ANALISIS KERAGAMAN GENETIK
Phytophfhora
palmivora
PENYEBAB BUSUK BUAH PADA KAKAO
DI INDONESIA
OLEH
:
ABU
UMAYAH
SEKOLAH
PASCASARJANA
INSTITUT PERTANIAN BOGOR
ANALISIS KERAGAMAN GENETIK
Phytoplrthora
paIrnz*vora
PENY EBAB BUSUK BUAH PADA KAKAO
DI INDONESIA
OLEH
:
ABU UMAYAH
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRAK
Abu Umayah. Analisis Keragaman genetik Phytophihora palmivora
penyebab busuk buah pada kakao di Indonesia. Di bawah bimbingan Meity Suradji Sinaga sebagai ketua, Sarsidi Sastrosumarjo, Sientje Mandang Sumaraw, Agus Purwan tara sebagai anggota.
Penelitian ini bertuj uan untuk menganalisis intensitas penyakit busuk buah kakao di kebun akibat infeksi I-'. puimivoru yang terjadi secara alami, meIakukan
identi fikasi species Phytuphrhoru yang menyerang tanaman kakao di Indonesia. melakukan evaluasi tingkat virulensi beberapa isolat
P.
pulmivora dm resistensi beberapa klon kakao serta melakukan analisis keragarnan genetik beberapa isolatP.
pulmivoru yang dikumpulkan dari berbagai provinsi penghasil kakao di Indonesia.Monitoring d m evaluasi penyakit dilaksanakan di Larnpung Selatan, Cianjur dan Jember menggunakan metode survei dengan teknik pengambilan contoh secara
purposive. Perkembangan penyakit diukur berdasarkan kejadian penyakit dan
keparahan penyakit yang terjadi di lapangan. Data juga dikumpul kan dengan bantuan daftar pertanyaan. Ident i fikasi P. palmivoru didasarkan pada si fat morfologi
(tipe koloni, pem bengkakan hi fa, ada tidaknya klamidospora, percabangan
sporangiofor, bentuk dan ukuran sporangia, cadukous, panjang pedisel dan papila) dan mole kuler menggunakan tekni k ITS (Inlerrwl 1 iumcribeci Spacer). Uj i virulensi
dan resistensi disusun menggunakan rancangan acak kelompok, dengan 24 perlakuan kombinasi antara enam isoIat P. puZmivorcr yajtu isolat Sumut, Lampung, Jabar,
Jatim, Sulsel dan Sultra dan empat klon kakao yaitu SCA 12,
DRC
16, DR 2 dan GC 7 dm diulang tiga kafi. Peubah yang diamati meliputi periodeinkubasi,
luas bercakdan laj u in feksi. Analisis keragaman geneti k P. pulmivora dilakukan menggunakan teknik RAPD dan AFLP.
Kej adian pen yaki t d m keparahan pen yaki t pada perkebunan kakao swasta di Cianjur lebih bear dibandingkan pada perkebunan kakao rakyat di Lampung Selatan dan perkebunan kakao negara di Jem ber. Kedua variabel tersebut sangat dipengaruhi
oleh kondisi lingkungan yang kondusi f untuk perkembangan penyaki t terutama oleh
curah hujan (>2500 mmltahun) dan kelembaban relatif yang tinggi (>900/b) dan suhu yang rendah
(*z?c).
Berdasarkan pada sifat-sifat morfologi dan molekuler 20 isolat Fang diuji diketahui adalah 1'. pulmivoru sebagai penyebab busuk buah dan kanker batang kakao di Indonesia. Tingkat virulensi isolat berturut-t urut adalah isolat Sultra, Jatim,Sumut,
Jabar, Lampung dan isofat Sulsel. Sedangkan tillgkat resistensi klon beflurut-turut adalah klon SCA 12, DRC 16, DR 2 , dan kIon GC 7.Adanya perbedaan tinghat periode inkubasi, luas bercak dan Iaju infeksi pada klon- klon kakao yang diuji mengindikasikan bahwa a& perkclaan ras fisiologik diantara isolat-isolat. Sedangkan isolal-isolat
P.
pulmivoru mernpunyai keragaman genetik yang rendah, walaupun letak geografisnya berjauhan. Analisis menggunakan teknik AFLP lebih efisien, konsisten dan lebih disknminatif dibandingkan menggunakan teknik RAPD.ABSTRACT
A bu Umayah. The analysis of genetic variations of Phytop/zthoru pulntivoru causing pod rot on cocoa in Indonesia. Under supervisions of Meity Suradji Sinaga as chairman, Sarsidi Sastrosuma j o , Sientje Mandang Surnaraw, Agus Purwantara as members.
The objectives of this research were to determine the disease intensity of pod rot in cocoa plantations in Lampung Selatan, Cianjur and Jember, to identify
Yl1ytophlhora species amclung cocoa in Indonesia, to evaluate the virulence of isolates of P/yiophthom sp. and the resistance of selected cocoa clones, and to analyse genetic variation of isolates of Phytophthorca sp. collected from several cocoa producing provinces in Indonesia.
Monitoring and evaluation of disease were carried out in Larnpung Selatan
(Lampung), Cianjur (Jawa Barat) and Jember (Jawa Timur) using survey method
l l t h purposive sampling technique. Development of disease was evaluated on the base of disease incidence and disease severity of cocoa pod caused by natural infection. Data were also collected through interview with farmers or plantation managers. The pathogen was identified based on morphological characteristics (colony type, hyphal swellings, production and diameter of chlamydospores,
sporangiophore branching, shape and size of sporangia, caducity, pedicel length and
papi llate) and on molecular c haractenstic using Internal Transcribed Spacer (ITS) technique. Test of virulence and resistance were conducted in the field using randomized complete block design, with 24 treatment combinations between six isolates of the pathogen from Sumatera Utara, Larnpung, Jawa Barat, Jawa
Timur,
Sulawesi Selatan and Sulaivesi Tenggara and four cocoa clcnes namely SCA12,DRC16, DR2 and GC7, and each replicated three times. Latent period, ex-tent of blight and infection rate were recorded to determine degree of virulence of the pathogen and resistance of cocoa clones. Genetic variation was analysed using Randomly Amplified Polymorphic DNA (RAPD) and Amplified Frabment Length Polymorphism (AFLP) techniques.
Isolates
of
P.
pulmivora showed low genetic variations, eventhough the isolates werefrom separated geographical origin. AFLP t e c h q u e was more efticient, consistent and discriminative
than
RAPD technique.SURAT
PERNYATAAN
Saya rnenyatakan dengan sebenar-benarnya bahwa segda pernyataan dalam
disertasi saya yang be j udul :
"Analisis Keragaman Genetik Phytophthora palmivora Penyebab Busuk
Buah Pada Krmkao di Indonaia"
rnerupakan gagasan atau hasil penelitian disertasi saya sendiri, dengan pembimbingan para komisi pembimbing, kecuali yang dengan jeIas ditunjukkan
rujukannya. Disertasi ini belum pernah diajukan untuk memperoleh gelar pada
program sejenis
1
perguruan tinggi lain.Semua data dan informasi yang digunakan telah dinyatakan secara jeIas dan
dapat dipri ksa ke benarannya.
Bog& mber 2004.
ANALISIS
KERAGAMAN GENETIK
Phytuphthora
palmivora
PENYEBAB BUSUK BUAH PADA KAKAO
DI INDONESIA
OLEEI : ABU UMAYAH
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Entomologi dan Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : ANALISIS KERAGAMAN GENETK Ykylophthora plmivora PENYEBAB BUSUK BUAH PADA
KAKAO
Dl INDONESIA
Nama : Abu Umayah
NRP
: 995234Program Studi : Entomologi
dan
FitopatologiMenyetujui,
1. Komisi Pembimbing
,
d
-
.
-Prof Dr. Ir. Sarsidi Sastrosummo
Ketua h g g o t a
Prof. Dr. Ir. Sienti e Mandang Surnaraw Dr.Ir.Agus Purwantara. APU
Ansgota Anggota
Mengetahui,
2. Ketua Program Studi ENT-FIT
gg&J&&
&A&
Dr.
Er.
Sri Hendrashrh Hiday at. MSc.IUWAYAT
HIDUP
Penulis dilahirkan di Kota Palembang Provinsi Sumatera Selatan pa&
tanggal 25 Nopember 1958, sebagai anak ke tujuh dari pasangan Bapak H.
