MANIPULASI KONDISI FISIOLOGIS DAN KEASAMAN SEMEN
MELALUI PENGATURAN PERBEDAAN KATION ANION
RANSUM DALAM UPAYA MEMPENGARUHI
KARAKTERISTIK SPERMATOZOA
PADA DOMBA GARUT
RAHMAT HIDAYAT
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI
Dengan ini saya menyatakan dengan sebenar-benarnya bahwa disertasi saya dengan judul: Manipulasi Kondisi Fisiologis dan Keasaman Semen Melalui Pengaturan Perbedaan Kation Anion Ransum dalam Upaya Mempengaruhi Karakteristik Spermatozoa pada Domba Garut adalah karya saya sendiri dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Mei 2009
ABSTRACT
RAHMAT HIDAYAT. Altering Physiological Conditions and Semen Acidity by Manipulating Dietary Cation-Anion Difference and Their Effect on Sperm Characteristics of Garut Breed Rams. Under the supervision of TOTO TOHARMAT, ARIEF BOEDIONO and IDAT GALIH PERMANA.
This study was carried out to evaluate the effect of dietary cation-anion difference (DCAD) and fish oil supplementation on dry matter intake, dry matter and organic matter digestibility, weight gain, acidity of urine, blood and semen, blood gas, plasma mineral, sperm characteristics and ratio of X:Y spermatozoa. The dietary treatmens were as follows: RN0= basal ration (DCAD +14) without fish oil, RNI= basal ration (DCAD +14) with 3% fish oil, RB0= base ration (DCAD +40) without fish oil, RBI= base ration (DCAD +40) with 3% fish oil, RA0= acid ration (DCAD -40) without fish oil, and RAI= acid ration (DCAD -40) with 3% fish oil. The rations contained 150 ppm of zinc and were offered to 18 of Garut breeed rams. The result indicated that: DCAD +40 and -40 decreased dry matter intake significantly. The ration with DCAD +40 had the lowest dry matter intake. No differences were observed for dry matter and organic matter digestibility. DCAD +40 and -40 decreased body weight of rams except RA0 ration, however, the body weight of rams was very low. Variation of urine pH followed the DCAD pattern. Blood pH, pCO2, and pO2 were not affected by DCAD, but cHCO3 and cBase were affected. DCAD affected plasma Mg and S, but did not affect K, Na, Zn and Cl. Semen pH after day 28 of experimental period were significantly different and that followed DCAD pattern. There were not differences for X:Y ratio, macroscopic and microscpic characteristic of sperm.
RINGKASAN
RAHMAT HIDAYAT. Manipulasi Kondisi Fisiologis dan Keasaman Semen Melalui Pengaturan Perbedaan Kation Anion Ransum dalam Upaya Mempengaruhi Karakteristik Spermatozoa pada Domba Garut. Dibimbing oleh TOTO TOHARMAT sebagai ketua, ARIEF BOEDIONO dan IDAT GALIH PERMANA sebagai anggota.
Penelitian ini menggunakan ransum dengan nilai perbedaan kation anion ransum (PKAR) +14, +40 dan -40 meq masing-masing ada yang ditambahkan minyak ikan 3% dan tidak. Tujuan penelitian ini yaitu untuk mengetahui : (1) Pengaruh PKAR dan suplementasi asam lemak esensial dalam ransum berkecukupan Zn terhadap konsumsi pakan, absorpsi dan metabolisme nutrien, metabolit dan kondisi kimia darah. (2) Keterkaitan PKAR suplementasi asam lemak esensial terhadap absorpsi dan metabolisme Zn, Na, K, Cl serta S. (3) Mengkaji pengaruh PKAR dan suplementasi asam lemak esensial dalam ransum berkecukupan Zn terhadap karakteristik spermatozoa dan rasio sperma X:Y.
Penelitian dilaksanakan di Fapet dan FKH IPB pada 9 Mei – 27 Oktober 2007. Ransum perlakuan, yaitu nilai PKAR +14, +40 dan -40 meq masing-masing ada yang ditambahkan minyak ikan 3% dan tidak. Ransum basa diperoleh dengan menambahkan Na2CO3 dan K2CO3 ke dalam ransum basal, masing-masing sebagai sumber kation Na dan K agar nilai perbandingan kation anion ransum (PKAR) menjadi +40. Kelompok ransum asam diperoleh dengan menambahkan MgCl2 dan MgSO4 ke dalam ransum basal masing-masing sebagai sumber kation Cl dan S agar nilai PKAR-nya menjadi -40. Ransum tersebut dianalis kandungan Na, K, Cl, dan S total, dilanjutkan dengan menghitung besarnya neraca kation-anion berdasarkan persamaan Tucker et al. (1992). Seluruh ransum mengandung Zn 150 mg/kg BK ransum dengan cara menambahkan ZnSO4. Ransum dicobakan selama 50 hari pada 18 ekor Domba Garut jantan berumur dua tahun yang dua minggu sebelumnya telah diberi ransum basal tanpa suplementasi ZnSO4. Rancangan yang digunakan adalah rancangan acak kelompok (RAK). Kelompok berdasarkan bobot badan, yaitu (I) bobot badan 32.5-37.5 kg, (II) 30.5-31.5 kg dan (III) 28.5-30 kg.
PKAR dan penambahan minyak ikan pada ransum percobaan nyata menurunkan konsumsi BK ransum. Selanjutnya perbandingan antara ransum netral dengan ransum basa menunjukkan bahwa terjadi penurunan konsumsi BK pada ransum dengan nilai PKAR basa jika dibandingkan dengan ransum normal. Hal ini juga terjadi pada konsumsi BK ransum dengan PKAR asam lebih rendah dari pada konsumsi BK ransum normal. Nilai PKAR basa dengan asam, hasilnya memperlihatkan bahwa konsumsi BK ransum basa lebih rendah dari pada ransum asam.
rata-rata pertambahan bobot badan (PBB) tertinggi yaitu 133 gram/hari dengan tingkat konsumsi 871±90 g BK/hari.
Nilai pH urin ternak percobaan pada hari ketujuh sampai hari ke-21 pada PKAR basal dan basa mempunyai nilai yang sama, sedangkan pH urin ternak yang diberi ransum PKAR asam mempunyai nilai yang lebih rendah. Nilai pH tersebut memperlihatkan bahwa kelebihan konsumsi ion negatif yang dikonsumsi ternak percobaan segera diekskresikan melalui urin yang dimanifestasikan melalui menurunnya nilai pH urin.
Hari ke-28-42 penelitian, pemberian ransum PKAR basa tampak jelas pengaruhnya terhadap nilai pH urin. Domba garut yang diberi ransum percobaan PKAR basa menghasilkan nilai pH yang lebih tinggi dibandingkan dengan domba yang diberi ransum basal maupun asam. Hal ini juga terjadi pada hari ke-35 dan 49 percobaan, walaupun pada perlakuan ransum basal dengan suplementasi minyak ikan mempunyai nilai pH yang relatif sama dengan yang diberi ransum basa. Ransum PKAR asam mempunyai pengaruh yang lebih kuat terhadap pH urin jika dibandingkan dengan ransum dengan PKAR basa.
Domba yang mendapat perlakuan PKAR asam tanpa suplementasi lemak mempunyai pH darah 7.33±0.07 tergolong ke dalam katagori pH asam. Domba yang diberi perlakuan ransum dengan nilai PKAR basa tanpa suplementasi lemak mempunyai pH darah tertinggi yaitu 7.44±0.04, walaupun demikian pH ini masih tergolong kategori normal. Sedangkan domba lainnya mempunyai nilai pH darah berkisar antara 7.36±0.08 sampai 7.40±0.02. Menurut Story et al. (2004), pH darah < 7.35 termasuk asam, 7.35-7.45 termasuk normal dan > 7.45 termasuk basa. Sementara itu, Frandson (1992) dan Anstey (2005) menyatakan bahwa nilai pH darah sebesar 7.06 termasuk ke dalam kategori sangat asam, 7.25-7.31 termasuk asam, 7.35-7.40 termasuk normal dan 7.47-7.52 termasuk basa.
Domba yang diberi perlakuan ransum basa tanpa suplementasi lemak mempunyai nilai cHCO3 paling besar, sedangkan yang diberi perlakuan ransum asam tanpa lemak paling kecil. Domba yang diberi perlakuan ransum basa mempunyai nilai cHCO3 yang lebih tinggi daripada domba yang diberi ransum asam maupun basal.
nilai normal yaitu 0.20±1.22–1.87±1.50 mmol/L. Senada dengan Domba Garut yang diberi ransum dengan nilai PKAR basa, suplementasi minyak ikan dalam ransum basal dapat menurunkan nilai cBase darah. Kondisi ini memperlihatkan bahwa suplementasi minyak ikan dalam ransum dengan PKAR basa dapat menurunkan nilai cBase darah.
PKAR dan suplementasi lemak pada ransum tidak mempengaruhi nilai BDM, BDP, PCV dan Hb darah Domba Garut. Suplementasi lemak pada ransum dengan PKAR basal dan basa menurunkan N differensiasi butiran darah putih (BDP), tetapi tidak terjadi dalam ransum dengan PKAR asam. Elemen lain seperti L, M, E dan B tidak terpengaruh oleh PKAR dan suplementasi lemak dalam ransum.
Konsentrasi mineral K+ dan Na+ plasma Domba Garut tidak terpengaruh oleh PKAR dan suplementasi lemak. Hal ini menunjukkan bahwa Domba Garut mampu mempertahankan homeostasi mineral K+ dan Na+ plasma dalam kondisi pemberian pakan dengan PKAR yang berbeda.
Semua ternak dalam penelitian ini mempunyai konsentrasi Na+ lebih tinggi dari K+. Hasil penelitian ini didapatkan perbandingan kation Na+ :K+ untuk RN0. RNI, RB0, RBI, RA0 dan RAI secara berturut-turut adalah 8:1, 7:1, 6:1, 6:1, 7:1 dan 7:1. Imbangan Na+ :K+ dalam tubuh ternak dilakukan untuk mencapai homeostasis.
Konsentrasi mineral Zn dalam plasma darah Domba Garut hasil penelitian berkisar antara 12.99±3.38 sampai 23.81±14.65 ppm. Konsentrasi tertinggi terdapat pada Domba Garut yang mendapat ransum basal tanpa suplementasi minyak ikan. Suplementasi minyak ikan dalam kondisi PKAR basa dapat menurunkan konsentrasi mineral Zn plasma. Sebaliknya, dalam kondisi PKAR asam, suplementasi minyak ikan justru meningkatkan konsentrasi Zn plasma. Walaupun demikian, konsentrasi mineral Zn dalam plasma semua domba penelitian berada dalam kisaran normal. Menurut McDonald et al. (1988), konsentrasi normal Zn dalam tubuh hewan adalah 10–50 ppm.
