PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Keragaman Genetik Bawang Merah (Allium cepa L.) Berdasarkan Marka Morfologi dan ISSR adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2016
Vebrita Sari
RINGKASAN
VEBRITA SARI. Keragaman Genetik Bawang Merah (Allium cepa L.) Berdasarkan Marka Morfologi dan ISSR. Dibimbing oleh MIFTAHUDIN dan SOBIR.
Bawang merah (Allium cepa L.) merupakan salah satu spesies dari marga
Allium dengan karakteristik morfologi umbi berlapis, memiliki aroma dan rasa yang unik. Bawang merah sangat populer dalam dunia kuliner yaitu sebagai bumbu masakan, saat ini bawang merah sedang dikembangkan sebagai obat tradisional. Pengembangan varietas bawang merah membutuhkan variasi genetik dari plasma nutfah bawang merah. Namun, informasi tentang keragaman genetik bawang merah lokal Indonesia masih terbatas. Penelitian ini bertujuan untuk menganalisis keragaman genetik dari 34 genotipe bawang merah koleksi Pusat Kajian Hortikultura Tropika Institut Pertanian Bogor (PKHT-IPB) berdasarkan marka morfologi dan ISSR. Hasil penelitian ini diharapkan dapat dijadikan informasi dasar dalam pengelolaan sumber daya genetik dan pemuliaan bibit unggul bawang merah di Indonesia.
Penelitian ini dilaksanakan pada bulan Oktober 2014 sampai dengan September 2015. Sebanyak 34 genotipe bawang merah yang berupa umbi dikoleksi dari beberapa daerah di Indonesia. Sejumlah 24 karakter morfologi dan agronomi diamati dalam penelitian ini. Marka Inter Simple Sequences Repeat
(ISSR) juga digunakan untuk menganalisis keragaman genetik bawang merah. DNA total diisolasi menggunakan metode CTAB. Amplifikasi PCR menggunakan
GoTaq® Green PCR Mix dengan 13 primer ISSR. Hasil PCR dielektroforesis dalam gel agarosa 1.2%, divisualilasikan dengan sinar ultraviolet dan didokumentasikan menggunakan kamera digital. Karakter morfologi diskoring menjadi data multistate dan pita polimorfik diskoring menjadi data biner. Hasil skoring data morfologi dan molekuler ditabulasi dalam matriks, kemudian dianalisis menggunakan program NTSys. Dendrogram dikonstruksi menggunakan metode UPGMA dengan koefisien kemiripan simple matching (SM). Principal component analysis (PCA) dikonstruksi menggunakan metode SIMINT dengan koefisien correlation. Analisis struktur genetika populasi menggunakan program GenAlex.
Pengamatan morfologi menghasilkan 19 karakter polimorfik dari 24 karakter yang diamati. Karakter informatif untuk menganalisis keragaman morfologi bawang merah antara lain: tingkat kepatahan daun, jumlah siung umbi dan morfologi pada umbi. Bobot basah umbi tertinggi dimiliki oleh varietas Bangkok (57.56 g/rumpun) dan terendah dimiliki oleh varietas Kuning (2.33 g/rumpun). Hampir semua genotipe bawang merah yang diamati pada penelitian ini termasuk dalam kategori mutu I yang disenangi oleh konsumen dan petani. Rata-rata diameter umbi yang diamati lebih dari 1.7 cm (mutu I), yaitu berkisar antara 1.51 cm (genotipe Solo 7) sampai 7.54 cm (varietas Bangkok). Dendrogram menunjukkan bahwa 34 genotipe bawang merah terbagi menjadi dua kelompok utama (kelompok I dan II) dengan koefisien kemiripan berkisar antara 0.68 sampai 0.95.
bawang merah berkelompok menjadi dua kelompok (kelompok I dan II) dengan nilai koefisien kemiripan genetik berkisar antara 0.62 sampai 0.89. Primer informatif untuk menganalisis keragaman genetik bawang merah antara lain: primer ISSRred 4, ISSRred 9 dan ISSRred 20.
Keseluruhan genotipe, baik berdasarkan marka morfologi maupun marka ISSR menghasilkan dua kelompok utama, namun pengelompokkan tidak berhubungan dengan asal geografi genotipe tersebut. Pengelompokan diduga karena proses budidaya dalam waktu yang lama dan juga dipengaruhi oleh distribusi perdagangan bawang merah. Berdasarkan nilai struktur genetika populasi dari populasi bawang merah yang diamati, populasi asal Solo memiliki keragaman genetik tertinggi (I= 0.44, h= 0.31 dan PLP= 77.39%) dan populasi asal Solok memiliki keragaman genetik terendah (I= 0.37, h= 0.24 dan PLP= 67.83%).
Genotipe bawang merah yang dikategorikan unggul diantaranya genotipe dengan diameter umbi terbesar yaitu varietas Bangkok (7.54 cm) dan Sembrani (4.99 cm). Selain itu genotipe yang memiliki jumlah siung umbi terbanyak yaitu genotipe asal Pekanbaru (27 siung/rumpun) dan Nganjuk (20 siung/rumpun). Varietas Bangkok dan Sembrani dapat disilangkan dengan genotipe asal Pekanbaru dan Nganjuk karena memiliki jarak genetik cukup jauh. Genotipe tersebut dapat dijadikan sebagai genotipe tetua yang potensial dalam merakit varietas unggul bawang merah Indonesia.
SUMMARY
VEBRITA SARI. Genetic Diversity of Shallot (Allium cepa L.) Based on Morphological Characters and ISSR Markers. Supervised by MIFTAHUDIN and SOBIR.
Shallot (Allium cepa L.) is a member of Allium genus with layered-bulbs as its specific morphological characteristic, unique aroma and taste. The shallot is very common cooking ingredient in culinary world and now being developed as traditional medicine. Development of shallot varieties requires genetic variation of shallot germplasm. However, information about the genetic diversity of local shallot in Indonesia is still lacking. This study aimed to analyze the genetic diversity of 34 shallot collection’s genotypes from Center for Tropical Horticulture Studies of Bogor Agricultural University (PKHT- IPB) based on morphological characters and ISSR markers. It is expected that the result of this research will be valuable information for shallot genetic resource management and breeding program in Indonesia.
This study was conducted in October 2014 until September 2015. The 34-shallot collection’s genotypes were collected from several regions in Indonesia in the form of the bulb. An amount of 24 marker morphological and agronomic characters were observed in this research. Inter Simple Sequences Repeat (ISSR) markers were also used to analyze genetic diversity of shallot. Total DNA were isolated based on CTAB methode. The PCR amplification was used GoTaq® Green PCR Mix with 13 ISSR primers. PCR products was electrophoresed in 1.2% agarose gel, visualized using ultraviolet light and documented with a digital camera. The morphological characters was scored into multistate data and the polymorphic DNA band was scored into binary data. The morphological and molecular scoring data were tabulated in data matrix, then analyzed using NTSys. Dendrogram was constructed using similarity coefficient of simple matching and UPGMA method. Principal component analysis (PCA) was constructed using SIMINT method with coeffisient of correlation. The genetic population structure was analyzed using GenAlex.
Although both morphological characters and ISSR markers grouped all genotypes into two main groups, the grouping is not related to the geographic origin of genotypes. The grouping is suggested due to long-term cultivation and trade distribution. Based on the genetic population structure of the observed shallot populations, Solo population has the highest genetic diversity (I= 0.44, h= 0.31; PLP= 77.39%) and Solok population has the lowest genetic diversity (I= 0.37, h= 0.24 dan PLP= 67.83%).
The genotypes of shallot that have good characters for shallot breeding program in Indonesia are Bangkok and Sembrani varieties because both genotypes have biggest bulb diameter (7.54 cm and 4.99 cm). Furthermore, genotypes from Pekanbaru and Nganjuk because both genotypes have most number of bulb (27 blub/clump and 20 bulb/clump). Bangkok and Sembrani varieties can be crossed with Pekanbaru and Nganjuk genotypes because those genotypes have largest genetic distance. Those genotypes can be used as a potential parents for shallot breeding program in Indonesia.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang
–
Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
KERAGAMAN GENETIK BAWANG MERAH (
Allium cepa
L.)
BERDASARKAN MARKA MORFOLOGI DAN ISSR
VEBRITA SARI
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biologi Tumbuhan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PRAKATA
Puji dan syukur penulis ucapkan kepada Allah subhanahu wa ta’ala atas segala rahmat dan karunia-Nya, sehingga tesis yang berjudul “Keragaman Genetik Bawang Merah (Allium cepa L.) Berdasarkan Marka Morfologi dan ISSR” dapat diselesaikan, sebagai syarat utama untuk memperoleh gelar Magister Sains pada Mayor Biologi Tumbuhan, Pascasarjana Institut Pertanian Bogor. Penelitian ini dilaksanakan sejak bulan Oktober 2014 hingga Sebtember 2015.