Muhammad Ali Pisol (Almarhum) dan Hj . Fatimah (Almarhumah). Penulis
rnenempuh pendidikan
S
1 di Fakultas Pertanian Universi tas Sriwijaya (UNSRI),Departemen Proteksi Tanaman,
dengan
mata ajaran pokok (major)Umu
PenyakitTumbuhan dan mata ajaran pilihan (minor) Ilmu Tanaman Tahunan (KT) dan lulus
tahun
1982.Pads
tahun 1986 penulis mendapat tugas belajar S2di
FakultasPascasarjana Universitas Gadjah Mda, Jurusan Ilmu-Ilmu Pertanian,
Program
StudiFitopatologi dan
IuIus
pada tahun 1 989. Kesempabn untuk rnelanjutkanke
programdoktor
pa& program studi Fitopatologi di Sekolah Pascasarjana IPB pa& tahunaj aran 1 999/2000, dengan beasiswa pendidikan
di
peroleh dari proyek DUE-Li keUniversitas Sriwijaya
Penulis mulai merintis men& pengalaman bekerja diawali dari tahun 1980
sebagai asisten dosen dalam praktikum mata kuliah ITT, Fisiologi
Tumbuhan,
Mikrobiologi, Mikologi, Bakteriologi dan Fitopatologi. Sejak tahun 1982, setelah
lulus S 1 penulis tercatat sebagai Dosen Faperta Unsri dan baru tahun 1985 diangkat
menjadi PNS sebagai Dosen Faperta Unsri sampai sekarang. Penulis pernah juga
memberi kuliah pada Fakultas Pertanian Universitas Sj akyakirti, Universi tas
Muhamadiyah, Universitas Palembang, Universitas Tamansiswa dm Program
PRAKATA
Puji syukur penulis panjatkan ke hadirat Allah
SWT
atas berkat rahmat clanhidayah-Nya, pendis dapat menyelesai kan disertasi yang be j udul Analisis
Keragaman geneti
k
Phyiophthora palmivora penyebab busuk buah pada kakao diIndonesia. Disertasi im disusun untuk meIengkapi syarat mernperoleh gelar Doktor
pada Sekolah Pascasarjana Institut Pertanian Bogor. Selain sebagai syarat
memperoleh gelar Doktor, harapan penulis disertasi ini dapat memberikan
sumbangan pernibran Mam rangka rneningkatjran produksi dan mutu bkao
Indonesia. Dalam disatasi ini diuraikan
dan
dijelaskan mengenai waiuasi penyalut,W e r i s t i k patogen, virulensi isolat, resistensi klon,
dan
keragaman genetikP.
paImivora sebagai penyebab penyalut
busuk
buahkakao,
yang kesemuanya sangatbermanfaat dalam menyusun program pengelolaan penyakit yang disebabkan oleh P.
palmivora pada perkehan
kakao
yang di kelola oleh rakyat, swasta clan pemerintah.Dengan sel esainya disertasi ini, penul is mengucapkan terima kasih kepada
semua pihak yang telah membantu mulai dari masa perkuliahan, selama penelitian
sampai akhir proses belajar di Institut Pertanian Bogor yaitu:
I . Dr.Ir. Meity Suradji Sinaga, MSc. sebagai ketua komisi pmbirnbing yang telah
memberikan bantuan bahan-bahan acuan, dorongan, bimbingan, pengarahan,
petunjuk clan saran-saran yang ilmiah, kritis dan teliti kepda penulis selama
2. Prof
Dr.
Lr.
Sarsidi Sastrosurnarjo sebagai anggota komisi pembimbingyang
telahmemberikan bimbingan, petunjuk dan saran-saran ilmiah yang loltis dan teliii
dalam penyelesaian disertasi ini
3. Prof.
Dr.
Ir.
Sientje Mandang Sumaraw sebagai anggota komisi pembimbing yangtelah memberikan bimbingan dan saran-sarannya mulai dari penyusunan
proposal, sampi pembuatan disertasi ini.
4. Dr.Ir.Agus Punvantara, APU sebgm anggota komisi pembimbing yang telah
memberikan bantuan
bahan-bahan
penelitian, Wan-bahm acuan, bimbingan,arahan-arahm serta petunjuk yang ilmiah, kritis clan teliti mulai dan rencana pembuatan proposal, selama penelitian sampi penyusunan
disertasi
ini.5 . Prof.
Dr.
1r.H. Zainal Ridho Djafar, Rektor Universitas Sriwijaya yang telahmemkrikan
izin d m dorongm kepada penulis untuk mengikuti program S3 dihstitut Pertanian Bogor.
6 . Ir-H-Lukman Hakirn Taslim,
MS
mantan Dekan Faperta Unsri,dan
1r.Hj.ZuljatiSjahrul, MSc. Dekan Faperta Unsri yang tefah memberikan izin dan dorongan
kepada penulis untuk mengikuti program S3
di
IPB Bogor.7. Dr.Ir.Rujito Agus Suwignyo, M. Agr. rnantan Direktur Eksekuti f Proyek DUE-
Like UNSRI, dan Drs.A.Rachman Ibrahim, M.Ed. Direktur Eksekutif Proyek
8. Dr.Ir.Supaman SHK dan Ir. Suwandi, M.Agr. kolega yang telah banyak
memberi
kan
bantuan dan domngan kepada penulis selama mengikuti program S3dl
htitut Pertmian Bogor.9. Ir.Basuki, MS mantan Kepala
UPBP,
dan Dr.Ir.Darmono Taniwiryono, MSc. kepalaBdai
Peneiitian Bioteknologi Pekebunan, Bogor yang telah memberikanizin dan fmilitas selama proses penelitian
dl
lembaga tersebut.10. Direktur Utarna PT. Intergreen Estate, Jakarta ymg
telah memberikan
izin
dm
bmtuan untuk rnelakukan penelitian di kebun kakao Cianj
ur,
Jawa Barat.1 1 . Direksi
PTPN
XU J1. Rajawali 44 Surabaya yang telah memkrikanizin
dan bantuan mtuk rnelakukan penelitiandi kebun
kakao
RentengJ1.
Gajah Mads249 Jemkr, Jawa Timur.
12. Kepla Pusat Penefitian Kopi
dm
Kakao Indonesia J1. P.B. Sudirman 90 Jember,Jaw Timur yang telah memberikan kin dan fasilitas dalarn pelaksanaan
penelitian di instansi tersebut.
13. Lr. Sri Sukamto Sugarto,
MP
yang telah memberikan bantuan terutama bahan-bahan acuan dm saran-sarannya yang sangat baik kepada penulis selama
penelitian
di
Puslit Kopi dan kakao, di Jember.14. Tolhas Hutabarat, Dipl. Kim., Dra. Nurhairni, MSi dan Neda yang telah banyak
rnemberi kan penjelasan, masukan, saran-saran seputar tekni
k
analisis molekulerdm bantuannya selama penulis melakukan penelitian di labomtoriurn Biologi
15. Bapak Mamak, Ddy, Supri, Rahrnat, Udin,
Main,
Hj. Em% dan Mamay ymgtelah memberikan bantuannya dalarn
pefaksanaan
penelitiandi
Laboratorium Mikmba BPBP, Bogor.16. Bapak Prof.Dr.lr.Sudiman Yahya, MSc. sebagai penguji luar komisi pada ujian
tertutup tanggal 18 Agustus 2004, Dr. lr. Darmono Taniwiryono, MSc. dan Dr.Ir.
Gede Suastika x b g m penguji luar kornisi pada ujian terbuka tanggal 28
September 2004 yang telah rnemberikan saran-saran, komentar m u m
dm
komentar spesifihya kepada pendis.
17. Istri tercinta
Ir.
Numini Arsianty, MSi yang klah memberikan dorongan moral, pengorbanan yang tak temilai clan do'a selama pendis mengkuti kuliah, melaksanakan penelitian clan penyusunan disertasi ini. Atas dorongannyalahpenulis timbd semangat untuk term melanjutkan studi program S3 in^.
Demikian juga anak-anak saya Rend Febrianda, W s t a Febra Nugraha,
dan
Marsian Gustnanda yang terus-menerus memberikan dukungan moral
d m
selaiutabah clan sabar menanti papanya dapat menyelesaikan tugas belajar ini dengan
ANALISES KERAGAMAN Phytophthora palmivora PADA
...
TANAMAN KAKAO DI INDONESIA
...
Pendahuluan... Bahan dm Metode
... Hasil
... Pembahasan
... Kesim pulan
... KESIMPULAN DAN SARAN
...
Kesimpulan
... Saran
...
DAFTAR TABEL
Halaman Katagori penilaian keparahan penyakit (1P) bus& buah kakao . . .
.
23Evaluasi kondisi penyakit, deslcripsi kebun dan desknpsi tanaman
kakao perkebunan rakyat di Lampung
.. . .
..
. . ...
. . .
..
. . . .. . .
26EvaIuasi kondisi penyakit, desknpsi kebun dan deskripsi tanaman
kakao perkebunan swasta milik PT Intergreen Estate Cianjur,
Jawa Barat
. . . ...
.. . .
, . . .. . .
. . .. . .
. . .. . .
28Evaluasi kondisi penyakit, deskri psi kebun dm deskripsi tanaman kakao pericebunan negara miIik
PTP
Nusantara XII Jernber JawaTimur
. . .
. . ..
.. . .
. . .. . .
. . .. . .
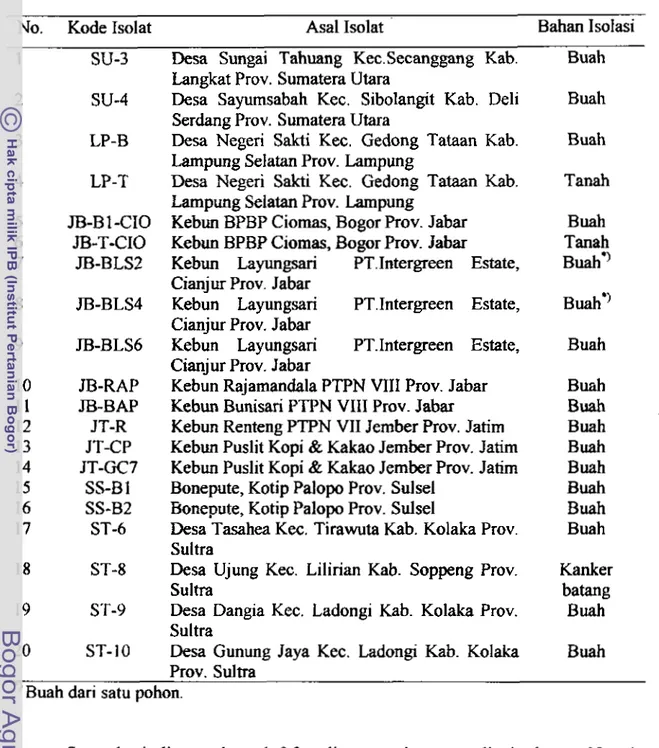
. 30Daftar isolat
P.
palmivoru asal enam provinsi di Indonesia ..
