PENGGUNAAN ZEOLIT BERAMONIUM DAN
ANALOG HIDROKSI METIONIN DALAM
RANSUM SAP1 PERAH LAKTASI
oleh
NUR-AENI SIGIT
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
RINGKASAN
NUR-AENI
SIGIT'
Penggunaan Zeolit Beramonium dan Analog Hidroksi
Metionin dalam Ransum Sapi Perah Laktasi (di bawah bimbingan Adi Sudono
(ketua), Toha Sutardi, Djokowoerjo Sastradipradja, Mirnawati Sudarwanto,,dan
Wasmen Manalu).
Persediaan pakan dipengaruhi kondisi dan fluktuasi hijauan. Lahan makin
'
menyempit menyebabkan ketersediaan hijauan terbatas. Proporsi hijauan menyusut
dan
konsentrat meningkat dalam ransum dapat menurunkan kadar lemak susu.
Protein dalam konsentrat umumnya tahan degradasi,
sehingga sering tidak
dapat memenuhi kebutuhan
N
untuk mikroba. Urea biasa digunakan sebagai sumber
N,
tetapi karena di dalam rumen cepat dirombak menjadi amonia maka penggunaannya
hams dilengkapi sumber karbohidrat mudah difermentasikan.
Zeolit adalah kristal aluminosilikat berstruktur tiga dimensi, berongga,
dan
dapat berperan sebagai penukar kation. Dengan teknologi amoniasi
dan
tukar kation,
zeolit dapat diisi oleh ion amonium menjadi zeolit beramonium
(Z-NH4).
Di dalam
rumen ion amonium zeolit akan dilepaskan secara bertahap sesuai dengan ketersediaan
ion penukar yang berasal
dari
saliva sehingga tidak
terjadi akumulasi amonia.
Metionin dapat meningkatkan lemak susu,
akan
tetapi metionin merupakan asam
amino pembatas pertama pada pakan. Metionin di dalam rumen ternyata mudah
didegradasi. Melalui proses transarninasi, kekurangan metionin kemungkinan dapat
dipenuhi oleh analog hidroksi metionin (AHM) yang lebih tahan degradasi.
Penelitian bertujuan mengkaji manfaat
Z-NH4
dan
AHM serta
kombinasinya
terhadap konsumsi pakan, metabolisme di rumen, kondisi tubuh, produksi dan
komposisi susu sapi perah, menduga produksi panas tubuh serta mengkaji manfaat
penggunaan zeolit terhadap kualitas susu. Penggunaan zeolit b e r a n o n i u m ~ ~ . ~ a n a l o g
hidroksi metionin diharapkan dapat memperbaiki komposisi dan meningkatkan mutu
susu.
Percobaan dilaksanakan di PT Baru Adjak Bandung, PT Kariyana Gita Uiama
Sukabumi, Laboratorium Ilmu Nutrisi Fakultas Peternakan, Laboratorium Fisiologi dan
Laboratorium Kesehatan Veteriner Fakultas Kedokteran Hewan, IPB. 2-NH4 dibuat
dengan merendam zeolit di dalam larutan NH40H 1N selama 1 minggu. AHM yang
digunakan buatan Sunco Amerika Serikat.
Percobaan disusun dalam rancangan acak kelompok pola faktorial 3 x 3. Faktor pertama 3 taraf 2-NH4, yaitu 056, 1.5 % dan 3.0% bahan kering konsentrat atau 0, 150 dan 300 g/ekor/hari. Faktor kedua taraf AHM, yaitu 0 % , 0.1 % dan 0.2% atau 0, 10 dan 20 glekorlhari. Perlakuan diulang 3 kali (sebagai kelompok). Ransum terdiri atas
konsentrat
dan
rumput Gajah dengan nisbah 75:25.Percobaan in M'vo dilakukan pada 27 ekor sapi perah awal laktasi. Peubah yang diukur adalah perubahan bobot badan, konsumsi zat makanan, produksi dan komposisi
susu, dan nilai dugaan produksi panas. Penentuan koefisien cerna dilakukan secara i n vitro. Konsentrasi N-amonia diukur dengan teknik mikrodifusi Conway dan konsentrasi
asam lemak atsiri (VFA) dengan penyuIingan uap. Sintesis protein mikroba diukur
dengan teknik radioisotop, menggunakan 3 2 ~ sebagai perunut. Percobaan in sit*, dengan teknik kantong nilon, mengukur degradasi bahan kering dan bahan organik.
