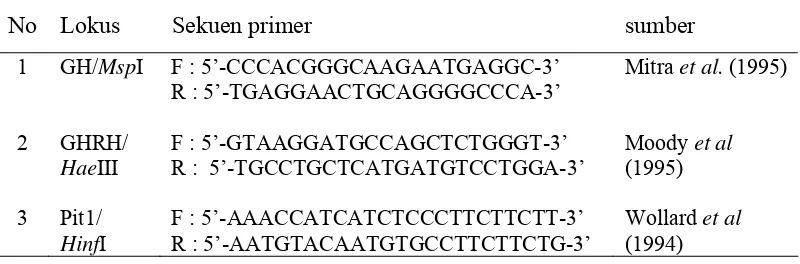KARAKTERISTIK UKURAN TUBUH DAN
POLIMORFISME GEN GH, GHRH DAN PIT-1
PADA POPULASI KERBAU DI BANTEN
ROHMAT DIYONO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Karakteristik Ukuran Tubuh dan
Polimorfisme Gen GH, GHRH dan Pit-1 pada Populasi Kerbau di Banten adalah karya saya sendiri dibawah arahan dan bimbingan para pembimbing. Karya ini
belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber
informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak
diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam
Daftar Pustaka di bagian akhir tesis ini.
Bogor, Agustus 2009
ABSTRACT
ROHMAT DIYONO. Body Measurements Characteristic and Polymorphism of GH, GHRH and Pit-1 Genes of Buffalo Population in Banten. Supervised by
CECE SUMANTRI and ACHMAD FARAJALLAH.
The objectives of this study were to get information about body measurements characteristic and molecular polymorphisms of Banten buffalo population consisted of Pandeglang and Lebak subpopulations. Five variables of body measurement which were body length (BL), wither height (WH), chest circumference (CC), chest depth (CD) and hip height (HH) were analyzed on body measurement study. While, three loci of growth hormone genes (GH/MspI, GHRH/HaeIII and Pit-1/HinfI) were used on molecular polymorphism study. The results showed that body measurements of Pandeglang generally was not differ from Lebak subpopulation according to t-test analysis (P>0.05). Molecular analysis showed that GH/MspI and GHRH/HaeIII loci of Banten population were polymorphic, while Pit-1/HinfI locus was monomorphic. Genetic polymorphisms of GH/MspI locus was low showed by expected heterosigosity value (He=0.0469).
While, genetic polymorphisms of GHRH/HaeIII locus was higher (He=0.4908).
FIS index showed negative value indicated that there was a random mating system
on Banten buffalo population. FIT value for GH/MspI locus near to 0 (-0.0207)
indicated that there was a balanced population according to Hardy-Weinberg principle. A bias of Hardy Weinberg principle was on GHRH/HaeIII locus showed with FIT value near to -1 (-0.7224). Population differentiation indicator,
FST index showed a small value (0.0024) indicated that differentiation of Banten
population to two subpopulations (Pandeglang and Lebak) only decreased a small number of genetic diversity (0.24%). Correlation analysis showed that GHRH/HaeIII genotypes was not significantly correlated with body measurements of Banten buffalo population (P>0.05).
RINGKASAN
ROHMAT DIYONO. Karakteristik Ukuran Tubuh dan Polimorfisme gen GH, GHRH dan Pit-1 pada Populasi Kerbau di Banten. Dibimbing oleh CECE SUMANTRI dan ACHMAD FARAJALLAH.
Penelitian ini bertujuan untuk: 1) mendapatkan informasi mengenai karakteristik kuantitatif ukuran-ukuran tubuh kerbau Banten (Pandeglang dan Lebak) meliputi panjang badan, tinggi pundak, lingkar dada, dalam dada dan tinggi pinggul, 2) mengidentifikasi polimorfisme gen GH, GHRH dan Pit-1 pada lokus GH/MspI, GHRH/HaeIII dan Pit-1/HinfI pada populasi kerbau Banten (Pandeglang dan Lebak), dan 3) menganalisis hubungan antara ukuran-ukuran tubuh kerbau dengan polimorfisme lokus GH/MspI, GHRH/HaeIII dan Pit-1/HinfI.
Penelitian dilakukan di dua Kabupaten di Banten yaitu Pandeglang dan Lebak. Data ukuran tubuh sampel ternak kerbau sebanyak 80 ekor dari Pandeglang dan 121 ekor dari Lebak diperoleh dengan pengukuran langsung dilapangan dari bulan September sampai November 2008. Identifikasi polimorfisme lokus GH/MspI, GHRH/HaeIII dan Pit-1/HinfI dilakukan dengan metode polymerase chain reaction-restriction fragment length polymorphisms (PCR-RFLP) dari bulan Desember 2008 sampai April 2009 di Laboratorium Zoologi Departemen Biologi Fakultas MIPA IPB. Karaktersitik ukuran tubuh kerbau dianalisis secara deskriptif meliputi nilai rataan, simpangan baku dan koefisien keragaman. Sampel darah ternak kerbau yang digunakan dalam penelitian ini berjumlah 77 sampel meliputi 44 sampel dari Pandeglang dan 33 sampel dari Lebak. Analisis polimorfisme meliputi frekuensi alel dan genotipe, heterosigositas pengamatan (Ho) dan heterosigositas harapan (He), keseimbangan
Hardy-Weinberg, dan nilai-nilai indeks fiksasi meliputi FIS, FIT, dan FST.
Selanjutnya dilakukan analisis hubungan antara genotipe dengan ukuran-ukuran tubuh kerbau terkoreksi umur 4 sampai 5 tahun jenis kelamin betina. Analisis hubungan dilakukan dengan menggunakan uji-t.
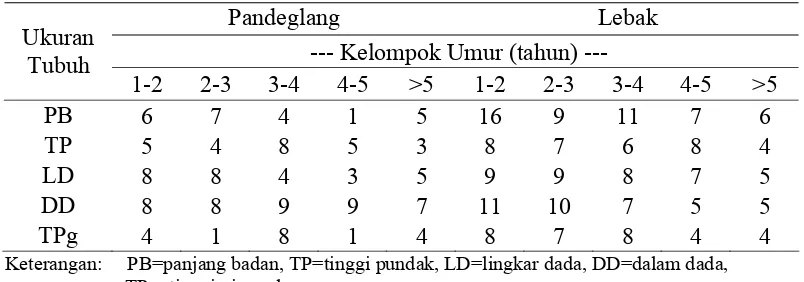
Hasil analisis uji-t menunjukkan bahwa karakteristik kuantitatif ukuran-ukuran tubuh kerbau Pandeglang dan Lebak secara umum tidak berbeda (P>0.05). Lokus GH/MspI dan GHRH/HaeIII bersifat polimorfik, sedangkan lokus Pit-1/HinfI bersifat monomorfik pada populasi kerbau Banten. Alel GH(-) pada lokus GH/MspIhanya ditemukan pada subpopulasi Pandeglang dengan frekuensi 0.04. Frekuensi genotipe GH(+/+) dan GH(+/-) pada subpopulasi Pandeglang adalah 0.92 dan 0.08. Pada subpopulasi Lebak, lokus GH/MspI bersifat monomorfik dengan hanya ditemukanya alel GH(+). Pada subpopulasi Pandeglang, frekuensi alel A dan B lokus GHRH/HaeIII adalah 0.41 dan 0.59, sedangkan pada subpopulasi Lebak adalah 0.43 dan 0.57. Pada subpopulasi Pandeglang, frekuensi genotipe AB dan BB lokus GHRH/HaeIII adalah 0.86 dan 0.14, sedangkan pada subpopulasi Lebak adalah 0.81 dan 0.19. Pada populasi total Banten Lokus GH/MspI mempunyai keragaman yang rendah (He = 0.0469) dan lokus
GHRH/HaeIII mempunyai keragaman yang tinggi (He = 0.4908).
Nilai FIS pada populasi Banten untuk semua lokus bernilai negatif yang
mengindikasikan adanya pola perkawinan acak. Nilai FIT untuk lokus GH/MspI
bahwa distribusi genotipe gen tersebut memenuhi prinsip keseimbangan Hardy-Weinberg. Penyimpangan keseimbangan Hardy-Weinberg terjadi pada lokus GHRH/HaeIII ditunjukkan dengan nilai FIT mendekati -1 (-0.7224). Populasi
kerbau Banten mempuyai nilai FST rataan sebesar 0.0024. Nilai FST yang kecil ini
mengindikasikan bahwa pemisahan populasi Banten menjadi dua subpopulasi Pandeglang dan Lebak hanya akan menurunkan keragaman genetik yang tidak signifikan yaitu sebesar 0.24%. Analisis hubungan genotipe dengan ukuran-ukuran tubuh hanya memungkinkan dilakukan untuk lokus GHRH/HaeIII. Hasil analisis uji-t menunjukkan bahwa keragaman lokus GHRH/HaeIII tidak berhubungan (P>0.05) dengan ukuran-ukuran tubuh kerbau di Banten.
©Hak Cipta milik IPB, tahun 2009
Hak cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusuan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
KARAKTERISTIK UKURAN TUBUH DAN
POLIMORFISME GEN GH,GHRH DAN PIT-1
PADA POPULASI KERBAU DI BANTEN
ROHMAT DIYONO
Tesis
sebagai salah syarat untuk memperoleh gelar Magister Sains pada
Mayor Ilmu Produksi dan Teknologi Peternakan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Karakteristik Ukuran Tubuh dan Polimorfisme Gen GH, GHRH dan Pit-1 pada Populasi Kerbau di Banten
Nama : Rohmat Diyono
NRP : D151070051
Mayor : Ilmu Produksi dan Teknologi Peternakan
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc Ketua
Dr. Ir. Achmad Farajallah, M.Si Anggota
Diketahui
Ketua Mayor Ilmu Produksi dan Teknologi Peternakan
Dr. Ir. Rarah Ratih A.M., DEA
Dekan Sekolah Pascasarjana IPB
Prof. Dr. Ir. Khairil Anwar Notodipuro, M.S.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala
karunia-Nya sehingga penulisan thesis ini dapat terselesaikan. Thesis ini berjudul
“Karakteristik Ukuran Tubuh dan Polimorfisme Gen GH, GHRH dan Pit-1 pada Populasi Kerbau di Banten”. Pemilihan judul penelitian dilatarbelakangi oleh kondisi populasi kerbau di Indonesia yang jumlahnya semakin menurun dari
tahun ketahun. Padahal pada masa mendatang, ternak kerbau mempunyai potensi
dalam pemenuhan kebutuhan daging. Salah satu sifat ekonomis yang penting pada
ternak kerbau adalah sifat pertumbuhan. Sifat pertumbuhan dapat dikarakterisasi
dengan mudah melalui pengukuran tubuh. Sifat pertumbuhan dikontrol oleh
banyak gen beberapa diantaranya tergabung dalam keluarga hormon pertumbuhan
(somatotropin). Harapan kedepan semoga hasil penelitian ini dapat memberikan informasi yang diperlukan sebagai langkah awal untuk melakukan program
pemuliaan kerbau.