.. . .
45Sekuen primer ITS4
dan
ITS 5 ... ......
... ... ... ...
... ... ...... ... ...
...
47 Campuran reaksi PCR untuk analisis ITS... . ..
..
. . ...
. .. . .
. . ... 48Karakteristik aseksual dua puluh isolat P. paImivora . . .
. . . .. . .
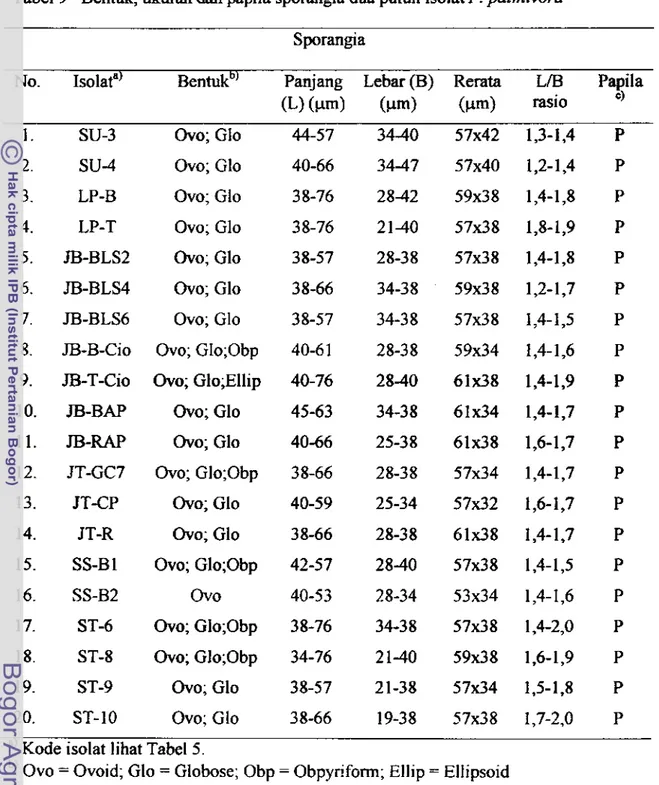
50Bentuk, ukuran clan papila spomga dua puluh isolat P.
plmivora
. . .
. . .
. . .. . .
. .. .
. . .. . .
5 1Profi l program PCR untuk reaksi arnplifi kasi selektif (GibcoBRL-
Life Technologies)
... ... ... ...
...
... ... ... ... ...... ...
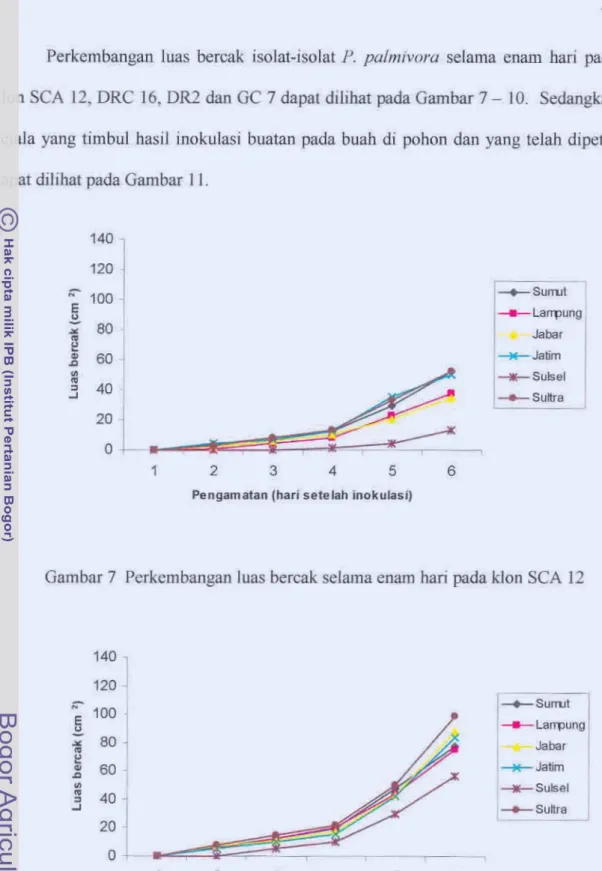
... ... ... ... ... ... . 66 Rerata luas bercak buah dan luas kanker batang hasil inokulasi 37isolat P. palmiwra
.. . .
. . ...
. . . .. . .. . ..
.. . . . .. . .. . .
..
. . . .. . . 70Rerata periode inkubasi (hari), luas bercak (cm2) dan laju infeksi (unitlhari) kombinasi perlakuan enam isolat P. palrnivora dan
ernpat klon kakao . .. . .
.
. . ..
. . . 74Ragarn genetik
(s2)
dm simpangan baku (S) enarn genotipe isolat14
.
Seleksi primer RAPD untukP
palmivora...
79
...
1 5
.
Jurnlah pita hasil amplifikasi dengan teknik
RAPD 80...
16
.
Seleksi pasangan primer AFLP untuk Ppalmivoru $4I 7
.
Jurnlah pita hasil ampiifikasi selekti f dengan teknikAFLP
Jurnlah...
DAFTAR GAMBAR
Buah kakao sehat (A), gejda penyakit busuk buah (B-E)
dan
kanker batang (F) pada tanaman kakao akibat infekiP.
palmivora ... 25
Rerata persentase kejadian penyaki t (KP), kepahan penyakit (IP) bus& buah dan curah hujan rerata 10 tahun terakhir
di
beberapa perkebunan kakao ... 3 1
Tipe koloni P. plmivora pada PDA umur 12 hari (A); sporangia mempunyai papila yang jelas, dengan tangkai
pendek
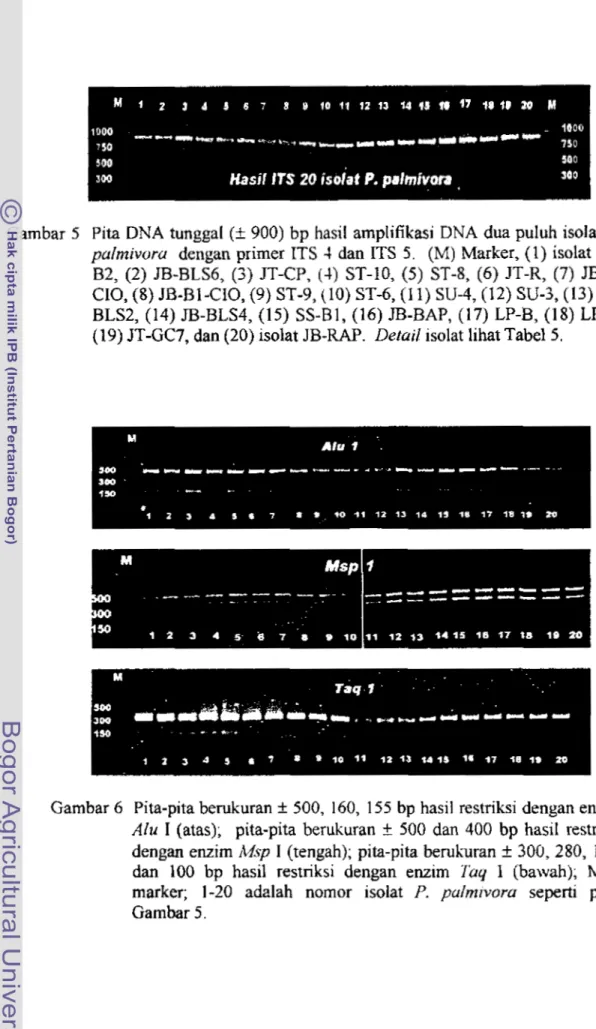
(+ hitam) dan ktamidospora (-+ merah) perbesaran 200 x (kanan, B-D)Pita DNA tunggal (+ 900) bp hasil amplifikasi DNA dua puluh isolat P. plmivora dengan primer ITS 4 dan ITS 5 . (M)
Marker, ( I ) iso1at SS-B2, (2) JB-BLS6, (3) JT-CP, (4) ST-10, ( 5 ) ST-8, ( 6 ) JT-R, (7) JB-T-CIO, (8) JB-B1 -CIO, (9) ST-9, (lo)
ST-
6 , ( 1 I ) SU-4, (12) SU-3, (13) JB-BLS2, (14) JB-BLS4, (15) SS- Bl, (16) IB-BAP, (17) LP-B, (18j LP-T, (19) ST-GC7, dan (20)
...
isolat JB-RAP. Derail isolat fihat Tabel 5 52
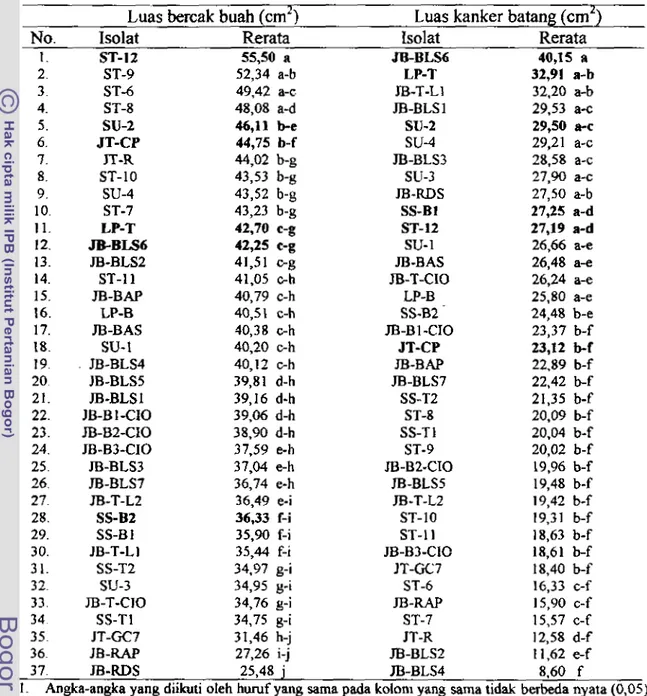
Pita-pita krukuran
+
500, 160, 155 bp hasil restriksi dengan enzim Alrr 1 (atas); pita-pita berukuran f 500 clan 400 bp hasil restnksi dengan enzim Msp 1 (tengah); pita-pita bemkuran k 300, 280, 1 50, dan I00 bp hasil restri ksi dengan enzim Tuq 1 (bawah);M
= marker; 1-20 adalah nomor isolat 1". paImivoru seperti padaGarnbar 5 ... 5 2
Perkembangan Iuas bercak seIama enam hari pada klon SCA 12 .. 71 Perkem bangan luas bercak selama enam hari pa& klon DRC 1 6
.