Percobaan pengamatan h a l i t a s air susu menguji 4 perlakuan, yaitu kontrol,
"teat dipping", zeolit dan "teat dipping"
+
zeolit. Perhitungan jumlah bakteri dalarn susu menggunakan metode cacah koloni setelah pembiakan pada media agar dalam cawan. Penghitungan dilakukan untuk cawan yang mengandung 25 - 250 koloni.Z-NH4 1.5 dan 3.0% atau AHM 10 clan 20 g/ekor/hari tidak mengakibatkan
bahan kering dan bahan organik tidak dipengaruhi oleh 2-NH4 atau oleh AHM maupun interaksinya. Baik 2-NH4 maupun AHM serta interaksinya tidak mempengaruhi degradasi ransum dalam kantong nilon.
2-NH4 meningkatkan produksi N-NH3 secara Iinier (P < 0.06). Produksi Nz NH3 yang dihasitkan oleh ransum ZO (kontrol), 21 dan 2 2 masing-masing adalah 5.46,
5.76 clan 6.07 mM. Penarnbahan AHM tidak mengubah produksi N-amonia. Produksi
VFA tidak berubah baik oleh 2-NH4 rnaupun AHM. Produksi VFA ransum yang mengandung 2-NH4 20, Z1 dan 22 masing-masing 59.48, 62.07 dan 64.14 mM.
Produksi VFA untuk AO, A1 dan A2 masing-masing 65.69, 62.58 dan 57.4 1 mM. Sintesis protein mikroba sangat nyata dipengaruhi oleh 2-NH4 (P
<
0.01)tetapi tidak dipengaruhi oleh AHM maupun interaksinya. Z N H 4 meningkatkan sintesis protein mikroba secara linier dengan persarnaan Y = 176.22
+
0.330 X(Y = Protein mikroba, X = Taraf Z-NH4). Protein mikroba yang disintesis oleh penambahan 2-NH4 150 dan 300 gram masing-masing ialah 252.8 dan 261.8 mg/l/jam.
2-NH4 dan AHM maupun kombinasinya tidak mengakibatkan produksi susu
dan produksi susu 4 % FCM berbeda. Produksi susu rataan hasil percobaan ini sebesar
16.61
k
0.42 kg/ekor/hari.Z-NH4 dan AHM sangat nyata ( P t 0.01) mempengaruhi kadar lemak susu, namun interaksinya tidak nyata. Antara ZO dan 2 1 kadar lemak susu naik sebesar 0.25 persen atau 5.81 persen unit, tetapi antara 21 dan 2 2 turun sebesar 0.52 persen atau
Terjadi perubahan kadar protein susu oleh AHM maupun interaksinya dengan
Z-NH4. 2-NH4 tidak nyata pengaruhnya terhadap kadar protein susu (%) d m produksi
protein susu (kglhari). AHM dapat meningkatkan kadar protein susu (PC0.05). Kadar
protein susu pada pemberian 10 dan 20 gram AHM meningkat masing-masing sebesar
0.42 dan 0.14 persen atau 10.91 dan 3.64 persen unit.
Interaksi antara Z-NH4 dan AHM dengan nyata (P<0.05) mempengaruhi kadar
protein susu. Kombinasi pemberian zeolit kramonium dengan AHM yang terbaik untuk kadar protein susu terjadi pada dosis Z-NH4 dan AHM masing-masing 300 dan
10 g/ekor/hari.
