F. cubense diisolasi oleh E. F. Smith (1910) dari bonggol terinfeksi Foc di Cuba, Jamaica. Di alam, cendawan ini memiliki konidium pada suatu badan yang diketahui dengan nama sporodokium. Sporodokium muncul dari tangkai dan permukaan daun pada tanaman yang terinfeksi. Sporodokium ini keluar melalui stomata yang terbuka, paling banyak pada epidermis bagian atas tangkai daun. Konidia muncul di ujung hifa, baik ujung maupun lateral dan terdiri atas dua jenis yang berbeda. Pertama, septa mikrokonidia yang muncul dari kumpulan konidia berbentuk bulat telur atau agak memanjang dengan ukuran 5-7x2,5-3 µm. Kedua, konidia berbentuk sabit, ukuran bervariasi dan sering ditemukan diselingi oleh beberapa mikrokonidia (Wardlaw, 1972).
vegetatif, namun meningkatkan pertumbuhan saprofit disisa-sisa tanaman dan berlangsung dalam tanah selama bertahun-tahun. Fusarium oxysporum
f.sp. cubense umumnya terdapat pada jaringan xilem. Parenkim yang mengelilingi jaringan vaskular akan mati, sebelumnya terjadi invasi cendawan dan selanjutnya berlangsung dalam lumen sel (Perez dan Vicente, 2004).
Vivotoksin merupakan zat yang dihasilkan tanaman inang yang terinfeksi Foc. Toksin tersebut bekerjasama dengan asam Fusarik dalam menyebabkan penyakit (Varma, 2012).
Gejala serangan
Gambar 2. Gejala serangan Foc pada permukaan daun (a), gejala pada pangkal batang (b), dan akar busuk (c).
Tanaman yang terserang tidak akan mampu berbuah atau buahnya tidak terisi. Lamanya waktu antara saat terjadinya infeksi penyakit sampai munculnya gejala penyakit
berlangsung kurang lebih 2 bula
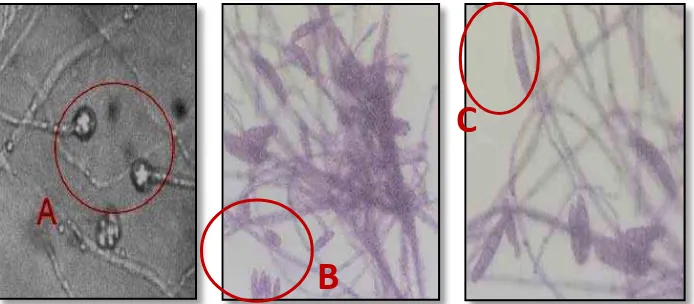
Mekanisme kelayuan pada tanaman disebabkan oleh cendawan yang dapat hidup di dalam tanah dapat menyerang tanaman. Selanjutnya berpenetrasi ke dalam akar, dari akar cendawan tumbuh dan berkembang hingga mencapai bonggol pisang (Gambar 3a, tanda ).
Gambar 3. Foc berpenetrasi kedalam akar (a), dan Foc berkolonisasi (b). (Perbesaran objek 400X).
Di dalam bonggol dan pembuluh xilem cendawan ini berkolonisasi dan menginfestasi secara cepat (Gambar 3b, tanda ). Akibatnya akar tanaman dan bonggol serta pembuluhnya terinfeksi Foc. Infeksi Foc pada tanaman pisang, akan menganggu
a b c
proses penyerapan, transportasi air dan zat makanan di dalam tanah, sehingga tanaman menjadi layu dan akhirnya mati (Maimunah, 1999).
Gejala yang paling khas adalah gejala dalam pangkal batang (pseudostem). Jika pangkal batang dibelah membujur, terlihat garis-garis coklat atau hitam menuju ke semua arah, dari batang (bonggol) ke atas melalui jaringan pembuluh ke pangkal daun dan tangkai. Perubahan warna pada berkas pembuluh paling jelas tampak pada batang. Berkas pembuluh akar biasanya tidak berubah warna, namun sering sekali akar tanaman sakit berwarna hitam dan membusuk (Semangun, 1996).
Keparahan penyakit yang ditimbulkan serangan Foc pada tanaman pisang Barangan berhubungan dengan virulensi patogen. Variasi virulensi selain disebabkan oleh variasi yang terjadi saat reproduksi aseksual cendawan, juga disebabkan oleh perbedaan produksi toksin dari masing-masing isolat. Fusarium oxysporum
f.sp. cubense dapat memproduksi asam fusarat, toksin ini mempengaruhi mitokondria, menghambat enzim katalase serta mempengaruhi sel yang mengakibatkan kebocoran ion dan kematian sel (Balio, 1981 dalam Jumjunidang et al.2011).
Faktor yang mempengaruhi perkembangan penyakit
Varietas rentan terhadap Foc seperti pisang Barangan yang telah diuji oleh Jumjunidang et al. (2011) memiliki masa inkubasi penyakit paling cepat, sering tidak memperlihatkan perbedaan yang nyata antar isolat dengan masa inkubasi 13-80 hari setelah inokulasi. Persentase serangan yang diakibatkannya mencapai 93,33%. Perbedaan virulensi pada isolat uji tersebut dapat disebabkan oleh perbedaan karakter biologi, kimia, dan genetik dari isolat cendawan. (Jumjunidang et al., 2011).
Radopholus similis Cobb.
Klasifikasi nematoda Radopholus similis Cobb.
Menurut Dropkin (1992) nematoda Radopholus similis Cobb. diklasifikasikan sebagai berikut:
Kingdom : Animalia Phylum : Aschelminthes Class : Nematoda Ordo : Tylenchida Family : Pratylenchidae Genus : Radopholus
Species : Radopholus similis Cobb. Morfologi
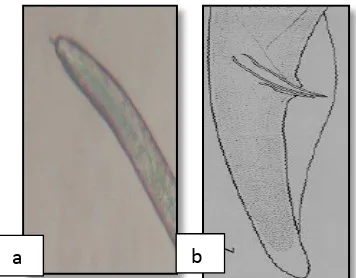
Betina
Jantan
Panjang nematoda jantan rata-rata 0,58 mm dan mengalami degenerasi, esofagus dan stiletnya tidak berkembang sempurna. Kepala nematoda jantan berbentuk membulat dan berlekuk yang sangat berbeda dengan betina. Mempunyai testis tunggal (Gambar 4b) dan bursa meluas sampai dua per tiga ekor (Dropkin,1992).
Gambar 4. Nematoda R.similis betina (a) (Perbesaran 100X), nematoda R. similis jantan (b) (Dropkin, 1992).
Biologi dan siklus hidup
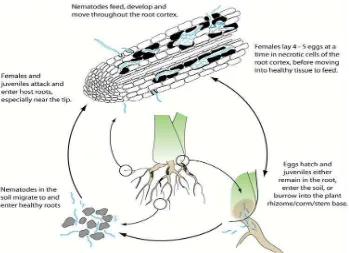
Radopholus similis adalah spesies nematoda endoparasitik yang berpindah-pindah yang mampu menyelesaikan daur hidupnya di dalam jaringan korteks akar. Nematoda betina bertelur untuk menghasilkan populasi baru selama melakukan perpindahan. Nematoda ini ditemukan pada semua tingkatan perkembangan akar tanaman dan pada tanah di sekitar perakaran pisang (Dropkin, 1992).