7 1...
Perkembangan luas bercak selarna enarn hm pa& klon DR 2 72
...
Gejata coklat kehitaman pada buah
di
pohonkakao
klon SCA 12 (A), DRC 16(B),
DR 2 (C) dan GC 7(D).
Gejala yang sarna pada buah yang telah dipetik hasil inokulasi buatan dengan isolat...
P. palmivora (E)Rerata periode inkubasi enam isolat
P.
paimivora pada empat klonkakao
...
Rerata luas bercak enarn isolat
P.
palmivora pada empat ldon...
kakaoRerata laju infeksi enam isoiat P. palmivora pada empat klon kakao ...
Tiga puluh empat primer RAPD yang diseleksi, yang diguaakan addah (14) OPH 12, ( 1 7) OPH 19, (26) OPB 1 1, (30) OPN 06,
...
dan (3 1 ) OPN 1 0
Dendrogram kesamaan genetik 20 isolat P. palmivora hasil analisis RAPD menggunakan 5 primer ...
Analisis komponen utama (Principal Component Analysis, PCA) kesamaan genetik 20 isolat
P.
palmivora menggunakan 5 primer...
RAPD
Sepuluh pasang primer AFLP yang digunakan ( 1 ) E-AC/M-CAT;
(2) E-ACIM-CTC, (3) E-ACIM-CTG, (4) E-AGM-CTG, ( 5 ) E- AGIM-CAG, (6) E-AGM-CTC, (7) E-ACIM-CAA, (8) E-
ACIM-CAC, (9) E-AG/M-CTT, ( 1 0) E-AT/M-CAC,
(M)
markerDendrograrn kesamaan genetik 20 isolat
P.
palmivora hasil analisis AFLP menggunakan 10 pasangan primer ...Anal isis kom ponen utama (Principul Component A nulysis, PCA) kesamaan genetik 20 isolat P. pulmivoru menggunakan 10
DAFTAR LAMPJRAN
Daftar pertanyam dalam s w e i penyakit busuk buah pada tanaman kakao ...
Analisis ragam pengaruh lokasi
kebun
di Lampung, Cianjur dan Jember terhadap kejadian penyakitbusuk
buah pada kakaoAnalisis ragam pengaruh lokasi kebun
di
Lampung, Cianjur danJember terhadap keparahan penyakit bus& buah pada
kakao
Analisis ragam pengaruh 37 jenis isolat P. palmivoru terhadap luas bercak yang ditimbulkan pa& buah kakao klon Amelonado
Analisis ragam penganrh 37 jenis isolat
P.
pulmivora terhadap luas kanker batang yang ditimbulkan pada pohon kakao kIon Arnelonaclo.. ...Analisis ragam pengaruh 24 p e r l h n isolat x Uon terhadap
p r i d e inkubasi (PI), luas bercak (LB) clan laju infeksi (LI)
penyaki t
bus&
buah kakao ...Prof11 pita DNA hasil amplifikasi
dua
puluh (1-20) isolatP.
pafmivora dengan primer OPi3 1 1,(M)
marker ...Profil pita DNA hasil amplifikasi dua puluh 11-20] isolat
P.
...
palmivclra dengan primer
OPH
19,(M)
markerProfil pita DNA hasil amplifikasi selektif sepuluh ( 1
-
10) isolat P....
pulmivora dengan pasangan primer E-AC/M-CAT,(M)
markerProfil pita DNA hasil amplifikasi selektif sepuluh (1 1-20) isolat
P.
palmivora dengan pasangan primer E-ACM-CAT, (M) markerProfil pita DNA hasil amplifikasi selehf sepuluh ( 1
-
10) isolat P.12. ProfilpitaDNAhasiIamplifilcasiselektifsepuluh(l1-20)isolatP.
palmivoru dengan pasangan primer E-ACIM-CTC,
(M)
marker. .
.
11813. Matrik hubungan kesamaan genetik dua puluh isolat P. pulmivorir
hasil RAPD
. . .
..
. . ..
..
.. . .
. . . 11914. Matrik hubungan kesarnaan geneti k dua pul uh isolat I! palmivora
PENDAHULUAN
Latar Belakang
Kakao (Theobrorna cucuu L. ) merupakan salah satu komoditi ekspor diluar
rnigas yang sangat penting sebagai sumber penghidupan bagi jutaan petani produsen
kakao
di
Indonesia, karena produknya merupakan bahan yang sangat diperlukansebagai sumber lemak nabati (kandungan lemak biji kakao k 53%) yang banyak
dibutuhkan oleh manusia dan juga pada industri kosmetik dan farmasi (Susanto 1994;
Iswanto 200 I ).
Kakao kebanyakan diproduksi di Afrika Barat, sebanyak 69 % produksi dunia hanya berasal dari empat negara yakni Ivory Coast, Ghana, Nigeria
dan
Cameraon.Indonesia merupakan produsen ketiga dengan 14 % produksi dunia setelah Ivory Coast dart Ghana, dimana 80 % produksi Indonesia berasal dari Sulawesi (Lambert 2001). Luas perkebunan kakao rakyat adalah 383,611 ha dengan produksi sebesar
274,732 ton, perkebunan kakao swasta 86,801 ha dengan produksi 2 1,934 ton dan
perkebunan kakao negara 62,355 ha dengan produksi 3 8,583 ton, dengan produksi
rerata nasional adalah 0,629 tonlhaltahun (BPS 1999; Eskes 1999).
Perkebunan kakao sering mengalami berbagai gangguan, diantaranya adalah
gangguan oleh kondisi cuacaliklim, hama dan penyakit tanaman. Saat ini, salah satu
gangguan penting yang perlu mendapat perhatian adalah peny akit busuk buah yang
disebabkan oIeh cendawan Phytuphthoraplmrvora (Butler) Butler. Penyakit busuk
buah dijumpai di semua perkebunan kakao di seluruh dunia dengan menyebabkan
Saat ini, bus& buah @od rot) adalah penyakit y ang terpenting dalam budidaya
kakao
di
Indonesia, bahkandi
kebanyakan negara penghasil kakao (Prior dan Smith198 1 ). Di Lndonesia, besarnya kehiiangan produksi akibat busuk buah sangat krbeda antara kebun yang satu dengan yang lain. Penyakit busuk buah dapat langsung
mengakibatkan penurunan produksi karena buah yang terserang sudah busuk sebelurn
dipanen. Persentase buah busuk adalah 26-56 % di Jawa Barat (Pawirosoernardjo dan Purwantara 1992), 32-52 % di Jawa Tengah (Soemomarto 1972),
dan
35-48 % di Jawa Timur (Situmomg dan Swjatno 1974). Biji yang berasal dsri buah busukapabila diproses iebih lmjut akan menghasilkan biji berwarna hitam dengan mutu
yang
rendah
(Away 1994).Laju perkembangan penyakit busuk buah
kakao
sangat dipengaruhi tingkatpatogenisitas P. palmivora, lingkungan fisik setempat, dan tanggap tanaman kakao
terhadap patogen tersebut (Prior 1978). Oleh karena itu, monitoring dan evaluasi
mengenai ha1 tersebut pada kebun-kebun kakao perlu dilakukan karena sangat
berguna dalam menetapkan strategi pengelolaan penyakit busuk buah kakao yang
spesifik lokasi.