Pada kesempatan ini penulis mengucapkan terimakasih kepada Prof.Dr.Ir.
Cece Sumantri, M.Agr.Sc selaku ketua komisi pembimbing, Dr.Ir.Achmad
Farajallah, M.Si selaku pembimbing anggota dan Prof.Dr.Ir.Ronny Rachman
Noor, M.Rur.Sc selaku penguji luar komisi. Secara khusus ucapan terimakasih
penulis sampaikan kepada Prof.Dr.Ir.Cece Sumantri, M.Agr.Sc selaku peneliti
utama yang telah membantu biaya penelitian melalui kerjasama dengan Badan
Penelitian dan Pengembangan Pertanian, Departemen Pertanian RI dalam program
Kerjasama Kemitraan Penelitian Pertanian dengan Perguruan Tinggi (KKP3T)
tahun 2008. Ucapan terimakasih juga penulis sampaikan kepada semua pihak
yang telah membantu baik terlibat secara langsung dan tidak langsung dalam
penelitian dan penulisan tesis ini. Akhirnya, penghargaan dan ucapan terimakasih
sebesar-besarnya disampaikan kepada orang tua penulis, Bapak Brahim dan Ibu
Wagini (Almh) yang telah memberikan doa, dukungan moral dan materil yang
tidak ternilai besarnya.
Bogor, Agustus 2009
RIWAYAT HIDUP
Penulis dilahirkan di Temanggung pada tanggal 11 Juli 1985 dari Bapak
Brahim dan Ibu Wagini (almh). Penulis merupakan anak keempat dari empat
bersaudara.
Pendidikan sarjana di tempuh di Program Studi Teknologi Produksi
Ternak, Fakultas Peternakan Institut Pertanian Bogor (IPB) dan lulus pada tahun
2007. Pada tahun yang sama, penulis terdaftar pada Mayor Ilmu Produksi dan
Teknologi Peternakan Sekolah Pascasarjana IPB dengan biaya sendiri. Minat
keilmuan yang diambil yaitu Genetika dan Pemuliaan Ternak. Selama kuliah
penulis aktif sebagai anggota sekaligus penggerak Kelompok Tani Ternak “Ngudi
Mandiri” di Desa Prangkokan, Kecamatan Bejen, Kabupaten Temanggung Jawa
KARAKTERISTIK UKURAN TUBUH DAN
POLIMORFISME GEN GH, GHRH DAN PIT-1
PADA POPULASI KERBAU DI BANTEN
ROHMAT DIYONO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Karakteristik Ukuran Tubuh dan
Polimorfisme Gen GH, GHRH dan Pit-1 pada Populasi Kerbau di Banten adalah karya saya sendiri dibawah arahan dan bimbingan para pembimbing. Karya ini
belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber
informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak
diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam
Daftar Pustaka di bagian akhir tesis ini.
Bogor, Agustus 2009
ABSTRACT
ROHMAT DIYONO. Body Measurements Characteristic and Polymorphism of GH, GHRH and Pit-1 Genes of Buffalo Population in Banten. Supervised by
CECE SUMANTRI and ACHMAD FARAJALLAH.
The objectives of this study were to get information about body measurements characteristic and molecular polymorphisms of Banten buffalo population consisted of Pandeglang and Lebak subpopulations. Five variables of body measurement which were body length (BL), wither height (WH), chest circumference (CC), chest depth (CD) and hip height (HH) were analyzed on body measurement study. While, three loci of growth hormone genes (GH/MspI, GHRH/HaeIII and Pit-1/HinfI) were used on molecular polymorphism study. The results showed that body measurements of Pandeglang generally was not differ from Lebak subpopulation according to t-test analysis (P>0.05). Molecular analysis showed that GH/MspI and GHRH/HaeIII loci of Banten population were polymorphic, while Pit-1/HinfI locus was monomorphic. Genetic polymorphisms of GH/MspI locus was low showed by expected heterosigosity value (He=0.0469).
While, genetic polymorphisms of GHRH/HaeIII locus was higher (He=0.4908).
FIS index showed negative value indicated that there was a random mating system
on Banten buffalo population. FIT value for GH/MspI locus near to 0 (-0.0207)
indicated that there was a balanced population according to Hardy-Weinberg principle. A bias of Hardy Weinberg principle was on GHRH/HaeIII locus showed with FIT value near to -1 (-0.7224). Population differentiation indicator,
FST index showed a small value (0.0024) indicated that differentiation of Banten
population to two subpopulations (Pandeglang and Lebak) only decreased a small number of genetic diversity (0.24%). Correlation analysis showed that GHRH/HaeIII genotypes was not significantly correlated with body measurements of Banten buffalo population (P>0.05).
RINGKASAN
ROHMAT DIYONO. Karakteristik Ukuran Tubuh dan Polimorfisme gen GH, GHRH dan Pit-1 pada Populasi Kerbau di Banten. Dibimbing oleh CECE SUMANTRI dan ACHMAD FARAJALLAH.
Penelitian ini bertujuan untuk: 1) mendapatkan informasi mengenai karakteristik kuantitatif ukuran-ukuran tubuh kerbau Banten (Pandeglang dan Lebak) meliputi panjang badan, tinggi pundak, lingkar dada, dalam dada dan tinggi pinggul, 2) mengidentifikasi polimorfisme gen GH, GHRH dan Pit-1 pada lokus GH/MspI, GHRH/HaeIII dan Pit-1/HinfI pada populasi kerbau Banten (Pandeglang dan Lebak), dan 3) menganalisis hubungan antara ukuran-ukuran tubuh kerbau dengan polimorfisme lokus GH/MspI, GHRH/HaeIII dan Pit-1/HinfI.
Penelitian dilakukan di dua Kabupaten di Banten yaitu Pandeglang dan Lebak. Data ukuran tubuh sampel ternak kerbau sebanyak 80 ekor dari Pandeglang dan 121 ekor dari Lebak diperoleh dengan pengukuran langsung dilapangan dari bulan September sampai November 2008. Identifikasi polimorfisme lokus GH/MspI, GHRH/HaeIII dan Pit-1/HinfI dilakukan dengan metode polymerase chain reaction-restriction fragment length polymorphisms (PCR-RFLP) dari bulan Desember 2008 sampai April 2009 di Laboratorium Zoologi Departemen Biologi Fakultas MIPA IPB. Karaktersitik ukuran tubuh kerbau dianalisis secara deskriptif meliputi nilai rataan, simpangan baku dan koefisien keragaman. Sampel darah ternak kerbau yang digunakan dalam penelitian ini berjumlah 77 sampel meliputi 44 sampel dari Pandeglang dan 33 sampel dari Lebak. Analisis polimorfisme meliputi frekuensi alel dan genotipe, heterosigositas pengamatan (Ho) dan heterosigositas harapan (He), keseimbangan
Hardy-Weinberg, dan nilai-nilai indeks fiksasi meliputi FIS, FIT, dan FST.
Selanjutnya dilakukan analisis hubungan antara genotipe dengan ukuran-ukuran tubuh kerbau terkoreksi umur 4 sampai 5 tahun jenis kelamin betina. Analisis hubungan dilakukan dengan menggunakan uji-t.
Hasil analisis uji-t menunjukkan bahwa karakteristik kuantitatif ukuran-ukuran tubuh kerbau Pandeglang dan Lebak secara umum tidak berbeda (P>0.05). Lokus GH/MspI dan GHRH/HaeIII bersifat polimorfik, sedangkan lokus Pit-1/HinfI bersifat monomorfik pada populasi kerbau Banten. Alel GH(-) pada lokus GH/MspIhanya ditemukan pada subpopulasi Pandeglang dengan frekuensi 0.04. Frekuensi genotipe GH(+/+) dan GH(+/-) pada subpopulasi Pandeglang adalah 0.92 dan 0.08. Pada subpopulasi Lebak, lokus GH/MspI bersifat monomorfik dengan hanya ditemukanya alel GH(+). Pada subpopulasi Pandeglang, frekuensi alel A dan B lokus GHRH/HaeIII adalah 0.41 dan 0.59, sedangkan pada subpopulasi Lebak adalah 0.43 dan 0.57. Pada subpopulasi Pandeglang, frekuensi genotipe AB dan BB lokus GHRH/HaeIII adalah 0.86 dan 0.14, sedangkan pada subpopulasi Lebak adalah 0.81 dan 0.19. Pada populasi total Banten Lokus GH/MspI mempunyai keragaman yang rendah (He = 0.0469) dan lokus
GHRH/HaeIII mempunyai keragaman yang tinggi (He = 0.4908).
Nilai FIS pada populasi Banten untuk semua lokus bernilai negatif yang
mengindikasikan adanya pola perkawinan acak. Nilai FIT untuk lokus GH/MspI
bahwa distribusi genotipe gen tersebut memenuhi prinsip keseimbangan Hardy-Weinberg. Penyimpangan keseimbangan Hardy-Weinberg terjadi pada lokus GHRH/HaeIII ditunjukkan dengan nilai FIT mendekati -1 (-0.7224). Populasi
kerbau Banten mempuyai nilai FST rataan sebesar 0.0024. Nilai FST yang kecil ini
mengindikasikan bahwa pemisahan populasi Banten menjadi dua subpopulasi Pandeglang dan Lebak hanya akan menurunkan keragaman genetik yang tidak signifikan yaitu sebesar 0.24%. Analisis hubungan genotipe dengan ukuran-ukuran tubuh hanya memungkinkan dilakukan untuk lokus GHRH/HaeIII. Hasil analisis uji-t menunjukkan bahwa keragaman lokus GHRH/HaeIII tidak berhubungan (P>0.05) dengan ukuran-ukuran tubuh kerbau di Banten.