Konsentrasi mineral Mg pada PKAR asam tanpa suplementasi asam lemak lebih tinggi dibanding dengan ransum perlakuan lain. Hal ini dikarenakan adanya penambahan mineral Mg dalam bentuk MgSO4 dan MgCl2. Kadar mineral Mg yang berlebih dalam pemberian ransum dengan PKAR asam dapat ditekan keberadaanya dengan adanya suplementasi lemak dalam ransum. Terbukti bahwa plasma darah dari domba yang diberi ransum dengan PKAR asam yang disuplementasi lemak mengandung kadar Mg lebih rendah dibandingkan dengan yang tanpa suplementasi.
Domba Garut yang diberi ransum PKAR asam tanpa suplementasi minyak ikan mempunyai konsentrasi ion S tertinggi yaitu 81.44±22.83 ppm. Peningkatan ion S dalam plasma ini disebabkan adanya penambahan garam MgSO4 ke dalam ransum. Keberadaan ion S dalam ransum PKAR asam dapat ditekan oleh suplementasi minyak ikan, sehingga domba yang diberi ransum asam dengan suplementasi minyak ikan mempunyai konsentrasi ion S plasma lebih rendah (26.78±1.39 ppm). Keberadaan minyak ikan dalam ransum mampu menekan konsenrasi ion S baik dalam ransum dengan nilai PKAR basal, basa maupun asam.
mempertahankan homeostasis ion Cl meskipun dalam ransum ditambahkan garam Cl yang berlebih. Rataan konsentrasi ion Cl plasma berkisar antara 4577±159.35 sampai 4851±226.93 ppm. Sama halnya dengan ion S, suplementasi minyak ikan dalam ransum mampu menekan konsenrasi ion Cl baik dalam PKAR basal, basa maupun asam.
Nilai pH semen domba yang diberi PKAR asam lebih rendah dari PKAR basal dan basa setelah penelitian berlangsung selama 28 hari. Hali ini memperlihatkan bahwa PKAR asam mempunyai peluang yang besar untuk memanipulasi kondisi semen Domba Garut. Kondisi pH semen yang lebih asam dari keadaan normal akibat PKAR asam, menyebabkan berkurangnya jumlah spermatozoa Y. Sebaliknya, kondisi pH semen yang lebih basa dari keadaan normal akibat PKAR basa akan menyebabkan berkurangna spermatozoa X. Hal ini disebabkan karena pada suasana asam spermatozoa Y akan mati sedangkan spermatozoa X lebih tahan pada suasana asam (Anonim 2006). Nurwansyah (2006) juga menyatakan bahwa spermatozoa Y akan lebih kuat dan lebih tahan hidupnya dalam suasana basa. Sebaliknya, spermatozoa X lebih kuat dan lebih tahan hidupnya pada suasana asam. Perubahan pH semen tersebut diduga akan merubah rasio spermatozoa X dan Y. Kondisi pH asam, menyebabkan persentase spermatozoa X lebih besar daripada Y, sebaliknya pada kondisi pH basa menyebabkan tingginya persentase spermatozoa Y. Dengan demikian, keadaan ini akan merubah proporsi jenis kelamin anak yang dilahirkan dari masing-masing ransum dengan PKAR yang berbeda.
Seluruh ternak percobaan menghasilkan volume, warna, kekentalan dan bau semen yang sama. Demikian pula halnya terhadap keadaan mikroskopis spermatozoa (gerakan massa, gerakan individu, konsentrasi, abnormalitas morfologi, persentase hidup spermatozoa) PKAR tidak banyak berpengaruh. Hal ini menunjukkan bahwa adanya prioritas pemanfaatan nutrisi pada Domba Garut jantan, ditujukan bagi produktivitas reproduksi sehingga mampu mempertahankan kualitas spermatozoa baik secara makroskopis maupun mikroskopis.
Persentase spermatozoa X dan Y domba garut yang diberi ransum dengan PKAR yang berbeda mempunyai perbandingan yang sama yaitu mendekati 50:50%.
PKAR mempengaruhi konsentrasi basa darah, cHCO3 dan konsentrasi mineral Mg dan S dalam plasma. Dibutuhkan waktu selama 28 hari untuk mempengaruhi pH semen sehingga mengikuti pola PKAR yang diberikan. Prioritas pemanfaatan nutrisi pada Domba Garut jantan, ditujukan bagi produktivitas reproduksi sehingga mampu mempertahankan kualitas spermatozoa baik secara makroskopis maupun mikroskopis.
© HAK cipta milik Institut Pertanian Bogor, tahun 2009 Hak cipta dilindungi undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
MANIPULASI KONDISI FISIOLOGIS DAN KEASAMAN SEMEN
MELALUI PENGATURAN PERBEDAAN KATION ANION
RANSUM DALAM UPAYA MEMPENGARUHI
KARAKTERISTIK SPERMATOZOA
PADA DOMBA GARUT
RAHMAT HIDAYAT
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Ilmu Ternak
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Manipulasi Kondisi Fisiologis dan Keasaman Semen Melalui Pengaturan Perbedaan Kation Anion Ransum dalam Upaya Mempengaruhi Karakteristik
Spermatozoa pada Domba Garut
Nama : Rahmat Hidayat
NIM : D061040011
Disetujui Komisi Pembimbing
Prof. Dr. Ir. Toto Toharmat, M.Agr.Sc. Ketua
Prof. Dr. drh. Arief Boediono Dr. Ir. Idat Galih Permana, M.Sc.
Anggota Anggota
Mengetahui
Ketua Program Studi Dekan Sekolah Pascasarjana IPB Ilmu Nutrisi dan Teknologi Pakan
Disertasi ini penulis persembahkan untuk:
Istriku tercinta : Nurrisky Murdjani, SE., Ak., M.Si.
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah SWT, karena berkat rahmat, petunjuk dan izin-Nya penulis dapat menyelesaikan disertasi ini dengan judul ‘Manipulasi Kondisi Fisiologis dan Keasaman Semen Melalui Pengaturan Perbedaan Kation Anion Ransum dalam Upaya Mempengaruhi Karakteristik Spermatozoa pada Domba Garut‘. Disertasi ini dibuat untuk memenuhi salah satu persyaratan dalam memperoleh gelar Doktor pada Sekolah Pascasarjana Institut Pertanian Bogor.
Penulis mengucapkan terima kasih dan penghargaan yang sebesar-besarnya kepada Bapak Prof. Dr. Ir. Toto Toharmat, M.Agr.Sc. sebagai ketua komisi pembimbing, Bapak Prof. Dr. drh. Arief Boediono dan Bapak Dr. Ir. Idat Galih Permana, M.Sc. sebagai Anggota Komisi Pembimbing atas segala bimbingan, arahan, masukan, perhatian dan nasihat mulai dari awal perencanaan sampai selesainya usulan penelitian ini.
Terima kasih dan penghargaan penulis sampaikan kepada Dr. Ir. Maradoli Hutasuhut, M.Sc., M.Ec. dan Dr. Ir. H. Suryahadi, DEA yang telah bersedia menjadi penguji luar komisi pada ujian terbuka. Dr. Ir. Komang G. Wiryawan selaku penguji luar komisi pada ujian tertutup. Prof. Dr. Ir. Wasmen Manalu, M.Sc. dan Dr. Ir. Jajat Jachja, M.Sc. sebagai penguji pada ujian prelium atas segala sumbangan pemikiran, masukan, saran, dan pengarahan penulisan disertasi mulai dari ujian tertutup sampai selesainya disertasi ini.
IPB. Ucapan yang sama penulis juga sampaikan kepada Pengelola Beasiswa Program Pascasarjana (BPPS) dan Tim Hibah Penelitian Pascasarjana Ditjen Dikti Depdiknas atas bantuan dana sehingga perkuliahan dan penulisan disertasi ini dapat berlangsung dengan baik.
Penulis juga mengucapkan terima kasih yang sebesar-besarnya kepada Dr. Dra. Iis Arifiantini, M.Si, Dr. Ir. Farida, M.Sc., Ir. H. Yudi Guntara Noor, H. Muhammad Saleh, SE, Mas Bondan, drh. Dedi, Mbak Dian, Bapak Ujang, Amir, Haer, Bapak Adi, Bapak Dimyati, Diah Anggreini S.Pt, M.Si., Bapak dan Ibu Anda, Drs. Nusyirwan M.Si, dan Dr. Ir. Dewi Apriastuti, MS atas segala bantuan baik immateri maupun materi yang telah diberikan sehingga penelitian dapat dilaksanakan dengan baik dan lancar.
Semoga semua amal baik tersebut di atas dapat diterima dan diridhoi oleh Alloh SWT dengan balasan berupa kebaikan yang banyak. Sebagai manusia biasa, tentunya penulis mempunyai banyak kesalahan selama berkomunikasi dan berinteraksi pada waktu melaksanakan pendidikan pascasarjana, terutama selama pelaksanaan penelitian ini. Oleh karena itu, penulis menghaturkan permohonan maaf yang sebesar-besarnya, lahir bathin, baik yang disengaja maupun yang tidak disengaja kepada semua fihak yang pernah berinteraksi dengan penulis.
Semoga disertasi ini bermanfaat dan semoga Alloh SWT senantiasa memberi hidayah, taufik, inayah, dan meridhoi amal perbuatan kita semua.
Bogor, Mei 2009
RIWAYAT HIDUP
Penulis dilahirkan di Sumedang pada 19 Februari 1969 sebagai anak ketujuh dari pasangan Bapak US Natamihardja, BA (Alm.) dan Ibu Eri Rohaeti. Menikah dengan Nurrisky Murdjani, SE. Ak., M.Si. pada 1996 dan sudah dikarunia tiga orang anak yaitu, Fikry Nurrahman, Azka Fitria dan Alya Zahira.
Pendidikan sekolah dasar sampai dengan sekolah menengah atas ditempuh di Sumedang. Pendidikan sarjana ditempuh di Fakultas Peternakan Universitas Padjadjaran Bandung masuk pada 1988 dan lulus pada 1994. Melanjutkan S2 di Program Pascasarjana Institut Pertanian Bogor masuk pada 1999 dan lulus 2002. Pada 2004 penulis diterima untuk Program Doktor di Program Studi Ilmu Ternak Sekolah Pascasarjana IPB.