Sebelumnya dilaporkan bahwa busuk buah kakao disebabkan oleh satu spesies
I'l~y~uplztlroru, yaitu
P.
paltnivom. Namun ternyata dij umpai juga beberapa specieslain se pert i 1'. megakayu, P. cup.vic I , P. cirrophthoru, P. heveae, P. megaspermu
yang masing-masing berbeda tingkat virulensinya untuk lokasi yang berbeda
merupakan patogen pada Iebih dari 150 spesies tanaman diantaranya tanaman
kakao,
kelapa, karet, durian, pepaya,
jeruk,
nangka, anggrek, lacla, vanili,dan manggis
@renth 200 1 ), sehingga identifi kasi spesies bai k secara morfologi
dan
molukulermenjadi sangat penting untuk konfirmasi penyebab busuk buah di Indonesia
Beberapa cara pengendalian penyakit busuk buah kakao telah dilakukan,
misalnya dengan mengurangi kelernbaban
kebun,
mernpertahankan seresah sebagaimulsa
di
sekitar pangkal batang, memanenbuah
yang masak clan sakitsecam
teratur,dan penggunaan fungisida pada waktu musim penghujan. Namun intensitas serangan
patogen masih tetap tinggi sampai sekarang. Kondisi ini tejadi diduga karena P.
palmivora yang ada sekarang menjadi lebih virulen akibat stimdasi penggunaan
kultivar-kultivar resisten yang tidak berdasarkan epidemiologikal penyakit. Menurut
Erwin dan Ri beiro ( I 996), Phytophthora bersifat dip1 oid, dalam keadaan diploid Phytophthora dapat mem bentuk progeni-progeni baru yang mempakan sumber
keragarnan baru yang lebi h vi rulen. Me kani sme variabil i tas lain pada cendawan dapat
terjadi karena mutasi, rekombinasi, heterokariosis, paraseksual isme, heteroploidi dan
pembentukan sektor pada koloni (Agrios 1997). Oleh karena itu, perlu dipelajari
apakah ada perbedaan tingkat virulensi
P.
pufmivoru pada suatu lokasi dengan lokasilainnya melalui uji virulensi isolai-isolat P. palmivuru dari beberapa lokasi pada
berbagai kultivar kakao yang berbeda tingkat ketahanannya, dan juga melakukan
analisis keragarnan genetik dan kekerabatan antar isolat untuk mengetahui
Virulensi rnempakm penanda yang paling banyak digunakan dalam andisis
keragaman geneti
k,
karena m e r u p h n si fat patogen yang paling penting dalamhubungan patogen-inang, meskipun hasilnya sangat dipengaruhi lingkungan. Metode
RAPD (Randomly AmpI@ed Polymorphic DNA) merupakan teknik analisis penanda
genetik yang sangat popular dan banyak digunakan untuk menganalisis keragaman
geneti k patogen, karena prosedumya lebih sederhana dan relatif lebi h cepat walaupun
hasilnya kurang konsisten. Namun, akhir-akhir ini telah digunakan penanda genetik
AFLP (Ampl~fied Fragment Length Polymorphism). Dengan teknik ini, akan didapat
inforrnasi yang lebih baik dan akurat untuk menganalisis tingkat keragaman genetik
dan kekerabatan suatu organisme dm hasilnya sangat konsisten (Vos et al. 1995).
Tujuan Penelitian
Berdasarkan pada uraian pernasalahan di atas, dilakukan penelitian yang
bertujuan untuk ( 1 ) menganalisis intensitas penyakit busuk buah kakao akibat infeksi
P. pulmivt~m yang terjadi secara aIami pada perkebunan kakao di Lampung Selatan,
Cianjur dan Jember, (2) melakukan identi
fi
kasi species Phytofllhoru yangmenyerang tanaman kaho di Indonesia, ( 3 ) rnelakukan evaluasi tingkat virulensi
beberapa isolat 1'. pulmivcrm dan resistensi beberapa klon kakao serta (4) melakukan
analisis keragaman genetik beberapa isolat I'. pu~mivora yang dikumpulkan dari
5
Eipoiesis
Dalam penelitian ini hipotesis yang diajukan adalah ( 1 ) intensitas penyakit busuk buah
kakao
akibat infeksiP.
palrnivora yang terjadi secara alami padaperkebunan kakao di Lampung Selatan, Cianjur dan Jember berbeda, (2) penyebab
penyakit busuk buah pada tanaman kakao di Indonesia disebabkan oleh beberapa
species Phytophrhoru, (3) virulensi isolat-isolat P. paltnivora dan ketahanan klon-
klon
kakao
berbeda dan (4) isolat-isolat P. palmivoru mempuny ai tingkat keragarnangenetik yang tjnggi.
Kegunaan Penelitian
Adapun kegunaan dari penel i tian ini dapat mem beri kan in fomasi yang
berharga &lam menyusun pengelolaan pen yakit busuk buah yang disebabkan oleh
cendawan P. pulmivoru pada perkebunan kakao di Indonesia berdasarkan spesifik
LANGKAH-LANGKAH
DAN
KEGUNAAN PENELITIAN
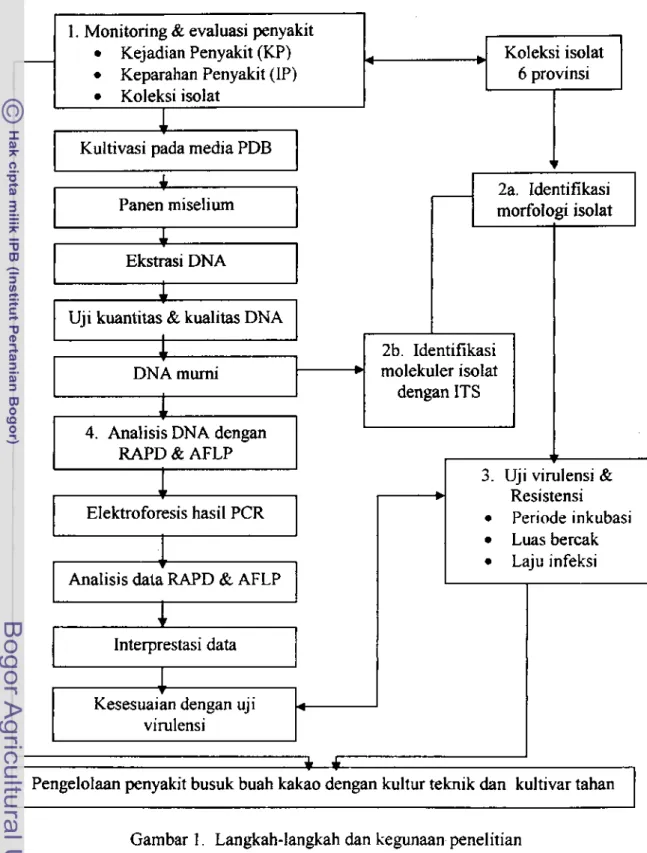
Gambar I . Langkah-langkah dan kegunaan penelitian 1. Monitoring & eval uasi pen yakit
KejadianPenyakit(KP) Keparahan Penyakit (IP)
Koleksi isolat
4 Koleksi isolat
6 provinsi
1
Kuitivasi pada media PDB
7
jl
Panen miseliurn morfologi isolat 2a, ldentifikasi
I
4
Ekstrasi DNA
6
Uji kuantitas & kualitas DNA
I
DNA mumi
&
2b. Identifikasi molekuler isolat
dengan
ITS
4. Anal i si s DNA dengan RAPD & AFLP
3. Uji virulensi &
Resi stensi
Periode inkubasi Luas bercak Laju infeksi
1
EIektroforesis hasil
PCR
J.
Analisis data RAPD & AFLP
+
i
Interprestasi data
I
TKesesuai an dengan uj i virulensi
4
f
TINJAUAN PUSTAKA
Phytophthora palmivora (Butler) Butler (1 9 19)
Sinonim Pl~ytophthoru pulnrivoru meliputi
P.
omnivom de Bary ( I 88 1 ),Pyflzium pultnivorum Butler ( 1 9071,
P.
fuberi Maublanc ( 1 9091, P. fheobromut.Coleman (1 9 101, Kuwukumiu curicu b r a (1 9 1 5 ) , P. Jici Hori ( 19 1 51, P. curica Hara
( 1 9 1 6), P. paImivora var. pperis Muller ( 1936), clan
P.
palmivora var. tl~eobrclmuc. (Coleman) Orellana (1959) (Holliday 1980; Erwin dan Ribeiro 1996). Dalamklasifikasi, P. pulmivom terrnasuk FamiIi Pythiaceae, Orde Peronosporales, KeIas
Oomycetes, Phylum Oomycota dm Kingdom Chromista (Hawksworth et al. 1995;
Alexopoulos e i al. 1 996).
Cendawan P. pulmivom &pat tumbuh baik pada media standar seperti
cornmeal agar, carrot ugur, lima bean ugur, dan V8 juice ugur pada suhu optimum
27-28°C (Drenth dan Sendall 200 1 ). Miselia turnbuh interseluler clan membentuk
haustoria di dalam sel tanaman inangnya. MiseIia benvarna putih, tidak bersekat dan
tumbuh baik pada 25-30°C (Siturnorang 1 983). Menurut Milndkur (1 96 1 ) cendawan ini mernbentuk miselia yang bercabang clan tidak bersekat ketika muda, dan
membentuk sekat pada hifa yang sudah tua yaitu pada saat pembentukan organ
reprodukti f.
Pada buah
kakao
cendaivan dapat membentuk sporangia (zoosporangia),berbentuk buah per, dengan ukuran 30-60 x 20-53 pm. Sporangia dapat berkecambah
8
berkecambah secara
tidak
langsung dengan rnembentuk zoospora atau spom kern barayang dapat berenang. Cendawan dapat membentuk klamidospora yang bulat, dengan
garis tengah 3040 pm (Erwin dm Ribeiro 1996).
Cendawan P. palmivoru bersifat heterotalik,
membentuk
oogonia dan anterediasecara alami atau buatan apabila strain-strain yang mernpunyai tipe kawin A ? dan A2
berpasangan dan pe teburan kedua tipe kawin tersebut menghasi 1 kan oospora (Brassier
dan Maddison 198 1). Dewasa ini di seluruh dunia dikenal adanya dua tipe kawin P.
pulmivora yaitu A1 dan A2. Di Jawa dikenal tipe kawin A2 yang berasal dari
tanaman inang lain bukan kakao (Cutlelya sp, dan Vanda sp.) (Zentmyer 1974).