©Hak Cipta milik IPB, tahun 2009
Hak cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusuan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
KARAKTERISTIK UKURAN TUBUH DAN
POLIMORFISME GEN GH,GHRH DAN PIT-1
PADA POPULASI KERBAU DI BANTEN
ROHMAT DIYONO
Tesis
sebagai salah syarat untuk memperoleh gelar Magister Sains pada
Mayor Ilmu Produksi dan Teknologi Peternakan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Karakteristik Ukuran Tubuh dan Polimorfisme Gen GH, GHRH dan Pit-1 pada Populasi Kerbau di Banten
Nama : Rohmat Diyono
NRP : D151070051
Mayor : Ilmu Produksi dan Teknologi Peternakan
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc Ketua
Dr. Ir. Achmad Farajallah, M.Si Anggota
Diketahui
Ketua Mayor Ilmu Produksi dan Teknologi Peternakan
Dr. Ir. Rarah Ratih A.M., DEA
Dekan Sekolah Pascasarjana IPB
Prof. Dr. Ir. Khairil Anwar Notodipuro, M.S.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala
karunia-Nya sehingga penulisan thesis ini dapat terselesaikan. Thesis ini berjudul
“Karakteristik Ukuran Tubuh dan Polimorfisme Gen GH, GHRH dan Pit-1 pada Populasi Kerbau di Banten”. Pemilihan judul penelitian dilatarbelakangi oleh kondisi populasi kerbau di Indonesia yang jumlahnya semakin menurun dari
tahun ketahun. Padahal pada masa mendatang, ternak kerbau mempunyai potensi
dalam pemenuhan kebutuhan daging. Salah satu sifat ekonomis yang penting pada
ternak kerbau adalah sifat pertumbuhan. Sifat pertumbuhan dapat dikarakterisasi
dengan mudah melalui pengukuran tubuh. Sifat pertumbuhan dikontrol oleh
banyak gen beberapa diantaranya tergabung dalam keluarga hormon pertumbuhan
(somatotropin). Harapan kedepan semoga hasil penelitian ini dapat memberikan informasi yang diperlukan sebagai langkah awal untuk melakukan program
pemuliaan kerbau.
Pada kesempatan ini penulis mengucapkan terimakasih kepada Prof.Dr.Ir.
Cece Sumantri, M.Agr.Sc selaku ketua komisi pembimbing, Dr.Ir.Achmad
Farajallah, M.Si selaku pembimbing anggota dan Prof.Dr.Ir.Ronny Rachman
Noor, M.Rur.Sc selaku penguji luar komisi. Secara khusus ucapan terimakasih
penulis sampaikan kepada Prof.Dr.Ir.Cece Sumantri, M.Agr.Sc selaku peneliti
utama yang telah membantu biaya penelitian melalui kerjasama dengan Badan
Penelitian dan Pengembangan Pertanian, Departemen Pertanian RI dalam program
Kerjasama Kemitraan Penelitian Pertanian dengan Perguruan Tinggi (KKP3T)
tahun 2008. Ucapan terimakasih juga penulis sampaikan kepada semua pihak
yang telah membantu baik terlibat secara langsung dan tidak langsung dalam
penelitian dan penulisan tesis ini. Akhirnya, penghargaan dan ucapan terimakasih
sebesar-besarnya disampaikan kepada orang tua penulis, Bapak Brahim dan Ibu
Wagini (Almh) yang telah memberikan doa, dukungan moral dan materil yang
tidak ternilai besarnya.
Bogor, Agustus 2009
RIWAYAT HIDUP
Penulis dilahirkan di Temanggung pada tanggal 11 Juli 1985 dari Bapak
Brahim dan Ibu Wagini (almh). Penulis merupakan anak keempat dari empat
bersaudara.
Pendidikan sarjana di tempuh di Program Studi Teknologi Produksi
Ternak, Fakultas Peternakan Institut Pertanian Bogor (IPB) dan lulus pada tahun
2007. Pada tahun yang sama, penulis terdaftar pada Mayor Ilmu Produksi dan
Teknologi Peternakan Sekolah Pascasarjana IPB dengan biaya sendiri. Minat
keilmuan yang diambil yaitu Genetika dan Pemuliaan Ternak. Selama kuliah
penulis aktif sebagai anggota sekaligus penggerak Kelompok Tani Ternak “Ngudi
Mandiri” di Desa Prangkokan, Kecamatan Bejen, Kabupaten Temanggung Jawa
DAFTAR ISI
Halaman
DAFTAR TABEL ... iii
DAFTAR GAMBAR ... iv
DAFTAR LAMPIRAN ... v
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan ... 2
TINJAUAN PUSTAKA ... 3
Domestikasi dan Tipe Kerbau ... 3
Populasi Kerbau di Indonesia ... 4
Produktivitas dan Karakteristik Ukuran Tubuh Kerbau ... 5
MAS (Marker Assisted Selection) ... 7
Kandidat Marker Sifat Pertumbuhan... 8
Gen Growth Hormone (GH) ... 9
Gen Growth Hormone Releasing Hormone (GHRH) ... 10
Gen Pituitary Transcription Factor-1 (Pit-1) ... 11
Analisis Polimorfisme Gen ... 11
MATERI DAN METODE ... 13
Tempat dan Waktu Penelitian ... 13
Kondisi Umum Propinsi Banten.. ... 13
Pengukuran Karakteristik Ukuran Tubuh ……….. 15
Materi ………. 15
Metode ……… 15
Analisis Data ………... 17
Identifikasi Polimorfisme Gen GH, GHRH dan Pit-1 ... 18
Materi ……….. 18
Metode ……… 19
Analisis Data ………... 21
HASIL DAN PEMBAHASAN ... 24
Koefisien Keragaman ... 24
Ukuran-Ukuran Tubuh Terkait dengan Performa
Produksi ... 25
Pengaruh Lingkungan terhadap Ukuran Tubuh Kerbau ... 28
Identifikasi Polimorfisme Lokus GH/MspI, GHRH/HaeIII
dan Pit-1/HinfI ……… 31
Polimorfisme Lokus GH/MspI, GHRH/HaeIII
dan Pit-1/HinfI ……... 35 Heterosigositas dan Indeks Fiksasi ... 39
Hubungan Genotipe dengan Ukuran Tubuh ... 42
Upaya Pemuliaan Kerbau Banten ... 44
KESIMPULAN DAN SARAN ... 48
Kesimpulan ... 48
Saran ... 48
DAFTAR PUSTAKA ... 49
DAFTAR TABEL
Halaman
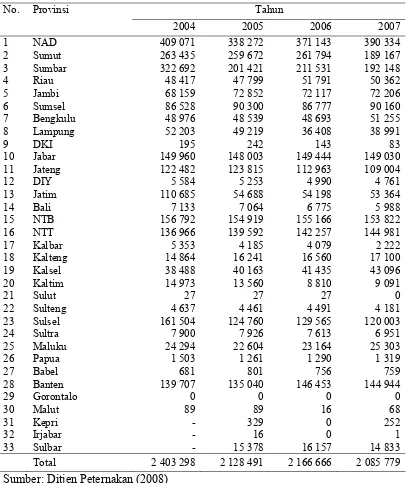
1. Populasi kerbau berdasarkan provinsi tahun 2004 sampai 2007... 5
2 Sampel ternak kerbau menurut umur, jenis kelamin dan asal ternak ….. 15
3 Kriteria penentuan umur kerbau berdasarkan pergantian gigi seri ……. 16
4 Informasi sekuen primer yang digunakan dalam penelitian ……… 18
5 Koefisien keragaman (%) ukuran-ukuran tubuh kerbau jantan
Pandeglang dan Lebak pada berbagai kelompok umur ... 24
6 Koefisien keragaman (%) ukuran-ukuran tubuh kerbau betina
Pandeglang dan Lebak pada berbagai kelompok umur ... 24
7 Frekuensi alel, frekuensi genotipe dan keseimbangan Hardy-Weinberg (χ2 dan p) pada subpopulasi Pandeglang dan Lebak ………... 37
8 Nilai heterosigositas pengamatan (Ho) dan heterosigositas harapan (He) 40
9 Heterosigositas dan indeks fiksasi pada populasi Banten ………... 41
10 Rataan ukuran tubuh dan standar eror (x + s.e) pada setiap genotipe
DAFTAR GAMBAR
Halaman
1 Peta lokasi penelitian di Provinsi Banten ……… 14
2 Skema pengukuran tubuh kerbau... 16
3 Perbandingan rataan panjang badan antara kerbau Pandeglang dan Lebak pada setiap kelompok umur dan jenis kelamin... 25
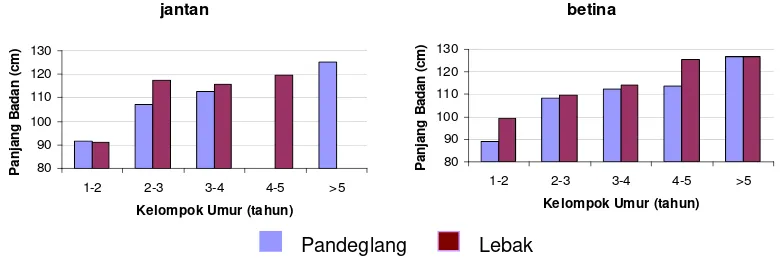
4 Perbandingan rataan tinggi pundak antara kerbau Pandeglang dan Lebak pada setiap kelompok umur dan jenis kelamin... 26
5 Perbandingan rataan lingkar dada antara kerbau Pandeglang dan Lebak
pada setiap kelompok umur dan jenis kelamin... 26
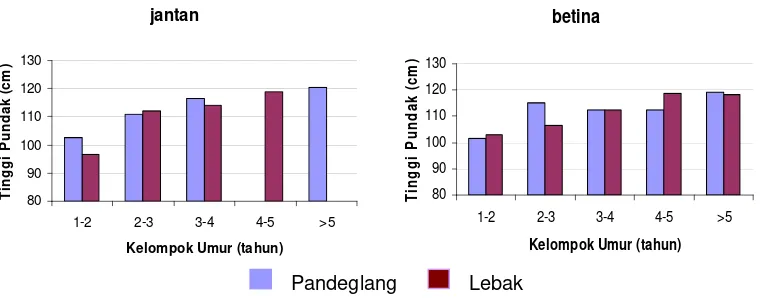
6 Perbandingan rataan dalam dada antara kerbau Pandeglang dan Lebak
pada setiap kelompok umur dan jenis kelamin... 27
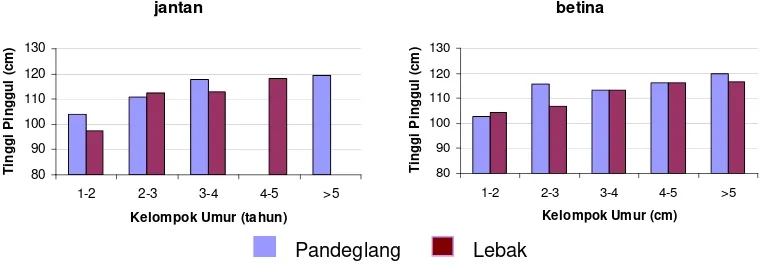
7 Perbandingan rataan tinggi pinggul antara kerbau Pandeglang dan Lebak pada setiap kelompok umur dan jenis kelamin... 28
8 Perkiraan sekuen gen GH target ……….
32 9 Pola pita pemotongan gen GH Sepanjang 327 pb pada gel
poliakrilamid 6%... 32
10 Perkiraan sekuen gen GHRH target ……….. 33
11 Produk PCR gen GHRH dengan panjang sekitar 451 pb, dan pola pita pemotongan dengan enzim restriksi HaeIII pada gel poliakrilamid 6%... 34
12 Perkiraan sekuen gen Pit-1 target ……….. 35
DAFTAR LAMPIRAN
Halaman
1. Rataan, standar eror (SE) dan koefisien keragaman (KK) ukuran-ukuran tubuh kerbau Pandeglang dan Lebak kelompok umur 1 sampai 2 tahun... 55
2 Rataan, standar eror (SE) dan koefisien keragaman (KK) ukuran-ukuran tubuh Kerbau Pandeglang dan Lebak kelompok umur 2 sampai 3 tahun .. 55
3 Rataan, standar eror (SE) dan koefisien keragaman (KK) ukuran-ukuran tubuh Kerbau Pandeglang dan Lebak kelompok umur 3 sampai 4 tahun .. 56
4 Rataan, standar eror (SE) dan koefisien keragaman (KK) ukuran-ukuran tubuh Kerbau Pandeglang dan Lebak kelompok umur 4 sampai 5 tahun... 56
5 Rataan, standar eror (SE) dan koefisien keragaman (KK) ukuran-ukuran tubuh Kerbau Pandeglang dan Lebak kelompok umur lebih 5 tahun …..