Penulis menyampaikan terima kasih dan apresiasi kepada 1) Dosen pembimbing (Dr Ir Miftahudin, MSi dan Prof Dr Ir Sobir, MSi) yang telah memberikan nasehat, saran, motivasi, waktu luang untuk konsultasi, serta solusi dari setiap permasalahan yang dihadapi penulis selama melaksanakan penelitian dan penyusunan tesis ini; 2) Dosen penguji luar komisi (Dr Ir Utut widyastuti, MSi); 3) Ketua Program Studi Biologi Tumbuhan (Dr Ir Miftahudin, MSi) atas kesediaannya untuk menguji dan mengoreksi penulisan tesis ini agar menjadi lebih baik; 4) DIKTI atas Beasiswa Pendidikan Pascasarjana Dalam Negeri tahun 2013 yang telah diberikan selama menempuh pendidikan; 5) Kepala LPPM-PKHT IPB Dr Ir Darda Efendi, MSi atas fasilitas yang telah diberikan selama penelitian; 6) Bapak Awang sebagai staf di kebun percobaan PKHT-IPB; 7) Mbak Sulassih, MSi sebagai staf peneliti di Pusat Kajian Hortikultura Tropika (PKHT) yang telah banyak membantu dan membimbing penulis; 8) seluruh dosen dan staf Program Studi Biologi Tumbuhan; 9) teman seperjuangan baik teman dari Program Studi Biologi Tumbuhan Pascasarjana IPB 2013 dan teman-teman seperantauan dari Riau atas kebersamaan yang berharga dan penuh kenangan serta 10) pihak lainnya yang tidak tersebutkan.
Ungkapan terima kasih yang tak terhingga penulis sampaikan kepada orang tua tersayang (Ayah Ir Zulkifli H dan Mamah Rahmi Hidayati), mertua (Mama Dahlia) dan adik-adik (Arief Munandar, ST dan Zidef Rizky), Ibu motivator saya (Dr Fitmawati Sofyan, MSi) serta seluruh keluarga besar atas segala doa, dukungan, motivasi dan kasih sayangnya selama ini. Secara khusus dan istimewa ucapan terima kasih penulis sampaikan kepada sahabat jiwa, suami tercinta Brigadir Rio Andria, SH yang selalu memberikan dukungan dalam segala hal, pengertian dan kesabaran yang luar biasa hingga studi yang panjang ini dapat terselesaikan. Semoga tesis ini selalu bermanfaat bagi kemajuan ilmu pengetahuan selanjutnya.
DAFTAR ISI
DAFTAR TABEL xv
DAFTAR GAMBAR xv
DAFTAR LAMPIRAN xvi
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
TINJAUAN PUSTAKA 3
Botani Bawang Merah (Allium cepa L.) 3
Varietas Bawang Merah di Indonesia 6
Marka Morfologi 7
Inter Simple Sequences Repeat (ISSR) 7
METODE PENELITIAN 9
Waktu dan Tempat Penelitian 9
Bahan Tanaman 9
Pengamatan Morfologi 10
Isolasi DNA Total 11
Elektroforesis 11
Amplifikasi DNA 12
Analisis Data 13
HASIL DAN PEMBAHASAN 15
HASIL 15
Keragaman Morfologi Pada Bawang Merah 15
Analisis Hubungan Genetik Bawang Merah Berdasarkan Marka Morfologi
19 Keragaman Genetik Bawang Merah Berdasarkan Marka ISSR 20 Analisis Hubungan Genetik Bawang Merah Berdasarkan Marka ISSR 22 Struktur Genetika Populasi Bawang Merah Asal Indonesia 23
PEMBAHASAN 24
KESIMPULAN 28
DAFTAR PUSTAKA 29
LAMPIRAN 33
DAFTAR TABEL
1 Varietas bawang merah yang telah dilepas Kementrian RI 6
2 Genotipe bawang merah yang dikoleksi PKHT 9
3 Karakter pengamatan morfologi pada bawang merah 10 4 Primer ISSR yang digunakan dalam penelitian 12
5 Tahapan PCR menggunakan primer ISSR 12
6 Profil pita hasil amplifikasi dari 34 genotipe bawang merah menggunakan primer ISSR
21
7 Struktur genetika populasi pada 8 populasi bawang merah di Indonesia 24
DAFTAR GAMBAR
1 Morfologi bawang merah (a) umbi; (b) bunga dan (c) daun 5 2 Kemampuan berbunga. (A) tidak berbunga pada varietas Manjung; (B)
berbunga pada genotipe asal Cirebon 15
3 Variasi pada tingkat kepatahan daun. (1) Absent or weak pada varietas Sembrani; (2) Intermediet pada varietas Kuning; (3) Strong pada genotipe asal Solo 3
16
4 Variasi panjang daun. (A) pendek pada genotipe Solo 2; (B) sedang pada varietas Tuk-tuk; (C) panjang pada varietas Bangkok 16 5 Variasi bentuk umbi dan bentuk leher umbi (garis kuning). (A) Bentuk
umbi flat-globe dan bentuk leher umbi broad pada varietas Sembrani; (B) Bentuk umbi rhomboid dan bentuk leher umbi narrow pada varietas Tuk-tuk; (C) Bentuk umbi broad-oval dan bentuk leher umbi medium pada varietas Trisula; (D) Bentuk umbi globe dan bentuk leher umbi medium
pada varietas Batu Ijo; (E) Bentuk umbi broad-elliptic dan bentuk leher
umbi very narrow pada genotipe Solo 3 17
6 Variasi bentuk ujung batang umbi. (A) flat pada varietas Sembrani; (B)
strongly-sloping pada genotipe Solo 3; (C) slightly sloping pada varietas Batu Ijo; (D) rounded pada genotipe Vietnam 3
17
7 Variasi bentuk ujung akar umbi. (A) round pada varietas Batu Ijo; (B)
weakly-tapered pada genotipe Cirebon; (C) strongly pada genotipe Solo 3; (D) flat pada varietas Sembarani; (E) depressed pada genotipe Bantaeng
8 Variasi jumlah siung umbi. (A) sedikit (2-5 siung); (B) sedang (6-9 siung); (C) banyak (>10 siung)
18
9 Variasi posisi diameter umbi. (A) mendekati ujung batang; (B) di bagian tengah; (C) mendekati ujung akar (tanda panah kuning)
18
10 Variasi warna kulit umbi. (A) merah; (B) merah keunguan; (C) merah kecoklatan; (D) merah tua; (E) coklat kekuningan
18
11 Variasi diameter umbi. (A) kecil; (B) sedang; (C) besar 19 12 Dendogram hasil analisis 34 genotipe bawang merah berdasarkan ciri
morfologi. V = Varietas yang sudah dilepas; BM = Koleksi PKHT
19
13 Matrix plot hasil Principal Component Analysis berdasarkan ciri morfologi 34 genotipe bawang merah, terbentuk dua kelompok yaitu kelompok I dan II. V = Varietas yang sudah dilepas; BM = Koleksi PKHT
20
14 Elektroforegram hasil amplifikasi Primer ISSRred 4 (CAG)6G sebagai representasi pita dengan persentase pita polimorfik tinggi. Genotipe (1) Solo 8, (2) Solo 9, (3) Varietas Trisula, (4) Bilitar, (5) Kediri, (6) Aceh, (7) Pekanbaru, (8) Solok, (9) Bukittinggi, (10) Varietas Brebes, (11) Varietas Batu Ijo, (12) Varietas Sembrani, (13) Varietas Bangkok, (14) Vietnam 1, (15) Vietnam 2, (16) Cirebon, (17) Bantaeng, (18) Tanduyung, (19) Solo 1, (20) Solo 3, (21) Solo 4 dan (22) Tungganamo
21
15 Dendogram hasil analisis 34 genotipe bawang merah berdasarkan 13 primer ISSR. V = Varietas yang sudah dilepas; BM = Koleksi PKHT
22
16 Matrix plot hasil Principal Component Analysis 34 genotipe bawang merah berdasarkan 13 primer ISSR. V = Varietas yang sudah dilepas; BM = Koleksi PKHT
23
DAFTAR LAMPIRAN
1 Karakter morfologi yang diamati pada bawang merah sebagai penjelasan untuk Tabel 3 (IPGRI 2001; UPOV 2008)
34
2 Proporsi karakter polimorfik (kualitatif dan kuantitatif) hasil pengamatan morfologi 34 genotipe bawang merah yang dianalisis
36
PENDAHULUAN
Latar Belakang
Bawang merah (Allium cepa L.) merupakan salah satu tanaman hortikultura penting yang multiguna, baik di Indonesia maupun di negara lainnya setelah tomat dan semangka (FAOSTAT 2015). Bawang merah dapat dimanfaatkan sebagai bumbu masakan (flavor), sayuran (acar dan salad) dan produk olahan contohnya bawang goreng (Chyau dan Mau 2001). Saat ini ekstrak bawang merah telah dikembangkan sebagai obat tradisional (antimikroba, antikanker dan antiinflamasi) (Motlagh et al. 2011). Sebagai komoditas hortikultura yang bermanfaat dan bernilai ekonomi tinggi, bawang merah dibudidayakan hampir di seluruh wilayah Indonesia. Potensi pengembangan bawang merah di Indonesia masih terbuka lebar, baik untuk kebutuhan dalam negeri maupun luar negeri. Namun produktivitas bawang merah di Indonesia saat ini masih rendah (10.16 ton ha-1) jika dibandingkan dengan negara lainnya seperti Cina (20 ton ha-1) dan India (15 ton ha-1) (BPS 2015a).
Peningkatan penduduk dan penurunan luas lahan penanaman menuntut tersedianya varietas baru yang berdaya hasil tinggi. Pengembangan varietas unggul merupakan salah satu strategi untuk meningkatkan produktivitas bawang merah di Indonesia. Dalam pengembangan varietas unggul dibutuhkan informasi keragaman genetik (plasma nutfah) yang cukup di antaranya informasi varietas dengan produktivitas tinggi, toleran terhadap cekaman biotik maupun abiotik, serta memiliki mutu umbi yang diterima oleh konsumen. Kondisi agroekosistem Indonesia yang beragam dapat menyebabkan tingginya keragaman genetik bawang merah sehingga melahirkan varietas-varietas lokal. Varietas lokal bawang merah Indonesia merupakan sumber plasma nutfah yang penting untuk tujuan pemuliaan. Namun penelitian dan informasi tentang keragaman genetik bawang merah lokal Indonesia masih terbatas. Pusat Kajian Hortikultura Tropika (PKHT) Institut Pertanian Bogor (IPB) memiliki koleksi genotipe bawang merah dari berbagai daerah di Indonesia yang belum dianalisis keragaman genetiknya, untuk itu perlu dilakukannya penelitian ini.