Kadar laktosa susu yang diperoieh lebih rendah dari kisaran normal, yaitu hanya
sekitar 2.66 - 3.00%. Penambahan Z-NH4 dan AHM ke dalam ransum sapi perah pada awal laktasi tidak mempengaruhi berat jenis susu.
Z-NH4 meningkatkan kadar bahan kering susu ( P t 0 . 0 5 ) dan nilai tertinggi
dicapai pada 2-NH4 150 g/ekor/hari (Zl). AHM meningkatkan bahan kering susu
(P
<
0.05), bahan kering tertinggi dihasilkan oleh pemberian 10 g/ekor/hari. Bahankering tanpa lernak (BKTL) tidak berubah oleh 2-NH4 maupun AHM.
Selama percobaan, sapi mengalami penyusutan bobot badan sebesar 0.275 f 0.093 kghari. Pemberian 2-NH4 dapat menekan penyusutan bobot badan sapi perah
pada awal laktasi (P c0.05). AHM tidak mempengaruhi perubahan bobot tubuh sapi.
Produksi panas tidak dipengaruhi oleh 2-NH4, AHM maupun kombinasinya.
Produksi panas akibat penambahan zeolit beramonium ZO, Z1 dan 2 2 masing-masing
68.24, 56.76 dan 50.66 MJIhari. Produksi panas karena penambahan AHM yaitu AO,
A l , A2 masing-masing 53.68, 62.11 dan 59.87 MJIhari. Hasil perhitungan produksi
B B O - ~ ~ pa& perlakuan 20, Z1 dan 22, sedangkan AO, A1 dan A2 menghasilkan panas t u b u h / B ~ O - ~ ~ masing-masing 0.63, 0.72 dan 0.70.
Efek penggunaan "teat dipping" dengan desinfektan, zeolit, maupun
kombinasinya terhadap penurunan jumlah bakteri susu tidak nyata. Tetapi pada umumnya jumlah bakteri susu menurun setelah satu bulan. Pada dua minggu pertama sapi yang mengalami "teat dipping" dengan desinfektan dan sapi yang mendapat zeolit
sudah memperlihatkan penurunan jumlah bakteri masing-masing sebesar 86.4 clan 43.5
persen. Pa& sapi yang mendapat kombinasi desinfektan dan zeolit penurunan jumlah bakteri sebesar 86.57 persen terjadi setelah empat minggu, sedangkan penurunan
PENGGUNAAN ZEOLIT BERAMONIUM DAN
ANALOG HIDROKSI METIONIN DALAM
RANSUM SAP1 PERAH LAKTASI
Oleh
NUR-AENI
SIGIT
Disertasi sebagai salah satu syarat
untuk memperoleh gelar doktor pada Program Pascasarjana
Institut Pertanian Bogor
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Penelitian : Penggunaan Zeolit Beramonium dan Analog Hidroksi Metionin &am Ransum Sapi Perah Laktasi
Nama Mahasi+swa : Nur-Aeni Sigit
Nomor Pokok : 94562
Ketua
n
Prof. Dr. D. SastradiDradiaAnggota pAnggOb /
Dr. Drh. H. Mimawati Sudarwanto Dr. Ir. Wasmen Manalu M.Sc
J'wzgota Anggota
RIWAYAT HIDUP
Penulis dilahirkan di Bogor pada tanggal 2 Januari 1944, anak pertama dari
0. Abdurachman Rafiudin dan U. Kalsum. Penulis menikah dengan D. Sigit dan
berpum tiga orang yaitu Aldy Nugraha, Meigi Melissa dan Rachmat Febriandi.