Secara in vitro, oospora dibentuk pada
suhu
rendah f20°C) dalam keadaan gelap clannutn'si yang sesuai. Pada keadaan alami, oospora dibentuk pa& jaringan berkayu atau
sisa-sisa tanaman yang terhindar dari cahaya. Oogonia buiat (21-40' pm), dibentuk
secara lateral atau terminal, berdinding tipis dan tidak berwarna waktu masih muda.
Sebaliknya jika su&h matang, oogonia akan berdinding tebal dan b e m a coklat
keemasan. Anteridia am figenus dan persisten, 1 0- 1 5 x 1 0- 1 7 y m juga dibentuk secara
lateral atau terminal, berdinding tipis dan tidak benvarna. Oospora berbentuk bulat,
berdinding tipis atau tebal, dengan garis tengah 16-30 pm tidak berwarna pada waktu
muda tetapi akan krwama kuning hingga coklat keemasan apabila telah matang
(Erwin dan Ribeiro 1996).
Di beberapa negara penghasil kakao
di
Afrika Barat diketahui adanya spesiesPhyfophihoru lain yang dapat menyebabkan penyakit busuk buah, yaitu P. megakurycl
I0
pengaruhnya terjadi secara tidak langsung melalui kebasahan permukaan buah
dan
kelembaban. P e w suhu terjadi S a r a tidak langsung melalui kelembaban nisbi
udara dan kebasahan permukaan buah. Makin rendah
suh
y rnakin tinggi kelembabannisbi udara dan makin lama pula bertahannya kebasahan pada permukaan buah.
PeIepasan clan perkecambahan zoospora terjah pada
suhu
15 sampai dengan 30°C,sedangkan infeksi @a buah kakao terjadi pada
suhu
20 sampai dengan 30°C.PeIepasan, perkecambahan, dan infeksi memerl
ukan
adany a air bebas minimumselama 3-4 jam (Purwantara I 990).
Teknik
budidaya tanaman, antara lain pemangkasan, kerapatan tanaman,pemberian mu1 sa, drainasi, pemupukan, dan pernungutan hasil sangat rnempengaruhi
perkembangan penyakit. Lapisan mulsa yang ada di sekitar pangkal batang akan
mencegah terjadinya percikan air yang membawa tanah yang terinfestasi cendawan.
Juga adanya mulsa ini akan meningkatkan kegiatan jasad-j asad renik saprofi t yang
bersifat antagonisti k terhadap 1'. pulmivora. Ada beberapa cendawan antagonis
terhada p
P.
pultnivuru y ait u Suillus luteus, /,uciuriu,s delic,to.~us, /,encopwillu.vcerealis var picein, Asprgi//u.s iumari, A. gigunteur., Penicillium purpurecens dan
Botryodiplodiu ~laeohromue. Di samping cendawan diketahui juga bakteri yang dapat
mengham bat perkem bangan 1'. pulmivom yai tu Pseudomonas uerzrginosu, Buc~llzrs
cereus dan B. subfrli.~ (Sukarnto 1995 ). Busuk buah Iebi h banyak terdapat pada pohon
yang le bat buahnya. Sering di katakan penyaki t busuk buah berbanding 1 urus dengan
1 1
Kakao dari kelompok CrioIlo sangat rentan terhadap
busuk
buah. KeIornpokForastero mempunyai ketahanan yang lebi h tinggi. Trinitario, merupakan hi brida dari
Criollo dan Forastero mempunyai ketahanan yang bervariasi. Sukarnto dan Mawardi
(1986) melaporkan bahwa klon DRI rentan; klon DR2, DR38
dan
GC7 agak rentan;klon
DRC9
dan SCA89 agak tahan; sedangkan klon SCA6, SCAl2, ICS6 danDRC
16tahan terhadap serangan P. paIm~vora penyebab penyakit busuk buah kakao.
Viru lensi Phytophtlrora pulmrmrvora
Virulensi adalah tingkat kemampuan suatu patogen untuk menimbulkan
pen yakit atau tingkat patogenisi tas dari suatu patogen. Sedangkan patogeni sitas
berarti kemampuan relati f dari suatu patogen unhk menimbulkan penyakit (Talboy s et
al. 1973; Shurtleff dan Averre I11 1997). Isolat
P.
palmivnru dari berbagai bagiantanaman kakao dan dari tanah rnampu menimbulkan penyakit busuk buah dan kanker
batang pada kakao. Virulensi isolat P. pulmivoru yang diambiI dari berbagai bagian
tanaman kakao dari dua kebun di Jawa Barat tidak berbeda. Virulensi tidak dapat
digunakan sebagai penanda untuk menentukan keragaman genetik P. pulmivoru
(Purwantara 200 1 ).
Motulo (2000) melaporkan bahwa isolat-isolat P. pu/mivuru yang berasosiasi
dengan penyakit gugur buah pada tanaman kelapa mempunvai keragaman genetik
yang tinggi. Isolat yang berasaI dari lokasi yang sama tidak selalu menunjukkan
juga tidak selalu menunjukkan hubungan kekerabatan yang jauh. Patogenisitas isolat-
isolat P. palmivora pada setiap kdtivar kelapa berbeda.
Hasil penelitian Kasim dan Prayitno (1980) membuktikan bahwa isoIat-isolat
P. cupsici (sebelurnnya di kenal sebagai P. pulmivora) penyebab penyakit busuk
pangkal batang pa& tanaman lada di Lampung Utara, Larnpung Selatan, dan Bangka
berbeda-beda dalam morfologi dan virulensinya. Isolat dari Lampung, khususnya
Lampung Selatan mempunyai virulensi yang lebih tinggi dari pada isoIat dari Bangka.
Beberapa isolai P. pulmivora yang berasal dari kakao mernpunyai tingkat
patogenisitas yang tinggi pada buah kabo bila dibandingkan dengan isolat asal
kelapa, lada dan vani ti. Hal ini dibuktikan dengan perkembangan diameter bercak
pada permukaan kulit buah dan jaringan di b a d permukaan kulit buah kakao
(Hendrawati 1 997). Ruzelfin ( 1989) j uga melaporkan bahwa cendawan
P.
puimivoraasztl busuk buah kakao, busuk bidang sadapan karet, dan gugur buah kelapa hibrida PB
12 1 menyebabkan gejala bercak pada kulit buah kakao dengan patogenisitas isoIat asal
kakao lebih tinggi dibandingkan isolat asal karet dan kelapa, sedangkan isolat asal lada tidak rnenunjukkan sifat patogenisitas (tidak mampu menimbulkan gejala) pa& buah
kakao. Dengan demi kian tanaman karet dan kelapa hibrida PB 1 2 1 yang terserang P.
Mekanisme Keragaman pada Cendawan
Pada cendawan ras-ras b m dapat terjadi meldui beberapa proses, yaitu
mutasi, hibridisasi seksual, heterokariosis, rekombinasi paraseksual dan adaptasi
(Brown 1980; Semangun I 996; Agrios 1997).
Mutasi adaIah perubahan secara mendadak pada sifat genetik yang te rjadi
karena adanya perubahan pada satu basa atau Iebih pada untaian nukleotida. Pada
kebanyakan mutasi, perubahan terjadi pada gen individual, meskipun kadang-kadang
juga karena a&nya perubahan pada krornosom DNA (Agrios 1997). Frekuensi
mutasi dapat meningkat karena adanya agensia fisik maupun kirniawi.
Hibridisasi s e k s d terjadi karena rekombinasi gen sebagai akibat percampuran
secara acak kromosom cendawan i nduk dan j uga karena terjadi nya crossing-over
selama meiosis. Sebagai akibatnya genotipe koloni keturunannya berbeda dengan
genotipe induknya. Diperkirakan bahwa banyak ras baru cendawan patogen timbul
karena hibridisasi seksual ini (Brown 1 980; Agrios 1 997).
Heterokariosis terjadi karena sel cendawan mengandung dm atau Iebi h inti
yang berbeda secara genetik. Keadaan heterokariotik ini dapat terjadi karena beberapa
cara anbra lain ( I ) terjadinya fusi atau anastomosis hifa-hifa yang rnenyebabkan
tercarnpurnya inti yang berbeda ke dalam suatil miselium, dan (2) te jadinya mutasi
pada hi fa homokariotik karena adanya inti mutan yang dapat bertahan di antara inti-
inti lama. Diperkirakan bahwa heterokariosis merupakan mekanisme variasi yang
penting pa& cendawan Phyfophfhora karena fase di kariot ik merupakan bagian besar
Rekombinasi parase ksual terjadi b i h gen-gen rnengadakan rekombinasi
di
Iuardaur seksual. Rincian daur ini belum banyak diketahui, namun agaknya daur ini
melewati I ima langkah penting: ( I ) pembentukan
rniseliurn
hetero karioti k clante j adinya fusi antara dua inti haploid yang berbeda untuk menjadi inti diploid; (2) Inti
dpIoid ini harm dapat memperbanyak diri bersama-sama dengan inti haploid
induknya. Dengan demikian miselium mengandung tiga tipe inti, dua inti haploid
yang berbeda dan inti diploid; (3) Berkembangnya miseli~m diploid yang
menghasil kan strain cendawan diploid; (4) Terjadmya crossing-over mitoti k yang
menyebabkan terjadmya rekombinasi gen pada kromosom ddam inti diploid; dan (5)
Terjadtnya haploidisasi vegetatif, kromosom yang mengandung gen rekombinasi
terdapat dalam inti haploid (Semmgun 1996; Agrios 1997).