57
6 Perbandingan rataan dan standar eror ukuran-ukuran tubuh kerbau Pandeglang dan Lebak jenis kelamin jantan kelompok umur 1 sampai 2 tahun ... 57
7 Perbandingan rataan dan standar eror ukuran-ukuran tubuh kerbau Pandeglang dan Lebak jenis kelamin betina kelompok umur 1 sampai 2 tahun ... 58
8 Perbandingan rataan dan standar eror ukuran-ukuran tubuh kerbau Pandeglang dan Lebak jenis kelamin jantan kelompok umur 2 sampai 3 tahun ... 58
9 Perbandingan rataan dan standar eror ukuran-ukuran tubuh kerbau Pandeglang dan Lebak jenis kelamin betina kelompok umur 2 sampai 3 tahun ... 59
10 Perbandingan rataan dan standar eror ukuran-ukuran tubuh kerbau Pandeglang dan Lebak jenis kelamin jantan kelompok umur 3 sampai 4 tahun ... 59
11 Perbandingan rataan dan standar eror ukuran-ukuran tubuh kerbau Pandeglang dan Lebak jenis kelamin betina kelompok umur 3 sampai 4 tahun ... 60
12 Perbandingan rataan dan standar eror ukuran-ukuran tubuh kerbau Pandeglang dan Lebak jenis kelamin betina kelompok umur 4 sampai 5 tahun ... 60
13 Perbandingan rataan dan standar eror ukuran-ukuran tubuh kerbau Pandeglang dan Lebak jenis kelamin betina kelompok umur lebih dari 5 tahun ... 61
PENDAHULUAN
Latar Belakang
Kerbau (Bubalus bubalis) merupakan ternak ruminansia yang digunakan sebagai salah satu sumber mata pencaharian bagi sebagian masyarakat petani di
pedesaan dan berpotensi sebagai salah satu sumber dalam pemenuhan kebutuhan
daging nasional. Kerbau mampu memanfaatkan pakan bermutu rendah dan
beradaptasi pada lingkungan yang cukup keras, spesifik lokasi dan sudah menyatu
dengan kehidupan sosial budaya masyarakat. Populasi kerbau di Indonesia
cenderung mengalami penurunan dari tahun ketahun. Pada tahun 2004 populasi
kerbau mencapai 2 403 298 ekor dan pada tahun 2007 menurun 13.21% menjadi 2
085 779 ekor (Ditjen Peternakan 2008). Penurunan populasi ini hampir terjadi di
semua wilayah nusantara yang merupakan basis pengembangan populasi kerbau,
termasuk di Provinsi Banten. Penurunan populasi yang tidak terkontrol
dikhawatirkan akan mengakibatkan hilangnya gen-gen penting yang terkait
dengan daya hidup dan kemampuan reproduksi serta gen-gen yang mengontrol
sifat ekonomis. Penurunan populasi kerbau disebabkan oleh semakin sempitnya
lahan penggembalaan, penggunaan mesin untuk mengolah lahan pertanian dan
penurunan preferensi peternak terhadap kerbau. Penurunan preferensi peternak
terhadap kerbau disebabkan oleh rendahnya produktifitas kerbau dibandingkan
ternak lain seperti sapi dan domba apabila dipelihara secara intensif. Upaya-upaya
untuk untuk meningkatkan produktivitas kerbau oleh karenanya perlu dilakukan
baik melalui perbaikan manajemen pemeliharaan maupun peningkatan mutu
genetik ternak.
Upaya peningkatan mutu genetik ternak dapat dilakukan melalui seleksi
terhadap sifat yang mempunyai nilai ekonomis tinggi. Salah satu sifat ekonomis
yang bernilai ekonomi tinggi yaitu sifat pertumbuhan. Upaya seleksi terhadap sifat
pertumbuhan dapat dilakukan dengan mudah berdasarkan ukuran-ukuran tubuh
kerbau. Ukuran-ukuran tubuh mempunyai kegunaan yang dapat dimanfaatkan
untuk menduga bobot badan dan menentukan harga jual ternak. Namun demikian
upaya seleksi berdasarkan ukuran tubuh akan menghasilkan respon seleksi yang
lambat karena variasi lingkungan yang tinggi. Terobosan baru berupa penerapan
untuk mendapatkan respon seleksi yang lebih cepat. Metode seleksi ini dapat
digunakan untuk mendeteksi sifat unggul dari seekor ternak dalam waktu yang
relatif lebih cepat. Kemajuan dalam bidang biologi molekuler berupa penerapan
teknik polymerase chain reaction (PCR) dan restriction fragment length polymorphism (RFLP), memungkinkan upaya seleksi dapat dilakukan dengan bantuan marker molekuler yang telah terbukti mengontrol sifat ekonomis. Upaya
tersebut dapat dilakukan melalui dua tahap yaitu identifikasi polimorfisme gen
yang mengontrol sifat ekonomis dan pencarian tipe gen (alel) yang terkait dengan
sifat unggul. Kandidat gen yang dapat digunakan yaitu gen yang tergabung dalam
keluarga hormon pertumbuhan diantaranya gen growth hormone (GH), growth hormone releasing hormone (GHRH), dan pituitary transcription factor (Pit-1). Gen-gen ini merupakan pengontrol sifat pertumbuhan yang keberadaan dan
polimorfismenya penting untuk mendukung seleksi terhadap sifat pertumbuhan.
Pada penelitian sebelumnya telah diketahui bahwa terdapat polimorfisme pada
ketiga ketiga gen tersebut khususnya pada sapi yaitu pada pada lokus GH/MspI (Zhang et al. 1992), GHRH/HaeIII (Moody et al. 1995) dan Pit-1/HinfI (Wollard et al. 1994).
Tujuan
Penelitian ini bertujuan untuk:
1. Mendapatkan informasi mengenai karakteristik kuantitatif ukuran-ukuran
tubuh kerbau Banten (Pandeglang dan Lebak) meliputi panjang badan, tinggi
pundak, lingkar dada, dalam dada dan tinggi pinggul,
2. Mengidentifikasi polimorfisme gen GH, GHRH dan Pit-1 pada lokus
GH/MspI, GHRH/HaeIII dan Pit-1/HinfI pada populasi kerbau Banten (Pandeglang dan Lebak),
3. Menganalisis hubungan antara ukuran-ukuran tubuh kerbau dengan
TINJAUAN PUSTAKA
Domestikasi dan Tipe Kerbau
Kerbau termasuk dalam sub-famili Bovinae, genus Bubalus. Menurut sejarah perkembangan domestikasi, ternak kerbau yang berkembang di seluruh
dunia berasal dari daerah sekitar India. Kerbau Asia dan Eropa termasuk dalam
genus Bubalis sedangkan Kerbau Afrika termasuk dalam genus Syncerus. Terdapat beberapa jenis kerbau liar yang masih dapat dijumpai, yaitu Anoa,
Kerbau Mindoro, Bubalus caffer dan Kerbau Merah. Kerbau Mindoro (Buballus mindorensis) terdapat di Filipina. Kerbau ini juga bertubuh kecil, menyerupai kerbau kerdil. Buballus caffer merupakan kerbau liar yang sangat kuat terdapat di Afrika Timur, dan beberapa di daerah Afrika Barat Daya, Transvaal dan Kongo.
Sesuai dengan namanya, kerbau merah berwarna merah dengan ukuran tubuh
kecil, tingginya 1.2 sampai 1.5 m dan terdapat di Afrika Barat, di daerah Tsad,
Niger hilir, Kongo dan Maroko Selatan. Kerbau Afrika, atau disebut juga dengan
kerbau liar, terdiri atas dua subspesies, yaitu Syncerus caffer caffer dan Syncerus caffer nanus. Kerbau Afrika belum pernah mengalami proses domestikasi dan masih berupa kerbau liar atau semi liar (Kikkawa et al. 1997).
Kerbau domestik (Bubalus bubalus) terdiri atas dua tipe yaitu tipe sungai (river buffalo) dan tipe rawa (swamp buffalo). Kedua tipe kerbau ini dibedakan berdasarkan pada penampakan, tingkah laku, pemanfaatan dan habitatnya. Kerbau
sungai mempunyai bentuk tanduk melingkar ke bawah dan kerbau rawa
mempunyai tanduk melengkung ke belakang. Kerbau sungai menunjukkan
kesenangan terhadap air mengalir yang bersih, sedangkan kerbau rawa suka
berkubang dalam lumpur, rawa-rawa dan air menggenang. Kerbau tipe rawa biasa
digunakan sebagai ternak kerja, untuk nantinya dipotong sebagi penghasil daging
dan tidak pernah sebagai penghasil susu, sedangkan kerbau sungai merupakan tipe
penghasil susu (Kikkawa et al. 1997). Kerbau rawa mempunyai 24 pasang kromosom (48 kromosom), sedangkan kerbau sungai 25 pasang (50 kromosom).
Menurut Chunchai (1981) selain adanya perbedaan dalam hal jumlah pasangan
grup kerbau tersebut. Pada kerbau lumpur, besar kromosom Y tidak melebihi 1/3
dari besar kromosom X, sedangkan pada kerbau sungai besar kromosom Y
mencapai sekitar ½ dari kromosom X.