Analisis keragaman genetik dari setiap sumber plasma nutfah yang tersedia perlu dilakukan untuk mendapatkan data deskripsi atau karakter spesifik dari masing-masing genotipe baik secara morfologi maupun molekuler. Informasi keragaman genetik diperlukan untuk mengetahui kemiripan atau hubungan genetik antar genotipe. Hubungan genetik atau jarak genetik tersebut dapat digunakan sebagai dasar untuk menentukan genotipe tetua yang akan digunakan dalam proses pemuliaan bawang merah. Persilangan antar genotipe yang memiliki jarak genotipe terjauh akan memberikan hasil terbaik pada tanaman menyerbuk silang seperti bawang merah (Degewione et al. 2011).
Inter Simple Sequences Repeat (ISSR) yang dikembangkan dari wilayah mikrosatelit. Keunggulan ISSR antara lain: tidak dipengaruhi oleh kondisi lingkungan dan musim, tidak membutuhkan informasi sekuen terlebih dahulu, berbasis Polymerase Chain Reaction (PCR), DNA yang digunakan lebih sedikit (5-50 ng per reaksi), wilayah sekuen tersebar diseluruh genom sehingga menghasilkan pola polimorfisme lebih tinggi dibandingkan dengan teknik
Random Amplified Polymorphic DNA (RAPD) dan dapat diaplikasi pada tingkat takson terendah (spesies dan varietas) (Son et al. 2012; Syahruddin 2012; Sulassih
et al. 2013).
Tujuan Penelitian
Penelitian ini bertujuan untuk: 1) menganalisis keragaman genetik dari 34 genotipe bawang merah koleksi PKHT-IPB; 2) menganalisis struktur genetika populasi bawang merah asal Indonesia dan 3) seleksi genotipe potensial sebagai tetua unggul. Penelitian ini diharapkan dapat dijadikan informasi dasar dalam pengelolaan sumber daya genetik dan pemuliaan bibit unggul bawang merah Indonesia.
TINJAUAN PUSTAKA
Botani Bawang Merah (Allium cepa L.)
Marga Allium merupakan tanaman herba perennial berumah satu (monoecious) dengan karakteristik morfologi memiliki umbi berlapis, berdaun sempit, bunga dengan 6 sepal atau tidak ada sepal dan memiliki aroma juga rasa yang unik karena kandungan sulfurnya (Li et al. 2010). Informasi mengenai karakter morfologi, anatomi dan molekuler marga Allium telah banyak dilaporkan. Namun karena kemiripan morfologi antar spesies dan data yang tersedia sangat bergantung pada sampel herbarium, masih banyak kesenjangan informasi mengenai taksonomi antar spesies yang berbeda (interspesies) dan dalam spesies yang sama (intraspesies), diferensiasi dan evolusi dalam marga Allium. Untuk itu penelitian mengenai marga Allium masih terus dilakukan hingga sekarang (Samiei
et al. 2015). Posisi marga Allium telah lama menjadi kontroversi, dalam klasifikasi awal angiosperma, Allium merupakan anggota dari suku Liliaceae
kemudian berdasarkan struktur bunga dan data molekulernya, marga Allium
menjadi anggota Amaryllidaceae, sub-suku Alliaceae, suku Allieae (Chase et al.
2009). Marga Allium merupakan salah satu marga terbesar dengan anggota lebih dari 900 spesies. Allium memiliki pusat keanekaragaman di daerah Timur Mediterania, Asia Tenggara dan Asia Tengah (Rabinowitch dan Currah 2002). Sebagian besar spesies dari anggota marga Allium dibudidayakan secara luas di dunia untuk dimanfaatkan sebagai rempah-rempah, sayuran, obat-obatan dan tanaman hias (Samiei et al. 2015).
Bawang merah (Allium cepa L.) merupakan salah satu spesies dari marga
Allium yang multiguna dan bernilai ekonomi tinggi. Bawang merah sangat populer dalam dunia kuliner yaitu sebagai bumbu masakan, saat ini bawang merah sedang dikembangkan sebagai obat tradisional. Bawang merah memiliki nama lain yang populer yaitu shallot. Selama beberapa tahun nama latin bawang merah dalam dunia taksonomi mengalami sedikit kekeliruan. Sebelumnya bawang merah diberi nama latin Allium ascalonicum karena merupakan salah satu spesies dari marga Allium yang tumbuh liar. Helm (1956) mengklasifikasikan shallot ke dalam takson Allium cepa, berdasarkan bentuk umbi dan pertumbuhannya. Helm (1956) mengelompokkan Allium cepa menjadi 4 varietas botani antara lain (1) var. cepa (common onion); (2) var. vivaparum (top onion); (3) var. aggregatum (multiplier onion atau shallot) dan (4) var. cepiform (the shallot-like). Jones dan Mann
auct. non strand; Allium cepa spp. orientale Kazak.; Allium cepa var. ascalonicum
Aggregatum dan satu spesies dengan bawang bombay (onion). Bawang merah dapat menghasilkan keturunan fertil jika disilangkan dengan bawang bombay, selain itu sitologi dan morfologi keduanya sangat mirip, sehingga keduanya dikelompokkan dalam satu spesies Allium cepa dengan nama latin Allium cepa L. Aggregatum group, namun nama latin yang sering digunakan saat ini adalah
Allium cepa L. (Rabinowitch dan Currah 2002).
Morfologi bawang merah telah banyak diteliti dan dilaporkan oleh para ahli taksonomi. Bawang merah merupakan tanaman herba perennial dengan tinggi tanaman mencapai 100 cm dan membentuk rumpun. Perakarannya berupa akar serabut namun tidak terlalu panjang. Daun bawang merah berwarna hijau muda sampai hijau tua, berbentuk bulat panjang dan berlubang seperti pipa (Gambar 1c). Bagian ujung daun bawang merah runcing sedangkan pangkalnya melebar memeluk batang semu (pseudostem). Pembentukan primordial daun dimulai dengan tonjolan pada permukaan atas umbi yang akan berkembang menjadi daun. Primordial daun berikutnya tumbuh di sisi berlawanan (the opposite side) dari daun sebelumnya. Setiap daun yang baru akan tumbuh di dalam daun sebelumnya sehingga daun pertama akan menyelimuti daun yang baru secara konsentris. Percabangan pada bawang merah merupakan hasil dari hilangnya dominansi apikal, dimana terjadi inisiasi lateral setelah perkembangan dua atau tiga daun. Pada titik ini meristem apikal terbagi menjadi dua bagian sehingga terbentuklah percabangan yang akan menumbuhkan daun-daun baru dan tunas lateral (Krontal
et al. 1998; Rabinowitch dan Currah 2002).
Bawang merah dapat menghasilkan bunga tetapi sangat sulit bahkan terkadang tidak menghasilkan biji. Inflorescence (perbungaan) bawang merah bersifat majemuk terdiri dari banyak rangkaian bunga dan berbentuk seperti payung yang disebut umbels (Gambar 1b). Umbels dapat terdiri dari 50 sampai 200 bunga yang tersususun melingkar. Tangkai rangkaian bunga lebih tinggi dibandingkan daun dan setiap kuntum bunga memiliki tangkai tetapi lebih pendek. Bunga bawang merah termasuk bunga sempurna dengan setiap bunga terdiri atas benang sari dan kepala putik. Biasanya terdiri atas 5 sampai 6 benang sari dan satu buah putik dengan kelopak bunga berwarna hijau bergaris putih, serta bakal buah duduk di atas bunga membentuk suatu bangun seperti kubah. Penyerbukan pada bunga bawang merah bersifat open-pollination yaitu dapat menyerbuk antar bunga dalam satu umbel atau antar umbel, antar bunga berbeda tanaman pada satu spesies bawang merah (intraspesifik) dan antar bawang merah dengan spesies yang berbeda tapi masih dalam satu marga Allium (interspesifik). Penyerbukan silang antar jenis dan antar varietas dapat terjadi secara alami, sehingga peluang munculnya varietas baru cukup tinggi.
Proses pembungaan pada bawang merah sama seperti anggota marga
Allium lainnya. Selama proses transisi dari perbanyakan secara vegetatif menuju ke generatif, terjadi perubahan pertumbuhan monopodial menjadi simpodial pada batang tetapi tidak mempengaruhi pertumbuhan dan perkembangan tanaman secara keseluruhan. Buah berbentuk bulat dengan ujung tumpul membungkus biji dan berjumlah 2 sampai 3 butir. Biji berbentuk pipih, biji muda berwarna bening atau putih, tetapi setelah masak menjadi hitam (Krontal et al. 1998; Brewster 2008; Block 2010).
(Allium cepa L.). Berbeda dengan bawang bombay, umbi bawang merah berukuran lebih kecil dan membelah secara lateral sehingga jumlah siung yang dihasilkan lebih banyak hingga mencapai 30 siung dalam satu rumpun. Proses pembentukan umbi pada bawang merah dipengaruhi oleh lamanya fotoperiode dan suhu, sehingga setiap varietas memiliki kondisi lingkungan yang berbeda dalam membentuk umbi. Umbi bawang merah merupakan umbi lapis dengan bentuk dan warna yang bervariasi (Gambar 1a). Umbi bawang merah mengandung senyawa antosianin (cyanidin dan peodin) dan flavonoid (quercitin) yang memberikan aroma juga rasa yang unik. Variasi warna umbi penting dalam menentukan preferensi konsumen dan juga dalam klasifikasi antar varietas. Pigmen pada umbi bawang merah tidak hanya mengendalikan warna umbi tetapi juga mempengaruhi daya simpan dan ketahanan terhadap penyakit. Setiap umbi tunggal berisi titik tumbuh (1-2) dan setiap umbi dilapisi 1 sampai 3 kulit pelindung. Dormansi umbi bawang merah berlangsung selama 2.5-5 bulan pada suhu 27-32 ºC (Arifin et al. 1999; Rabinowitch dan Currah 2002; Brewster 2008).