Adaptasi sebagai suatu m e h i s m e keragaman pada cendawan, walaupun agak
sul i t di buktikan &lam percobam. Beberapa cendawan patogen dapat beradaptasi
terhadap lingkungan tertentu karena memproduksi enzim-enzim adaptif. Adaptasi
nampaknya merupakan h a i l seleksi dari strain-strain mutan yang mempunyai
keuntungan kompetitif untuk keragaman (Brown 1980).
Marker Molekuler
Potensi penggunaan marker sebagai dat untuk rnelakukan karakterisasi genetik
dalam program pemuliaan telah dikenal sejak puluhan tahun yang lalu. Marker bisa
dikatagorikan sebagai marker morfologi, sitologi, dan yang terbaru adalah marker
tanaman, virulensi merupakan marker yang paling
banyak
digunakan(Puwan&ra
2001). .
Marker
rnorfologi merupakan marker yang telah banyak digunakan, baikda1am program mendasar geneti ka maupun dalam program praktis pemuliaan tanarnan
karena marker ini dapat dengan mudah diarnati, seperti warna bunga, warm batang,
warna kulit biji, bentuk biji, clan sebagainya. Narnun marker ini rnemililu kelernahan
karena &pat dipengaruhi oleh kondisi lingkungan, memperlihatkan sifat menurun
dominankesesi f, dm mempunyai tingkat keragaman (polimorfisme) rendah atau
jumlah yang sedikit (Tanksley et a/. 1989). Marker sitolog adalah marker yang
berhubunp dengan kromosom. Contoh marker sitologi yang telah dipergmakcin
di
daIam membantu pernul iaan tanaman adalah jurnlah kromosom, ukuran kromosom
dan morfologi set kromosorn (Sessions 1996).
Pada saat ini, kemajuan ddam bidang biologi berkembang sangat pesat.
Biologi molekuler merupakan salah satu cabang ilmu yang mernpelajari organisme
pada tingkat DNA. Teknik di bidang biologi molekuler sangat membantu pemulia
tanaman daiam melakukan studi genetik dengan ketepatan yang lebih akurat. Untuk
mendapatkan informasi genetik dapat dilakukan analisis dengan menggunetkan marker
molekuler, se perti isozym, RFLP (restriction fragment length po(vmorphism), RAPD
(randomly amplfled pobmorphic DNA), AFLP (ampdifled fragment length
polymorphism), dan yang Iainnya (Kongluatngam et 02. 1 995; Powell et 01. 1996; Karp
Marker rnolekuler &pat rnemberi gambaran yang cukup t inggi tentang
perbedaan genetik individu, baik pada tingkat spesies maupun dengan kerabat
jauhnya. Menurut Tanskley (1983) marker molekuler dapat mendeteksi variasi
genetik pada tingkat jaringan atau seluler, dm polimorfisrnenya tidak dipengaruhi oleh
lingkungan. Marker molekuler yang pertam dan yang paling sederhana dikenal
dengan penanda protein yang lazim disebut isozyrn. Polimorfisme protein dideteksi
dengan cara eIektroforesis, dan perbedaan yang terdeteksi
antar
ale1 bergantungpada
pergantian asam-asam amino yang berrnuatan. Untuk pencirian dan analisis gen yang
j umIahnya beragam, aplikasi marker isozym
mem
punyai keterbatasan karenaj
umlahlokus yang bisa digunakan terbatas (Murphy el al. 1996 j.
Perkembangan dan penggunaan marker RFLP relati
f
baru dibandingkandengan isozim, meski pun demi kian prinsip interpretasi anal isis genetik dari isozyrn
sama dengan RFLP. Akhir-akhir ini RFLP mendapat perhatian yang Iebih besar dari
pakar genetika molekuler dart pemulia tanaman. Hal ini karena RFLP dapat
mengungkapkan perbedaan-perbedaan yang lebih banyak antar individu-individu
di bandingkan i s o w . Marker RFLP mendasarkan pada perbedaan &lam ukuran
fragmen DNA nukleus (kromosom), organel, atau total DNA yang dihasilkan dari
pernotongan dengan enzim restri ksi. Individu-individu yang mempun yai perbedaan
sekuen DNA akan mempunyai perbedaan distribusi dan situs restriksi untuk suatu
enzim restriksi. Fragmen DNA hasil restriksi dipimhkan menurut ukuran berat
molekul pada gel elektroforesis, kernudian dipindahkan ke mernbran niIon dan
17
DNA yang terbentuk p d a membran nilon yang dihibridisasi dengan suatu probe berbeda antar individu yang diuji (McCouch dan Tanskley 199 1).
Semenjak diperkenalkm oleh Williams et al. (1990), teknik RAPD menjadi
salah satu cara yang banyak digunakan untuk krbagai penelitian di bidang biologi
molekuler. Dibandingkan dengan RFLP, teknik ini lebih sederhana karena DNA tidak
perlu lpotong dengan enzim resbiksi, sampei DNA yang diperIukan relatif sedikit,
tidak
memerlukan pemindahan DNAke
membran nilon,tidak memerlukan
hibridisasiDNA,
clan
ti& memerlukan prosedur labeling. Telmik RAPD mendasarkan padaamplifikasi DNA secara in vitro dengan PCR (polymerase chain reaction), yaitu
dengan rnengatur variasi
suhu
pada mesin PCR selarna pengulangan siklus
denaturasi,pertautan primer, dan perpanjangan pita DNA dengan bantuan enzirn Tag DNA
polirnerase. Teknik ini memerlukan primer yang panjangnya 10 basa untuk segrnen
pem ula dalam pem bentukan fragmen tertentu dari DNA (Nair 1 993 ).
Di bandingkan dengan te kni
k
RFLP,
RAPD mempun yai beberapa kemudahanyaitu; pengetahuan latar klakang genom ti& diperlukan, hasil RAPD dapat
diperoleh secara cepat ji ka dibandingkan dengan analisis RFLP yang memerlukan
banyak tahapan
dan
beberapa jenis primer arbitrari dapat dibeli dan digunakan untukanalisis genom semua jenis organisme, sedangkan keterbatasannya adalah sangat
sensitif terhadap kondisi reaksi dan profil suhu (Williams el ul. 1990; Vos er a/. 1995).
Di samping itu marker RAPD bersifat dominan, yaitu dalam populasi yang
bersegregasi, individu yhqg homozigot dengan individu yang heterozigot tidak dapat
18
atau yang heterozigot akan sama-sama memberikan hasil pita DNA untuk suatu
marker RAPD tertentu (Ronning et al. 1 995).
Teknik AFLP merupakan penggabungan dari RFLP clan RAPD, yang
mendasarkan pada amplifikasi PCR selektif fragmen restriksi dari pernotongan total
DNA genomik. Teknik ini meliputi tiga tahapan yaitu restriksi DNA dan ligasi
adapter oligonukleoti& amplifikasi selektif set fragmen restriksi dan malisis gel dari
fragmen restriksi. Dengan rnenggunakan tekni k ini informasi geneti
k
yang didapatkanlebih akurat, narnun pelaksanaannya lebih sulit dan memerlukan biaya yang tinggi
EVALUASI PENYAKlT BUSUK BUAH KAKAO
PADA
BERBAGAI PENGELOLAAN
KEBUN
DI LAMPUNG
SELATAN,
CIANJUR
DAN
JE-MBER
Pendahuluan
Busuk buah kakao adalah penyakit yang tcrpenting dalam budidaya kakao di
Indonesia. Penyakit ini &pat timbul pada berbagai umur buah, sejak buah masih kecil
sampai rnenjelang masak. Warna buah berubah, umumnya mulai dari dekat tangkai
atau ujung
buah,
yang dengan cepat meluas ke seluruh buah clan akhirnya buahmenjadi coklat kehitaman. Pada bercak tersebut terbentuk banyak sporangiofor dan
sporangia cendawan
P.
plmivora penyebab penyakit ini. Jika buah yang terserang P.puimivora tidak segera diptik, cendawan ini akan berkembang melalui tangkai
buab
dart menginfeksi kulit batang atau cabang menirnbulkan kanker batang. Kelak,
patogen yang metlimbul kan kan ker batang dapat kembali mengi nfeksi buah melalui
tangkai buah atau secara tidak langsung melalui spora yang dihasil kan (Semangun
2000).
Keparahan penyakit busuk buah kakao pada suatu lokasi atau kebun ditentukan
oleh banyak faktor, antara !ain kelembaban udara. curah hujan, cara bercocok tanam,
banyaknya buah pada pohon dan kultivarklon kakao yang ditanam. Kelembaban
yang tinggi akan membantu pembentukan spora dan meningkatkan infeksi. Infeksi
hanya dapat terjadi kalau pada permukaan buah terdapat air, baik yang berasal dari air
hujan,
dan
dapat j uga air yang terbentuk karena pengem bunan uapair
pada pemukaanpermukaan buah merupakan syarat penting untuk perkembangan penyakit, mengingat
ketersediaan air adalah salah satu kondisi yang diperlukan untuk perkecambahan spora
dan proses infeksi pada
b d .
Hujan akan membantu penyebaran spora, di sampingmeningkatkan
kelernhaban
kebun (Purwantara 1 990). Fluktuasi keparahan penyakitcenderung sama dengan fluktuasi curah hujan (Purwantara dan Pawirosoemardjo
1990).
Cam
b e m k
tanam, antara lain pemangkasan, kerapatan tsnaman, pem berianmulsa, drainase, pemupukan dan pernungutan hasiI sangat mempengasuhi penyakit.
Lapisan mulsa yang ada di sekitar pangkal batang &an rnencegah te rjadinya perci kan
air yang membawa tanah yang terinfestasi cendawan. Juga adanya mulsa ini akan
meningkatkan kegiatan jasad-jasad renik saprofit yang bersifat antagonistik terhadap
P.
plmivoru. Busuk buah lebih banyak terdapat pada pohon yang lebat buahnya(Semangun 2000).