Pada 1995 sampai dengan 1996 penulis bekerja di Dinas Peternakan Kabupaten Bogor di bagian Seksi Bina Program. Pada 1996 sampai sekarang penulis bekerja sebagai Staf Pengajar di Jurusan Ilmu Nutrisi Ternak, Fakultas Peternakan, Universitas padjadjaran. Bidang ilmu yang pernah diajarkan adalah Nutrisi Ternak Dasar, Pengetahuan Bahan Makanan Ternak, Nutrisi Ternak Ruminansia dan Mikrobiologi Nutrisi.
Bogor, Mei 2009
Penulis,
Penguji diluar komisis pembimbing :
1. Penguji pada ujian tertutup : Dr. Ir. Komang G. Wiryawan
2. Penguji pada ujian terbuka : Dr. Ir. Maradoli Hutasuhut, M.Sc., M.Ec.
DAFTAR ISI
Deskripsi Domba Garut ... 8Anatomi Sistem Reproduksi Domba Jantan ... 10
Testis ... 10
Spermatogenesis ... 12
Pematangan Spermatozoa ... 15
Pengendalian Spermatogenesis oleh Hormon ... 17
Semen ... 18
Spermatozoa ... 18
Plasma Semen ... 20
Karakteristik Semen Domba ... 21
Penentuan Jenis Kelamin ... 22
Keseimbangan Kation-Anion ... 24
Peran Mineral Na, K, Cl dan S ... 29
Mineral Seng ... 39
Metabolisme Seng dalam Tubuh Makhluk Hidup ... 40
Manifestasi Defisiensi Seng ... 41
Suplementasi Seng Bagi Ternak Ruminansia ... 41
Lemak ... 42
Asam Lemak Esensial ... 43
MATERI DAN METODE PENELITIAN Lokasi dan Waktu Penelitian ... 45
Metode Penelitian ... 45
Peubah yang Diamati ... 48
Analisis Data ... 54
HASIL DAN PEMBAHASAN Pengaruh Perlakuan terhadap Peubah Nutrisi (Konsumsi Bahan Kering, Kecernaan Bahan Kering dan Kecernaan Bahan Organik) dan Pertambahan Bobot Badan ... 56
Pengaruh Perlakuan terhadap pH Urin Ternak Percobaan ... 60
Pengaruh Perlakuan terhadap Gas Darah ... 64
Pengaruh Perlakuan terhadap Status Mineral Plasma Darah .... 69
Pengaruh Perlakuan terhadap Volume Semen ... 75 Pengaruh Perlakuan terhadap pH Semen ... 77 Pengaruh Perlakuan terhadap Gerakan Massa Spermatozoa .... 78
Pengaruh Perlakuan terhadap Gerakan Individu Spermatozoa ... 81 Pengaruh Perlakuan terhadap Persentase Spermatozoa Hidup . 81 Pengaruh Perlakuan terhadap Normalitas Spermatozoa ... 83 Pengaruh Perlakuan terhadap Konsentrasi Spermatozoa ... 83 Pengaruh Perlakuan terhadap Komposisi Spermatozoa X dan
Y ...
85
SIMPULAN DAN SARAN
Simpulan ... 89 Saran ... 89
DAFTAR PUSTAKA 90
DAFTAR TABEL
Tabel Halaman
1. Berat molekul, nilai valensi, berat equivalen dan berat miligram equivalen mineral-mineral yang digunakan untuk menghitung keseimbangan kation-anion ransum ...
27 2. Perbandingan konsentrasi bahan-bahan di luar dan dalam
membran sel mamalia ... 32
3. Jenis ransum perlakuan ... 45
4. Komposisi dan kandungan nutrien ransum basal ... 46
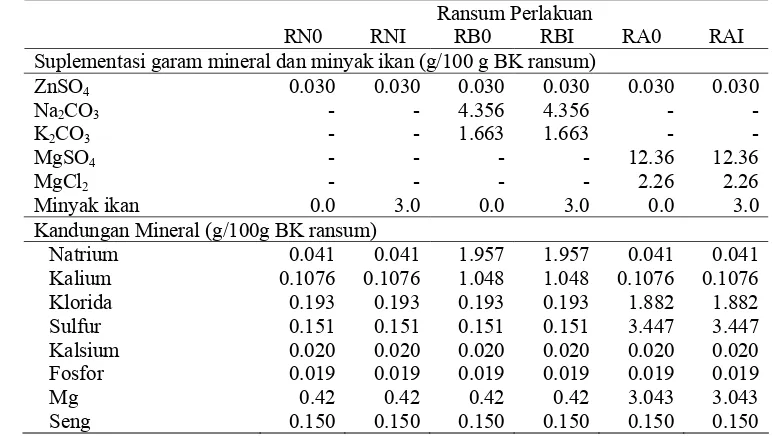
5. Suplementasi garam-garam mineral dan minyak ikan 47
6. Rataan konsumsi harian bahan kering (BK), kecernaan bahan kering (KCBK), kecernaan bahan organik (KCBO) ransum dan pertambahan bobot badan domba dengan nilai PKAR
ransum yang berbeda ... 57
7. Rataan harian pH urin Domba Garut yang mendapat ransum
dengan nilai PKAR yang berbeda ... 63
8. Rataan pH, gas dan haemoglobin darah Domba Garut yang
mendapat ransum dengan nilai PKAR berbeda ... 65
9. Rataan differensiasi butiran darah putih (BDP) Domba Garut
yang mendapat ransum dengan nilai PKAR berbeda ... 68
10. Rataan mineral plasma dan semen domba yang diberi ransum
dengan nilai PKAR berbeda... 71
11. Warna semen Domba Garut yang mendapat ransum dengan
nilai PKAR berbeda ... 74
12. Kekentalan spermatozoa Domba Garut yang mendapat
ransum dengan nilai PKAR berbeda ... 76
13. Rataan perkembangan volume semen Domba Garut yang
mendapat ransum dengan nilai PKAR berbeda ... 79
14. Rataan perkembangan pH semen Domba Garut yang
15. Rataan perkembangan gerakan masa spermatozoa Domba
Garut yang mendapat ransum dengan nilai PKAR berbeda ... 82
16. Rataan perkembangan gerakan individu spermatozoa Domba
Garut yang mendapat ransum dengan nilai PKAR berbeda ... 84
17. Rataan persentase spermatozoa hidup Domba Garut yang
mendapat ransum dengan nilai PKAR berbeda ... 84
18. Rataan perkembangan spermatozoa normal (normalitas) Domba
Garut yang mendapat ransum dengan nilai PKAR berbeda ... 86
19. Rataan perkembangan konsentrasi spermatozoa Domba Garut
yang mendapat ransum dengan nilai PKAR berbeda ... 86
20. Rataan persentase spermatozoa X Domba Garut yang
DAFTAR GAMBAR
Gambar Halaman
1. Domba Garut Jantan dewasa ... 9
2. Anatomi sistem reproduksi domba jantan ... 10
3. Tahap-tahap perkembangan sel spermatosit ... 14
4. Scanning elektron mikrograph spermatozoon ... 15
5. Morfologi spermatozoa ... 20
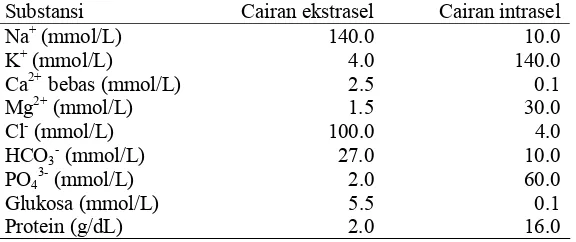
6. Keadaan cairan intraseluler dan ekstraseluler ... 31
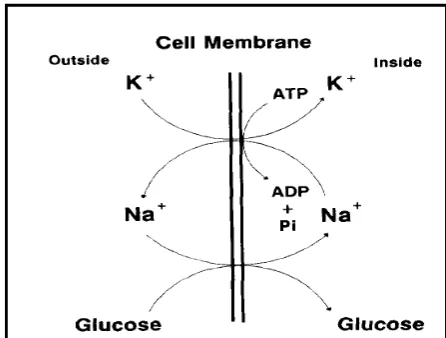
7. Mekanisme pemompaan yang bergantung pada Na+-K+-ATP pada waktu difusi glukosa ke dalam sel ... 33
8. Nefron dan duktus dalam sistem ekskresi urin ... 35
9. Tata letak satuan percobaan pada penelitian ... 48
10. Tahapan pengukuran luas area kepala spermatozoa mempergunakan perangkat lunak Image J ... 55
DAFTAR LAMPIRAN
Lampiran Halaman
1. Perhitungan penambahan mineral seng (Zn) dalam ransum .... 100
2. Perhitungan perbedaan kation anion ransum ... 101
3. Metode pewarnaan William ... 104
4. Tahapan pengukuran luas area kepala spermatozoa ... 105
PENDAHULUAN
Latar Belakang
Kondisi fisiologis dan tujuan pemeliharaan ternak menentukan jumlah dan jenis nutrien yang dibutuhkan. Ternak dewasa, membutuhkan nutrien selain untuk maintenance, produksi juga membutuhkan nutrien tertentu dengan jumlah yang mencukupi bagi fungsi reproduksinya. Kecukupan nutrien pokok seperti energi, protein, mineral dan vitamin dapat mempengaruhi reproduksi, sebaliknya bila kekurangan dapat menyebabkan performa reproduksi suboptimal.
Pakan yang baik adalah pakan yang dapat memenuhi semua kebutuhan nutrisi bagi ternak yang mengkonsumsinya. Hal ini meliputi kecukupan protein, karbohidrat dan mineral-mineral esensial sesuai dengan status fisiologis ternak. Ternak yang berbeda status fisiologisnya membutuhkan kandungan nutrisi yang berbeda pula dalam ransumnya.
Mineral merupakan zat makanan yang jumlahnya relatif sedikit dalam tubuh mahluk hidup, namun demikian zat tersebut merupakan mikro nutrien penting dalam kehidupan. Fungsi mineral dalam organisme sangat bervariasi dan berhubungan erat dengan kondisi dan bentuknya. Fungsi utamanya meliputi : keikutsertaannya dalam pembentukan jaringan; pemeliharaan homeostasi dalam cairan internal; pemeliharaan keseimbangan membran sel; pengaktifan reaksi biokimia dalam sistem enzim; berefek langsung atau tidak langsung pada kelenjar endokrin dan mempengaruhi simbiotik mikroflora saluran pencernaan.
penting untuk pembelahan sel, sintesis dan stabilitas DNA juga dalan diferensiasi sel (Devenson et al. 1993).