Penulis menyelesaikan Sekolah Rakyat, Sekolah Menengah Pertama
dan
SekolahMenengah Atas di Bogor pada tahun 1956, 1959 dan 1962. Penulis menyelesaikan Sarjana
Peternakan Institut Pertanian Bogor tafiun 1969 dan Magister Sains Institut Pertanian Bogor
tahun 1982, enam tahun kemudian penulis mandapat kesempatan untuk mengikuti program S3 di Institut Pertanian Bogor. Sejak lulus S1 penulis bekerja sebagai staf pengajar di Fakultas
KATA
PENGANTARSegala puji syulcur dipersembahkm ke hadiirat AUah S.W.T. yang telah
melimpahkan rachrnat dan nikmatnya kepada penulis, sehingga disertasi ini dapat
diselesaikan, karena tiada satu usahapun akan berhasil tanpa perkenanNya.
Karya tulis ini sangat tidak berarti tanpa petunjuk dan pengarahan para
pembmbmg yang dengan gigih mendorong, menempa d m mongobarkan semangat serta
membina kepercayaan diri penulis. Dengan rasa hormat penulis mengucapkan t e r b m h s i i dan penghargaan yang setulus-tulusnya kepada komisi pernbimbing:
Prof. Dr. Drh. Adi Sudono, M.Sc. (Ketua)
Prof Dr. Drh. Toha Sutardi, M.Sc. (anggota)
Prof Dr. Drh. Djokowoeqo Sastradipradja (anggota)
Dr. Drh. Hj. Mimawati Sudarwanto (anggota)
Dr. Ir. Wasrnen Manalu, M.Sc. (anggota)
Kepada pimpinan fakultas Peternakan IPB, pimpinan Institut Pertanian Bogor, pirnpinan Program Pascasarjana beserta staf d m Tim Manajernen Program Doktor, penulis
mengucapkan terimakasih atas kesempatan yang diberikan untuk menernpuh pendidikan S3.
Penulis mengucapkan terima kasih kepada pimpinan serta karyawan PT. Baru
Adjak, Lembang Bandung dan pirnpinan serta karyawan PT. Kariyana Gita Utama, Cicurug Sukabumi atas segala fasilitas yang digunakan penulis selama penelitian. Terimakasih atas sumbangan zeolit dari C.V. Wonder Pharmatheucal Indonesia. Penulis juga mengucapkan terimakasih kepada ketua dan staf lab. Dinas Petemakan Kodya
Bandung, GKSI komda Bandung, lab. Ilrnu Nutrisi Fakultas Peternakan, lab. Fisiologi dan
lab. Kesehatan Masyarakat Veteriner Fakultas Kedokteran Hewan, IPB atas bantuannya
selama penelitian.
Sebagai seorang isteri dan ibu dari anak-anak, penulis telah banyak menyita waktu
dan perhatian mereka selama mengikuti program S3. Kepada suami dan anak-anaku
tercinta penulis ingin mempersembahkan karya tulis ini sebagai bekal pengalaman di rnasa
yang pada masa hidupnya selalu mendampingi penulis ddam suka dan duka. Kepada ayah
dan bibi yang menggantikan kehilangan tersebut, dengan rasa hormat penulis
mengucapkan terimakasih.
Kepada Dr. Drh. Lily Amalia Sofyan, M.Sc., Dr. Ir. Erwanto dan semua pihak yang telah membantu penulis selama persiapan, pelaksanaan clan penyelesaian karya ilmiah ini, penulis mengucapkan terimakasih. Semoga amal dan budi baik mereka rnendapat imbalam yang sepadan dari Allah S.W.T.
Akhir kata penulis berharap semoga sumbangan yang sederhana ini dapat dmanfaatkan untuk pengembangan ilmu pengetahuan. Penulis sangat menyadari
kelemahan dan kekurangan karya ilmiah
ini.