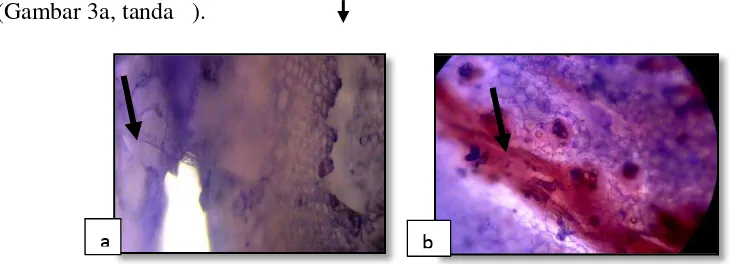
Histopatologi akar tanaman pisang yang terserang R.similis telah diteliti bahwa penetrasi nematoda tersebut ke dalam akar, biasanya terjadi di dekat dengan ujung akar, tetapi nematoda tersebut dapat melakukan serangan di seluruh panjang akar. Nematoda betina dan larva merupakan stadium yang infektif, sedangkan yang jantan secara morfologi mengalami degenerasi (tidak mempunyai stilet) dan mungkin tidak bersifat parasitik. Setelah masuk ke dalam jaringan akar tanaman pisang nematoda tersebut menempati ruang-ruang interseluler di parenkim (Gambar 5) dan korteks tempat
nematoda tersebut memperoleh makanannya yaitu sitoplasma, sel-sel yang berada di dekatnya dan menimbulkan rongga-rongga yang kemudian menjadi satu membentuk saluran-saluran di dalam jaringan tersebut. Invasi ke dalam stele tidak pernah dijumpai walaupun akar terserang berat (Blake, 1966).
Gambar 5. R. similis didalam sel parenkim (Perbesaran objek 400X).
Perpindahan dan peletakkan telur dipengaruhi oleh faktor makanan, misalnya nematoda betina berpindah tempat dari luka pada akar untuk mencari jaringan akar sehat. Di dalam jaringan yang terinfeksi nematoda betina meletakkan telur. Daur hidupnya dari telur ke telur generasi berikutnya membutuhkan waktu 20 sampai 25 hari pada suhu berkisar 240C sampai 320C (Luc et al., 1995).
Nematoda betina menghasilkan 4-5 butir telur setiap hari selama 10 -12 hari. Telur menetas 8 sampai 10 hari dan stadia juvenil secara keseluruhan
Gambar 6. Siklus hidup R. similis R.similis in banana roots, diakses tanggal 24 Mei 2012).
Radopholus similis terdapat secara luas di daerah tropik dan sub tropik, merupakan patogen penting pada pisang pada daerah penanaman pisang. Nematoda ini berbentuk seperti cacing, panjang 0,65 mm dan lebar 25μm. Nematoda ini hidup dan bereproduksi di dalam rongga korteks akar. semua larva dan dewasa dapat menginfeksi akar (Dropkin, 1992).
Gejala serangan
Gejala kerusakan yang paling jelas akibat serangan R. similis pada pertanaman pisang ialah rebahnya batang pisang atau mudahnya tanaman dicabut khususnya pada waktu tanaman berbuah (Gambar 7). Tingkat kerusakan yang berat ditandai dengan makin panjangnya pertumbuhan vegetatif sampai berkurangnya berat tandan secara drastis. Hal tersebut menunjukkan terdapat dua tipe kerusakan yang dapat ditimbulkan pada pertanaman pisang yaitu mempengaruhi tegak berdirinya tanaman pisang dan kemampuan menyerap air dan hara (Luc et al., 1995).
Gambar 7. Tanaman pisang rebah akibat serangan R. similis.
Nematoda sering diketahui terlibat dalam kekomplekan penyakit dimana nematoda menjadi penyebab penyakit primer dan beberapa spesies cendawan atau bakteri menjadi penyebab penyakit sekunder. Keduanya dapat menimbulkan kerugian dan penyakit secara sendirinya tetapi ketika bersatu ada efek sinergi mengakibatkan kerusakan yang lebih besar dengan perubahan gejala dan keterkaitan dalam memparasiti inangnya. Nematoda memberikan jalan masuk kepada patogen-patogen lain (Singh, 2000).
dengan jumlah kerusakan yang disebabkan oleh masing-masing patogen tersebut apabila menyerang secara sendiri-sendiri. Juga varietas yang sebenarnya tahan terhadap cendawan, akan terinfeksi oleh cendawan tersebut setelah sebelumnya diinfeksi oleh nematoda (Agrios, 1996).
Nematoda ini menyediakan jalan masuk untuk patogen lemah, khususnya patogen tanaman pisang seperti Fusarium spp., Cylindrocladium spp., Rhizoctonia spp.
yang memungkinkan jamur ini memperpanjang nekrosis di bagian akar dan akhirnya akar mati (INIBAP, 1994)
Pengelolaan penyakit
Penggunaan kultivar resisten atau genotip tahan telah terbukti menjadi ukuran utama. Tindakan budaya dan karantina diimplementasikan untuk menghindari masuknya patogen. Peralatan yang digunakan di daerah yang terinfeksi tidak boleh digunakan di daerah-daerah bebas kecuali telah dengan hati-hati dicuci dan didesinfeksi. Secara kimia biasanya tanah difumigasi dengan metil bromida. Kelemahan secara kimia yaitu bisa terinfeksi kembali dalam dua atau tiga tahun kemudian (Perez dan Vicente, 2004).
Cendawan endofit
Cendawan endofit tergolong pada Ascomycotina atau Deuteromycotina. Cendawan endofit dapat menginfeksi tumbuhan dan hidup secara simbiosis mutualistik dengan tanaman inangnya. Dalam simbiosis ini, cendawan dapat membantu proses penyerapan unsur hara yang dibutuhkan oleh tanaman untuk proses fotosintesis serta melindungi tanaman inang dari serangan penyakit, dan hasil fotosisntesis dapat dipergunakan cendawan untuk mempertahankan hidupnya (Lingga, 2009).
Penggunaan antagonis merupakan cara aman jangka panjang. Sebagian besar penelitian yang sedang berlangsung pada biokontrol layu Fusarium pada pisang telah diarahkan pada pengurangan inokulum dalam tanah dengan penggunaan antagonis endofit (Kidane dan Laing, 2010).