Keberadaan mineral dalam bentuk garam yang larut dalam medium sel, cairan interestitial, darah dan limpa, berpartisipasi langsung atau tidak langsung dalam menjaga homeostasi. Garam ketika dilarutkan dalam air, sebagian atau seluruhnya terpecah menjadi kation dan anion.
Ion diklasifikasikan sebagai grup dari satu atau beberapa atom yang mempunyai muatan listrik berdasarkan penambahan atau kehilangan elektronnya. Muatan listrik yang dibawa elektrolit ini akan mempengaruhi keseimbangan asam basa dan mempengaruhi metabolisme mineral. Ion dapat bermuatan positif atau negatif. Jumlah kation dan anion di dalam tubuh sangatlah banyak dan sangat responsif terhadap fungsi-fungsi biologis. Kondisi asam basa tubuh sangat ditentukan oleh jumlah kation dan anion yang dikonsumsi dari pakan. Kondisi ini sangat menentukan fungsi fisiologis tubuh. Fungsi fisiologis tubuh akan berubah jika keseimbangan asam basa dalam tubuh berubah. Hal ini akan mengakibatkan pergeseran fungsi fisiologis sesuai dengan keseimbangan asam basa yang ada.
Mineral merupakan ion yang sangat penting di dalam tubuh. Kation meliputi kalsium, potasium, sodium, magnesium dan lain-lain, sedangkan anion meliputi sulfur, oksigen, klor dan lain-lain. Sodium, potasium, sulfur dan klor merupakan ion-ion monovalen yang sangat kuat. Keseimbangan kation anion dalam pakan dinyatakan dalam equivalent atau milliequivalent.
Dietary cation-anion different (DCAD), yaitu perbedaan miliequevalen antara kation dan anion tertentu dalam ransum dengan cara pengurangan miliequivalen anion dari miliequivalen kation dalam seluruh ransum. Pada umumnya, mineral yang sering digunakan dalam perhitungan DCAD, yaitu dua macam kation (Na dan K) dan dua macam anion (Cl dan S). Perhitungan nilai DCAD yang digunakan oleh Harris and Beede (1993), Moore et al. (2000), Roche et al. (2003a), Roche et al. (2003b), dan Borucki Castro et al. (2004), berdasarkan persamaan Tucker et al. (1992) berikut ini.
Manipulasi pakan melalui perbedaan keseimbangan kation anion pada ternak ruminansia tidak banyak mempengaruhi perubahan pH darah, karena ginjal dan tulang dapat mempertahankan keseimbangan kation anion dan mempertahankan pH darah dalam keaadaan normal. Pakan yang “asam” akan mendorong mobilisasi tulang (osteocytic resorpsion) sebab tulang bekerjasama dengan ginjal bertindak sebagai buffer terhadap kondisi asam sistemik. Pakan yang “asam” mempunyai pengaruh yang kecil terhadap absorpsi kalsium dalam usus. Selain itu pakan ini juga dapat meningkatkan produksi 1.25 dihydroxy vitamin D per unit peningkatan parathyroid hormon.
Mineral merupakan unsur yang sangat penting dalam sistem reproduksi terutama yang berhubungan dengan spermatogenesis. Ion kalsium, kalium, natrium, klor dan fosfat merupakan konstituen cairan elektrolit dalam epididimis. Keseimbangan elektrolit dalam epididimis ini sangat penting untuk mendukung epitel epididimis sebagai tempat pematangan fisiologis spermatozoa (Wong and Yeung 1978; Brooks 1981).
Enzim ATP-ase yang sangat penting untuk mengkatalisis ATP selama proses kontraksi dan relaksasi protein kontraktil dinein aksonema flagelum spermatozoa, memerlukan fosfat anorganik, kalium-natrium dan kalsium (Mitchell et al. 1976; Brooks 1981). Struktur membran spermatozoa mengandung fosfolipid, dengan demikian fosfat sangat diperlukan agar membran spermatozoa berfungsi normal (Mitchell et al. 1976; White et al. 1976; Pederson and Fawcet 1976).
Salah satu plasma nutfah Indonesia yang memiliki peluang untuk dikembangkan dalam ruang lingkup nasional adalah Domba Garut. Domba ini dikelompokkan kedalam tipe medium dengan kualitas daging yang baik. Selain itu, kulit Domba Garut dikenal memiliki kualitas terbaik dibanding domba tipe medium yang lain.
Diperkirakan 50 persen dari populasi domba nasional (4.6 juta ekor) ada di Jawa Barat. Populasi terbesar (80 persen) adalah jenis Domba Garut atau merupakan persilangan Domba Garut dengan bangsa domba lain. Peranan Domba Garut sangat penting bagi masyarakat Jawa Barat karena ditinjau dari aspek sosio-ekonomi ternak ini memiliki nilai yang sangat penting misalnya untuk seni ketangkasan/kesenian domba tangkas.
Di Jawa Barat, konsumsi daging rata-rata 4.30 kg/kapita/tahun atau baru mencapai 42.57% dari norma gizi (10.1 kg/kapita/tahun). Ini berarti setara dengan 267 ribu ekor sapi atau 2.5 juta ekor domba. Untuk skala nasional, jumlah tersebut akan jauh lebih besar. Kontribusi Domba Garut sebagai sumber protein hewani nampaknya belum optimal. Oleh karena itu, untuk mencapai norma gizi sesuai dengan harapan maka masih sangat dibutuhkan ternak potong baik bagi Jawa Barat maupun nasional. Sebagian besar kebutuhan daging nasional masih sangat tergantung dari impor sehingga kebijakan tersebut harus dievaluasi kembali.
Di bidang reproduksi, Domba Garut dikenal memiliki sifat prolifikasi yang baik yaitu rata-rata 175%. Kualitas pakan yang baik sangat penting untuk mendapatkan efisiensi produksi dan efisiensi ekonomi yang baik. Dengan demikian perlu kajian mendalam dalam pengembangan aplikasi teknologi terpadu antara pakan dan reproduksi. Kualitas semen sangat ditentukan oleh kesehatan reproduksi pejantan. Pakan yang baik dan aplikasi bioteknologi reproduksi akan sangat bermanfaat untuk mendapatkan anak yang sehat, memiliki daya hidup tinggi dan pertumbuhan yang baik.
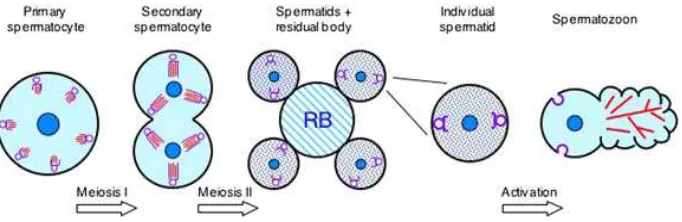
Spermatozoa terbentuk dalam testes melalui serangkaian pembelahan yakni spermatogenesis. Proses spermatogenesis terdiri dari tiga fase yaitu (1) spermatocytogenesis, proses spermatogonia berkembang menjadi spermatosit, (2) meiosis, tahap matang dari spermatosit yang menghasilkan spermatid dengan jumlah kromosom berkurang (haploid), dan (3) spermiogenesis, proses transformasi dari spermatid menjadi spermatozoa. Spermatocytogenesis dikendalikan oleh FSH dari adenohyphophysis. Spermiogenesis berada di bawah pengaruh LH dan testosteron (Toelihere 1985). Hasil pembentukan spermatozoa dilepaskan dengan proses yang disebut spermiasi dari sel-sel sertoli dan memasuki lumen tubuli seminiferi menuju rete testis. Pada domba, keseluruhan proses spermatogenesis berlangsung antara 46-49 hari (Salisbury and Van Demark 1985; Bearden and Fuquay 2000).
Pakan di Indonesia rata-rata mengandung seng yang rendah yaitu berkisar antara 20-38 mg/kg dari bahan kering (Little 1986), sedangkan kebutuhan seng untuk ternak ruminansia berkisar 40-50 mg/kg dan kebutuhan untuk mikroba rumen yaitu antara 130-220 mg/kg (Arora 1989). Dosis letal Seng bagi ternak domba adalah 750 ppm (Salt Institut 2001). Mineral seng terdapat pada semua jaringan tubuh dan esensial bagi ternak. Seng memiliki berbagai fungsi dalam tubuh. Seng mempengaruhi pertumbuhan, perkembangan, fungsi reproduksi, pembentukan darah dan tulang dan metabolisme asam nukleat, protein dan karbohidrat. Dalam proses ini, seng berhubungan dengan enzim dimana seng sebagai komponen esensial atau aktivator dari enzim (Georgievskii et al. 1982). Seng dalam rumen memacu pertumbuhan mikroba rumen (Putra 1999), meningkatkan penampilan ternak (Hartati 1998) dan immunitas pada domba (Hernaman et al. 2003).
(poly unsaturated fatty acid = FUFA), yang mengandung asam lemak arakhidonat (C20:4n-6) cukup tinggi (21.97%) dan dapat dipakai sebagai prekursor prostaglandin terutama PGE2 yang paling dominan berperan dalam peningkatan absorbsi seng. Disamping itu asam lemak yang terkandung dalam minyak lemuru juga dapat digunakan sebagai sumber energi, karier vitamin D, agensia defaunasi dan mereduksi emisi metan. Salah satu sumber asam lemak esensial adalah minyak ikan lemuru (Sardinella longiceps). Ditinjau dari segi produksi, ikan lemuru cukup potensial sebagai sumber minyak ikan.
Suplementasi mineral Seng (Zn) dan asam lemak esensial asal minyak ikan lemuru bagi ternak Domba Garut jantan diharapkan mampu dijadikan sebagai bahan stimulan peningkatan kemampuan reproduksinya.
Penelitian ini mengkaji tentang peran suplementasi asam lemak esensial dan keseimbangan anion kation dalam ransum untuk mendukung reproduksi ternak Domba Garut. Kajian ini diharapkan dapat mengungkapkan peran asam lemak esensial dan keseimbangan anion kation dalam ransum terhadap kondisi fisiologis dan karakteristik spermatozoa Domba Garut.
Tujuan Penelitian Tujuan penelitian ini yaitu untuk :
1. Mengetahui pengaruh nilai neraca kation-anion dan suplementasi asam lemak esensial dalam ransum berkecukupan Zn terhadap konsumsi pakan, metabolisme nutrient, keadaan fisiologis dan kondisi kimia darah.