Bogor, 28 September 1995
DAFTAR IS1
RIWAY AT HIDUP KATA PENGANTAR
DAFTAR IS1 DAFTAR TABEL
DAFTAR GAMBAR PENDAHULUAN
TINJAUAN PUSTAKA
Produksi Susu dan Kebutuhan Nutrisi
Sapi Awal Laktasi
Komposisi dan Kualitas Susu
Metabolisme Protein dan Energi
Zeolit dan Penggunaannya dalam Ransum Metionin dan Analog Hidroksi Metionin d a m Ransum Sapi Perah Laktasi
BAHAN DAN CARA PERCOBAAN
Penelitian I
Penelitian I1
HASIL DAN PEMBAHASAN
Konsumsi Ransum dan Zat-zat Makanan Kecemaan dan Hasil Metabolisme di Rumen Produksi clan Komposisi Susu
DAFTAR TABEL
Nomor Judul Tabel
1 . Kapasitas Tukar Kation Beberapa Zeolit A l m Indonesia
2 . Efek Zeolit Beramonium terhadap Kadar N-NH3, pH dan Sintesis Mikroba Rumen
3. Komposisi Asam Amino Protein Pakan, Mikroba Rumen, Digesta Abomasum dan Susu
4. Efek Carnpuran Asam Amino terhadap Kandungan Protein Susu
5 . Mum Protein Beberapa Limbah Tanaman
6 . Efek Infusi Metionin ke dalam Abomasum terhadap .Produksi dan Komposisi Susu
7 . Kombinasi Perlakuan Zeolit Beramonium dan Analog Hidroksi Metionin
8. Komposisi Ransum Percobaan
9. Konsumsi Zat Makanan dan Ransum Percobaan 10. Kecernaan dan Hasil Metabolisme di Rumen
11. Kecernaan dan Hasil Metabolisme di Rumen pada Taraf ikolit Beramonium dan Analog Hidroksi Metionin Berbeda
12. Sintesis Protein Mikroba Rumen
13. Produksi
dan
Komposisi Susu14. Produksi dan Komposisi Susu pada Taraf Zeolit
Beramonium
dan
Analog Hidroksi Metionin yang Berbeda15. Efek Taraf -lit Beramonium &in Analog Hidroksi Metionin pa& Kadar Lemak Susu
16. Efek Taraf Zeolit Beramonium dan Analog Hidroksj Metionin pada Kadar Protein Susu
17. Efisiensi Penggunaan Ransum, Protein dan Energi 18. Efek Penambahan Zeolit Beramonium dan Analog
DAFTARTABEL (mj utan)
Nomor Judul Tabel
19. Nitrogen Urin, Retensi Nitrogen
dan
Katabolisme Protein pada Taraf Zeolit Berarnonium dan Analog Hidroksi Metionin yang Berbeda20. Pembahan Bobot Badan, Retensi Karboil dan Produksi Panas pada Taraf Zeolit Beramonium dan Analog Hidroksi Metionin yang Berbeda
DAFTAR GAMBAR
Nomor Judul Gambar
1. Metabolisme MET, CYSH dan CYS
2. Formula Analog Hidroksi Metionin dan L-Metionin
3. Konversi Analog Alfa Hidroksi Metionin Menjadi L-Metionin
4. Histogram antara Kadar Asam Lemak Atsiri clan Arnonia dengan Protein Mikroba Rumen pada PerIakuan Zeolit Beramonium
5 . Histogram Interaksi Zeolit B e m n i u m dengan Analog =droksi Metionin pa& Kadar Protein Susu
6 . Efek Zeolit Beramonium terhadap Komposisi Susu
7. Efek Analog Hidroksi Metionin terhadap Komposisi Susu
PENDAHULUAN
Issu peningkatan sumber daya manusia yang berkualitas berkembang menjelang
akhir Pelita
V.
Peningkatan kualitas manusia perlu didukung oleh pangan yang cukup,bermutu dan aman. Produk peternakan seperti dagi~ig, susu dan telur merupakan
pangan yang bermutu tinggi tetapi keamanan produk tersebut terutama susu bagi
konsumen belum sepenuhnya te jamin.
Peternakan sapi perah terus dikembangkan sesuai dengan tujuan Pembangunan
Nasional untuk mencapai target pemenuhan gizi dari protein hewani. Melalui berbagai
program, sejak awal Pelita 111 sampai pertengahan Pelita V, populasi sapi perah
ditingkatkan dari 125 000 ekor menjadi 325 000 ekor, namun produksi susu dalam negeri hanya rnarnpu menyediakan separuh dari permintaan susu yang terus meningkat.