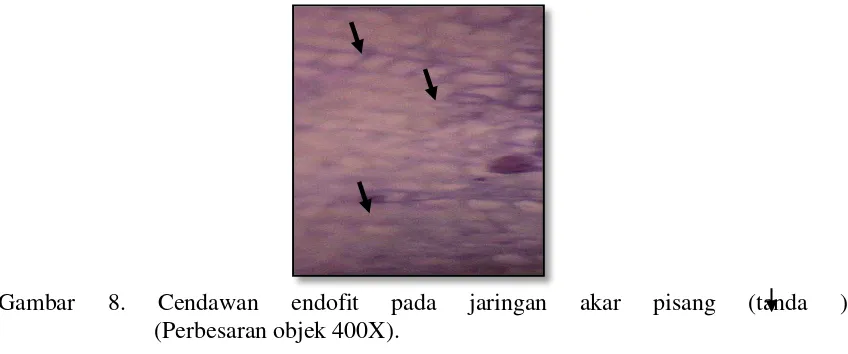
Gambar 8. Cendawan endofit pada jaringan akar pisang (tanda ) (Perbesaran objek 400X).
Potensi cendawan endofit dalam mengendalikan penyakit layu fusarium dan R. similis pada tanaman pisang
Globodera. Masalah nematoda pada beberapa tanaman penting di Indonesia dan teknologi pengendalian yang sudah diperoleh sampai saat ini (Mustika, 2010)
Selaras dengan program Badan Litbang Pertanian 2005-2009 yang berkaitan dengan rekayasa dan pemanfaatan teknik biologi molekuler dan rekayasa genetik untuk perbaikan tanaman dan ternak, serta pemanfaatan kultur in vitro untuk perbanyakan tanaman, perbaikan varietas dan produksi metabolit sekunder. Penelitian pengendalian nematoda diarahkan pada bioteknologi dan pertanian berkelanjutan. Penelitian terutama ditujukan untuk mengantisipasi tuntutan konsumen yang makin peduli terhadap masalah lingkungan, dan juga sejalan dengan sistem pertanian yang lestari (sustainable agriculture) (Badan Penelitian dan Pengembangan Pertanian, 2004).
Endofit mendapat perhatian besar akhir-akhir ini antara lain karena keberadaannya sangat melimpah dan beragam, serta ditemukan dalam seluruh famili tanaman, baik tanaman pertanian maupun rumput-rumputan. Selain itu juga diketahui asosiasi endofit dengan tanaman inang bersifat mutualisme. Simbiosis mutualistik ini menyebabkan berkurangnya kerusakan pada sel atau jaringan tanaman, meningkatkan kemampuan bertahan hidup dan fotosintesis sel jaringan tanaman yang terinfeksi patogen tanah. Dalam simbiosis ini, cendawan endofit membantu tanaman lebih toleran terhadap faktor abiotik dan biotik (IPB, 2012).
Beberapa jenis cendawan endofit dapat membuat perangkap atau penjerat pada saat larva bergerak mengenai hifa. Cendawan tersebut juga mampu mengeluarkan zat kemoantraktan dan enzim kutikula, sehingga larva melekat dan selanjutnya terjadi penguraian lapisan kutikula yang menyebabkan kematian larva nematoda tersebut. Cendawan oportunistik dapat mengkolonisasi nematoda betina sebelum nematoda tersebut bertelur (Sayre, 1971 dalam Lingga, 2009).
BAHAN DAN METODE
Tempat dan waktu penelitian
Penelitian ini dilakukan di Laboratorium Penyakit Tumbuhan Program Studi Agroekoteknologi dan Rumah Kaca Fakultas Pertanian Universitas Sumatera Utara, Medan, dengan ketinggian + 25m dpl. Penelitian ini dilaksanakan mulai Juni-Desember 2012.
Bahan dan alat penelitian
Bahan yang digunakan dalam penelitian ini adalah isolat cendawan endofit koleksi Laboratorium Penyakit Tumbuhan Program Studi Agroekoteknologi Universitas Sumatera Utara yaitu E1 (5BSBH), E2 (4BSU), E3 (1BJH), E4 (1BTAH), E5 (1BSHT), isolat F. oxysforum f.sp. cubense isolat 7 (foc 7) koleksi Laboratorium Penyakit Tumbuhan Program Studi Agroekoteknologi Universitas Sumatera Utara, nematoda R. similis, tanaman pisang Barangan hasil kultur jaringan, media beras, media jagung, media potato dextrose agar (PDA), media wortel steril, air steril, NaOCl, tanah+pasir steril (1:1).
Alat yang digunakan dalam penelitian ini adalah pipet tetes, beacker glass,
corong Baermann, botol kultur, cawan petri, shaker, polybag, jarum ose, autoklaf, lampu bunsen, timbangan analitik, kantong plastik, aluminium foil.
Metode Penelitian
Penelitian ini dilakukan dengan menggunakan Rancangan Acak Lengkap Non Faktorial dengan 13 perlakuan dan 3 ulangan yaitu:
K : Kontrol ( tidak di inokulasi R. similis dan F. o. f.sp. cubense). Rs : inokulasi 300 R. similis
F : inokulasi F. o. f.sp. cubense .
E1RsW1FW2 : inokulasi E1, 1 minggu kemudian diinokulasi dengan 300 R. similis, 2 minggu kemudian diinokulasi dengan Foc.
E2RsW1FW2 : inokulasi E2, 1 minggu kemudian diinokulasi dengan 300 R. similis, 2 minggu kemudian diinokulasi dengan Foc.
E3RsW1FW2 : inokulasi E3, 1 minggu kemudian diinokulasi dengan 300 R. similis, 2 minggu kemudian diinokulasi dengan Foc.
E4RsW1FW2 : inokulasi E4, 1 minggu kemudian diinokulasi dengan 300 R. similis, 2 minggu kemudian diinokulasi dengan Foc.
E5RsW1FW2 : inokulasi E5, 1 minggu kemudian diinokulasi dengan 300 R. similis, 2 minggu kemudian diinokulasi dengan Foc.
RsFW1E1W2 : inokulasi 300 R. similis, 1 minggu kemudian diinokulasi dengan Foc 2 minggu kemudian diinokulasi dengan E1.
RsFW1E2W2 : inokulasi 300 R. similis, 1 minggu kemudian diinokulasi dengan Foc 2 minggu kemudian diinokulasi dengan E2.
RsFW1E4W2 : inokulasi 300 R. similis, 1 minggu kemudian diinokulasi dengan Foc 2 minggu kemudian diinokulasi dengan E4.
RsFW1E5W2 : inokulasi 300 R. similis, 1 minggu kemudian diinokulasi dengan Foc 2 minggu kemudian diinokulasi dengan E5.
Ket : E1, E2, E3, E4 dan E5 (Cendawan Endofit) Rs (Radopholus similis)
F (F. oxysforum f.sp. cubens)
Untuk menentukan banyaknya ulangan ditentukan dengan rumus: t (r-1) > 15
r > 2,071
Jumlah ulangan yang dilakukan pada penelitian ini adalah 3 ulangan setiap perlakuan, setiap perlakuan terdiri dari 4 tanaman. Maka jumlah keseluruhan tanaman yaitu 156 tanaman.
Data hasil penelitian dianalisis dengan menggunakan sidik ragam berdasarkan model sebagai berikut:
Yij : µ + σi + εij Dimana :
Yij = Nilai pengamatan pada suatu percobaan yang disebabkan perlakuan ke-i dan ulangan ke-j
µ = Rataan
σi = Pengaruh perlakuan ke-i pada ulangan ke-j εij = Pengaruh error perlakuan ke-i dan ulangan ke-j
(Bangun, 1994).