Gambar 1 Morfologi bawang merah (a) umbi; (b) bunga dan (c) daun. (dokumentasi pribadi)
Bawang merah telah lama dibudidayakan di beberapa daerah di Indonesia dengan kondisi agroekosistem yang beragam. Berdasarkan data Dirjen Hortikultura (2015) terdapat 34 sentral produksi bawang merah yang tersebar di beberapa wilayah Indonesia. Budidaya bawang merah di Indonesia secara umum memerlukan bulan kering sekitar 4 sampai 5 bulan dengan musim tanam optimal pada akhir musim hujan (Maret-April) atau akhir musim kemarau (Mei-Juni). Curah hujan untuk budidaya bawang merah di Indoneisa berkisar 1000 sampai 1500 mm tahun-1. Suhu lingkungan tanam bawang merah berkisar antara 25-32 ºC dengan pH tanah 5.6-6.5. Tanah yang digunakan memiliki kesuburan dan drainase yang baik, tekstur tanah remah, lempung berpasir dan tidak ternaungi (Erythrina 2012).
Varietas Bawang Merah di Indonesia
Varietas merupakan subdivisi dari spesies berdasarkan International Code of Botanical Nomenclature. Varietas unggul dapat berasal dari varietas lokal, varietas liar, varietas introduksi, galur homozigot juga mutan yang mempunyai potensi unggul dan sesuai dengan target pemuliaan yang diinginkan. Varietas unggul adalah varietas yang memiliki karakter unik, seragam dan stabil (distictness, uniformity dan stability) juga telah lulus uji seleksi dan daya hasil. Untuk menghasilkan varietas unggul dengan sifat-sifat yang diinginkan harus melalui prosedur pemuliaan yang sistematik. Koleksi plasma nutfah sangat berperan dalam program pemuliaan varietas unggul tentunya informasi keragaman genetik sangat diperlukan (Brown dan Caligari 2008). Bawang merah merupakan komoditas unggulan utama yang dikembangkan di Indonesia. Saat ini telah dilepas 24 varietas bawang merah unggul oleh Kementrian Pertanian Republik Indonesia (Tabel 1) (Dirjen Hortikultura 2012).
Tabel 1 Varietas bawang merah yang telah dilepas Kementan RI
No. Varietas No. Kepmentan Asal
1 Bima Brebes 594/Kpts/TP.240/8/1984 Lokal Brebes
2 Medan 595/Kpts/TP.240/8/1984 Lokal Samosir
3 Keling 596/Kpts/TP.240/8/1984 Lokal Maja
4 Bima Maja Cipanas 597/Kpts/TP.240/8/1984 Lokal Cipanas
5 Super Philip 65/Kpts/TP.240/2/2000 Lokal Nganjuk
6 Bauji 66/Kpts/TP.240/2/2000 Introduksi dari Philipina
7 Bangkok 68/Kpts/TP.240/2/2000 Introduksi dari Thailand
8 Kramat-1 225/Kpts/TP.240/4/2001 Maja Cipanas x Bawang Bombay
9 Karamat-2 226/Kpts/TP.240/4/2001 Maja Cipanas x Bawang Bombay
10 Kuning 227/Kpts/TP.240/4/2002 Lokal Brebes
11 Tiron 498/Kpts/TP.240/8/2003 Bantul, DIY
12 Keta Monca 529/Kpts/PD.210/10/2004 Bima, NTB
13 Batu Ijo 368/Kpts/LB.240/6/2004 Batu Malang, Jatim
14 Palasa 480/Kpts/LB.240/6/2004 Parigi Moutg, Sulteng
15 Tinombo 481/Kpts/LB.240/6/2004 Parigi Moutg, Sulteng
16 Tuk-Tuk 361/Kpts/SR.120/5/2006 PT.East West Seed Philipina
17 Sembrani 304/Kpts/SR.120/5/2007 Balitsa Lembang
18 Katumi 305/Kpts/SR.120/5/2007 Balitsa Lembang
19 Manjung 703/Kpts/SR.120/5/2007 Pamekasan, Jatim
20 Biru Lancor 2830/Kpts/SR.120/7/2009 Probolinggo, Jatim
21 Lembah Palu 11843/Kpts/SR.120/4/2011 Lembah Palu, Sul
22 Rubaru 2525/Kpts/SR.120/5/2011 Sumenep, Jatim
23 Trisula 4580/Kpts/SR.120/11/2011 Balitsa Lembang
24 Pancasona 4581/Kpts/SR.120/11/2011 Balitsa Lembang
25 Mentes 4582/Kpts/SR.120/11/2011 Balitsa Lembang
berwarna kuning pucat dan varietas medan berwarna merah tua (Erythrina 2012). Selain varietas yang telah dilepas oleh Kementan RI, masih banyak genotipe bawang merah lokal yang belum dikarakterisasi dan diidentifikasi keragaman genetiknya sebagai potensi sumber daya genetik bawang merah Indonesia.
Marka Morfologi
Keragaman genetik tanaman dapat dianalisis menggunakan marka morfologi dan molekuler. Marka morfologi didasarkan pada karakter fenotipe yang bisa langsung diamati, baik pada organ vegetatif maupun organ generatif (Sulassih 2011). Karakter morfologi merupakan hasil interaksi antara gen dan lingkungan, sehingga memiliki kelemahan yaitu bersifat tidak konsisten. Perbedaan lingkungan adaptasi atau kondisi egroekosistem yang beragam akan mempengaruhi keragaman karakter morfologi suatu tanaman. Karakter morfologi terbagi menjadi dua jenis yaitu karakter yang bersifat kuantitatif dan karakter yang bersifat kualitatif (Syahruddin 2012).
Karakter kuantitatif merupakan karakter yang tidak dapat dibedakan secara sederhana namun harus diukur dengan alat ukur tertentu yang hasilnya bersifat kuantitatif karena karakter kuantitatif dikendalikan oleh banyak gen. Karakter kuantitatif yang digunakan untuk menganalisis keragaman genetik bawang merah diantaranya tinggi tanaman, panjang daun, lebar daun, bobot basah umbi, diameter umbi dan tinggi umbi. Sedangkan karakter kualitatif merupakan karakter yang dapat dibedakan secara tegas dan sederhana tanpa ada proses pengukuran karena karakter kualitatif dikendalikan oleh gen yang sederhana (umumnya satu gen). Karakter kualitatif yang digunakan untuk menganalisis keragaman genetik bawang merah diantaranya tingkat kepatahan daun, warna daun, bentuk umbi, bentuk leher dan batang umbi, warna kulit dan warna daging umbi. Karakter warna dan bentuk dikendalikan oleh gen sederhana yang dapat dipengaruhi oleh lingkungan sehingga bersifat tidak konsisten dan menghasilkan informasi yang sedikit bias (Syahruddin 2012).
Analisis keragaman menggunakan marka morfologi tidak mudah untuk tanaman yang berada pada takson di bawah spesies (varietas). Hasil analisis morfologi yang didasarkan dari kemiripan karakter diharapkan dapat menggambarkan hubungan kekerabatan diantara takson atau individu suatu tanaman (Rustiami et al. 2011). Analisis keragaman genetik bawang merah lokal Indonesia masih terbatas. Namun, analisis keragaman genetik bawang merah pada 10 genotipe Allium cepa L. di India menggunakan marka morfologi telah dilakukan oleh Akter et al. (2015). Sepuluh genotipe bawang merah asal India tersebut dilaporkan memiliki variasi pada karakter umbi dan daunnya.
Inter Simple Sequences Repeat (ISSR)
Marka molekuler telah banyak digunakan untuk mengungkap keragaman genetik suatu tanaman dengan menggunakan DNA. Penggunaan marka molekuler dapat mendukung atau menutupi kelemahan dari marka morfologi, khususnya pada tingkat takson rendah (spesies dan varietas). Kelemahan dari marka morfologi yaitu bersifat tidak konsisten, polimorfisme yang dihasilkan cukup rendah dan dipengaruhi oleh peristiwa epistasis atau pleiotropi (Weising et al.
Reaction) atau non PCR. Marka yang berbasis non PCR yaitu RFLP (Restriction Fragment Length Polymorphism) sedangkan marka yang berbasis PCR diantaranya RAPD (Random Amplified Polymorphism DNA), AFLP (Amplified Fragment Length Polymorphism), SSR (Simple Sequence Repeat) dan ISSR (Inter Simpel Sequences Repeat). Marka molekuler yang digunakan diharapkan menghasilkan polimorfisme yang tinggi, terdapat diseluruh genom, biaya yang dibutuhkan sedikit, analisis data cepat juga mudah dan bersifat reproducible
(Kumar 2009).