2. Mendapatkan informasi keterkaitan pengaruh nilai neraca kation-anion (negatif, netral dan positif) dan suplementasi asam lemak esensial terhadap metabolisme Zn, Na, K, Cl serta S.
3. Mengkaji pengaruh nilai neraca kation-anion dan suplementasi asam lemak esensial dalam ransum berkecukupan Zn terhadap karakteristik spermatozoa dan rasio sperma X:Y yang dihasilkan ternak ruminansia jantan.
Zn, Na, K, Cl dan S, kondisi fisiologis, karakteristik spermatozoa dan rasio sperma X:Y.
Hipotesis Penelitian
TINJAUAN PUSTAKA
Deskripsi Domba Garut
Domba merupakan salah satu jenis ternak ruminansia kecil yang sangat
populer di masyarakat Indonesia. Selain mudah dipelihara, domba juga disenangi
masyarakat karena dapat berfungsi sebagai tabungan yang mudah
diperjualbelikan. Menurut Sudarmono dan Sugeng (2005), di Indonesia terdapat
berbagai tipe domba, yaitu (1) Domba asli Indonesia atau disebut dengan domba
kampung atau lokal. Ciri-cirinya, berbadan kecil, lambat dewasa, warna bulu
tidak seragam, dan karkasnya rendah. (2) Domba Priangan atau disebut dengan
Domba Garut yang merupakan persilangan antara domba asli, merino, dan ekor
gemuk dari Afrika Selatan. Domba Garut banyak terdapat di Garut sebagai
domba laga dengan ciri-ciri sebagai berikut: mempunyai tubuh besar dan lebar (60
kg untuk jantan dan 35 kg untuk betina); jantan bertanduk dan melengkung ke
belakang; daun telinga ramping; warna bulu kombinasi putih hitam dan cokelat
atau warna campuran. (3) Domba ekor gemuk banyak terdapat di Jawa Timur,
Madura, Sulawesi, dan Lombok. Ciri-cirinya, bentuk badan besar (50 kg untuk
jantan dan 40 kg untuk betina), bertanduk pada yang jantan, dan berekor panjang
(pada bagian pangkalnya besar dan menimbun lemak yang banyak, ujung ekornya
kecil tidak berlemak).
Domba (Ovis aries) menurut sistematikanya diklasifikasikan sebagai berikut :
Kingdom : Animalia
Filum : Chordata
Kelas : Mammalia
Ordo : Artodactyla
Famili : Bovidae
Genus : Ovis
Spesies : Ovis aries
Domba Garut merupakan bangsa domba yang berasal dari Jawa Barat
yaitu Kabupaten Garut. Domba Garut disebut juga Domba Priangan termasuk
1864 merupakan hasil persilangan tiga bangsa yaitu Domba Merino dari
Australia, Domba Kapstad dari Afrika Barat Daya dan domba lokal (Devendra
and McLeroy 1982).
Ciri khas Domba Garut adalah sebagai berikut: propil kepala cembung,
bentuk muka bagian atas lebar, pendek dan sedikit cembung. Daun telinga
memiliki kombinasi antara kuping rumpung/rudimenter (di bawah 4 cm) atau
ngadaun hiris (4-8 cm). Bentuk ekor lebar dan berlemak pada pangkal ekor dan mengecil ke ujung (ngabuntut beurit atau ngabuntut bagong) . Domba jantan mempunyai tanduk besar, kokoh, kuat dan melingkar, sedangkan betina tidak
bertanduk. Bobot hidup jantan dewasa antara 50-80 kg, sedangkan betina dewasa
35-40 kg. Ternak betina prolifik tinggi dan mempunyai selang kelahiran pendek.
Warna bulu bervariasi hitam, putih, coklat dan belang (Heriyadi 2006; Devendra
and McLeroy 1982; Mulyono 2003).
Keturunan hasil persilangan ini baik karena memiliki daya produksi wool
dari Merino dan tinggi badan lebih tinggi dari Kaapstaad. Pusat pengembangan
Domba Garut yang ada dewasa ini berada di daerah Garut, hasilnya disebar ke
daerah lain seperti Sumedang, Bandung, Tasikmalaya dan sekitarnya.
Berdasarkan dari tinggi badannya, domba ini cocok sebagai domba aduan
(Merkens and Soemirat 1979). Seni Ketangkasan Domba Garut diperkirakan
telah berlangsung sejak terbentuknya bangsa domba ini. Domba Garut memiliki
penampilan yang menarik sehingga sering digunakan sebagai hewan peliharaan
(Gambar 1.)
Anatomi Sistem Reproduksi Domba Jantan
Organ reproduksi domba jantan terdiri atas tiga komponen yaitu : (1)
organ kelamin primer yaitu testes, (2) kelenjar-kelenjar kelamin pelengkap yaitu
kelenjar vesikularis, kelenjar prostat, kelenjar bubourethalis dan saluran-saluran
epididimis serta duktus deferen, (c) alat kelamin luar yaitu penis (Toelihere 1993).
Anatomi sistem reproduksi domba jantan dapat dilihat pada Gambar 2.
Komponen-komponen yang penting pada gonand jantan adalah tubulus seminiferus yang mensekresikan spermatozoa dan sel leydig yang terdapat pada jaringan interstitial yang mensekresikan androgen. Androgen mempertahankan
sifat seks sekunder (jengot, suara, tanduk, jengger, agresivitas dan sebagainya)
dan kelenjar aksesoris (kelenjar-kelenjar prostat, vesikula seminalis dan kelenjar
Cowper). Sekresi kelenjar aksesori merupakan komponen essensial semen.
Gambar 2 Anatomi sistem reproduksi domba jantan (Kott 2008).
Testis
Testis selain merupakan kelenjar endokrin karena menghasilkan hormon,
juga berfungsi sebagai kelenjar eksokrin karena menghasilkan sel-sel
spermatozoa. Testes pada hewan mamalia, terletak dalam kantong skrotum dan
mempunyai dua lobus. Masing-masing lobus terbungkus oleh kapsula fibrosa
membujur epitel selom, sebelah kiri dan kanan mesenteri dorsal yang biasanya
dinamakan tonjolan genital (genital ridge). Sel gamet (spermatozoa) berkembang dari sel germinal primordium (SGP) yang berasal dari luar gonad. SGP mula-mula ditemukan pada kantong kunir, kemudian bergerak menuju usus belakang
(hind gut) dan melalui mesenteri dorsal sampai kepada tonjolan genital (Suhana and Rafiah 1982). Selanjutnya epitel selom mengelilingi sel-sel mesenkim dan
SGP, membentuk bakal tubulus (sex cord) yaitu tubulus seminiferus yang masih sederhana. Sel-sel interstisial antara bakal tubulus berkembang menjadi sel
Leydig yang dapat membentuk hormon androgen, sedangkan bakal tubulus akan membentuk tubulus seminiferus (Johnson and Everitt 1980). Proses pembentukan
tetis memerlukan hormon testosteron yang dihasilkan oleh sel Leydig. Sel ini terus menerus membuat androgen sampai beberapa bulan setelah kelahiran,
kemudian berhenti berproduksi dan akan aktif kembali ketika masa pubertas
sampai dewasa (Johnson and Everitt 1980).
Ditinjau secara histologis, testis pada hewan mamalia umumnya terdiri
atas jaringan epitel seminiferus, yaitu jaringan pengikat dinding tubulus
seminferus, jaringan pengikat intertubuler dan jaringan pengikat padat
pembungkus testis. Epitel seminiferus terdiri atas dua macam sel yaitu sel
penunjang atau sel sertoli dan sel-sel kelamin dari berbagai tingkat
perkembangan. Sel sertoli terdapat sepanjang membran basalis yang mudah
dibedakan dari sel kelamin, karena bentuk selnya torak, inti oval,
nukleoplasmanya homogen dan anak intinya jelas. Jumlah sel sertoli pada
mamalia dewasa umumnya tetap dan sangat resisten terhadap zat-zat yang dapat
membunuh atau merusak sel-sel kelamin (Oakberg 1959; Rowley and Heller
1971). Oleh karena itu, sel sertoli dapat digunakan sebagai faktor pembanding
yang konstan dalam analisis kuantitatif dari epitel seminiferus.
Sel sertoli mempunyai fungsi penting yang erat kaitannya dengan
kelangsungan hidup sel kelamin, antara lain:
1. Sebagai sumber nutrisi bagi sel-sel kelamin, karena ada hubungan yang erat
antara jumlah glikogen dalam sertoli dengan perkembangan sel kelamin pada
2. Sebagai sel sekresi antara lain menghasilkan protein pengikat androgen (ABP)
yang berperan sebagai alat pengangkut androgen ke sel-sel kelamin (Frecnh
and Ritzen 1973), inhibin (Steinberger 1979) dan estrogen (Lacy 1967).
3. Fagositosis terhadap sel-sel kelamin yang mengalami degenerasi atau rusak
dan memfagositosis sisa protoplasma spermatozoa dewasa (residual bodies)
yang banyak terdapat dalam tubulus (Carr et al. 1968).
4. Berfungsi sebagai penghalang darah testis (blood-testis barrier). Secara ultrastruktur, sitoplasma sel sertoli membentuk tonjolan-tonjolan yang
kemudian bercabang-cabang seperti tangan. Cabang-cabang tersebut
menyebar diantara sel-sel kelamin dan memberi kesan melindungi
perkembangan sel-sel kelamin. Cabang sitoplasma sel sertoli yang berdekatan
akan saling bertautan erat sekali, sehingga menghambat darah masuk ke dalam
tubulus seminiferus. Pertautan cabang sel sertoli yang berdekatan disebut
sertoli-sertoli junction (Dym and Fawcet 1970; Dym 1973; Fawcett 1975). Sel-sel kelamin pada epitel seminiferus terdiri atas berbagai tingkat
perkembangan. Letak sel-sel kelamin dalam tubulus seringkali dihubungkan
dengan tingkat perkembangannya. Makin dewasa tingkat perkembangan sel
kelamin, makin dekat letaknya ke arah lumen. Sebaliknya, sel-sel kelamin yang
tingkat perkembangannya belum maju, misalnya spermatogonia, letaknya pada
membran basalis (Soeradi 1987).
Spermatogenesis
Spermatogenesis merupakan rangkaian proliferasi dan diferensiasi sel
spermatogonia di dalam tubulus seminiferus testis membentuk spermatozoa.