Bibit pisang yang digunakan adalah kultivar pisang Barangan hasil perbanyakan kultur jaringan umur 2 bulan setelah aklimatisasi. Bibit ditanam serentak di dalam pot plastik berdiameter 30 cm yang berisi 5 kg media steril (tanah dan pasir 1:1). Ditempatkan di rumah kaca sesuai dengan perlakuan. Dalam pemeliharaan digunakan pupuk NPK (15:15:15) sebanyak 1 gram per pot pada awal tanam dan 30 hst dengan cara menaburkan pupuk di sekeliling batang tanaman.
Perbanyakan cendawan endofit pada media jagung
Lima isolat cendawan endofit koleksi Laboratorium Penyakit Tumbuhan digunakan dalam penelitian ini. Semua isolat diperbanyak dalam media jagung. Jagung giling direndam selama 24 jam, dicuci dan dikukus sampai lunak. Ditimbang masing-masing 100 gr, dimasukkan media ke dalam kantong plastik tahan panas, disterilkan dalam autoklaf pada suhu 100-1210C selam 15 menit. Media didinginkan kemudian diinokulasikan inokulum endofitdengan menggunakan jarum ose yang dilakukan dalam
Laminar air flow. Diinkubasi dalam ruangan bersih dengan suhu 250C-270C selama 7-14 hari.
(Dinas pertanian dan Tanaman Pangan Jawa Barat, 2012). Perbanyakan nematoda Radopholus similis
a. Pembuatan media wortel steril
b. Perbanyakan Radopholus similis
Radopholus similis yang diisolasi dari sampel akar dan tanah tanaman pisang diektraksi di laboratorium dengan menggunakan metode modifikasi corong Baermann. Inokulum yang diperoleh diperbanyak dengan menggunakan metode Huettel (1985) menggunakan media wortel steril. Nematoda R. similis yang telah diisolasi disterilisasi menggunakan larutan HgCl, 0,01% dan Streptomycin sulfat 0,1% selama 30 detik kemudian dibilas dengan air steril dan diinokulasikan menggunakan pipet steril pada potongan wortel. Biakan diinkubasi pada suhu 270C selama +2 bulan. Biakan ini digunakan sebagai sumber inokulum.
Perbanyakan cendawan Fusarium oxysporum f.sp.cubense
Fusarium oxysporum f.sp. cubense yang digunakan adalah isolat 7 (Foc 7) koleksi Laboratorium Penyakit Tumbuhan. Perbanyakan dilakukan menggunakan media beras. Beras direndam selama 24 jam, dicuci dan dikukus sampai lunak. Ditimbang media masing-masing 100 gr, dimasukkan media kedalam kantong plastik tahan panas, disterilkan dalam autoklaf pada suhu 100-1210C selam 15 menit. Media didinginkan kemudian diinokulasikan inokulum Foc pada media beras dengan menggunakan jarum ose yang dilakukan di Laminar air flow. Dinkubasi dalam ruangan bersih dengan suhu 250C-270C selama 7-14 hari.
(Dinas pertanian dan Tanaman Pangan Jawa Barat, 2012). Aplikasi perlakuan
a. Aplikasi Radopholus similis
tiga hari berturut-turut pada lubang yang dibuat di sekeliling batang semu pisang dengan kedalaman 3 cm (Spijer dan De Waele 1997 dalam Jumjunidang et al., 2009).
b. Aplikasi cendawan endofit
Inokulum cendawan endofit yang telah diperbanyak pada media jagung di infestasikan 10 g biakan endofit (kerapatan konidia 106) ke dalam media tanam (Maimunah, 1999).
c. Aplikasi F. oxysporum f.sp cubense
Aplikasi Foc yang telah diperbanyak pada media beras diinfestasikan 10 g
biakan Foc tersebut (kerapatan konidia 106) ke dalam media tanam (Maimunah, 1999).
Pemeliharaan tanaman
Pemeliharaan tanaman dilakukan seperti: penyiraman, pemupukan dan penyiangan.
Peubah amatan
1. Periode inkubasi (hsi)
Diamati pada masing-masing perlakuan sejak sehari setelah inokulasi sampai munculnya gejala pertama layu fusarium pada tanaman uji.
2. Kejadian penyakit (%)
Kejadian penyakit pada tanaman dihitung dengan pengamatan gejala eksternal, dilakukan pada 60 hsi. Perhitungan dilakukan dengan menggunakan metode Abbott (1925) yaitu dengan rumus sebagai berikut:
Persentase kejadian penyakit =∑ tanaman terserang pada tiap perlakuan
∑tanaman yang diamati × 100%
3. Keparahan penyakit (%)
Gejala serangan Foc pada bonggol ditandai dengan terdapatnya bercak nekrosis warna coklat, hitam atau kemerahan. Indeks nekrosis jaringan bongol dihitung dengan metode INIBAP (1994), menggunakan kriteria sebagai berikut:
0 : tidak ada nekrosis (diskolorisasi) 1 : ada sedikit diskolorisasi
2 : diskolorisasi sampai 1/3 berkas pembuluh 3 : diskolorisasi sampai 1/3-2/3 berkas pembuluh 4 : diskolorisasi > 2/3 berkas pembuluh
5 : berkas pembuluh (bonggol) penuh dengan diskolorisasi
Keparahan penyakit diamati pada 15, 30, 45 dan 60 hsi dengan mengamati diskolorisasi yang terdapat pada bonggol tanaman pisang secara internal dengan menggunakan metode Townsend & Hueberger (1948) dengan rumus sebagai berikut:
Kp =∑ ( ni x vi)
Z x N × 100%
Ket: Kp : keparahan penyakit Z : skor tertinggi
N : jumlah tanaman yang diamati
vi : nilai skor penyakit dari I = 0,1,2,… sampai I t- skor tertinggi
ni : jumlah tanaman dengan skor ke-i (Townsend & Hueberger, 1948 dalam Lisnawita et al., 1998). 4. Persentase nekrosis jaringan akar (%)
kemudian dibelah memanjang selanjutnya jaringan akar yang mengalami nekrosis dihitung.
5. Persentase akar yang mati (%)
Semua akar dipisahkan dari bonggol, kemudian dihitung jumlah akar primer yang mati dan jumlah akar yang masih hidup, persentase akar mati dihitung dengan rumus:
Persentase akar yang mati =∑ akar yang mati pada tiap perlakuan
∑akar yang diamati × 100%
(Jumjunidang, 2009) 6. Berat basah akar (g).
Berat basah akar dihitung pada 15, 30, 45, dan 60 hsi. 7. Populasi akhir nematoda R. similis (ji)
Populasi akhir adalah total populasi nematoda dari akar, bonggol dan tanah. Contoh akar dan bonggol masing-masing diambil sebanyak 10 g secara komposit setelah akar dan bonggol dicacah, kemudian dilakukan diekstraksi, sedangkan sampel tanah 10 g diekstraksi menggunakan modifikasi corong Baermann. Perhitungan faktor reproduksi dilakukan pada akhir penelitian dengan membandingkan populasi akhir dengan populasi awal atau Rf= Pf/Pi (Speijer dan Waele 1997).