Marka molekuler yang digunakan pada penelitian ini adalah Marka Inter Simpel Sequences Repeat (ISSR). Marka ISSR merupakan marka yang dikembangkan dari daerah wilayah mikrosatelit atau Simple Sequences Repeat
(SSR). SSR merupakan daerah sekuen nukleotida berulang atau STR (Short Tandem Repeat) yang tersebar diseluruh genom sedangkan ISSR merupakan daerah inter-SSR. ISSR merupakan daerah bukan gen yang tidak mengkode protein (non coding region) dan berada diantara dua lokus mikrosatelit. Marka ISSR menggunakan primer tunggal yang pada bagian ujung 3’ dan 5’ terdapat penambahan sekuen nukleotida. Wilayah amplifikasi ISSR yaitu daerah inter-SSR. Produk amplifikasi akan berupa pola pita ganda dan polimorfik yang dapat digunakan untuk studi variasi genetik pada organisme, analisis hubungan kekerabatan, identifikasi genetik tetua, pembentukan klon atau galur dan studi hubungan asal tanaman dengan pusat penyebarannya (Ng dan Tan 2015).
Setiap marka molekuler tentunya memiliki kekurangan dan kelebihan. Marka ISSR memiliki banyak kelebihan yaitu lebih murah, mudah dan cepat digunakan dibandingkan RFLP dan AFLP, sekuennya tersebar diseluruh genom sehingga menghasilkan polimorfisme lebih tinggi dibandingkan RAPD, tidak memerlukan informasi sekuen terlebih dahulu, DNA yang digunakan lebih sedikit (5-50 ng per reaksi), bersifat lebih reproducible dibandingkan marka RAPD dan dapat mengungkap keragaman genetik pada tingkat takson rendah (spesies dan varietas) (Son et al. 2012; Syahruddin 2012; Sulassih et al. 2013). Marka ISSR telah banyak digunakan dalam analisis keragaman genetik tanaman. Marka ISSR telah digunakan untuk mengungkap hubungan kekerabatan pada 13 aksesi marga
METODE PENELITIAN
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan pada bulan Oktober 2014 sampai dengan September 2015. Penanaman bawang merah di Kebun Percobaan Institut pertanian Bogor (IPB) Pasir Sarongge, untuk analisis data morfologi dan molekuler (ISSR) dilakukan di Laboratorium Molekuler Pusat Kajian Hortikultura Tropika (PKHT) IPB Baranangsiang.
Bahan Tanaman
Bawang merah yang digunakan berupa umbi sebanyak 34 genotipe. Genotipe tersebut merupakan koleksi PKHT dari hasil eksplorasi beberapa daerah di Indonesia. Sampel terdiri dari 20 genotipe merupakan koleksi PKHT, 5 genotipe impor Vietnam dengan berbeda generasi tanam (generasi 0, 1 dan 2) dan 9 genotipe merupakan varietas yang telah dilepas oleh Kementrian Pertanian Republik Indonesia (Tabel 1).
Tabel 2 Genotipe bawang merah yang dikoleksi oleh PKHT No. Genotipe Asal Koleksi No. Genotipe Asal Koleksi
1 BM ACH Aceh 18 BM SL4 Solo, Jateng
Keterangan: Var. = Varietas dan G = Generasi
pada minggu ketiga setelah tanam menggunakan pupuk urea dan KCl (1:1) dengan dosis 10 gr per individu tanaman. Penyiraman dilakukan satu kali setiap dua hari, fumigasi dengan Previcur (1-2 cc per Liter) dua kali selama proses penanaman dan penyiangan rumput atau gulma dilakukan satu kali seminggu. Pemanenan dilakukan pada 60-90 hari tanam, dengan ciri-ciri tanaman cukup tua dimana 60-90% batang lemas atau rebah, daun menguning dan umbi tersembul kepermukaan tanah.
Pengamatan Morfologi
Pengamatan karakter morfologi dan agronomi menggunakan panduan deskriptor Allium spp. (IPGRI 2001) dan deskriptor varietas Allium (UPOV 2008) dengan beberapa modifikasi karakter sehingga didapatkan 24 karakter morfologi yang diamati (Tabel 3 dan penjelasan pada Lampiran 1).
Tabel 3 Karakter pengamatan morfologi pada bawang merah No. Karakter Sifat Karakter
1 Umur panen 1=genjah (<59 hari); 2=sedang (60-90 hari);
3=dalem (>91 hari)
2 Kemampuan berbunga 1=berbunga; 2=tidak berbunga
3 Jumlah bunga dalam karangan
1=banyak (>31); 2=sedikit (<30); 3=tidak ada 4 Warna mahkota bunga 1=putih; 2=krem; 3=kuning;3=hijau
5 Warna anther bunga 1=kuning; 2=orange; 3=orange kehijauan; 4=hijau; 5=abu-abu 6 Tinggi tanaman 1=pendek (< 15 cm); 2=sedang (16-45 cm); 3=tinggi (>46 cm)
7 Warna daun 1=hijau muda; 2=hijau kekuningan; 3=hijau tua
8 Kerapatan daun 1=tinggi; 2=sedang; 3=rendah
9 Panjang daun 1=pendek (<15 cn); 2=sedang (16-30 cm);
3=panjang (> 31 cm)
10 Jumlah daun 1=sedikit (<3 helai); 2=sedang (4-6 helai); 3=banyak (>7 helai)
11 Tingkat kepatahan daun 1=lemah; 2=sedang;3= kuat
12 Diameter daun 1=kecil (<0,5 cm); 2=sedang (0,6-0,8 cm); 3=lebar (>0,9 cm) 13 Bentuk umbi 1=flat; 2=flat-globe; 3=rhomboid; 4=broad-oval; 5=globe;
6=broad-elliptic; 7=ovate-elongated oval; 8=spindle; 9= high top; 10=lainnya
14 Jumlah siung umbi 1=sedikit (1-3 siung); 2=sedang (4-9 siung); 3=banyak (>10 siung)
15 Warna kulit umbi 1=merah; 2=merah muda; 3=merah keunguan; 4=merah tua; 5=merah kecoklatan; 6=coklat muda; 7=coklat kekuningan 16 Ketebalan kulit umbi 1=tipis; 2=sedang ; 3=tebal
17 Diameter umbi 1=kecil (<1 cm); 2=sedang (2-3 cm); 3=besar (>4 cm) 18 Posisi diameter maksimum muda; 5=merah kecoklatan; 6=merah keunguan; 7=merah tua 21 Bobot umbi basah/rumpun 1=ringan (<9 gr); 2=sedang (10-20 gr); 3=berat (>21 gr) 22 Tinggi umbi 1=pendek (<1 cm); 2=sedang (2-3 cm); 3=tinggi (>4 cm) 23 Bentuk ujung akar umbi 1=depressed; 2=flat; 3=round; 4=weakly-tapered;
5=strongly-tapered
24 Bentuk ujung batang umbi 1=depressed; 2=flat; 3=slightly-raised; 4=rounded;
Isolasi DNA Total
Isolasi DNA total 34 genotipe bawang merah menggunakan metode Cetyl Trimethyl Ammonium Bromide (CTAB) oleh Doyle dan Doyle (1987) dengan sedikit modifikasi (Son et al. 2012). Sampel daun muda dipotong sebanyak 0.2 g, kemudian digerus menggunakan mortar dengan menambahkan sedikit PVP (polyvinyl-pyrrolidone) dan pasir kuarsa sampai menjadi bubuk. Setelah halus ditambahkan buffer lysis secukupnya (CTAB 10%, EDTA (Ethylene Diamine Tetra-acetic Acid) 0.5 M pH 8.0, Tris-HCl 1 M pH 8.0, NaCl 5 M dan aquadest steril) dan dipindahkan ke dalam tabung eppendorf kemudian diinkubasi dalam
water bath (65 °C) selama 60 menit, dimana setiap 15 menit dibolak-balik. Setalah diinkubasi, ditambahkan larutan CIAA (Chloroform Iso-amyl Alkohol
24:1) sebanyak satu kali volume larutan sampel, kemudian larutan dihomogenisasi menggunakan vortex selama satu menit. Untuk memisahkan DNA dengan komponen sel lainnya sampel disentrifuse (11.000 rpm) selama 10 menit.
Hasil sentrifuse yang berupa supernatan dipindahkan ke tabung eppendorf
baru secara perlahan kemudian ditambahkan isopropanol dingin sebanyak satu kali volume supernatan yang didapatkan, sambil dibolak-balik tiga kali dan diinkubasi dalam freezer selama satu malam agar proses presipitasi DNA sempurna. Setelah diinkubasi semalaman, larutan DNA disentrifuse (11.000 rpm) selama 15 menit dan supernatan dibuang untuk menghasilkan endapan DNA (pellet DNA) di ujung tabung. Pellet DNA dicuci dengan menggunakan 200 µl ethanol 70% dingin kemudian disentrifuse kembali (11.000 rpm) selama 10 menit. Larutan alkohol dibuang dan endapan DNA dikeringkan menggunakan desikator. Endapan DNA yang telah kering kemudian ditambahkan 100 µl TE-buffer (Tris -EDTA pH 8.0) sambil disentil-sentil dan disimpan dalam freezer (-20 °C) sebagai stok DNA. DNA total yang didapatkan diuji kualitasnya terlebih dahulu. Uji kualitas DNA total secara kualitatif dan kuantitatif menggunakan DNA lamda dengan cara dielektroforesis dalam gel agarosa yang akan dijelaskan pada tahap elektroforesis.