Di Indonesia terdapat tiga kelompok bahan tanaman kakao, yaitu kelompok
Trin i tario (klon-klon
DR),
kelornpok Amelonado (West African Amelonado), dankelompok Amazon (Ilpper Amazon HybridICrAH). Trini tario rnenghasil kan kakao
mulia, sedang Amelonado dan Amazon rnenghasilkan kakao iindak (Wardojo 1991).
Dari ketiga kelornpok bahan tanaman kakao tersebut, kakao mulia rnempunyai
ketahanan yang lebi f? rendah terhadap infeksi
Y.
palmivorudi
bandingkan dengankakao lindak (Semangun 1 987).
Berdasarkan pernasalahan yang ada di Indonesia secara umum maka
2 1
buah kakao akibat infeksi
P.
palrnivora yang te jadi secara alami p d a perkebunankakao rakyat di Lampung Selatan, perkebunan kakao swasta di Cianjur dan
perkebunan kakao negara di Jemkr. Hipotesis yang diaj ukan ddarn penelitian ini
adalah intensitas penyaki t busuk buah kakao akibat infeksi
P.
palrnivora yang te jadisecara alarni pada perkebunan kakao rakyat di Lampung Selatan, perkebunan kakao
swasta di Cianjur dan perkebunan kakao negara di Jember berbeda. Hipotesis ini
didasarkan pa& adanya perbedam pen& kondisi lingkungan kebun, kultivar/klon
kakao
ymg ditanam dan pengelolaan penyalut yang diteraph oleh ptam clanpekebun baik pada perkebunan kakao rakyat, perkebunan
kakao
swasta rnaupun padaperkebunan kakao negara.
Bahan dan Metode
Penelitian ini dilaksanakan di tiga Iokasi yaitu
di
kebun-kebun kakao milikrakyat di Desa Negeri Sakti Kecamatan Gedong Tataan Kabupaten Lampung Selatan,
Lampung; perkebunan swasta nasional milik PT Inter Green Estate, Cianjur, Jawa
Barat dan perkebunan negara milik PTP Nusantara XII, Jember, Jawa Timur, dari
bulan Pebruari sampai bulan Juni tahun 2002. Bahan-bahan y ang digunakan daIam penelitian ini berupa tanaman dan buah kakao rnulia (klon DRl, DR2, DR 38) dan
tanaman dan buah kakao lindak (Upper Amuzon Hybrid (UAH)/Hibrida Amazon
Hulu) yang ada pada kebun-kebun contoh.
Penilaian penyakit diIaksanakan dengan metode survei, untuk mendapatkan
22
Pacia kebun-kebun kakao yang dij adi kan lokasi penelitian
di
buat petak-petakpengamatan secara diagonal sebanyak 1 i
ma
petak. Masing-masing petak berj umlah100 tanaman kakao dengan tanaman sampel sebanyak 10 tanaman per petak atau 50
tanaman sampel per Iokasi penef itian yang ditentukan secara purposive dan masing-
masing diulang tiga kali.
Kerusakan aki bat penyaki t busuk buah diniIai berdasarkan kejadian penyakit
dan keparahan penyakit yang terjadi secara dami. Data lainnya yang mendukung
penelitian ini di kum pulkan rnelalui wawancara dengan 1 0 petanupekebun
menggunakan daftar pertanyaan (Lam piran I ). Kejadian penyaki t di hitung menggunakan forrnul a yai t u :
Keterangan :
KP = Kejadian penyakit (%)
n = Jumlah pohon yang menunjukkan gcjala busuk buah
N
= Jumlah pohon yang diamatiKeparahan penyakit dihitung dengan menggunakan formuIa Townsend dan
Keterangan :
IP
= Keparahan penyakit (%)ni = Jurnlah po hon yang menunj
ukkan
katagori seranganvi = Nilai numerik dari setiap katagori
Z = Nilai numerik katagori serangan tertinggi
N
= Jumlah pohon yang diamatiPohon-pohon yang dijadikan sampel dibagi dalam enam katagori penilaian
seperti disajikan pada Tabel 1.
Tabel 1 Katagori penilaian keparahan penyakit
(IP)
busuk buah kakao--
Katagori Kisaran serangana'
0 X = 0%
1 0 < X I 2,5%
2 2,5
< X l
5 %3 5
< X I
7,5 %4 7,5 < X 5 1 0 %
5
X >
10%Pengamstan Gejala Penyakit Busuk Buah Kakao
Hasil pengamatan gejala penyakit busuk buah dan kanker batang yang tejadi
secara alami dapat dilihat pada Garnbar 2. BiIa buah-buah kakao sehat (A) yang terin feksi P. pulmivora menj adi sakit dengan menunj ukkan perubahan warm menjadi
coklat kehitaman, urnumnya gejala banyak terdapat mulsi dari dekat tangkai buah
(proksimd)
(B)
atau dari ujung buah (distal) (C) dan ada juga yang dimulai dari bagian tengah buah (lateral) (D). Pada permukaan Mit buah kakao yang sakitterdapat tanda penyakit berupa lapisan tepung b e m a putih
(E).
ApabiIa buahkakao
sakit dibiarkan masi h tergantung di pohon dengan tangkai buah yang telahbusuk, umumnya penyakit sudah berkembang meluas ke kulit batang atau cabang
menyebabkan gejala kanker
(F).
Buah-buah kakao sakit yang masih tergantung dipohon merupakm sumber inokulum penyakit busuk buah
kakao.
Penilaian Penyakit Busuk Bush Kakao di h m p u n g Selatan
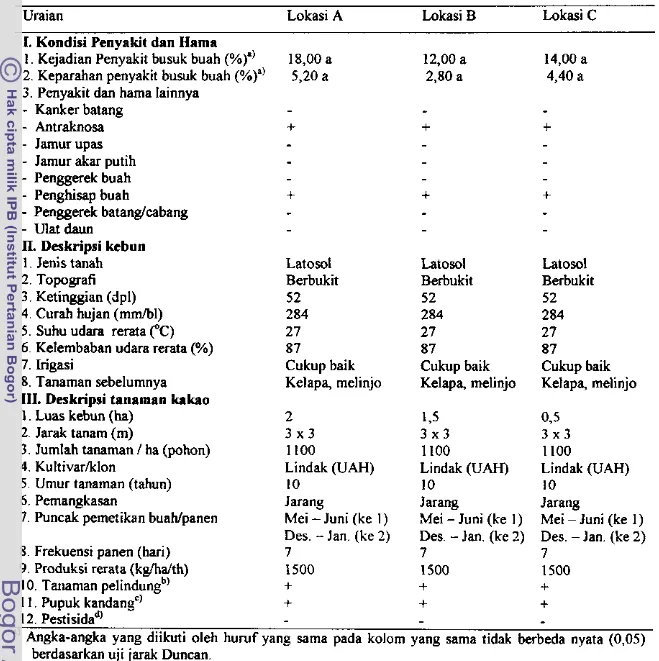
Rerata kejadian penyakit dan keparahan penyakit busuk buah pada tiga lokasi
kebun kakao rakyat di Lampung Selatan tidak bertKda nyata (Tabel 2). Hasil analisis
ragam kejadian penyaki t dan keparahan penyakit busuk buah disaj ikan pada Lam piran
P e ~ e l b m m
mnaman
k a h
dm@ -&WD j m gjuga
dengan
&naman pdimbg, pada l o b Aj d a b
tanmanp e b b g
termtam
kelapa
lebih
banyak
&ban- pa& l o h iB dan
C.T a m a m
pisangd i g u d m
sebaga~tunpang
sari
cEan
pelindung
pgds waldam
mih
mu&.
?:maman
k a hymg
ditanam
addah t i p W l W &(UAH) y m g
bsrssal
dpriM&,
S~~~~~bsntukbiii
hthlo.
Pada
mnur
I0 Mtm,ptodddvitasnya Eulcup tinggi yaitu 1500
k@haM~
(Tabel
2).Umumnya
peagetah- p b m m a p m i penyakit
bmuk buah,
p y a k tdan
hama h y a ymgTabel 2. Evaluasi kondisi penyalut, deskripsi kebun dan deskripsi tanaman kakao perkebunan rakyat
di
Lampung SefatanUraian Lokasi A Lokasi 3 Lokasi C
I. Kondisi Penyakit dan Harna
1 . Kejadian Penyakit busuk buah (%)') 18,OO a 12,00 a 14,00 a
2. Keparahan penyakit busuk buah (%)" 5,20 a 2,80 a 4,40 a
3. Penyakit dan hama Iainnya
-
Kanker batang- Antraknosa I-
+
+
- Jamur upas - Jamur akar putih
- Penggwek buah
- Penghisap buah
+
+
+
-
Penggerek batangfcabang- Ulat daun
II. Deskripsi ktbun
1. Jenis tanah Latosot Latosol Latosol
2. Topografi Berbukit Berbukit Berbukit
3 . Ketinggian (dpl) 52 52 52
4 . Curah hujan (mm/bl) 284 284 284
5 . Suhu udara rerata CC) 27 27 27
6. Kelernbaban udara rerata (%) 87 87 87
7. Irigasi Cukup baik Cukup baik Cukup baik
8. Tanarnan sebelurnnya Kelapa, melinjo Kelapa, melinjo Kelapa, melinjo
111. Deskripsi tanaman kakao
1. Luas kebun (ha) 2 1,s O,5
2. Jarak tanam (m) 3 x 3 3 x 3 3 x 3
3. Jumlah tanaman