Jenis kerbau yang ada di Indonesia dan beberapa negara Asia Tenggara
lainnya adalah jenis kerbau rawa (swamp buffalo) yang biasanya digunakan sebagai ternak kerja dan penghasil daging. Bobot dewasa kerbau rawa rata-rata
450 sampai 650 kg untuk kerbau jantan dan 350 sampai 450 kg untuk kerbau
betina Umur dewasa berkisar antara 4 sampai 5 tahun (Chantalakhana &
Skunmum 2002).
Populasi Kerbau di Indonesia
Populasi ternak kerbau di Indonesia hanya sekitar 2% dari populasi dunia.
Populasi kerbau di Indonesia sebagian besar merupakan kerbau lumpur dan hanya
sedikit kerbau sungai di Sumatera Utara yaitu kerbau Murrah yang dipelihara oleh
masyarakat keturunan India dan digunakan sebagai penghasil susu. Populasi
kerbau di Indonesia dari tahun 2004 sampai 2007 disajikan pada Tabel 1.
Kerbau rawa yang berkembang dan dibentuk menurut agroekosistem
memunculkan berbagai tipe kerbau. Di Toraja ada kerbau Tedong Bonga, di
daerah Alabio ada kerbau Rawa, di Tapanuli Selatan ada kerbau Binanga, di
Kalimantan Selatan ada kerbau Kalang dan di Maluku ada kerbau Moa.
Disamping itu di daerah Taman Nasional Baluran didapatkan pula kerbau liar.
Potensi pengembangan ternak kerbau di Indonesia cukup besar, mengingat
populasi pada tahun 2007 cukup banyak sebesar 2 085 799 ekor dengan
penyebaran yang tidak merata antar daerah. Populasi kerbau terbanyak ada di
Aceh, Sumatera Utara, Sumatera Barat, Sumatera Utara, Nusa Tenggara Barat,
Jawa Barat, Nusa Tenggara Timur, Banten, Sulawesi Selatan dan Jawa Tengah,
dengan jumalh mencapai 76.4% dari total populasi nasional (Ditjen Peternakan
Tabel 1 Populasi kerbau berdasarkan provinsi tahun 2004 sampai 2007
Tahun No. Provinsi
2004 2005 2006 2007
1 NAD 409 071 338 272 371 143 390 334
2 Sumut 263 435 259 672 261 794 189 167
3 Sumbar 322 692 201 421 211 531 192 148
4 Riau 48 417 47 799 51 791 50 362
5 Jambi 68 159 72 852 72 117 72 206
6 Sumsel 86 528 90 300 86 777 90 160
7 Bengkulu 48 976 48 539 48 693 51 255
8 Lampung 52 203 49 219 36 408 38 991
9 DKI 195 242 143 83
10 Jabar 149 960 148 003 149 444 149 030
11 Jateng 122 482 123 815 112 963 109 004
12 DIY 5 584 5 253 4 990 4 761
13 Jatim 110 685 54 688 54 198 53 364
14 Bali 7 133 7 064 6 775 5 988
15 NTB 156 792 154 919 155 166 153 822
16 NTT 136 966 139 592 142 257 144 981
17 Kalbar 5 353 4 185 4 079 2 222
18 Kalteng 14 864 16 241 16 560 17 100
19 Kalsel 38 488 40 163 41 435 43 096
20 Kaltim 14 973 13 560 8 810 9 091
21 Sulut 27 27 27 0
22 Sulteng 4 637 4 461 4 491 4 181
23 Sulsel 161 504 124 760 129 565 120 003
24 Sultra 7 900 7 926 7 613 6 951
25 Maluku 24 294 22 604 23 164 25 303
26 Papua 1 503 1 261 1 290 1 319
27 Babel 681 801 756 759
28 Banten 139 707 135 040 146 453 144 944
29 Gorontalo 0 0 0 0
30 Malut 89 89 16 68
31 Kepri - 329 0 252
32 Irjabar - 16 0 1
33 Sulbar - 15 378 16 157 14 833
Total 2 403 298 2 128 491 2 166 666 2 085 779
Sumber: Ditjen Peternakan (2008)
Produktivitas dan Karakteristik Ukuran Tubuh Kerbau
Ternak kerbau memiliki kemampuan dalam hal memanfaatkan pakan yang
kurang berkualitas (hijauan berprotein rendah dan serat kasar tinggi). Hal ini
dimungkinkan karena karakteristik fisiologi pencernaan dan kapasitas perut ternak
kerbau yang relatif besar. Ternak kerbau memiliki potensi yang relatif mudah dari
lingkungan yang bervariasi (Suhubdy 2007). Ternak kerbau tahan terhadap
tekanan dan perubahan lingkungan yang sangat ekstrim misalnya perubahan
temperatur (head load) atau perubahan fenologi padang rumput. Karakteristik kerbau terhadap lingkungan menunjukkan bahwa sifat produksi dan reproduksi
kerbau sangat responsif apabila habitat dan manajemen pemeliharaanya diperbaiki
(Suhubdy 2007). Salah satu sifat produksi kerbau yang mempunyai nilai ekonomis
penting adalah sifat pertumbuhan yang dapat dikarakterisasikan dengan ukuran
tubuh.
Ukuran-ukuran tubuh ternak dapat digunakan sebagai penduga yang
menyeluruh dari bentuk tubuh dan deskripsi khas dari berbagai gambaran tubuh
(Wiley 1981). Selanjutnya Warwick et al. (1983), bahwa ukuran-ukuran tubuh sangat berguna untuk menentukan asal-usul dan hubungan filogenetik antara
spesies, bangsa dan tipe ternak yang berbeda. Karakter atau sifat kuantitatif
dikontrol oleh banyak gen yang aksinya bersifat aditif. Biasanya hubungan antar
alel yang paling umum adalah kodominan atau dominan tidak penuh (Noor 2004).
Ukuran-ukuran tubuh seperti panjang badan, tinggi pundak dan lingkar dada
merupakan sifat kuantitatif. Karakter ini diperoleh dengan cara pengukuran.
Martojo (1983) menyatakan bahwa ukuran tubuh secara kuantitatif
menimbulkan perbedaan-perbedaan individu dalam populasi dan sering digunakan
dalam seleksi dan sebagai petunjuk morfogenetik dari jenis ternak tertentu.
Ukuran-ukuran tubuh kerbau bermanfaat untuk menduga sifat produksi misalnya
bobot badan dan juga dapat digunakan sebagai parameter untuk kegiatan seleksi
terhadap sifat pertumbuhan. Sifat pertumbuhan yang mempunyai nilai ekonomis
tinggi pada ternak termasuk ternak kerbau salah satunya adalah bobot badan.
Pengukuran bobot badan masih sangat sulit dilakukan secara langsung untuk
ternak besar seperti kerbau di Indonesia. Oleh karena itu, ukuran-ukuran tubuh
dapat digunakan untuk menduga bobot badan kerbau. Bhattacharya (1993)
menyatakan bahwa ukuran lingkar dada, tinggi badan dan panjang badan dapat
digunakan untuk menduga bobot badan ternak kerbau dengan tepat. Selain untuk
tujuan seleksi, informasi mengenai ukuran tubuh dapat digunakan sebagai alat
tubuh untuk pendugaan bobot badan akan sangat bermanfaat untuk sistem usaha
pengembangan ternak kerbau. Bila bobot badan kerbau telah diketahui maka
pemberian pakan untuk kerbau dapat diberikan secara tepat sesuai kebutuhan.
Bobot badan kerbau juga dapat digunakan sebagai standar dalam menentukan
harga jual kerbau.
Berdasarkan bobot badan, kerbau rawa memiliki karakteristik yang
spesifik tergantung lokasi tempat berkembangnya. Kerbau rawa di Thailand
mempunyai bobot badan dewasa 350 sampai 650 kg (Chantalakhana & Skunmum
2002), bobot badan kerbau rawa di Cina sekitar 250 kg, di Mianmar sekitar 300
kg dan di Laos sekitar 500 sampai 600 kg (Shackleton & Harestad 2003). Bobot
badan kerbau dapat diduga melalui ukuran-ukuran tubuh. Ukuran-ukuran tubuh
kerbau domestik yang ada sekarang mempunyai ukuran-ukuran tubuh yang lebih
kecil dibandingkan pada awal domestikasinya. Kerbau liar dalam proses
domestikasinya hanya digunakan sebagai hewan pekerja, hewan buruan serta
dikebiri dan disembelih, tetapi tidak digunakan untuk tujuan pembibitan, sehingga
berakibat menurunya ukuran-ukuran tubuh kerbau diberbagai negara seperti di
Indonesia dan Thailand. Hal ini disebabkan oleh hilangnya gen-gen tertentu yang
mempengaruhi sifat pertumbuhan (Shackleton & Harestad 2003). Oleh karena itu
perlu adanya upaya seleksi untuk meningkatkan produktivitas kerbau khususnya
sifat pertumbuhan. Upaya seleksi tersebut dapat dilakukan dengan mudah
berdasarkan fenotipe ukuran tubuh. Selain itu, seleksi dapat juga dilakukan
dengan bantuan marker molekuler yang berhubungan dengan sifat kuantitatif
ukuran tubuh. Upaya seleksi dengan bantuan marker molekuler disebut dengan
marker assisted selection.
MAS (Marker Assisted Selection)
Metode seleksi sederhana dengan menggunakan informasi fenotipik telah
berhasil dilakukan untuk perbaikan produktivitas ternak. Namun demikian
terdapat beberapa keterbatasan seperti perbedaan jenis kelamin dan sifat-sifat
yang sulit atau mahal untuk diukur dan diamati (Vischer et al. 2000). Munculnya gagasan penggunaan MAS adalah terdapat gen yang memiliki hubungan nyata
MAS adalah suatu harapan yang optimis, tetapi penerapan MAS akan lebih tepat
dilakukan pada skala industri pemuliaan ternak atau industri peternakan sehingga
keberhasilan penerapannya memerlukan strategi terpadu dan menyeluruh untuk
skala usaha peternakan besar (Dekkers 2004). Penerapan MAS dapat dilakukan
atau penting untuk kegiatan seleksi pada sifat yang mempunyai ketepatan seleksi
dan heritabilitas rendah. Ketepatan seleksi diharapkan dapat lebih baik sejak
periode anak bahkan pada fase embrional sekalipun (Meuwissen 2004).
Penerapan MAS memerlukan sebuah marker atau penanda molekuler yang
spesifik. Salah satu teknik yang telah dikembangkan dan secara luas digunakan
untuk mencari marker molekuler tersebut adalah restriction fragment length polymorphisms (RFLP). Penggunaan teknik RFLP menjadi semakin intensif setelah dikembangkannya teknik amplifikasi fragmen DNA yaitu polymerase chain reaction (PCR). Kombinasi teknik PCR dan RFLP merupakan teknik pertama yang dikembangkan untuk memvisualisasikan perbedaan level DNA
yang didasarkan pada penggunaan enzim pemotong (restriction enzymes) yang dapat memotong pada sekuen nukleotida spesifik (Montaldo & Herera 1998).