Lamanya spermatogenesis adalah waktu yang diperlukan untuk menyelesaikan
seluruh proses tersebut, termasuk tahapan dalam rangka menambah jumlah
spermatogonia. Rangkaian tahapan spermatogenesis dibagi menjadi tiga tahapan.
Tahap pertama, sel spermatogonia mengadakan pembelahan mitosis menghasilkan
spermatosit dan sel induk spermatogonia (stem cell). Tahap kedua adalah tahap pembentukan spermatid yang haploid dari spermatosit melalui pembelahan
reduksi (meiosis). Tahap ketiga adalah pembentukan spermatozoa dari spermatid
Secara morfologis, dinamika epitel seminferous dapat dibagi dalam aspek
sitologis dan histologis. Aspek sitologis antara lain meliputi proliferasi,
pembaruan (renewal), diferensiasi spermatogonia, meiosis spermatosit, spermiogenesis dan spermiasi. Selanjutnya untuk memperoleh gambaran aspek
histologis tersebut, maka epitel seminiferus secara keseluruhan harus dilihat
sebagai satu kesatuan yang utuh. Dengan cara demikian akan dapat dibedakan
berbagai kumpulan sel-sel epitel seminiferus (asosiasi sel) yang terbesar di
sepanjang tubulus.
Menurut Steinberger and Steinberger (1975), ada beberapa hal penting
dari kinetika spermatogenesis, antara lain:
1. Siklus epitel seminiferus
Epitel seminiferus pada hewan mamalia dewasa terdiri atas sel sertoli
dan sel-sel kelamin (sel germinal) yang berada pada perkembangan tertentu.
Adanya tingkatan perkembangan sel-sel germinal pada tubulus seminferus
tersebut, terutama disebabkan oleh perbedaan waktu proliferasi dan
diferensiasi dari sel induk spermatogonia.
2. Spermiogenesis
Segera setelah terbentuk spermatosit primer, akan memasuki profase I
dari pembelahan reduksi, dengan urutan menuju ke arah lumen adalah
leptoten, zigoten, pakiten, diploten dan diakinesis. Setelah itu, spermatosit
primer mengalami pembelahan reduksi pertama untuk membentuk spermatosit
sekunder. Kemudian mengalami pembelahan reduksi kedua membentuk
sprmatid yang haploid. Spermatid yang terbentuk akan mengalami
serangkaian perubahan morfologi yang rumit membentuk spermatozoa,
dinamakan proses spermiogenesis. Rangkaian perubahan tersebut dapat dibagi
menjadi empat fase yaitu : fase golgi, fase tudung (cap), fase akrosom, dan fase pematangan atau maturasi. Beberapa tahapan perkembangan sel
Gambar 3 Tahap-tahap perkembangan sel spermatosit (Smith 2006).
Pada spermatid yang baru terbentuk, terdapat badan golgi yang terletak
pada sitoplasma dekat inti. Tanda diferensiasi pertama yang terlihat adalah
adanya granula dalam badan golgi. Granula ini disebut dengan proakrosom
yang banyak mengandung karbohidrat. Pada perkembangan selanjutnya
granula-granula akan bersatu membentuk granula akrosom dalam suatu
kantong akrosom yang melekat pada sisi luar inti. Pada tingkat perkembangan
seperti ini spermatid tersebut berada dalam fase golgi. Pada fase tudung,
membran kantong akrosom memperluas area perlekatannya pada membran
inti, dan akhirnya melingkupi seluruh belahan anterior dari inti (Clermont and
Leblon 1955; Steinberger dan Steinberger 1975).
Pada fase akrosom, terjadi kondensasi nukleoplasma dan pemanjangan
spermatid, serta terjadi redistribusi materi akrosom. Pada fase ini materi
akrosom yang tadinya terkumpul pada ujung anterior, mulai menyebar ke
dalam lipatan membran kantong akrosom membentuk tudung akrosom. Pada
fase pematangan, sistem akrosom tidak mengalami perkembangan berarti,
tetapi materi ini terus mengalami perkembangan sampai membentuk masa
padat yang homogen. Pada awal perkembangan kantong akrosom, sentriol
bergerak ke ujung berlawanan dari spermatid. Di sini sentriol distal terletak
tegak lurus pada permukaan sel dan membentuk flagel spermatozoa (Clermont
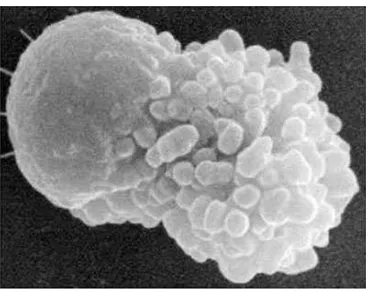
and Leblon 1955; Steinberger dan Steinberger 1975). Hasil scanning elektron
Gambar 4 Scanning elektron mikrograph spermatozoon (Smith 2006).
Pematangan Spermatozoa
Secara garis besar kemampuan fungsional spermatozoa meliputi dua hal
utama yaitu kemampuan gerak dan kemampuan dalam melakukan fertilisasi
(Soeradi 1990). Setelah terbentuk di dalam tubulis seminiferus, spermatozoa
belum memiliki kemampuan tersebut. Kemampuan ini diperoleh setelah
spermatozoa ke luar meninggalkan tubulus seminiferus testis, dan mengalami
pematangan secara perlahan dan bertahap, dimulai dari kaput, korpus, dan
berakhir di kauda epididimis. Pada umumnya proses maturasi spermatozoa di
dalam epididimis memerlukan waktu 14 hari (Steinberger dan Steinberger 1975).
Tanpa hormon androgen, epididimis akan mengecil dan beratnya menurun
hingga 25% dari normal. Pemberian testosteron pada hewan kastrasi,
mengakibatkan berat epididimis meningkat tiga kali dibanding kontrol. Jika
androgen dicegah masuk ke dalam lumen epididimis maka proporsi protein
terlarut dan fosfolipid menjadi rendah, demikian juga kandungan RNA. Hal ini
menunjukkan bahwa untuk menjalankan fungsinya, epididimis sangat tergantung
pada ambang androgen (high threshold of androgen) (Jindal and Panda 1980). Sel-sel epitel epididimis melakukan transfort aktif elektrolit dalam
menjalankan fungsinya sebagai tempat maturasi, Elektrolit utama yang
didistribusikan melintasi membran sel ialah Na+, K+, Ca+ HPO4 dan Cl-. Air
diabsorpsi dari lumen sehingga sekitar 98% cairan testis diserap ke dalam
Senyawa-senyawa yang disintesis dan diekskresikan oleh epitel epididimis
ialah protein, glikoprotein, fosfolipid, gliseril fosforilkolin, karnitin, asam sialat
dan inositol. Fosfolipid penting sebagai substrat maturasi, yaitu untuk
menstabilkan sistem membran spermatozoa. Bahan ini juga sebagai prekursor
gliseril fosforilkolin, yang penting untuk metabolisme spermatozoa. Jika plasma
semen tiba pada saluran kelamin betina, gliserol dilepaskan dan ATP dipakai
sebagai sumber energi, disamping glukosa dan fruktosa (Riar et al. 1973).
Karnitin diperlukan untuk transfort asam lemak berupa senyawa fatty acyl
ke dalam mitokondria spermatozoa dan digunakan pula sebagai sumber energi,
sedangkan fosfolipid sebagai substrat maturasi bersama inositol. Adenosin
trifosfat (ATP) bagi spermatozoa berasal dari pemecahan karbohidrat (glukosa
dan fruktosa) dan juga dari lipid. Pemecahan karbohidrat tersebut dapat
berlangsung secara aerob maupun anaerob. Di dalam epididimis spermatozoa
berada dalam lingkungan anaerob. Untuk memperoleh energi, ATP dipecah oleh
enzim ATP-ase pada dinein aksonema flagelum spermatozoa, menjadi ADP dan
Pi dengan membebaskan energi (Lewin et al. 1976).
Enzim ATP-ase pada spermatozoa ada yang memerlukan ion Ca+ dan ada
pula yang memerlukan ion Na+ dan K+. Ini berarti konsentrasi elektrolit tersebut
diperlukan dalam jumlah yang cukup, agar metabolisme ATP lancar, sehingga
energi untuk maturasi cukup. Gangguan transfort zat elektrolit dapat disebabkan
oleh defisiensi androgen, hal ini dapat menghambat metabolisme ATP, sehingga
bahan yang bersifat antiandrogen bermanfaat sebagai antimaturasi. Berbagai
enzim dalam epididimis juga telah diidentifikasi, dan meliputi
glutamat-oksaloasetat transaminase (GOT), laktat dehidrogenase (LDH), beta-Nasetil
glukosaminidase, alfa-amidase, fosfatase asam, dan fosfatase basa. Adanya
enzim-enzim ini dalam jumlah yang mencukupi, akan tetap mempertahankan
fungsi epididimis sebagai tempat maturasi dan penampungan sementara
spermatozoa. Sebaliknya jika ada gangguan terhadap jumlah dan kualitas
Pengendalian Spermatogenesis oleh Hormon
Spermatogenesis merupakan proses yang kompleks dan sangat bergantung
pada kadar hormon reproduksi. Kelenjar hipotalamus mensekresi GnRH
(Ginadotropin Releasing Hormone), hormon ini kemudian merangsang kelenjar hipofisis untuk melepaskan LH dan FSH (Franchimont et al. 1978). Kedua hormon peptida ini penting dalam mengontrol proses seluler pada sel target,
antara lain; aliran ion, aktivitas enzim, sintesis protein, sintesis dan sekresi
hormon steroid, proliferasi dan diferensiasi sel germinal, motilitas sel, dan
komunikasi antar sel (Means et al. 1980). Pentingnya hormon tersebut dalam memelihara dan mempertahankan spermatogenesis, yang dikenal dengan proses
pengendalian fertilitas melalui poros hipotalamus-hipofisis-testis.
LH merangsang sel Leydig untuk mensintesis dan mensekresikan
testosteron. Fungsi FSH yaitu merangsang sel sertoli untuk mensintesis ABP,
memelihara pertumbuhan sel germinal, meningkatkan sensitivitas sel Leydig
terhadap LH pada proses steroidogenesis, memulai spermatogenesis tahap-tahap
mitosis dan menyempurnakan spermatogenesis dalam fase spermiogenesis dan
spermiasi (Means et al. 1980; Rommerts 1990).