Pada kurun waktu yang sama produksi susu meningkat dari 25 000 ton menjadi 306
000 ton sedangkan konsumsi meningkat dari 499 000 ton menjadi 795 000 ton, sehingga untuk memenuhi kebutuhan pada tahun 1992 masih diperlukan impor susu
sebesar 5 14 000 ton (Suhadji, 1994).
Kenyataan bahwa produksi susu rata-rata sapi perah di Indonesia masih rendah,
diduga disebabkan oleh manajemen yang kurang baik. Perbailcan rnanajemen dapat
diupayakan melalui pendekatan agribisnis yang a n m lain mencakup perbailcan pakan.
Pakan sapi perah tidak terlepas dari kondisi dan fluktuasi hijauan. Pada kondisi lahan yang makin menyempit porsi hijauan dalam ransum yang umumnya tinggi perlu
dipertimbangkan. Penambahan porsi konsentrat merupakan salah satu cara penanggulangan masalah tersebut. Peningkatan proporsi konsentrat terhadap hijauan
akan mengakibatkan perubahan ekosistem di dalam rumen dan penurunan kadar lemak
susu. Padahal kadar lemak susu m e ~ p a k a n salah satu faktor penentu harga susu. Peningkatan porsi konsentrat dalam pakan sapi perah lebih ditujukan kepada kecukupan
Strategi pemberian pakan sumber protein pada ruminansia mencakup tiga hal
yaitu memaksimalkan sintesis protein mikroba, meningkatkan pasokan protein 1010s
degradasi yang mudah dicerna di usus dan melengkapi asam amino esensial. Sumber nitrogen untuk mikroba rumen yang paling murah dan sering digunakan adalah urea.
Namun pemberian urea selain memerlukan surnber karbohidrat yang mudah
diferrnentasikan juga dapat mengakibatkan keracunan pa& ternak. Urea sangat cepat
dirombak menjadi amonia dan akumulasi amonia sering mengganggu ekosistem di
dalam rumen. Agar penggunaan urea atau nitrogen bukan protein
(NPN)
lain, optimum maka pelepasan amonia hams diperlambat sesuai dengan pelepasan energi darikarbohidrat dan sesuai dengan kemampuan sintesis protein mikroba.
Zeolit adalah mineral d a m berstruktur tiga dimensi dan berpori, yang mempunyai sifat sebagai penukar kation, sebagai bufer dan mempunyai daya serap yang tinggi. Meldui proses penjenuhan dengan ion amonium (amoniasi), zeolit
diharapkan dapat dimanfaatkan sebagai pemasok nitrogen yang dapat memperlambat
pelepasan amonia. Di dalam rumen ion amonium akan ditukarkan secara bertahap dengan ion Na+ dan K + yang berasal dari saliva. Penggunaan -lit beramonium diharapkan menjadi lebih bermakna karena selain sebagai pemasok N bagi mikroba juga
sebagai pengendali pH rumen.
Metionin mempakan asam amino pembatas pertama pada sebagian besar bahan
pakan penyusun ransum sapi perah yang pada umumnya terdiri atas bungkil-bungkilan.
Metionin juga rnempakan pembatas pertama pada sapi perah awal laktasi. Padahal
menurut beberapa laporan metionin berperan dalam peningkatan kadar lemak susu yang
biasanya rendah pa& saat awal laktasi. Kecuali mahal harganya, metionin di dalam
rumen lebih mudah didegradasikan sehingga penambahan asam amino tersebut &lam
dapat dipenuhi oIeh analog hidroksi metionin (AHM) yang lebih murah dan lebih tahan degradasi di dalam rumen. Melalui proses transaminasi oleh enzim transaminase di
dalam tubuh ternak, analog hidroksi metionin diubah menjadi metionin sehingga
mungkin dapat menutupi kekurangan metionin terutama pada bungkil-bungkilan.