Jumlah dan ukuran fragmen DNA hasil pemotongan dengan enzim restriksi,
memiliki pola pita ada atau tidak ada tempat restriksi. Apabila tidak terpotong ada
indikasi terjadi mutasi pada situs tersebut sehingga tidak ada variasi hasil
pemotongan dan ekspresinya bersifat kodominan (Meghen et al. 1995). Hasil fragmen potongan DNA tersebut dapat divisualisasikan melalui teknik
elektroforesis yang hasilnya menunjukkan ada atau tidaknya polimorfisme pada
suatu individu dalam populasi (Nei & Kumar 2000).
Kandidat Marker Sifat Pertumbuhan
Prinsip dasar penerapan MAS adalah adanya hubungan yang kuat secara
statistik antara tipe gen (alel) tertentu dengan sifat ekonomis unggul. Kebanyakan
sifat-sifat ekonomis penting merupakan sifat kuantitatif yang dikontrol oleh
banyak gen dan masing-masing gen memberikan sedikit kontribusi pada sifat
tersebut (Noor 2004). Namun demikian, diprediksi terdapat beberapa gen utama
yang memberikan kontribusi lebih pada variasi suatu sifat. Gen semacam ini
atau quantitative traits loci (QTL). Gen mayor yang dapat digunakan sebagai kandidat dalam program MAS yaitu apabila gen tersebut mempunyai fungsi dan
pengaruh biologis yang nyata terhadap sifat kuantitatif. Salah satu sifat kuantitatif
yang bernilai ekonomis pada ternak kerbau adalah sifat pertumbuhan. Sifat
pertumbuhan merupakan sifat yang masih menjadi prioritas sebagai tujuan seleksi
untuk ternak-ternak di Indonesia, khususnya ruminansia. Bila ditinjau dari sisi
hormonal, hormon yang tergabung dalam keluarga hormon pertumbuhan
(somatotropic axis) mempunyai peran yang sangat penting selama beragam fase pertumbuhan ruminasia (Owens et al. 1993). Beberapa gen yang berperan dalam proses pertumbuhan diantaranya gen growth hormone (GH), growth hormone releasing hormone (GHRH) dan pituitary transcription factor-1(Pit-1).
Gen Growth Hormone (GH)
Hormon pertumbuhan sapi (bGH) merupakan sebuah polipetida yang
terdiri dari 191 asam amino (Etherton & Bauman 1998). Gen GH ditranskripsikan
dan diterjemahkan di kelenjar pituitari anterior. Proses transkripsi diregulasi oleh
dua faktor pengikat, yaitu Pit-1 dan Prop-1. Sekresi GH oleh pituitari di stimulus
oleh GH-releasing hormone yang terdapat di hipotalamus, tetapi juga dihambat oleh somatostatin yang juga disekresikan oleh hipotalamus (Giustina & Veldhuis
1998; Pfaffle et al. 1999; Fodor et al. 2006). Sintesis dan sekresi gen GH diregulasi oleh hypothalamic releasing factors dan somatotrophic transcription factors (Fodor et al. 2006).
Seleksi genetik untuk meningkatkan pertumbuhan dan kualitas karkas
telah dihubungkan dengan peningkatan sekresi GH oleh pituitari (Bunger & Hill
1999). Penyuntikan rekombinan hormon GH rekombinan juga mampu
meningkatkan laktasi dan kualitas karkas (Etherton & Bauman 1998). Baik secara
langsung ataupun tidak, gen GH melalui peran gen insulin growth factor-1 (IGF-1) merupakan regulator utama dalam pertumbuhan sel somatic posnatal, stimulasi
proses anabolis seperti pembelahan sel, pertumbuhan tulang dan sintesis protein.
Gen GH telah digunakan sebagai kandidat gen dalam mencari keterkaitan antara
atas fungsinya dalam proses laktasi, metabolisme karbohidrat (Ohlsson et al.1998).
Pada ternak sapi, gen GH terletak pada kromosom 19, Lagziel et al. (2000) mengungkapkan bahwa polimorfisme skuen pada intron 4 dapat
digunakan untuk membedakan ada atau tidaknya punuk pada sapi, yaitu Bos indicus (berpunuk) dan Bos taurus (tidak berpunuk). Perbandingan lain bangsa sapi ini, sebagai contoh adalah sapi Brahman dan Angus mempunyai perbedaan
mencolok pada sifat karkasnya (Morrison 2005). Sapi Brahman memiliki kadar
lemak dan konsentrasi serum yang lebih rendah dibandingkan sapi Angus pada
karkasnya (Lopez et al. 2006). Beberapa penelitian tentang penggunaan polimorfisme gen yang dihubungkan dengan sifat fenotipe menunjukkan adanya
pengaruh yang bersifat aditif. Namun demikian pada populasi silangan Bos indicus dan Bos taurus, genotipe yang heterosigot cenderung menunjukkan performa yang lebih superior dalam pengukuran fisiologis dan sifat produksi
(Pereira et al. 2005). Mutasi basa C-G ditemukan pada kodon ke-127 gen GH sapi (GH/AluI) yang berakibat pada perubahan asam amino leusin (alel L) menjadi valine (alel V) (Lucy et al. 1991). Perubahan asam amino ini telah dihubungkan dengan beberapa sifat produksi pada ternak sapi, seperti pertumbuhan, komposisi
dan kualitas karkas (Regitano et al. 1999).
Gen Growth Hormone Releasing Hormone (GHRH)
Gen GHRH mengkodekan sebuah hormon dengan panjang sekitar 40-44
asam amino. GHRH terekspresikan pada kelenjar hipotalamus, saluran
gastrointestinal, gonad, jaringan kekebalan, dan Plasenta. Fungsi utama GHRH
adalah menstimulasi sekresi gen GH. Selain itu, GHRH meningkatkan konsentrasi
serum GH (Lovendal et al. 1991) dan meningkatkan produksi susu (Lapierre et al. 1988). Vanderkool et al. (1995) melaporkan bahwa GHRH menstimulasi sintesis susu melalui mekanisme yang sama dengan GH.
Pada ternak sapi, gen GHRH terdiri dari 5 ekson (Frohman & Kineman
1999), terletak pada kromosom 13 dan terpaut dengan mikrosatelit CSSM30
HaeIII. Hasilnya menunjukkan bahwa frekuensi genotipe AA rendah (0.08) dan mampu meningkatkan persentase lemak karkas. Cheong et al. (2006) juga melaporkan keterkaitan antara polimirfisme gen GHRH dengan kualitas karkas
pada sapi Korea. Hubungan polimorfisme gen GHRH/HaeIII pada produksi susu telah diteliti oleh Marek et al. (2007). Penelitian yang dilakukan pada populasi sapi perah di Polandia ini menghasilkan tiga genotipe yaitu AA, AB dan BB
dengan frekuensi masing-masing 0.10; 0.37 dan 0.53, dan frekuensi alel A dan B
masing-masing 0.28 dan 0.72. Sapi dengan genotipe AA mempunyai produksi
susu lebih tinggi dibandingkan dengan kedua genotipe lainya. Pada Kerbau,
Rajamurugan et al. (2007) melaporkan bahwa gen GHRH lokus HaeIII pada 60 ekor kerbau perah Murrah adalah monomorfik.
Gen Pituitary Transcription Factor-1(Pit-1)
Gen Pit-1 yang juga dikenal dengan nama growth hormone factor 1 (GHF1) merupakan faktor transkripsi spesifik pituitari yang berperan untuk
perkembangan pituitari dan ekspresi hormon pada mamalia (Cohen et al. 1997). Pit-1 merupakan anggota domain POU yang mengandung protein, yaitu kelompok
regulator transkripsi yang mempunyai peran kunci dalam diferensiasi dan
pembelahan sel (Mangalam et al. 1989). Secara in vivo, kebanyakan dari protein POU berperan penting dalam proses perkembangan yang terkait dengan system
saraf. Pada hewan ternak, gen Pit-1 berfungsi dalam pengaturan transkripsi gen
GH dan prolaktin (Bodner et al. 1988). Gen Pit-1 juga berfungsi sebagai aktivator gen-gen pituitary, termasuk gen Pit-1 itu sendiri (Chen et al. 1990) dan gen GHRH (Lin et al. 1992).
Analisis Polimorfisme Gen
Pemilihan gen yang akan digunakan sebagai kandidat marker untuk
kegiatan seleksi, selain mempunyai hubungan biologis dan fisiologis dengan sifat
kuantitatif juga disyaratkan adanya informasi polimorfisme pada gen kandidat
tersebut. Teknik polymerase chain reaction (PCR) yang dikombinasikan dengan teknik restriction fragment length polymorphisms (RFLP) dapat digunakan untuk mendapatkan informasi ada atau tidaknya polimorfisme pada setiap daerah atau
Ada atau tidaknya polimorfime pada gen atau lokus yang diamati dapat
diketahui dari nilai frekuensi alel. Gen dikatakan bersifat polimorfik yaitu apabila
salah satu alelnya mempunyai frekuensi kurang dari 99% (Nei & Kumar 2000)
atau 95% (Hartl 1988). Sebaliknya, gen dikatakan monomorfik apabila tidak
memenuhi kriteria polimorfik diatas. Keragaman genetik digunakan untuk
menginvestigasi hubungan genetik suatu spesies antar subpopulasi. Prinsipnya
adalah kemungkinan adanya alel bersama yang dimiliki antar subpopulasi yang
disebabkan oleh migrasi. Alel bersama ini juga mengindikasikan adanya asal-usul
atau tetua yang sama (Hartl 1988). Keragaman genetik dapat dihitung secara
kuantitatif dengan menggunakan nilai frekuensi alel. Frekuensi alel adalah
proporsi jumlah suatu alel terhadap jumlah total alel dalam suatu populasi pada
lokus yang sama (Nei & Kumar 2000). Berdasarkan nilai frekuensi alel, maka
selanjutnya dapat dibandingkan perbedaan antar gen, baik didalam maupun antar
populasi. Perhitungan frekuensi alel dapat dilakukan dengan mudah menurut
petunjuk Nei (1987).