Hormon FSH dan testosteron bekerja sinergis pada sel sertoli untuk
menghasilkan zat-zat makanan yang diperlukan untuk proliferasi dan diferensiasi
sel-sel germinal dalam rangka menghasilkan spermatozoa yang fungsional. Di
samping itu, testosteron yang berdifusi ke sel-sel peritubuler diperlukan untuk
menghasilkan sejenis protein yang berfungsi sebagai faktor pemicu sel sertoli
(promote mudulation Sertoli), yang penting untuk meningkatkan aktivitas sel sertoli guna menghasilkan zat makanan bagi sel germinal dalam tubulus
seminiferus (Rommerts 1990).
Protein pengikat androgen (ABP) yang dihasilkan oleh sel sertoli di bawah
pengendalian FSH tersebut, berfungsi sebagai alat pengangkut androgen dari
jaringan interstisial ke epitel germinal melalui sel Sertoli. Hal ini diperkirakan
merupakan salah satu cara FSH untuk meningkatkan aktivitas androgenik dari
testosteron (efek sinergis) terhadap spermatogenesis (Means et al. 1980).
jumlah yang mencukupi di dalam lumen tubulus seminiferus testis, selain
berperan mengikat androgen untuk spermatogenesis, juga akan membawa
androgen dari tubulus seminiferus testis ke dalam epididimis. Androgen sangat
diperlukan juga pada proses pematangan fisiologis spermatozoa selama berada di
dalam tubulus epididimis (Franchimont et al. 1978; Means et al. 1980; Rommerts 1990).
Semen
Semen diekskresikan oleh organ reproduksi jantan. Semen terdiri dari
spermatozoa dan plasma. Spermatozoa yaitu suatu suspensi yang mengandung
sel-sel gamet yang mengandung game jantan, dihasilkan oleh testes. Plasma
semen merupakan bagian cairan semen yang diproduksi oleh epididimis,
vesikularis dan prostat (Hafez 1987 and Toelihere 1985). Kandungan semen
meliputi spermatozoa, air, gula sederhana (sebagai sumber energi bagi sperma),
alkali (untuk melindungi sperma dari suasana asam pada saluran jantan uretra atau
vagina) prostaglandin (substansi yang menyebabkan terjadinya kontraksi pada
uterus, tuba falopii), vitamin C, mineral Zn dan kolesterol. Selanjutnya Toelihere
(1985) menyatakan bahwa sebagian sifat fisik dan kimiawi semen ditentukan oleh
plasma semen.
Spermatozoa
Spermatozoa terbentuk dalam testes melalui serangkaian pembelahan sel
yakni proses spermatogenesis. Proses spermatogenesis terdiri atas tiga fase yaitu
(1) spermatocytogenesis, yaitu proses berkembangnya spermatogonia menjadi spermatosit, (2) meiosis, tahap matang dari spermatosit yang menghasilkan spermatid dengan jumlah kromosom berkurang (haploid), (3) spermiogenesis,
yaitu proses transformasi dari spermatid menjadi spermatozoa.
Spermatocytogensis dikendalikan oleh FSH dari adenohyphophysis dan
spermiogenesis berada di bawah pengaruh LH dan testosteron ( Toelihere 1993).
Hasil pembentukan spermatozoa yang baru kemudian akan dilepaskan dengan
proses yang disebut spermiasi dari sel-sel sertoli dan memasuki lumen tubuli
berlangsung antara 46-49 hari (Salisbury and VanDemark 1985; Bearden and
Fuquay 2000).
Ejakulat pada domba dan kambing dapat berbeda-beda secara kuantitas
maupun kualitas. Jumlah spermatozoa per ejakulat tergantung pada volume dan
konsentrasi semen. Motilitas dan morfologi spermatozoa termasuk dalam
karakteristik kualitas. Motilitas spermatozoa sangat penting dan erat kaitannya
dengan fertilitas. Bagaimana pun motilitas tidak selalu diindikasikan sebagai
kemampuan untuk membuahi dengan baik. Spermatozoa normal kehilangan
kemampuan membuahi sebelum motilitasnya hilang, namun spermatozoa yang
abnormal dapat memperlihatkan pergerakan yang normal tapi untuk selanjutnya
tidak mampu untuk membuahi (Garner and Hafez 1993).
Abnormalitas spermatozoa merupakan suatu keadaan dimana terjadi
kelainan pada kepala, ekor maupun kepala dan ekor yang terpisah. Abnormalitas
dapat digolongkan atas tiga kategori yaitu abnormalitas primer, sekunder dan
tersier. Abnormalitas primer terjadi karena kegagalan dalam proses
spermatogenesis, abnormalitas sekunder terjadi selama perjalanan spermatozoa ke
dalam epididimis, sedangkan abnormalitas tersier terjadi selama atau sesudah
ejakulasi dan pada waktu proses penanganan semen (Hastono 2001).
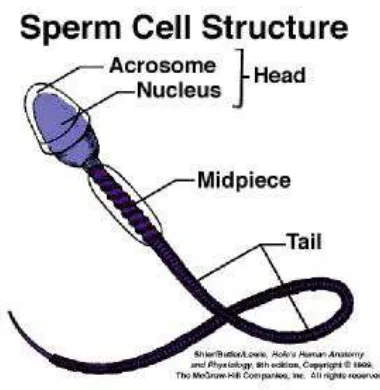
Spermatozoa yang normal terdiri dari kepala dan ekor. Bagian utama dari
kepala yaitu nukleus yang tertutupi oleh akrosom pada bagian depan dan
membran postnuclear pada bagian belakang. Sedangkan pada baian ekor terdapat dua fibril sentriol (pada bagian tengah) yang dikelilingi sebuah cincin yang terdiri
dari sembilan pasang fibril perifer (Faulkner and Pineda 1975).
Panjang kepala spermatozoa sapi, domba dan babi berkisar antara 8-10 μm
x 4-4.5 μm, dan tebalnya sekitar 0.5-1.5 μm (Toelihere 1985). Panjang total
spermatozoa domba atau kambing sekitar 60 μm dan panjang kepalanya saja
adalah 8-10 μm, lebarnya 4 μm dan tebalnya 1 μm (Evans and Maxwell 1987).
Bagian kepala spermatozoa mengandung materi inti, di dalamnya terdapat
kromosom yang terdiri dari DNA (deoxyribonucleid acid) yang bersenyawa dengan protein. DNA membawa informasi genetik (Foote 1980). Melalui
pembelahan reduksi selama proses spermatogenesis, spermatozoa hanya
untuk seks betina dan Y untuk seks jantan. Morfologi spermatozoa dapat dilihat
pada gambar 5.
Gambar 5 Morfologi spermatozoa (Kott 2008).
Plasma Semen
Plasma semen merupakan cairan ekstraseluler yang menyediakan media
dan transfort untuk spermatozoa. Cairan ini disekresikan oleh kelenjar reproduksi
pekengkap seperti kelenjar vesica seminalis, kelenjar prostat dan kelenjar
bulbourethralis (Mann 1964; Partodihardjo 1987).
Plasma semen mempunyai fungsi yang penting pada ejakulasi dan pada
fase awal pengangkutan spermatozoa. Selain itu, plasma semen berfungsi sebagai
medium suspensi dan dapat juga menjadi substrat metabolisme yang penting bagi
spermatozoa dengan kandungan khususnya seperti fruktosa, asam sitrat dan
inositol (Mann 1974). Menurut Bearden and Fuquay (2000), plasma semen
berperan sebagai buffer dan sumber energi bagi spermatozoa. Cairan ini juga mengandung bahan organik dan anorganik dan mempunyai tekanan osmotik
plasma sama dengan tekanan darah (ekuivalen dengan NaCl 0.9%). Derajat
Karakteristik Semen Domba
Volume satu ejakulat semen domba berkisar antara 0.7-2 ml dengan
konsentrasi antara 2-5 juta sel/ μl (Mann 1964). Sedangkan menurut Toelihere
(1993), volumenya berkisar antara 0.5-2.5 ml dengan konsentrasi 1.5 juta – 3 juta
sel/ml semen dan 90% spermatozoa hidup. Semen dengan konsentrasi yang tinggi
bereaksi agak asam, sedangkan semen dengan konsentrasi rendah bereaksi agak
basa. Semen domba yang normal berwarna krem dan konsistensinya kental.
Gerakan massa dari spermatozoa dengan motilitas yang tinggi dapat diobservasi
dengan mata telanjang. Semen yang mengandung spermatozoa mati dalam
jumlah banyak akan berwarna kecoklat-coklatan. Konsentrasi yang rendah
menyebabkan warna semen seperti air susu atau lebih encer lagi (Terril 1964;
Toelihere 1993).
Spermatozoa terdiri atas kepala dan ekor (terdiri dari body dan tail) yang keseluruhannya diselubungi oleh membran plasma (Johnson et al. 1997). Pada bagian kepala terdapat inti sel dengan DNA-nya yang merupakan materi genetik
jantan dan akrosom dengan enzim-enzim hidrolitik yang diperlukan untuk
menembus dinding ovum pada saat fertilisasi. Akrosom tersebut berasal dari
aparatus golgi yang banyak mengandung enzim-enzim pencerna protein dan gula.
Sebaliknya, pada bagian ekor yang menghubungkan dengan kepala membentuk
pilinan mitokondria yang mengandung selubung berserat dan mengandung
mikrotubulus tunggal tanpa selubung (Gilbert 1988).
Komponen utama suatu inti sel, yaitu kromatin yang mengandung DNA
pada saat maturasi, sebagian besar organel pada sitoplasma spermatozoa akan
menghilang dan tinggal beberapa organel termodifikasi yang tetap melangsungkan
fungsi spermatozoa. Selain itu, inti haploid mengalami perubahan, sehingga
pilinan serat DNA kromatin mengalami pemadatan menjadi pendek dan tebal.
Sebutan kromatin berubah menjadi kromosom (badan berwarna) (Gilbert 1988).
Hafez (1987) menyatakan bahwa kromosom yang dikandung spermatozoa disebut
dengan kromosom kelamin. Kromosom spermatozoa dibagi menjadi dua bagian
yaitu kromosom X (spermatozoa X) dan kromosom Y (spermatozoa Y).
fertilisasi. Kromosom X disebut kromosom betina, sedangkan kromosom Y
disebut kromosom jantan.
Penentuan Jenis Kelamin
Penentuan jenis kelamin (sexing) atau prenatal sex determination didasari oleh teori genetika yaitu jenis kelamin pada suatu mahluk hidup ditentukan pada
saat fertilisasi. Hewan betina mempunyai dua seks kromosom yang sama (X dan
X), sedangkan hewan jantan mempunyai dua seks kromosom yang berbeda X dan
Y). Sel somatik diploid dari betina (homogametic) mengandung sepasang kromosom X, tetapi sel somatik jantan mempunyai seks kromosom XY. Genetik
sex ditentukan dalam oviduct pada saat fertilisasi dan seks dari keturunan
ditentukan oleh kromosom seks dalam spermatozoa.