Elektroforesis
Eletroforesis untuk uji kualitas dan kuantitas DNA total menggunakan 0.8% gel agarosa(0.32 g agarosa TopVisioTM Fermentan dalam40 ml buffer TAE 1x). Gel agarosa dicetak terlebih dahulu sehingga memiliki sumur gel kemudian direndam dalam electrophoresis chamber (Bio-Rad Laboratories, Hercules) yang berisi larutan buffer TAE 1x (100 ml TAE 10x dilarutkan dalam 900 ml aquades
steril). DNA total sebanyak 5 µl yang ditambahkan 2 µl larutan loading dye
dimasukkan ke dalam sumur gel menggunakan pipet mikro bersama 2 µl λ (lamda) DNA (promega) pada sumur yang lain, kemudian running selama 15 menit dengan tegangan 50 volt. Setelah dielektroforesis selama 15 menit, gel agarosa direndam dalam larutan etidium bromida (EtBr) (10%) selama 5 menit kemudian divisualisasikan dengan sinar UV (Ultraviolet transiluminator) dan didokumentasikan menggunakan kamera digital merek Sony Optical Steady-Shot
DSC-W730, 16.1 mega pixels 8x optical zoom.
adalah 5 µl. Sehingga ketika pita DNA yang dihasilkan antara DNA total dengan lamda DNA sama dapat disetarakan bahwa konsentrasi 1 µl DNA adalah 91.4 ng µl-1 (didapatkan dari 457 ng µl-1/ 5 µl). Sementara konsentrasi DNA total yang dibutuhkan untuk tahapan PCR yaitu 20 ng, maka stok DNA total harus diencerkan sebanyak 5x (didapatkan dari 91.4 ng/ 20 ng). Pengenceran 5x (1:5) yaitu 1 µl stok DNA ditambahkan 5 µl air bebas ion. Elektroforesis untuk hasil PCR menggunakan 1.2% gel agarosa (0.48 gr dilarutkan dalam 40 ml buffer TAE 1x) selama 48 menit dengan tegangan 50 volt dan menggunakan DNA ladder 1 kb (promega) sebagai pembanding
Amplifikasi DNA
Amplifikasi DNA dengan 13 primer ISSR (Tabel 4) menggunakan mesin PCR-GeneAmp merek Applied Biosystems 2720. Volume total mix PCR yang akan diamplifikasi yaitu sebesar 15.5 µl yang terdiri dari 6 µl master mix Go Taq® Green, 1 µl primer ISSR, 2.5 µl sampel DNA total, dan 6 µl air bebas ion (ddH2O). Proses PCR dibagi dalam beberapa tahap (Tabel 5) yaitu denaturasi awal, denaturasi, penempelan primer (annealing), pemanjangan primer yang komplemen dengan DNA bawang merah (elongation), final extention dan penurunan suhu. Hasil amplifikasi dielektroforesis dalam gel agarosa, divisualisasikan dengan sinar UV dan didokumentasikan menggunakan kamera digital seperti yang telah dijelaskan pada prosedur elektroforesis.
Tabel 4 Primer ISSR yang digunakan dalam penelitian
No. Kode Primer Sekuen Primer (5'-3') Suhu Annealing (°C)
Tabel 5 Tahapan PCR menggunakan primer ISSR
Analisis Data
Data hasil pengamatan morfologi diterjemahkan ke dalam data multistate
dan pita DNA polimorfik hasil elektroforesis diterjemahkan menjadi data biner yaitu satu (1) apabila terdapat pita pada tingkat migrasi yang sama dan nol (0) apabila tidak terbentuk pita. Pengamatan pita dibantu dengan marker ladder (1 kb
promega) yang dinyatakan dalam satuan panjang basa (pb). Analisis hubungan genetik (dendrogram) dan analisis komponen utama (Principal Components Analysis)menggunakan program Numerical Taxonomy and Multivariate Analisys System (NTSys) versi 2.1.1a (Rohlf 2003).
Hasil skoring data pengamatan morfologi dan molekuler ditabulasi dalam matriks data. Dendrogram dikonstruksi berdasarkan nilai kemiripan antar genotipe yang dihasilkan melalui prosedur similarity of qualitative data (SIMQUAL) dengan koefisien kemiripan simple matching (SM) dan analisis Sequential Agglomerative Hierarchial and Nested Clustering (SAHN) menggunakan metode
Unweighted Pair Group Method with Arithmatic Average (UPGMA) pada program NTSys. PCA menggunakan metode SIMINT dengan koefisien CORR (correlation) dan mengekstrak eigenvector dari eigenvalue (EIGEN) utama melalui proses analisis ordination untuk memproyeksikan PCA ke dalam plot matriks dua dimensi pada program NTsys.
Pita hasil amplifikasi DNA dengan ukuran tertentu merupakan sifat fenotipe, sedangkan frekuensi alelnya diasumsikan sebagai ciri genotipe. Hasil pengamatan molekuler dengan marka ISSR pada suatu populasi untuk data biner, frekuensi alel diperoleh berasal dari nilai frekuensi pita yang muncul (p) dan frekuensi pita yang tidak muncul (q atau 1-p). Satu pita yang muncul pada pengamatan elektroforegram hasil amplifikasi DNA dianalogikan sebagai suatu lokus dimana lokus merupakan fragmen DNA berukuran tertentu yang terletak dalam genom (Riupassa 2016). Ada dan tidak adanya pita merupakan dua alel, yang kemunculannya dalam populasi dihitung sebagai frekuensi alel. Analisis parameter genetik populasi diantaranya jumlah alel (Na), jumlah alel efektif (Ne), indeks informasi Shannon (I), keragaman genetik (h) dan persentase lokus polimorfik (PLP) menggunakan Software GenAlEx 6.5-Genetic Analysis in Excel
(Peakall dan Smouse 2012). Rumus dan deskripsi parameter dijelaskan sebagai berikut:
a. Jumlah alel (Na) adalah rata-rata jumlah alel (ada pita amplifikasi) dan alel nol (tidak ada pita). Jumlah alel (Na) disebut pula sebagai alel aktual. b. Jumlah alel efektif (Ne) adalah nilai dugaan variabilitas dari alel aktual
dalam populasi (Kimura dan Crow 1964).
Untuk data biner, p = frekuensi pita yang muncul dan q = 1 - p. c. Indeks informasi Shannon (I) (Lewontin 1975).
�= −1 × [ �� + �� ]
d. Keragaman genetik (h)
ℎ= 1− ( 2 + 2)
Di mana: p = frekuensi pita yang muncul; dan q = 1 - p.
tidak ada pita. Perhitungan menggunakan data skor pita dengan angka 1 atau 0, yang dihitung menurut Ruiz et al. (2000)
hi = 2fi (1 - fi)
Dimana: hi adalah h untuk penanda ke-i; fi adalah frekuensi alel atau pita yang dapat diamplifikasi (muncul pita); dan (1-fi) adalah frekuensi alel nol (tidak muncul pita). Nilai h berkisar dari 0 sampai 0.5.
e. Persentase Lokus Polimorfik
HASIL DAN PEMBAHASAN
HASIL
Keragaman Morfologi Pada Bawang Merah
Hasil pengamatan morfologi menunjukkan bahwa terdapat keragaman morfologi pada 34 genotipe bawang merah koleksi PKHT yang diamati. Karakterisasi yang dilakukan menghasilkan 19 karakter yang beragam dari 24 karakter yang diamati. Keragaman morfologi yang dimaksud antara lain: umur panen, kemampuan berbunga, tinggi tanaman, warna daun, panjang daun, diameter daun, jumlah daun, tingkat kepatahan daun, bentuk umbi, bentuk ujung batang umbi, bentuk ujung akar umbi, warna daging umbi, warna kulit umbi, diameter umbi, posisi diameter maksimum umbi, bentuk leher umbi, tinggi umbi, jumlah siung umbi dan bobot umbi. Proporsi karakter hasil pengamatan morfologi disajikan pada Lampiran 2 dan deskripsi 34 genotipe bawang merah yang dianalisis dijelaskan pada Lampiran 3.
Umur panen genotipe bawang merah yang diamati berkisar antara 55 hari (varietas Batu Ijo) sampai 90 hari dan sebagian besar genotipe memiliki umur panen kategori sedang (60-90 hari). Tinggi tanaman yang didapatkan berkisar antara 23 cm (varietas Ilokos) sampai 51.67 cm (genotiep asal Bantaeng). Secara umum tanaman bawang merah mampu menghasilkan bunga, namun pada penelitian kali ini ditemukan 3 genotipe bawang merah yang tidak mampu menghasilkan bunga yaitu genotipe asal Bukittinggi, Kediri dan varietas Manjung (Gambar 2).
Morfologi daun pada 34 genotipe bawang merah yang diamati memiliki keragaman. Tingkat kepatahan daun pada bawang merah yang diamati bervariasi yaitu absent or weak, intermediet dan strong (Gambar 3). Warna daun yang ditemukan juga bervariasi yaitu hijau kekuningan, hijau muda dan hijau tua. Panjang daun yang ditemukan berkisar antara 21 cm (varietas Ilokos) sampai 49.50 cm (varietas Bangkok) dengan diameter daun berkisar antara 0.47 cm (varietas Trisula dan varietas Kuning) sampai 1 cm (varietas Bangkok) (Gambar 4). Jumlah daun yang ditemukan mulai dari 2 helai (Genotipe Solo 3) sampai 12 helai (varietas Tuk-tuk).
Gambar 2 Kemampuan berbunga. (A) tidak berbunga pada varietas Manjung; (B) berbunga pada genotipe asal Cirebon
Gambar 3 Variasi pada tingkat kepatahan daun. (A) Absent or weak pada varietas Sembrani; (B) Intermediet pada varietas Kuning; (C) Strong
pada genotipe asal Solo 3
Gambar 4 Variasi panjang daun. (A) pendek pada genotipe Solo 2; (B) sedang pada varietas Tuk-tuk; (C) panjang pada varietas Bangkok
Karakter morfologi umbi pada 34 genotipe bawang merah yang diamati memiliki keragaman. Bentuk umbi yang ditemukan bervariasi yaitu flat-globe,
rhomboid, broad-oval, globe dan broad-elliptic dan bentuk leher umbi bervariasi yaitu very narrow, narrow, medium dan broad (Gambar 5). Bentuk ujung batang umbi yang ditemukan yaitu flat, rounded, slightly-sloping dan strongly-sloping
sedangkan bentuk ujung akar umbi yang ditemukan yaitu depressed, flat, round,
V Sem V Kun BM SL3
A B C
BM SL2 V Tuk-Tuk V BGK
weakly-tapered dan strongly (Gambar 6 dan 7). Jumlah siung umbi yang ditemukan berkisar antara 2 siung (genotiep asal Tanduyung) sampai 27 siung (Genotipe asal Pekanbaru) (Gambar 8).