HASIL DAN PEMBAHASAN
Periode inkubasi (hsi) dan Kejadian penyakit (%)
Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap periode inkubasi dan kejadian penyakit pada 60 hsi dapat dilihat dalam Tabel 1.
Perlakuan Hari setelah inokulasi (hsi) Kejadian penyakit
Keterangan: Tanda (-) tidak terdapat gejala serangan Foc pada tanaman pisang sampai akhir penelitian.
(a)
. Tanaman terlihat kekuningan dan layu. (b)
. Tanaman terlihat kekuningan tidak layu.
Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata menurut uji jarak Duncan pada taraf 5%. Angka di dalam kurung adalah angka hasil transformasi ��+ 0,5.
Penelitian menunjukkan bahwa pada aplikasi Foc secara tunggal diperoleh kisaran masa inkubasi 29 hsi. Pengamatan dilakukan dengan mengamati gejala visual yang tampak pada tepi daun bagian bawah berwarna kuning tua kemudian menjadi coklat dan mengering (Gambar 2). Perlakuan Foc secara tunggal memperlihatkan gejala serangan lebih lama tetapi secara perlahan tanaman layu dan mati. Hasil ini sama dengan penelitian yang dilakukan Jumjunidang (2011) yaitu masa inkubasi Foc yang diaplikasikan pada pisang Barangan sekitar 30,27 hsi. Lebih lamanya periode inkubasi pada perlakuan Foc tunggal disebabkan karena patogen ini merupakan patogen lemah yang hanya mampu berpenetrasi secara pasif sehingga diperlukan waktu yang lebih lama agar pathogen dapat berpenetrasi ke dalam jaringan inang.
dua minggu kemudian aplikasi endofit (RsFW1EW2) yaitu 8 hsi. Pada aplikasi R. similis dan Foc lebih awal terjadi sinergisme yaitu dengan adanya pelukaan oleh R. similis akan membantu patogen Foc masuk kedalam jaringan tanaman. Hal yang sama juga pada penelitian Lisnawita (1998) yang menyatakan bahwa di dalam korteks akar pisang, R. similis dapat menciptakan sumber makanan berupa asam amino bagi patogen lemah seperti Fusarium sehingga cendawan lebih cepat berkembang dan kerusakan akan menjadi lebih parah.
Perlakuan aplikasi endofit lebih awal tidak memperlihatkan gejala serangan Foc. Hal ini disebabkan infestasi endofit lebih awal dan waktu untuk beradaptasi dengan baik. Sehingga kondisi fisik dan kimia rizosfer dapat mempengaruhi kemampuan endofit dalam menekan perkembangan patogen dalam tanah. Sama dengan penelitian Sariyanto (2006) yang menyatakan bahwa aplikasi endofit lebih awal pada tanaman sebelum aklimatisasi akan mendapatkan hasil yang lebih baik dalam menekan perkembangan patogen dalam tanah. Demikian juga dengan penelitian Carrol (1988) yaitu cendawan endofit menghasilkan mikotoksin atau metabolit lainnya yang menyebabkan perubahan fisiologi dan biokimia inang sehingga keberadaan endofit dalam jaringan tanaman dapat berperan langsung menghambat perkembangan patogen dalam tanaman.
dengan penelitian Maimunah (1999) yang menyatakan bahwa patogen Foc menyerang jaringan empulur batang melalui akar yang luka atau terinfeksi. Batang yang terserang akan kehilangan banyak cairan dan berubah warna menjadi kecoklatan, tepi bawah daun menjadi kuning tua (layu), merambat ke bagian dalam secara cepat sehingga seluruh permukaan daun tersebut menguning.
Penelitian menunjukkan perbedaan yang nyata antara perlakuan Foc secara tunggal dengan perlakuan lainnya (Tabel 1). Pada perlakuan cendawan endofit yang di aplikasikan diawal tidak terdapat gejala kejadian penyakit hingga akhir penelitian, sebaliknya pada perlakuan cendawan endofit setelah aplikasi Foc dan R. similis tampak persentase kejadian penyakit sebanyak 11%. Hal ini disebabkan karena cendawan endofit bersifat simbiosis mutualisme dengan tanaman inang. Salah satunya berpengaruh positif terhadap proses fotosintesis tanaman, sehingga tanaman lebih toleran terhadap faktor abiotik dan faktor biotik. Sesuai dengan pernyataan IPB (2012) yang menyatakan bahwa asosiasi endofit dengan tanaman inang bersifat mutualisme. Simbiosis mutualistik ini menyebabkan berkurangnya kerusakan pada sel atau jaringan tanaman, meningkatkan kemampuan bertahan hidup dan fotosintesis sel jaringan tanaman yang terinfeksi patogen tanah. Dalam simbiosis ini, cendawan endofit membantu tanaman lebih toleran terhadap faktor abiotik dan biotik.
Keparahan penyakit (%)
Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap tingkat keparahan penyakit (%) dapat dilihat dalam Tabel 2.
Tabel 2. Pengaruh inokulasi R. similis dan Foc dan cendawan endofitterhadap tingkat keparahan penyakit (%).
Perlakuan 15 hsi 30 hsi 45 hsi 60 hsi
K 0,00 (0,71) b 0,00 ( 0,71) c 0,00 (0,71) c 0,00 (0,71) b
Rs 0,00 (0,71) b 0,00 (0,71) c 0,00 (0,71) c 0,00 (0,71) b
E1RsW1FW2 0,00 (0,71) b 0,00 (0,71) c 0,00 (0,71) c 0,00 (0,71) b
Keterangan: Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata menurut uji jarak Duncan pada taraf 5%. Angka di dalam kurung adalah angka hasil transformasi ��+ 0,5.
Tingkat keparahan penyakit tanaman diamati dengan membandingkan bagian tanaman sakit dari tanaman sampel. Pada penelitian ini, keparahan penyakit pada perlakuan Foc secara tunggal berbeda nyata dengan perlakuan lain (Tabel 2). Pada perlakuan Foc secara tunggal keparahan penyakit lebih tinggi dibanding dengan perlakuan lainnya. Hal ini disebabkan pada perlakuan Foc tunggal tidak ada hambatan bagi patogen untuk menginfeksi dan berinvasi di dalam jaringan tanaman. Infeksi Foc
pada akar tanaman pisang yang rentan dapat berkembang ke xilem dan berlanjut ke bonggol dan batang semu yang mengakibatkan gangguan transportasi air. Sehingga muncul gejala penguningan pada daun (Lampiran 31). Jumjunidang (2011) menyatakan keparahan penyakit berawal dari infeksi akar, berlanjut ke dalam bonggol, dan batang semu yang mengakibatkan gangguan transportasi air sehingga muncul gejala penguningan pada daun.
adalah gejala dalam. Jika pangkal batang dibelah terlihat garis-garis coklat atau hitam, paling jelas tampak pada batang, pada akar biasanya menyebabkan akar busuk.