Kromosom adalah unsur utama dalam penentuan jenis kelamin. Pada
mamalia terdapat 46 kromosom yang terdiri atas dua macam kromosom, yaitu XY
pada jantan dan XX pada betina. Hafez (1987) menyatakan bahwa kromosom
yang dikandung spermatozoa disebut dengan kromosom kelamin, yaitu kromosom
X (sperma X) membawa sifat-sifat kebetinaan dan kromosom Y (sperma Y)
membawa sifat-sifat kejantanan. Kromosom spermatozoa merupakan faktor
penentu pada jenis kelamin anak hasil fertilisasi setelah terjadi penggabungan
antara kromosom spermatozoa dan kromosom ovum. Oleh karena itu, jenis
kelamin anak ditentukan oleh kromosom spermatozoa mana yang berhasil
membuahi kromosom sel telur induk. Informasi genetik yang dibawa oleh sperma
diterjemahkan dan disimpan di dalam molekul DNA yang tersusun oleh
nukleotida-nukleotida. Pada waktu mengalami spermatogenesis, spermatozoa
hanya mengandung setengah jumlah DNA pada sel-sel somatik dari spesies yang
sama dan terbentuklah dua macam spermatozoa; spermatozoa yang membawa
kromosom X akan menghasilkan embrio betina, sedangkan yang membawa
kromoson Y akan menghasilkan embrio jantan.
Perbedaan antara spermatozoa Y dan X pada manusia, yaitu kandungan
DNA spermatozoa Y lebih rendah 2.78% daripada DNA spermatozoa X (Kaneko
X (Shettles 1970; Revay et al. 2004). Spermatozoa Y mempunyai kepala yang panjang, besar, leher dan ekornya lebih kecil daripada spermatozoa X (Cui 1977,
diacu dalam Jongbloet 2004).
Anonim (2006) menyatakan bahwa spermatozoa Y pada manusia secara
fisik mempunyai bobot yang ringan, pergerakannya cepat, waktu hidup pendek.
Kebalikannya, spermatozoa X mempunyai bobot yang lebih besar, pergerakan
lebih lamban, dan masa hidup lebih panjang. Lebih lanjut dijelaskan, apabila
sekresi vagina dan uterus bersuasana asam, maka anak yang dihasilkan akan
berjenis kelamin perempuan. Hal ini karena pada suasana asam spermatozoa Y
akan mati, sedangkan spermatozoa X lebih tahan pada suasana asam. Apabila
sekresi vagina dan uterus bersuasana netral atau basa, maka anak yang dihasilkan
akan berjenis kelamin laki-laki. Pada suasana netral atau basa, spermatozoa Y
gerakannya lebih cepat karena ukurannya kecil. Nurwansyah (2006) juga
menyatakan bahwa spermatozoa Y akan lebih kuat dan lebih tahan hidupnya
dalam suasana basa. Sebaliknya, spermatozoa X lebih kuat dan lebih tahan
hidupnya pada suasana asam.
Rasio kelamin keturunan dipengaruhi oleh hormon, pH vagina, makanan
(Budinurdjaja 2007), tingkah laku seksual, konsentrasi hormon, bencana alam,
polusi lingkungan, gangguan endokrin, genetik (Krackow 1997; Pergament et al. 2002), waktu inseminasi, laju koitus, dan bagian tempat ovulasi (James 2004).
Dijelaskan oleh James (2004) bahwa apabila ovulasi di bagian kanan, maka
anaknya akan laki-laki karena di bagian kanan sekresi estradiol dan testosteron
lebih banyak. Makanan yang tinggi lemak meningkatkan hormon steroid
termasuk testosteron sehingga anak lebih banyak jantan; infeksi parasit
(Heligmosomoides polygyrus) menurunkan level testosteron; dioxin menurunkan rasio testosteron/gonadotropin sehingga anaknya banyak perempuan. Rasio
kelamin jantan lebih besar daripada betina apabila perkawinan dilaksanakan di
akhir estrus pada hamster, pH darah lebih tinggi pada tikus, diberi suntikan
progesteron di hari ke-8 kebuntingan pada tikus, diberi gonadotropins pada waktu
dilakukan penggertakan ovulasi, saluran reproduksi betina mempunyai pH yang
lebih tinggi (pH vagina dan pH serviks), suhu tidak panas, viskositas cairan
Rosenfeld and Roberts (2004) menyatakan bahwa jenis kelamin dipengaruhi
oleh makanan dan jika diberi makanan yang mengandung zat lemak (60 kkal%),
karbohidrat (20 kkal%), maka peluang untuk mendapatkan anak jantan sekitar
0.51-0.71% (pada tikus). Sebaliknya, jika diberi makanan mengandung
karbohidrat (70 kkal%), lemak (10 kkal%), maka peluang untuk mendapatkan
anak betina < sekitar 0.35-0.48%. Menurut Wauters et al. (1995), induk kijang akan beranak jantan sekitar 75% jika diberi makanan yang tinggi kandungan
energinya, sedangkan frekuensi beranak jantan akan turun menjadi sekitar 46%
jika diberi makanan yang mengandung energi rendah. Selanjutnya, Tamimi et al.
(2003) menjelaskan bahwa jika seorang ibu sedang mengandung bayi laki-laki,
maka kebutuhan konsumsi energi meningkat 10% daripada mengandung bayi
perempuan. Menurut Papa et al. (1993), elektrolit dalam ransum induk mungkin mempengaruhi rasio seks keturunan pada manusia. Induk yang mengkonsumsi
ransum tinggi Na+ dan K+ tetapi rendah Ca2+ mempengaruhi rasio seks
turunannya. Penambahan NaCl dari 0.08 menjadi 4.00% akan menurunkan rasio
seks jantan dari 57+0.03 menjadi 44+0.03%, tetapi meningkatkan jumlah anak
yang dilahirkan sebesar 12.8+0.4 menjadi 13.8+0.6% (Bird and Contreras 1986).
Keseimbangan Kation-Anion
Keseimbangan asam-basa dalam organisme tetap terjaga. Keseimbangan
asam-basa menyangkut pertukaran ion H+ dan komponen-komponen media
internal yang mampu menyumbangkan atau menerima ion. Substansi yang
mampu menyumbangkan ion adalah asam, sedangkan yang mampu mengikat
hidrogen adalah basa. Faktor lain dalam pelepasan ion H+ untuk menjaga
keseimbangan asam-basa dengan cara pembentukan amonia yang intensif dalam
organisme dan pemasukan sejumlah besar garam-garam potasium. Cairan
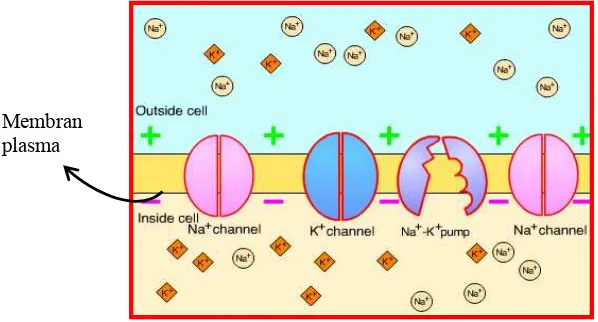
intraseluler mengandung ion kation sodium (Na+) dan anion klorida (Cl-) dalam
jumlah yang sedikit, tetapi banyak mengandung ion potasium (K+) dan anion
organik alami yang tidak bisa melewati membran. Proporsi ion individu dijaga
dengan mekanisme yang disebut pompa Na, ion Na+ dipompa keluar dari sel. Pompa ini mencegah perpindahan air yang berlebihan masuk ke dalam sel.
internal membran menjadi negatif. Pada waktu yang sama pompa memaksa ion
K+ masuk ke dalam sel, sehingga konsentrasi K+ lebih tinggi dari sekitarnya.
Transfer aktif ion Na+ dan K+ terjadi di dalam eritrosit (berhubungan
dengan fungsi pernapasan), di dalam epitel saluran ginjal, dan dalam membran
organel tertentu. Ion K+ dalam sel darah merah mempengaruhi transportasi O2
dan CO2 oleh haemoglobin. Mineral K juga sepertinya berhubungan langsung
dengan proses pembentukan protein. Jika ditambahkan K ke dalam ransum yang
kekurangan protein, maka akan menambahkan bobot tubuh anak dan memperbaiki
kegunaan makanan. Penetapan mekanisme, terutama homeostasis K dalam tubuh
adalah pada ginjal. Regulasinya meliputi mineral corticoids-aldosterone dan
deoxycorticosterone. Hormon-hormon ini berpengaruh pada pengeluaran K+ yang merupakan pengaruh kemungkinan kedua, karena kehadiran hormon tersebut
akibat adanya penyerapan kembali ion Na+ dalam saluran-saluran ginjal. Namun
demikian, kedua proses tersebut saling berhubungan karena hormon aldosteron
dipicu/didorong hanya jika level Na+ dalam plasma menurun dan level K+
meningkat secara bersamaan. Pengeluaran kelebihan ion K+ melalui ginjal dengan
mekanisme regulasi mempertahankan rasio Na:K agar konstan dalam cairan
ekstraseluler.
Yingst et al. (2001) menyatakan bahwa peningkatan konsentrasi K dari 4 menjadi 10 mM pada tikus mengakibatkan peningkatan aktivitas pump-Na dan konsentrasi ion Ca2+ bebas dalam darah. Kemungkinannya, K meningkatkan
pump Na dalam rangka meningkatkan konsentrasi Ca, sehingga meningkatkan pula Ca2+ bebas dalam darah yang ditingkatkan oleh pump-Na dalam beberapa sel. Dijelaskan juga oleh Burnay et al. (1994) dan Capponi et al. (1987) bahwa penambahan konsentrasi K memungkinkan untuk meningkatkan sekresi
aldosterone oleh depolarisasi membran yang membuka saluran tegangan Ca serta untuk membiarkan masuknya Ca ekstraseluler. Pada bagian lain, polarisasi
mempengaruhi konsentrasi K untuk diturunkan dalam rangka melakukan
keseimbangan K (Quinn et al. 1987). Walaupun penambahan konsentrasi K yang berlebihan akan meningkatkan sekresi aldoseteron, juga akan meningkatkan pump Na oleh kehadiran peningkatan jumlah konsentrasi K yang menyelimuti pump