Berdasarkan nilai frekuensi alel dan frekuensi genotipe dapat diduga nilai
keragaman genetik yaitu frekuensi heterosigositas pengamatan (Ho) dan
heterosigositas harapan (He) (Nei 1987). Pola perkawinan dan seleksi serta
struktur populasi yang berlangsung pada populasi yang diamati dapat diduga
dengan menggunakan nilai-nilai indeks fiksasi yang dikenal dengan uji-F statistic
yang terdiri dari tiga parameter yaitu FIS, FIT dan FST. Nilai Indeks fiksasi dapat
digunakan untuk mengetahui pola perkawinan dan pola seleksi yang terjadi di
dalam populasi. Nilai indeks fiksasi bisa positif atau negatif, hal tersebut
dipengaruhi oleh adanya inbreeding, seleksi dan kawin yang tidak acak. FIS dan
FIT adalah korelasi antara dua gamet secara acak pada sub populasi dan populasi
total, sedangkan FST adalah kolerasi antar gamet secara acak pada masing-masing
subpopulasi dan digunakan untuk mengukur derajat diferensiasi genetik
subpopulasi (Nei 1987). Nilai FST berkisar dari 0 sampai 1. Wright (1978)
mengelompokkan nilai-nilai FST sebagai berikut: diferensiasi genetik kecil (FST 0
sampai 0.05), diferensiasi genetik moderat (FST 0.05 sampai 0.15), diferensiasi
genetik besar (FST 0.15 sampai 0.25), diferensiasi genetik sangat besar (FST lebih
MATERI DAN METODE
Tempat dan Waktu Penelitian
Penelitian ini terdiri dari dua tahap yaitu pengambilan data ukuran-ukuran
tubuh di lapangan dilanjutkan dengan analisis molekuler di laboratorium. Lokasi
penelitian lapangan ditentukan dengan metode purposive sampling yaitu Kabupaten Pandeglang dan Lebak. Pengambilan data ukuran tubuh dilakukan dari
Bulan September sampai November 2008. Identifikasi polimorfisme lokus
GH/MspI, GHRH/HaeIII dan Pit-1/HinfI dilakukan di Laboratorium Zoologi, Departemen Biologi Fakultas MIPA IPB dari bulan Desember 2008 sampai April
2009.
Kondisi Umum Propinsi Banten
Propinsi Banten terletak pada 5o7’50”-7 o1’11’’ Lintang Selatan (LS) dan 05 o1’11”-106 o’12” Bujur Timur (BT). Luas wilayah propinsi Banten yaitu 9 160.7 km2. Posisi Banten sangat strategis yaitu sebagai penghubung jalur perdagangan Sumatera-Jawa. Secara geografis, batas-batas wilayah propinsi
Banten adalah sebagai berikut:
- Sebelah utara berbatasan dengan Laut Jawa
- Sebelah barat berbatasan dengan Selat Sunda
- Sebelah selatan berbatasan dengan Samudera Hindia
Iklim wilayah Banten dipengaruhi oleh Angin Monson dan gelombang La
Nina atau El Nino. Pada saat musim penghujan (November-Maret) cuaca
didominasi oleh angin barat (dari Samudera Hindia) yang bergabung dengan
angin dari Asia yang melewati Laut Cina Selatan. Pada musim kemarau
(April-Oktober), cuaca didominasi oleh angin timur yang menyebabkan wilayah Banten
mengalami kekeringan yang keras terutama diwilayah bagian selatan (BPS Banten
2008).
Kabupaten Pandeglang
Pandeglang memiliki dataran yang sebagian besar merupakan dataran rendah,
yaitu di daerah bagian tengah dan selatan, dengan variasi ketinggian antara 0
sampai 1 778 m di atas permukaan laut (mdpl). Suhu udara minimum dan
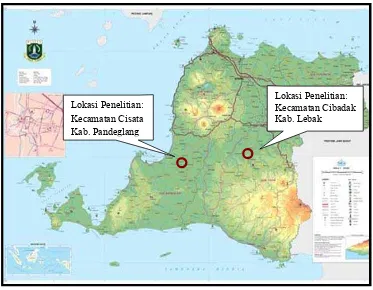
[image:39.595.122.495.213.503.2]maksimum yang terjadi di wilayah Kabupaten Pandeglang berkisar antara 22.5 oC sampai 27.9 oC dengan suhu udara rata-rata untuk dataran rendah adalah 22.9 oC, dan untuk dataran tinggi adalah 22.5 oC (BPS Banten 2008).
Gambar 1 Peta lokasi penelitian di Provinsi Banten.
Kabupaten Lebak
Kabupaten Lebak memiliki luas 304 472 ha. Kondisi Topografi Kabupaten
Lebak bervariasi antara dataran sampai pegunungan dengan ketinggian antara 0
sampai 200 mdpl di wilayah pantai selatan, ketinggian 201 sampai 500 mdpl di
wilayah Lebak tengah, ketinggian 501 sampai 1000 mdpl lebih di wilayah Lebak
timur dengan puncaknya Gunung Sanggabuana dan Gunung Halimun. Keadaan
suhu rata-rata dataran rendah mencapai 27.9 oC dan di dataran tinggi 25 oC dengan kisaran suhu udara antara 24.5 oC sampai 29.9 oC. Rata-rata curah hujan dalam kurun waktu 13 tahun terakhir 3 089 mm/tahun (BPS Banten 2008).
Lokasi Penelitian: Kecamatan Cibadak Kab. Lebak
Pengukuran Karakteristik Ukuran Tubuh
Materi
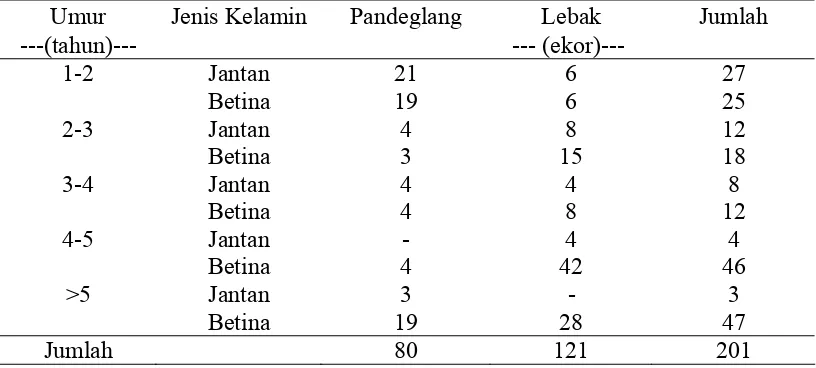
Ternak yang digunakan dalam penelitian ini adalah kerbau rawa (swamp buffalo). Jumlah ternak yang digunakan sebanyak 201 ekor, masing-masing 80 ekor dari Pandeglang dan 121 ekor dari Lebak. Jumlah sampel ternak kerbau
[image:40.595.108.516.248.432.2]menurut umur, jenis kelamin dan asal ternak, disajikan pada Tabel 2.
Tabel 2 Sampel ternak kerbau menurut umur, jenis kelamin dan asal ternak
Umur Jenis Kelamin Pandeglang Lebak Jumlah
---(tahun)--- --- (ekor)---
1-2 Jantan 21 6 27
Betina 19 6 25
2-3 Jantan 4 8 12
Betina 3 15 18
3-4 Jantan 4 4 8
Betina 4 8 12
4-5 Jantan - 4 4
Betina 4 42 46
>5 Jantan 3 - 3
Betina 19 28 47
Jumlah 80 121 201
Peralatan yang digunakan dalam penelitian ini adalah tongkat ukur dan
pita ukur dalam satuan centimeter (cm).
Metode
Data ukuran tubuh diperoleh melalui pengukuran langsung di lapangan.
Penentuan lokasi dan sampel dilakukan dengan metode purpossive sampling. Pada penelitian ini lokasi yang dituju yaitu Provinsi Banten, dengan pertimbangan
bahwa wilayah ini telah ditetapkan sebagai daerah pengembangan ternak kerbau.
Sedangkan pemilihan dua lokasi yaitu Kabupaten Pandeglang dan Kabupaten
Lebak juga dilakukan secara purpossive sampling yaitu tertuju pada lokasi dengan kepadatan ternak kerbau tinggi.
Kerbau dikelompokkan dalam 5 kelompok umur dan jenis kelamin.
Penentuan umur kerbau berdasarkan informasi dari peternak dan berdasarkan
pergantian gigi seri dengan kriteria menurut Lestari (1986) yang disajikan pada
Tabel 3. Kriteria penentuan umur kerbau berdasarkan pergantian gigi seri
Gigi Seri Umur (tahun)
Belum ada gigi tetap (I0) 1
Sepasang gigi tetap (I1) 2
Dua pasang gigi tetap (I3) 3
Tiga pasang gigi tetap (I4) 4
Empat pasang gigi tetap (I5) >5
Peubah yang Diamati
Peubah yang diamati pada penelitian ini adalah karakteristik fenotipe yang
ada hubunganya dengan sifat kuantitatif, diantaranya adalah tinggi pundak (TP),
panjang badan (PB), lingkar dada (LD), dalam dada (DD) dan tinggi pinggul
(TPg).
Gambar 2 Skema pengukuran tubuh kerbau, 1) panjang badan, 2) tinggi pundak, 3) lingkar dada, 4) dalam dada, dan 5) tinggi pinggul.
Adapun metode pengukuran kelima kriteria tersebut adalah:
1. Tinggi pundak (TP) diukur dari permukaan tanah sampai titik tertinggi
pundak (scapula) dengan menggunakan tongkat ukur dalam satuan cm. 2. Panjang badan (PB) diukur dari sendi bahu (humerus) sampai tulang duduk
(Tuber ischii) dengan menggunakan tongkat ukur dalam satuan cm.
[image:41.595.162.433.359.560.2]4. Dalam dada (DD) diukur dari titik tertinggi pundak dan tulang dada, diukur
dengan menggunakan tongkat ukur, satuan dalam cm.
5. Tinggi pinggul (TPg) diukur dari titik tertinggi pinggul secara tegak lurus ke
tanah, diukur dengan menggunakan tongkat ukur, satuan dalam cm.
Analisis Data
Analisis terhadap data ukuran-ukuran tubuh kerbau dilakukan secara
statistik deskriptif, meliputi nilai rataan, simpangan baku dan koefisien
keragaman. Formula yang digunakan adalah sebagai berikut (Steel & Torrie
1993):
;
;
Keterangan : = nilai rataan,
xi = ukuran ke-i dari peubah X,
n = jumlah contoh ternak kerbau yang diamati dalam populasi, KK = koefisien keragaman,
SE = standar eror, dan s = simpangan baku.
Perbandingan ukuran-ukuran tubuh antar dan dalam populasi kerbau
dianalisis dengan menggunakan uji-t dengan formula sebagai berikut:
keterangan : th = nilai hasil uji-t,
= nilai rataan sampel dari populasi pertama,
= nilai rataan sampel dari populasi kedua,
= simpangan baku gabungan,
n1 = jumlah sampel dari populasi pertama, dan
n2 = jumlah sampel dari populasi kedua.
Analisis uji-t ini dilakukan dengan bantuan perangkat lunak MINITAB 14.