Penelitian ini bertujuan untuk mempelajari manfaat zeolit beramonium dan analog hidroksi metionin maupun kombinasinya baik terhadap metabolisme cli dalam
rumen dan kondisi tubuh, maupun terhadap produksi dan komposisi susu sapi perah serta mengkaji pengaruh penggunaan zeolit terhadap kualitas susu yang dinyatakan
sebagai jumlah bakteri dalam susu segar. Penggunaan zeolit beramonium dan analog
hidroksi metionin diharapkan dapat memperbaiki komposisi dan kualitas serta
TINJAUAN
PUSTAKA
Produksi Susu dan Kebutuhan Nutrisi Sapi AwaI Laktasi
Produksi Susu
Pada awal laktasi produksi susu meningkat dengan cepat menjadi 50-80% dari
produksi maksimum dan mencapai puncak laktasi pada minggu ketiga sampai kedelapan
setelah beranak (Holmes dan Wilson, 1984; NRC. 1988). Produksi susu sapi perah di Indonesia selama dua bulan pertama setelah beranak sekitar 145 persen dari produksi rata-rata masa laktasi tahun bersangkutan, selanjutnya menurun menjadi 120 persen pada bulan ketiga sarnpai keempat dan mencapai rata-rata pada bulan kelima sampai
keenam. Produksi susu pada bulan ketujuh sampai kedelapan sebesar 78 persen dan menjelang beranak produksi susu kurang dari 70 persen dari produksi rata-rata (Sutardi, 1982). Sementara itu selarna tiga minggu setelah beranak selera makan sapi perah rendah dan konsumsi menurun 15-18% (Garnsworthy, 1988;
NRC,
1988), kemudian konsumsi meningkat dan mencapai puncak pada 2-3 bulan setelah beranak (NRC, 1988: Holmes dan Wilson, 1984). Pada saat itu konsumsi sapi laktasi 30-60% lebih tinggi dibandingkan dengan sapi tidak laktasi (Holmes dan Wilson, 1984).Peningkatan produksi susu yang tidak diimbangi oleh peningkatan konsumsi
pakan pada awal laktasi mengakibatkan kesenjangan pasokan zat nutrisi sehingga te jadi
mobilisasi cadangan zat makanan dari tubuh. MobiIisasi cadangan tubuh juga
disebabkan oleh kecepatan peningkatan konsumsi pada awal laktasi lebih lambat
daripada kecepatan peningkatan produksi susu, karena puncak produksi (7-8 minggu) dicapai lebih dahulu daripada puncak konsumsi (16 minggu setelah beranak).
yang kondisi tubuhnya baik (gemuk) akan mengalami penyusutan bobot tubuh yang
lebih besar dibandingkan dengan sapi yang lebih kurus. Perlemakan tubuh mengurangi
rasa lapar, dengan demikian makin gemuk sapi waktu beranak makin rend& nafsu
makannya. Dengan lain perkataan makin gemuk sapi pada waktu beranak makin besar
jarak antara puncak produksi dengan puncak konsumsi (Sutardi, 1980; Garnsworthy,
1988).
Pada awal laktasi mobilisasi protein dari jaringan tubuh sapi perah berproduksi
tinggi (30-40 kg/hari) dapat mencapai 150-300 glhari (Garnsworthy, 1988) atau 15 kg
selama dua bulan setelah beranak (NRC, 1988). Sementara itu lemak yang terkuras dari
depot lemak sebesar satu kglhari (Garnsworthy,l988). Menurut Holmes dan Wilson (1984) pada awal laktasi sapi perah bisa kehilangan bob- satu kg/hari.