Pengamatan tingkat keparahan penyakit (%) (Tabel 2) menunjukkan bahwa tingkat keparahan penyakit pada perlakuan kombinasi R. similis dan Foc lebih awal dan diikuti aplikasi endofit (RsFW1EW2) tampak pengurangan proporsi keparahan penyakit dari 15 hsi, 30 hsi, 45 hsi hingga 60 hsi. Hal ini disebabkan cendawan endofit menghasilkan mikotoksin yang dapat menghambat perkembangan patogen dalam tanah. Carrol (1988) mengungkapkan bahwa cendawan endofit menghasilkan mikotoksin atau metabolit lainnya yang menyebabkan perubahan fisiologi dan biokimia inang sehingga keberadaan endofit dalam jaringan tanaman dapat berperan langsung menghambat perkembangan patogen dalam tanaman.
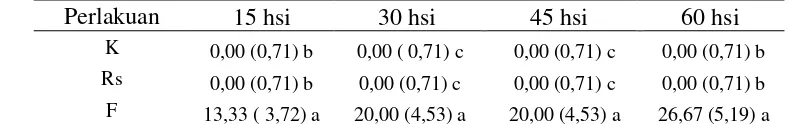
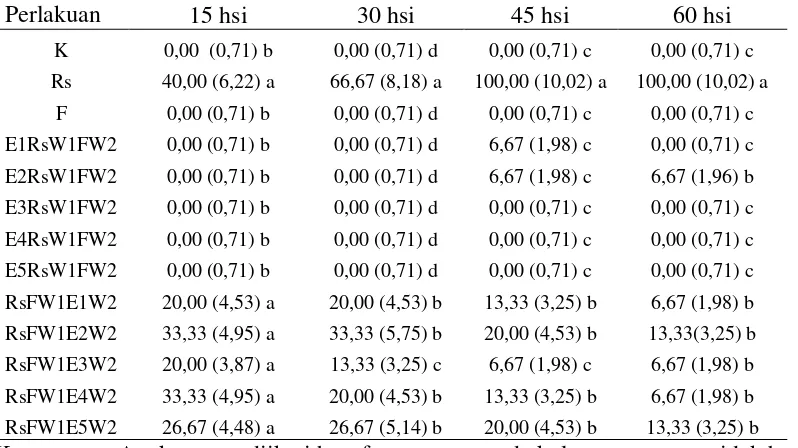
Nekrosis jaringan akar (%)
Hasil analisis statistika menunjukkan bahwa pengamatan 60 hsi terdapat perbedaan yang nyata pada perlakuan Rs (aplikasi nematoda R. similis secara tunggal) dengan perlakuan lainnya (Tabel 3). Pada pengamatan 15 hsi nekrosis jaringan akar oleh R. similis mencapai 40% dan pada pengamatan 60 hsi mencapai 100%. Hal ini disebabkan peran R. similis di dalam jaringan tanaman adalah bertelur dan menghasilkan populasi baru selama melakukan perpindahan. Histopatologi akar tanaman pisang yang terserang R. similis pada penelitian ini diperoleh bahwa penetrasi nematoda terjadi di ujung akar (rambut-rambut akar), setelah masuk ke dalam jaringan akar tanaman pisang, nematoda menempati ruang-ruang interseluler di parenkim dan korteks, sehingga tampak nekrotik pada jaringan akar tanaman apabila diamati secara visual maupun dibelah (Gambar 5). Nematoda yang bersifat parasit adalah
sebagai alat pelubang akar untuk mencari jaringan akar tanaman sehat. Dropkin (1992) menyatakan bahwa R. similis adalah nematoda endoparasit yang menyelesaikan daur hidupnya di dalam jaringan tanaman.
Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap nekrosis jaringan akar (%) dapat dilihat dalam Tabel 3.
Tabel 3. Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap nekrosis jaringan akar (%).
Perlakuan 15 hsi 30 hsi 45 hsi 60 hsi
K 0,00 (0,71) b 0,00 (0,71) d 0,00 (0,71) c 0,00 (0,71) c
Rs 40,00 (6,22) a 66,67 (8,18) a 100,00 (10,02) a 100,00 (10,02) a
F 0,00 (0,71) b 0,00 (0,71) d 0,00 (0,71) c 0,00 (0,71) c
E1RsW1FW2 0,00 (0,71) b 0,00 (0,71) d 6,67 (1,98) c 0,00 (0,71) c
E2RsW1FW2 0,00 (0,71) b 0,00 (0,71) d 6,67 (1,98) c 6,67 (1,96) b
E3RsW1FW2 0,00 (0,71) b 0,00 (0,71) d 0,00 (0,71) c 0,00 (0,71) c
E4RsW1FW2 0,00 (0,71) b 0,00 (0,71) d 0,00 (0,71) c 0,00 (0,71) c
E5RsW1FW2 0,00 (0,71) b 0,00 (0,71) d 0,00 (0,71) c 0,00 (0,71) c
RsFW1E1W2 20,00 (4,53) a 20,00 (4,53) b 13,33 (3,25) b 6,67 (1,98) b
RsFW1E2W2 33,33 (4,95) a 33,33 (5,75) b 20,00 (4,53) b 13,33(3,25) b
RsFW1E3W2 20,00 (3,87) a 13,33 (3,25) c 6,67 (1,98) c 6,67 (1,98) b
RsFW1E4W2 33,33 (4,95) a 20,00 (4,53) b 13,33 (3,25) b 6,67 (1,98) b
RsFW1E5W2 26,67 (4,48) a 26,67 (5,14) b 20,00 (4,53) b 13,33 (3,25) b
Keterangan: Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata menurut uji jarak Duncan pada taraf 5%. Angka di dalam kurung adalah angka hasil transformasi ��+ 0,5.
minggu kemudian aplikasi Foc) berbeda nyata dengan Rs. Hal ini disebabkan oleh infestasi endofit dilakukan di awal dan diberi waktu seminggu sebelum aplikasi R. similis sehingga endofit dapat beradaptasi dengan baik. Sayre (1971); Lingga (2009) menyatakan bahwa cendawan endofit dapat membuat perangkap atau penjerat pada saat larva bergerak mengenai hifa. Cendawan mampu mengeluarkan zat kemoantraktan dan enzim kutikula, sehingga larva melekat dan selanjutnya terjadi penguraian lapisan kutikula yang menyebabkan kematian larva nematoda tersebut. Cendawan juga dapat mengkolonisasi nematoda betina sebelum nematoda tersebut bertelur.