Posisi diameter maksimum umbi yang ditemukan beragam yaitu umbi dengan diameter maksimumnya pada daerah mendekati ujung batang, mendekati ujung akar dan pada bagian tengah (Gambar 9). Warna daging umbi yang ditemukan bervariasi yaitu merah keunguan, merah kecoklatan, merah muda, merah tua, ungu keputihan, ungu muda dan ungu tua sedangkan warna kulit umbi juga bervariasi yaitu merah muda, merah, merah tua, merah kecoklatan, merah keunguan, coklat kekuningan dan coklat tua (Gambar 10). Tinggi umbi yang ditemukan berkisar antara 0.97 cm (genotipe asal Vietnam 4) sampai 6.01 (varietas Bangkok). Bobot basah umbi tertinggi dimiliki oleh genotipe dari varietas Bangkok (57.56 g/rumpun) dan bobot basah umbi terendah dimiliki oleh genotipe varietas Kuning (2.33 g/rumpun). Diameter umbi yang diamati berkisar antara 1.51 cm (genotipe solo 7) sampai 7.54 cm (varietas Bangkok) (Gambar 11).
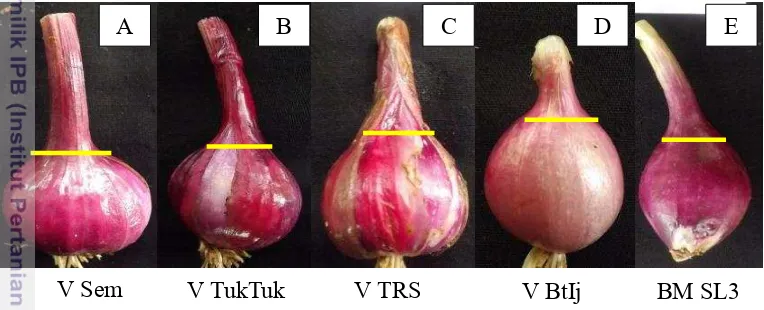
Gambar 5 Variasi bentuk umbi dan bentuk leher umbi (garis kuning). (A) Bentuk umbi flat-globe dan bentuk leher umbi broad pada varietas Sembrani; (B) Bentuk umbi rhomboid dan bentuk leher umbi narrow pada varietas Tuk-tuk; (C) Bentuk umbi broad-oval dan bentuk leher umbi
medium pada varietas Trisula; (D) Bentuk umbi globe dan bentuk leher umbi medium pada varietas Batu Ijo; (E) Bentuk umbi broad-elliptic dan bentuk leher umbi very narrow pada genotipe Solo 3
Gambar 6 Variasi bentuk ujung batang umbi. (A) flat pada varietas Sembrani; (B) strongly-sloping pada genotipe Solo 3; (C) slightly sloping pada varietas Batu Ijo; (D) rounded pada genotipe Vietnam 3
V Sem BM SL3 V BtIj BM VT3
A B C D
V Sem V TukTuk V TRS V BtIj BM SL3
Gambar 7 Variasi bentuk ujung akar umbi. (A) round pada varietas Batu Ijo; (B)
weakly-tapered pada genotipe Cirebon; (C) strongly pada genotipe solo 3; (D) flat pada varietas Sembarani; (E) depressed pada genotipe Bantaeng
Gambar 8 Variasi jumlah siung umbi. (A) sedikit (2-5 siung); (B) sedang (6-9 siung) ; (C) banyak (>10 siung)
Gambar 9 Variasi posisi diameter umbi. (A) mendekati ujung batang; (B) di bagian tengah; (C) mendekati ujung akar (tanda panah kuning)
Gambar 10 Variasi warna kulit umbi. (A) merah ; (B) merah keunguan; (C) merah kecoklatan; (D) merah tua; (E) coklat kekuningan
BM BTE V Sem
V BtIj BM CRI BM SL3
A B C D E
A B C
A B C D E
Kelompok I terdiri dari 27 genotipe yang memiliki kemiripan, 18 genotipe merupakan hasil eksplorasi dari berbagai daerah di Indonesia, 5 genotipe merupakan impor dari luar (Vietnam) dan 4 genotipe lainnya merupakan varietas yang telah dilepas oleh Kementan RI (varietas Brebes, Manjung, Kuning dan Trisula). Kelompok II terdiri dari 7 genotipe yang memiliki kemiripan yaitu varietas Bangkok (impor), varietas Batu Ijo, Sembrani, Ilokos, Tuk-tuk (varietas yang sudah dilepas oleh Kementan RI) dan 2 genotipe lainnya merupakan hasil eksplorasi PKHT (Kediri dan Cirebon).
Analisis PCA 34 genotipe bawang merah berdasarkan karakter morfologi menghasilkan dua kelompok utama (kelompok I dan II) dengan nilai keragaman kumulatif 41.55% (33.41% PC1 dan 8.14% PC2) (Gambar 13). Hasil pengelompokan dengan PCA mendukung dendrogram, namun terdapat sedikit perbedaan pada anggota kelompok antara PCA dengan dendogram. Pada PCA untuk varietas Ilokos termasuk kedalam kelompok I sedangkan pada dendrogram varietas Ilokos termasuk kedalam kelompok II. Kelompok I mengelompok karena memiliki jumlah siung umbi sedikit (1 sampai 4 siung), bentuk leher umbi medium, bobot umbi sedang (10 sampai 20 g per rumpun) dan diameter daun lebar (>0.9 cm). Sedangkan kelompok II berkelompok karena memiliki umur panen <60 hari, tinggi tanaman >30 cm dan bentuk ujung akar umbi round.
Gambar 13 Matrix plot hasil Principal Component Analysis berdasarkan ciri morfologi 34 genotipe bawang merah, terbentuk dua kelompok yaitu kelompok I dan II. V = Varietas yang sudah dilepas; BM = Koleksi PKHT
Keragaman Genetik Bawang Merah Berdasarkan Marka ISSR
berhasil mengamplifikasi DNA dari 34 genotipe bawang merah. Jumlah pita yang dihasilkan berkisar antara 6-16 pita dengan ukuran pita yang bervariasi mulai dari 250 sampai 2000 pb (pasang basa) (Tabel 6). Total jumlah pita yang teramati dan terdeteksi adalah 115 pita, dengan rata-rata jumlah pita per primer yaitu 8.85 atau 9 pita per primer.
Gambar 14 Elektroforegram hasil amplifikasi Primer ISSRred 4 (CAG)6G sebagai representasi pita dengan persentase pita polimorfik tinggi. Genotipe (1) Solo 8, (2) Solo 9, (3) Varietas Trisula, (4) Bilitar, (5) Kediri, (6) Aceh, (7) Pekanbaru, (8) Solok, (9) Bukittinggi, (10) Varietas Brebes, (11) Varietas Batu Ijo, (12) Varietas Sembrani, (13) Varietas Bangkok, (14) Vietnam 1, (15) Vietnam 2, (16) Cirebon, (17) Bantaeng, (18) Tanduyung, (19) Solo 1, (20) Solo 3, (21) Solo 4 dan (22) Tungganamo.
(Na), jumlah alel efektif (Ne), keragaman genetik (h), indeks informasi Shannon (I) dan persentase lokus polimorfik (PLP). Populasi yang dianalisis struktur genetika populasinya sebanyak 8 populasi, karena hanya 8 populasi yang memenuhi syarat untuk dianalisis dengan jumlah individu hidup paling banyak (N= 5 sampai 8 individu). Populasi lainnya memiliki jumlah individu yang sangat sedikit, sehingga tidak dianalisis struktur genetika populasinya. Struktur genetika populasi yang dihasilkan berdasarkan data ISSR menggunakan program GenAlex dapat dilihat pada Tabel 7.
Nilai keragaman genetik (h) 8 populasi bawang merah yang dikoleksi dari beberapa daerah di Indoneisa secara umum cukup tinggi. Jumlah alel yang diamati berkisar antara 1.43 sampai 1.58, jumlah alel efektif berkisar antara 1.41 sampai 1.53, nilai indeks informasi Shannon berkisar antara 0.37 sampai 0.44 dan nilai keragaman genetik berkisar antara 0.24 sampai 0.31. Persentase lokus polimorfik yang dihasilkan bervariasi berkisar antara 67.83% sampai 77.39% dengan nilai rata-rata persentase lokus polimorfik 72.72%. Populasi Solo memiliki nilai PLP tertinggi (77.39%) sedangkan populasi Solok memiliki nilai PLP terendah (67.83%). Nilai keragaman genetik (h), indeks informasi Shannon dan persentase lokus polimorfik pada populasi bawang merah asal Solo adalah tertinggi dan pada populasi bawang merah asal Solok adalah terendah dibandingkan populasi lainnya. Hal ini menunjukkan bahwa populasi Solok memiliki keragaman genetik terendah dan populasi Solo memiliki keragaman genetik tertinggi di antara populasi bawang merah lainnya.