Identifikasi Polimorfisme Gen GH, GHRH dan Pit-1
Materi
Sampel Ternak Kerbau
Sampel ternak kerbau yang digunakan untuk analisis keragaman lokus
GH/MspI, GHRH/HaeIII dan Pit-1/HinfI sebanyak 77 ekor yang terdiri dari 44 ekor dari Pandeglang dan 33 ekor dari Lebak. Pengambilan sampel dilakukan
secara purpossive sampling yaitu tertuju pada lokasi dengan kepadatan ternak kerbau tinggi.
Primer
Primer adalah DNA utas tunggal dengan ukuran pendek, biasanya 18
sampai 25 pb (pasang basa), yang akan menempel pada DNA cetakan pada tempat
yang spesifik. Pasangan primer digunakan untuk mengapit sekuen DNA target
pada reaksi PCR. Informasi primer yang digunakan pada penelitian ini
[image:43.595.110.510.591.723.2]ditunjukkan pada Tabel 4.
Tabel 4 Informasi sekuen primer yang digunakan dalam penelitian
No Lokus Sekuen primer sumber
1 GH/MspI F : 5’-CCCACGGGCAAGAATGAGGC-3’ R : 5’-TGAGGAACTGCAGGGGCCCA-3’
Mitra et al. (1995)
2 GHRH/
HaeIII
F : 5’-GTAAGGATGCCAGCTCTGGGT-3’ R : 5’-TGCCTGCTCATGATGTCCTGGA-3’
Moody et al
(1995)
3 Pit1/
HinfI
F : 5’-AAACCATCATCTCCCTTCTTCTT-3’ R : 5’-AATGTACAATGTGCCTTCTTCTG-3’
Wollard et al
(1994)
1
x
2
x
2 1 x
Metode
Beberapa tahapan yang dilakukan meliputi pengambilan sampel darah,
isolasi DNA, amplifikasi gen dengan teknik polymerase chain reaction (PCR), genotiping dengan teknik restriction fragment lenght polymorphisms (RFLP), dan visualisasi pita DNA.
Tahap 1. Pengambilan Sampel Darah
Darah diambil dari vena jugularis sekitar 2 ml dengan menggunakan tabung vacutainer berheparin. Sampel darah tersebut selanjutnya direndam dalam
etanol 95% untuk menghindari kerusakan sel-sel darah.
Tahap 2. Isolasi DNA Total
DNA diisolasi menggunakan metode fenol kloroform (Sambrook et al.1989). Sampel darah total yang disimpan dalam etanol 95% disentrifugasi 3500 rpm selama 5 menit. Endapan sel-sel darah yang diperoleh dicuci dengan buffer
TE sebanyak 2 kali. Sekitar 100 µl sel-sel darah yang telah bebas dari etanol
disuspensikan dengan 1xSTE sampai volume mencapai 350 µl. Sel-sel darah
kemudian dilisis dengan 20 µl proteinase K (10 mg/ml) dan 40 µl 10% SDS.
Campuran ini dikocok pelan-pelan selama 2 jam pada suhu 55 oC.
Pemurnian DNA dilakukan dengan metode fenol-kloroform, yaitu dengan
menambahkan 1/10 volume 5 M NaCl, 1 x volume larutan fenol, dan 1 x volume
kloroform iso amil alkohol (24:1), kemudian dikocok pelan-pelan pada suhu ruang
selama 2 jam. Fase DNA dipisahkan dari fase fenol dengan sentrifugasi pada
kecepatan 7000 rpm selama 5 menit. Molekul DNA diendapkan dengan
menambahkan 1/10 x volume 5 M NaCl dan 2 x volume etanol absolut. Endapan
DNA yang dihasilkan selanjutnya dicuci dengan etanol 70% kemudian
diendapkan lagi dengan kecepatan 7000 rpm selama 5 menit. Sisa etanol dibuang
dan diuapkan dengan menggunakan pompa vakum. DNA selanjutnya dilarutkan
dengan 80 µl 80% bufer TE.
Tahap 3. Amplifikasi Gen dengan Teknik PCR
Teknik PCR dilakukan untuk memperbanyak (amplifikasi) fragmen gen
perbanyakan ini maka fragmen gen target dapat divisualisasikan pada gel
elektroforesis.
Reaksi PCR dilakukan dengan volume total 25 µl dari campuran larutan
yang terdiri dari 10 sampai 100 ng DNA genom, 1 U enzim taq polimerase dan
10X bufernya (New England Biolab); 2 mM dNTP mix; 2.5 mM MgCl2 dan dH2O
steril. Sedangkan kondisi reaksi PCR dalam mesin thermocycler dirancang dengan suhu pradenaturasi 94 oC selama 4 menit, selanjutnya 30 siklus reaksi yang terdiri dari denaturasi 94 oC selama 10 detik, annealing (suhu spesifik primer) selama 1 menit, perpanjangan 72 oC selama 2 menit. Pemanjangan akhir pada suhu 72 oC selama 7 menit. Suhu annealing untuk primer lokus GH/MspI, GHRH/HaeIII, dan Pit1/HinfI secara berurutan yaitu 62 oC, 60 oC dan 60 oC.
Tahap 4. Genotiping Teknik RFLP
Produk PCR selanjutnya dipotong dengan enzim restriksi yang spesifik
dengan gen tersebut. Enzim restriksi yang digunakan untuk gen GH, GHRH dan
Pit-1 secara berurutan yaitu MspI, HaeIII dan HinfI. Enzim restriksi MspI mengenali situs restriksi C*CGG, sedangkan enzim restriksi HaeIII dan HinfI masing-masing mengenali situs GG*CC dan G*ANTC. Kondisi pemotongan
dilakukan menurut produsen enzim yaitu 2 µl produk PCR dicampur dengan 1
sampai 2 unit enzim restriksi dalam 1xBufer (New England Biolabs) dan
selanjutnya diinkubasi pada suhu 37 oC selama minimal 4 jam.
Tahap 5. Visualisasi pita DNA
Visualisasi pola pita hasil RFLP menggunakan elektroforesis gel
poliakrilamid 6% yang diikuti dengan metode pewarnaan perak (silver stainning). Gel dibuat dengan cara mencampurkan 12 ml air destilata, 4 ml larutan 5 x TBE,
4 ml larutan akrilamid 30%, 15 µl larutan TEMED, dan 160 µl APS 10%.
Sebanyak 2 µl produk RFLP dicampur dengan + 6 µl loadying dye (bromthymol blue 0.01%, xilene cyanol 0.01 dan gliserol 50%). Elektroforesis dilakukan pada
tegangan konstan 200 mVolt selama 40 menit. Setelah elektroforesis selesai, gel
diambil untuk dilakukan pewarnaan perak.
Tahapan pewarnaan perak yaitu gel dicuci secara bertahap sebagai berikut:
dengan larutan CTAB 0.2 gram/200 ml air destilata selama 8 menit, air destilata
perak nitrat (AgNO3) selama 10 menit dan air destilata selama 2 x 2 menit. Untuk
memunculkan pita dalam gel, gel direndam dalam larutan yang terdiri atas
Na2CO3 dan 38% formadehid. Setelah pita muncul, larutan asam asetat
dituangkan untuk menghentikan aktifitas oksidasi perak oleh formadehid.
Analisis Data
Analisis data molekuler lokus GH/MspI, GHRH/HaeIII dan Pit-1/HinfI pada populasi kerbau Banten meliputi frekuensi alel, keseimbangan
Hardy-Weinberg, nilai heterosigositas meliputi heterosigositas pengamatan (Ho) dan
heterosigositas harapan (He), dan nilai-nilai indeks fiksasi meliputi FIS, FIT dan
FST.
Frekuensi Alel
Frekuensi alel untuk setiap lokus dihitung berdasarkan petunjuk Nei
(1987) dengan formula sebagai berikut:
n n n
x j i
ij ii i 2 2 ⎟⎟ ⎠ ⎞ ⎜⎜ ⎝ ⎛ + =
∑
≠ Keterangan :xi = frekuensi alel,
nii = jumlah genotipe dari alel ke-i, dan
nij = jumlah alel ke-i terpaut alel ke-j (j≠i).
Keseimbangan Hardy-Weinberg
Keseimbangan Hardy-Weinberg diuji dengan chi-square (χ2) menurut Hartl (1988), sebagai berikut:
Keterangan:
χ2
= nilai chi-square,
Obs = jumlah pengamatan genotipe ke-i, dan
Exp = jumlah harapan genotipe ke-i
∑
≠−
=
j iExp
Exp
Obs
X
22
(
)
Nilai Heterozigositas Pengamatan (Ho)
Nilai heterosigositas teramati (Ho) dan heterosigositas harapan (He) dapat
digunakan untuk menduga nilai koefisien inbreeding pada suatu kelompok ternak. Perhitungan nilai Ho dan He dilakukan menurut petunjuk Hartl (1988) dengan
formula sebagai berikut:
∑
≠=
j iN
ij
N
Ho
1 , Keterangan :Ho = heterosigositas pengamatan,
N1ij = jumlah individu heterosigot pada lokus ke-1, dan N = jumlah individu yang diamati.
Nilai Heterozigositas Harapan (He)
∑
=−
=
n i iP
He
1 2 11
Keterangan :He = heterosigositas harapan,
P1i = frekuensi alel ke-i pada lokus ke-1, dan n = jumlah alel pada lokus-1.
Indeks Fiksasi
Nilai indeks fiksasi yang dikenal dengan uji F-statistik dapat digunakan
untuk mengetahui sistem perkawinan dan pola seleksi pada populasi yang diamati.
Indeks fiksasi terdiri atas tiga parameter yaitu FIS, FIT dan FST yang dihitung
berdasarkan petunjuk Nei (1987) dengan formula sebagai berikut:
Keterangan:
= rataan frekuensi genotipe homosigot,
= rataan frekuensi alel yang sama,
= rataan kuadrat frekuensi alel, dan
= kuadrat rataan frekuensi alel.
Uji Hubungan Genotipe dengan Ukuran Tubuh
Uji hubungan genotipe dengan ukuran tubuh kerbau dilakukan dengan
analisis uji-t. Data ukuran-ukuran tubuh untuk uji genotipe dikelompokkan menjadi tiga kelompok umur yaitu kelompok umur 2 sampai 3 tahun, 4 sampai 5
tahun, dan lebih dari 5 tahun. uji-t dilakukan dengan bantuan perangkat lunak
MINITAB 14.
Sebelum dilakukan uji hubungan genotipe dengan ukuran-ukuran tubuh,
data ukuran tubuh dikoreksi terlebih dahulu. Adapun koreksi dilakukan melalui
dua tahapan koreksi. Pertama dilakukan koreksi dengan patokan kerbau betina
umur 4 sampai 5 tahun untuk setiap daerah. Kedua, data ukuran tubuh kerbau
Lebak yang telah terkoreksi ke data kerbau betina umur 4 sampai 5 tahun,