Nekrosis jaringan akar pada perlakuan RsFW1EW2 (aplikasi nematoda R. similis, semingggu kemudian aplikasi Foc, dua minggu kemudian aplikasi endofit) pada 15 hsi menunjukkan tingginya tingkat nekrosis pada jaringan akar. Hal ini disebabkan sinergisme nematoda R. similis sebagai pelubang akar yang menyediakan jalan masuk patogen Foc ke dalam jaringan tanaman, sehingga Foc dapat dengan cepat memperpanjang nekrosis di bagian akar. Perbandingan aplikasi endofit lebih awal dan aplikasi R. similis dan Foc lebih awal, terdapat perbedaan persentase nekrosis jaringan akar dari 15 hsi, 30 hsi, 45 hsi, dan 60 hsi (Tabel 3) yaitu kerusakan jaringan akar lebih besar pada aplikasi R. similis dan Foc lebih awal karena infeksi oleh Foc telah diinfeksi nematoda. Tetapi dengan aplikasi endofit dua minggu kemudian menyebabkan berkurangnya nekrosis jaringan akar pada 30 hsi, 45 hsi, dan 60 hsi. Cendawan endofit berperan dalam menekan perkembangan nematoda R. similis dalam jaringan tanaman
kemoantraktan dan enzim kutikula, sehingga larva melekat dan selanjutnya terjadi penguraian lapisan kutikula yang menyebabkan kematian larva nematoda tersebut. Akar mati (%)
Analisis statistika menunjukkan bahwa pengamatan 15 hsi terdapat perbedaan yang nyata persentase akar mati pada perlakuan RsFW1EW2 (aplikasi Rs, seminggu kemudian aplikasi Foc, dua minggu kemudian aplikasi endofit) dengan perlakuan ERsW1FW2 (aplikasi endofit, seminggu kemudian aplikasi R. similis, dua minggu kemudian aplikasi Foc), K (kontrol), F (Foc tunggal) (Tabel 4). Hal ini disebabkan pada aplikasi RsFW1EW2 terjadi sinergisme antara nematoda R. similis
dan Foc sedangkan aplikasi endofit pada pengamatan 15 hsi belum menunjukkan reaksi terhadap tanaman uji karena endofit yang diaplikasikan belum beradaptasi dengan baik pada tanaman sehingga toleransi tanaman terhadap kerusakan akar belum tampak.
Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap persentase akar mati (%) dapat dilihat dalam Tabel 4.
Keterangan: Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata menurut uji jarak Duncan pada taraf 5%. Angka di dalam kurung adalah angka hasil transformasi ��+ 0,5.
Analisis statistika dapat dilihat bahwa pada pengamatan 30 hsi, 45 hsi hingga 60 hsi (Tabel 4) terjadi pengurangan persentase akar mati yaitu pada perlakuan endofit. Pada penelitian ini dengan adanya endofit dapat meningkatkan toleransi tanaman terhadap kerusakan jaringan akar oleh nematoda. Tingkat pengurangan kerusakan bervariasi diantara isolat yang diperlakukan. E3 (1BJH) merupakan isolat endofit yang memberikan hasil terbaik terhadap pengurangan akar mati. Dalam hal ini dengan pengaplikasian endofit mampu meningkatkan ketersediaan nutrisi dan menghasilkan hormon pertumbuhan. Hal ini sesuai dengan penelitian Lingga (2009) yang menyatakan bahwa keunggulan cendawan endofit sebagai agen pengendali hayati yaitu mampu meningkatkan ketersediaan nutrisi, menghasilkan hormon pertumbuhan dan mengendalikan penyakit tumbuhan serta menginduksi ketahanan tanaman.
Perlakuan tanpa aplikasi endofit yaitu Rs, F (R. similis, Foc) terjadi peningkatan persentase akar mati. Hal ini disebabkan kondisi tanaman yang rentan sehingga tidak mampu menghambat perkembangan patogen Foc dalam tanaman dan masuknya R. similis kedalam jaringan tanaman. Dapat dilihat bahwa persentase akar mati pada K (Kontrol) dengan Rs (R. similis tunggal) dan F (Foc tunggal) berbeda nyata, karena pada K tidak dilakukan aplikasi R. similis, Foc dan endofit, sedangkan pada F terjadi pembusukan akar, dalam hal ini Foc mampu menyerang jaringan vaskular yang dapat mempercepat pembusukan dan kematian akar. Wardlaw (1961) dalam Jumjunidang et al. (2009) menyatakan jamur Foc dapat menginfeksi melalui akar luka maupun akar sehat.
Data pengamatan menunjukkan bahwa berat basah akar dapat dihubungkan dengan nekrosis jaringan akar dan persentase akar mati, yaitu semakin tinggi berat basah akar maka semakin rendah persentase akar mati (Tabel 5) dan (Tabel 4). Hal ini disebabkan pengaruh infeksi patogen Foc pada tanamn pisang akan mengganggu proses penyerapan transportasi air dan zat makanan di dalam tanah, menyebabkan tanaman layu dan mati. Maimunah (1999) mengungkapkan bahwa infeksi
Foc pada tanaman pisang, akan menganggu proses penyerapan, transportasi air dan zat makanan di dalam tanah, sehingga tanaman menjadi layu dan akhirnya mati.
Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap berat basah akar dapat dilihat dalam Tabel 5.
Tabel 5. Pengaruh inokulasi R. similis, Foc dan cendawan endofitterhadap berat basah akar (g).
Populasi akhir nematoda R. similis (ji)
Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap populasi akhir nematoda R. similis (ji) dapat dilihat dalam Tabel 6.
Tabel 6. Pengaruh inokulasi R. similis, Foc dan cendawan endofit terhadap populasi akhir nematoda R. similis (ji).
Perlakuan Populasi akhir nematoda R. similis 60 hsi (total populasi di
dalam tanah, akar dan bonggol) (ji) Pf/Pi
K - -
Keterangan: Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata menurut uji jarak Duncan pada taraf 5%. Angka di dalam kurung adalah angka hasil transformasi ��+ 0,5.
bertelur untuk menghasilkan populasi baru selama melakukan perpindahan. Nematoda ini ditemukan pada semua tingkatan perkembangan akar tanaman dan pada tanah di sekitar perakaran pisang. Hal ini juga dinyatakan oleh Dropkin (1992) yang menyatakan bahwa R. similis adalah spesies nematoda endoparasitik yang berpindah-pindah yang mampu menyelesaikan daur hidupnya di dalam jaringan korteks akar tanaman pisang.