Tabel 7 Struktur genetika populasi pada 8 populasi bawang merah di Indonesia
No. Populasi N Na Ne I h PLP (%)
bukan gen yang tidak mengekspresikan suatu karakter morfologi tertentu, sehingga kemungkinan untuk sama dengan marka morfologi sangat jauh. Keragaman morfologi ditunjukkan dengan adanya 19 karakter polimorfik pada pengamatan morfologi yang dilakukan. Keragaman morfologi bawang merah juga ditemukan pada 49 aksesi bawang merah asal Ethiopia, dimana terdapat variasi pada karakter tinggi tanaman, panjang daun, diameter umbi, bobot umbi dan umur panen (Degewione et al. 2011). Akter et al. (2015) juga melaporkan 11 genotipe bawang merah asal India memiliki variasi morfologi pada karakter umbi dan daun.
Keragaman dapat terjadi pada tingkat spesies yang disebabkan oleh faktor genetik dan lingkungan (Hartati dan Darsana 2015). Henry (1997) menyatakan bahwa penggunaan marka molekuler dalam analisis keragaman adalah untuk mendukung juga mengevaluasi hasil analisis marka morfologi karena marka molekuler bersifat lebih konsisten atau tidak dipengaruhi oleh kondisi lingkungan. Jumlah pita DNA hasil amplifikasi yang dihasilkan cukup tinggi (115 pita) dimana semakin banyak pita yang dihasilkan oleh masing-masing primer menunjukkan bahwa sekuen pada primer tersebut tersebar melimpah dalam genom tanaman (Powell et al. 1996). Persentase pita polimorfik yang dihasilkan sedikit lebih tinggi dibandingkan hasil penelitian Khar et al. (2011) pada 46 aksesi bawang merah di India menggunakan 30 primer EST-SSR (53.3%) dan Mallor et al. (2014) pada 85 sampel bawang merah di Spanyol menggunakan 18 primer SSR (75%). Primer yang informatif dalam mengungkap keragaman genetik pada 34 genotipe bawang merah yang dianalisis diantaranya primer PKBT 4, ISSRred 4, ISSRred 9 dan ISSRred 10 karena menghasilkan persentase pita polimorfik paling tinggi (90-100%).
Nilai keragaman yang dihasilkan cukup rendah (< 50%) karena genotipe berasal dari spesies yang sama dan dibudidayakan secara vegetatif (umbi). Berdasarkan dendrogram yang dihasilkan marka morfologi koefisien kemiripan morfologi terendah (0.68) atau jarak genetik terjauh (0.32) dijumpai antara bawang merah varietas Tuk-tuk dengan genotipe asal Solok, sedangkan koefisien kemiripan morfologi tertinggi (0.95) atau jarak genetik terdekat (0.05) dijumpai antara genotipe bawang merah asal Solo 7 dengan Vietnam 5 (genotipe Vietnam generasi kedua). Sedangkan marka ISSR menghasilkan dendogram dengan koefisien kemiripan genetik terendah (0.62) atau jarak genetik terjauh (0.38) dijumpai antara bawang merah varietas Batu Ijo dengan genotiope Tanduyung, sedangkan koefisien kemiripan genetik tertinggi (0.89) atau jarak genetik terdekat (0.11) dijumpai antara genotipe bawang merah varietas Trisula dengan Tuk-tuk.
sebagai dasar untuk menentukan genotipe tetua yang akan digunakan untuk proses pemuliaan bawang merah. Berdasarkan analisis PCA marka molekuler, bawang merah varietas Bangkok dan Sembrani yang memiliki diameter umbi terbesar dapat disilangkan dengan bawang merah asal Pekanbaru dan Nganjuk, karena memiliki jarak genetik yang cukup jauh. Persilangan antar genotipe tersebut diharapkan akan menghasilkan genotipe bawang merah yang memiliki sifat unggul dari kedua genotipe tetuanya.
Analisis PCA juga dapat digunakan untuk mengetahui pola pengelompokan dan karakter utama yang menentukan pengelompokan tersebut. Pengelompokkan 34 genotipe bawang merah yang dianalisis baik menggunakan marka morfologi maupun molekuler (ISSR) terlihat tidak berhubungan dengan asal geografi genotipe tersebut, namun berkelompok menjadi satu kelompok yang sama walaupun berasal dari geografi yang berbeda. Hasil penelitian ini sesuai dengan Arifin et al. (2000) yang melaporkan 65 aksesi bawang merah asal Indonesia dan Jepang menggunakan 12 primer RAPD menghasilkan dua kelompok utama yang pengelompokkannya tidak berdasarkan asal geografi aksesi bawang merah tersebut.
Keragaman morfologi dan genetik pada genotipe bawang merah yang diamati diduga merupakan hasil proses budidaya dalam waktu lama dengan kondisi lingkungan beragam. Selain itu diduga dipengaruhi oleh distribusi perdagangan, terlihat dari pola pengelompokan yang tidak behubungan dengan letak geografi atau daerah asal genotipe tersebut. Bawang merah yang dikoleksi dari pulau Sumatera (Aceh, Solok, Bukittinggi dan Pekanbaru) berkelompok dengan bawang merah asal pulau Jawa (Blitar, Solo, Tungganamo, Sumbawa, Kediri, Nganjuk, Yogja dan Brebes) juga pulau Sulawesi (Bantaeng). Hal ini terjadi karena petani bawang merah di pulau Sumatera selain menggunakan bibit lokal juga menggunakan bibit yang berasal dari luar. Petani Aceh selain menggunkan bibit dari lokal Aceh juga menggunkan bibit dari Sumatera Utara untuk memenuhi kebutuhan bibit. Sementara itu, petani di Sumatera Utara bibitnya diperoleh dari Jawa Tengah sehingga terdapat kemiripan antara bawang merah asal Aceh dengan bawang merah asal Jawa Tengah (Solo). Bawang merah asal Bantaeng (Sulawesi Selatan) memiliki kemiripan morfologi dengan bawang merah asal Tungganamo dan Sumbawa (Nusa Tenggara), hal ini terjadi karena bawang merah hasil produksi Nusa Tenggara dipasarkan ke wilayah Sulawesi Selatan (Arsanti 2013; BPS 2015b).
Keragaman genetik suatu individu dalam suatu populasi mewariskan sifat dari tetuanya. Faktor yang menyebabkan keragaman tersebut dapat karena perkawinan secara acak. Faktor lain yang dapat menyebabkan keragaman yaitu mutasi pada lokus berukuran besar dimana efek mutasi sulit dibedakan secara konvensional. Dua alel dikatakan identik hanya jika kedua alel tersebut tidak mengalami mutasi pada pewarisan generasi selanjutnya, namun pada faktanya hasil analisis genetik memiliki frekuensi alel yang beragam (Riupassa 2016). Jika faktor genetik muncul sebagai interaksi gen, maka suatu ciri morfologi yang berbeda dapat muncul pada suatu spesies sebagai karakter kasat mata yang mungkin dapat menjadi potensi plasma nutfah yang terpendam
dalam suatu populasi. Nilai keragaman genetik (h) dan persentase lokus polimorfik (PLP) yang dihasilkan lebih rendah dibandingkan hasil penelitian Baldwin et al. (2012). Baldwin et al. (2012) melaporkan 24 populasi bawang merah di Afrika Barat memiliki nilai h = 0.19-0.49 dan PLP = 20-83% menggunakan marka SSR. Nilai persentase lokus polimorfik (PLP) yang dihasilkan >50% (77.39%) sehingga dapat disimpulkan bahwa primer ISSR yang digunakan pada penelitian ini sangat informatif dan layak digunakan untuk mengungkap keragaman genetik bawang merah di Indonesia. Analisis struktur genetika 8 populasi bawang merah yang diamati menunjukkan bahwa populasi Solo memiliki keragaman genetik tertinggi dan populasi Solok memiliki keragaman genetik terendah (Tabel 7).
Keragaman morfologi dan genetik merupakan modal awal dalam proses pemuliaan tanaman (Surahman et al. 2009). Variasi morfologi yang dihasilkan dapat digunakan sebagai informasi dasar dalam proses perakitan varietas unggul bawang merah Indonesia. Genotipe bawang merah yang dikategorikan unggul diantaranya memiliki diameter umbi besar dan jumlah siung umbi banyak sehingga menghasilkan bobot umbi dan produktivitas panen yang tinggi. Azmi et al. (2011) mengkategorikan bawang merah dengan diameter umbi >1.7 cm termasuk dalam kategori mutu I dan bawang merah dengan diameter umbi <1.3 cm termasuk dalam kategori II. Hampir semua genotipe bawang merah yang diamati pada penelitian ini termasuk dalam kategori mutu I yang disenangi oleh konsumen dan petani kecuali genotipe solo 7 dan varietas Kuning (1.51 cm dan 1.56 cm).
KESIMPULAN
Marka morfologi menghasilkan 19 karakter informatif dengan koefisien kemiripan berkisar antara 0.68-0.95. Marka ISSR menghasilkan 103 pita DNA polimorfik dari 13 primer dengan koefisien kemiripan berkisar antara 0.62-0.89. Tingkat keragaman genetik yang dihasilkan kedua marka yaitu sebesar 27%. Pola pengelompokkan yang dihasilkan kedua marka tidak berhubungan dengan asal geografi genotipe.
Populasi bawang merah Indonesia memiliki keragaman genetik yang cukup berdasarkan nilai struktur genetika populasinya. Populasi bawang merah asal Solo memiliki keragaman genetik tertinggi (I= 0.44, h= 0.31 dan PLP= 77.39%). Sedangkan populasi bawang merah asal Solok memiliki keragaman genetik terendah (I= 0.37, h= 0.24 dan PLP= 67.83%).