Dari hasil pengamatan 60 hsi diperoleh populasi R. similis lebih tinggi pada perlakuan R. similis secara tunggal yaitu mencapai 2028 ji dengan faktor reproduksi 6. Hal ini disebabkan oleh kompetisi antar nematoda di dalam tanah dan korteks akar dalam ketersediaan tempat hidup dan makanan yang berpengaruh terhadap kelangsungan hidup nematoda itu sendiri. Akar tanaman yang mendapat perlakuan R. similis dan Foc lebih awal terdapat populasi nematoda yang tinggi hingga 230 ji (RsFW1E3W2). Pada perlakuan kombinasi R. similis dan Foc, akar pisang mengalami pembusukan. Pembusukan ini menyebabkan R. similis tidak dapat berkembang biak dan mati, sebab R. similis parasit obligat. Pada penelitian ini juga terjadi peran antagonis antara kehadiran Foc dan R. similis di dalam jaringan akar. Cendawan Foc
mengeluarkan asam fusarik yang bersifat nematoksin yang akan mempengaruhi reproduksi dan perkembangan nematoda. Hal ini sama dengan penelitian Lisnawita (1998) yang menyatakan bahwa kehadiran Foc dan R. similis secara bersamaan dapat memberikan pengaruh yang antagonistik terhadap reproduksi nematoda. Cendawan layu mengeluarkan toksin berupa asam fusarik yang secara tidak langsung bersifat nematoksin.
membuat perangkap terhadap larva nematoda. Cendawan juga mengeluarkan zat kemoantraktan dan enzim kutikula, sehingga menyebabkan kematian larva nematoda Sayre (1971) dan Lingga (2009) mengungkapkan bahwa cendawan endofit dapat membuat perangkap atau penjerat pada saat larva bergerak mengenai hifa. Cendawan tersebut juga mampu mengeluarkan zat kemoantraktan dan enzim kutikula, sehingga larva melekat dan selanjutnya terjadi penguraian lapisan kutikula yang menyebabkan kematian larva nematoda tersebut.
KESIMPULAN DAN SARAN
Kesimpulan
1. Cendawan endofit yang diuji memiliki kemampuan dalam menghambat perkembangan layu fusarium dan nematoda R. similis.
2. Isolat E3 (1BJH) memiliki kemampuan terbaik terhadap pertumbuhan tanaman (persentase akar mati).
3. Periode inkubasi tercepat terjadi pada aplikasi RsFW1E2 (aplikasi R. similis, seminggu kemudian aplikasi Foc, dua minggu kemudian aplikasi endofit) yaitu 24 hsi dan penghambatan terjadi setelah aplikasi endofit.
4. Aplikasi cendawan endofit lebih awal pada tanaman dapat menghambat pertumbuhan patogen didalam tanah.
5. Kehadiran patogen Foc dan endofit di dalam jaringan akar tanaman dan tanah dapat mempengaruhi perkembangan nematoda R. similis (Populasi akhir nematoda R. similis).
DAFTAR PUSTAKA
Abbott, 1925. The basic principles of crop protection field trials. Planzenschutz-Nachrichten Bayer AG, Leverkusen. Didalam Lisnawita, M.S. Sinaga, S. Mulyati, dan I. Mustika. 1998. Analisis potensi sinergisme Radopholus similis Cobb. dan Fusarium oxysporum Schlecht, f.sp. cubense (E.F. Smith) Snyd. & Hans. dalam perkembangan layu fusarium pada pisang. Buletin Hama dan Penyakit Tumbuhan Fakultas Pertanian IPB. 10(2): 11-17.
Agrios, G. N. 1996. Ilmu penyakit tumbuhan. Busnia M, Penerjemah Yogyakarta: Gajah Mada University Press. Terjemahan dari: Plant Patology 3nded.
Badan penelitian dan Pengembangan Pertanian. 2004.
agriculture. diakses pada tanggal 30 April 2012.
Balio, A. 1981. Structure activity relationship. New York Academic Press. Di dalam Jumjunidang., Hermanto dan Riska. 2011. Virulensi Isolat Fusarium
oxysporum f. sp. cubense VCG 012113/16 pada pisang Barangan dari varietas pisang dan lokasi yang berbeda. J. Hort. 21(2): 145-151. Balai Penelitian Tanaman Tropika. Solok.
Bangun, M.K, 1994. Rancangan percobaan. Fakultas Pertanian Universitas Sumatara Utara, Medan.
Bappenas. 2000. Pisang (Musa sp.). Jakarta: Ristek. http: www. ristek. go. id (28 maret 2012).
Blake, C. D. 1966. The Histological changes in banana roots caused by Radopholus similis and Helicotylenchus multicinctus. Nematologica 12:129-137.
Carrol, G. C. 1988. Fungal endophytes in stems and leaves. From latent pathogens to mutualistic symbiont. Ecology. 69:2-9. Di dalam IPB, 2008. Bab II bahan dan Metode stp. http://www.Ipb respository bahan dan metode.co.id. diakses pada tanggal 30 april 2012.
Clay, K. 1988. Fungal endophytes of grasses: a devensive mutualism between plants and fungi. Ecology 69 (1): 10-16. Di dalam Lingga, R. 2009. Uji nematisidal jamur endofit tanaman padi (Oryza sativa L.) terhadap nematoda puru akar (Meloidogyne sp.). Skripsi. Universitas Sumatera Utara. Medan.
Departemen pertanian. 2000. Pisang (Musa sp.). Jakarta: Riset dan teknologi. http: www. ristek. go.id diakses pada tanggal 28 maret 2012.
Dinas pertanian Sumatera Utara, 2012. Analisa daerah rawan serangan OPT Fusarium pada tanaman Pisang Di Propinsi Sumatera Utara musim tanam 2011. Balai proteksi tanaman pangan dan hortikultura I. Sumatera Utara. Medan.
Djatnika. I., Hermanto dan Elisa. 2003. Pengendalian hayati layu fusarium pada tanaman pisang dengan Pseudomonas Fluorescens dan Gliocladium sp. Balai Penelitian Tanaman Buah. Sumatera Barat. J. Hort. 13(3):205-211.
Dropkin, V. H. 1992. Pengantar nematologi tumbuhan. 2nd ed. Gadjah Mada University Press, Yogyakarta.
Harni, R., Abdul, M., Supraman., Mustika, I. 2007. Potensi bakteri endofit pengendali nematoda peluka akar (Pratylenchus brachyurus) pada Nilam. Hayati Journal of Boisciences 14 (1): 7-12. Research Institute of Medicinal and Aromatic Crop. Bogor.
http://www. Cultures microconidia of Fusarium oxysporum f.sp.cubense, 2012. diakses pada tanggal 30 April 2012.
R.similis in banana roots, 2012. diakses pada tanggal 30
April 2012.
Huettel RN. 1985. Carrot disc culture. In: Zukermant BM, Mai WF, Harrison (ed).
Plant Nematology Laboratory Manual. Massachusetts: The Universitv of Massachusetts Agricultural Experiment Station. p 153-154. Didalam Harni, R., Abdul, M., Supraman., Mustika, I. 2007. Potensi bakteri endofit pengendali nematoda peluka akar (Pratylenchus brachyurus) pada Nilam. Journal of Boisciences 14 (1): 7-12.
INIBAP, 1994. IMTP Phase II Technical guidelines for Fusarium wilt sites. International Network for The Improvement of Banana and Plantain. 10(2): 11-17.
IPB, 2012. Bab II bahan dan Metode stp. http://www.Ipb respository bahan dan metode.co.id. diakses pada tanggal 30 april 2012.
Jumjunidang, 2009. Efikasi isolat cendawan Mikoriza Arbuskula Indogenous pisang terhadap nematoda Radopholus similis pada pisang Ambon Hijau. J.Horti. 19(2):186-191. Balai Penelitian tanaman tropika. Solok.
Jumjunidang., Hermanto dan Riska. 2011. Virulensi isolat Fusarium oxysporum f. sp.