Pada puncak Laktasi kadar protein, lemak dan bahan kering susu tanpa lemak mencapai titik terendah, selanjutnya pada bulan ketiga sampai keenam komposisi susu
akan stabil dan produksi susu mencapai rata-rata masa laktasi (Sutardi, 1982). Dikaitkan dengan selera makan sapi perah yang rendah pada awal laktasi, Garnsworthy
(1984) menyarankan pemberian ransum yang kaya
akan
propionat dan ransum seperti ini hanya dapat dicapai dengan meningkatkan porsi konsentrat dan menurunkan porsihijauan di dalarn ransum. Peningkatan proporsi konsentrat akan menyebabkan pH
rumen dan laju aliran digesta menurun. Kondisi seperti ini tidak menguntungkan bagi
bakteri pencerna selulosa karena dapat mengakibatkan produksi asetat sebagai prekursor
lemak susu berkurang sehingga kadar Iemak susu menurun (NRC,1988). Menurut
Holmes clan Wilson (1984) peningkatan produksi ]em& susu pada awal laktasi bukan disebabkan oIeh peningkatan kadar (persen) Iemak tetapi disebabkan oleh produksi susu
yang meningkat. Di Indonesia faktor penentu harga susu tidak didasarkan kepada
Kebutuhan Nutnsl
. .
Komponen yang paling penting harus cukup dalam ransum adalah energi, Kekurangan energi a s d karbohidrat akan mengakibatkan perombakan zat organik
lainnya menjadi energi sehingga keefisienannya akan berkurang. Energi juga
merupakan kunci utama untuk produksi susu. Kekurangan energi akan menurunkan
produksi susu, walaupun nutrien laimya cukup. Energi pada ruminansia tidak
bersumber pada giukosa tetapi pada asam l e d terbang yang diproduksi di dalam rumen. Glukosa darah ruminansia selalu rendah tetapi kebutuhan glukosa meningkat
tiga kali lipat pada sapi laktasi. Delapan puluh persen glukosa darah digunakan untuk
sintesis laktosa (Preston
dan
Leng, 1987; Holmes dan Wilson, 1984).Kebutuhan glukosa clipenuhi melalui perombakan glikogen clan melalui
pembentukan glukosa (glukonwgenesis) temtama dari propionat dan
asam
amino.Peningkatan giukonwgenesis melibatkan enzim-enzim piruvat karboksilase,
fosfoenolpiruvat karboksilase, fruktosa difosfatase
dan
glukosa-6-fosfatase (Vernon,1988). Peningkatan glukoneogenesis juga memerlukan suplai prekursor glukonwgenik
yang biasanya terjadi dengan peningkatan penyerapan propionat dan asam amino,
pelepasan asam laktat dan asam amino dari otot, dan gliserol dari jaringan deposit
(Vernon, 1988).
Asarn amino protein pakan kecuali dibutuhkan untuk sintesis protein susu juga diperlukan untuk sintesis laktosa melalui proses glukonwgenesis sedangkan laktosa
adalah komponen susu yang erat kaitannya dengan produksi sum. Protein tidak dapat
digantikan oleh zat lain dan di dalam tubuh selalu mengalami "turn over" sehingga tidak
tersedia dalam bentuk cadangan seperti lemak. Sintesis protein dari asam amino juga
mahal, membutuhkan energi 50 persen lebih tinggi dibandingkan dengan sintesis
akan di sintesis maka asam amino yang bukan pembatas akan dideaminasi dan
rnenghasilkan urea. Pembentukan dan pengeluaran urea sebagai ampas katabolisme
protein ke dalam urin memerlukan energi (Holmes dan Wilson, 1984) yang terbuang
sebagai panas.
Komposisi dan Kualitas Susu
Komposisi (susunan) susu dibedakan dengan kualitas susu dalam ha1 komponen yang terdapat di dalam susu dengan sifat susu. Komponen yang menyusun susu seperti
umumnya terdapat dalam bahan pangan lainnya terdiri atas air, protein, l e d ,
karbohidrat (laktosa = gula susu), mineral clan vitamin. Semua padatan yang terdapat
dalam susu yang meliputi bahan organik dan mineral, bakteri, leukosit
dan
sel-selsekretori kecuali lemak, dikelompokkan ke dalam bahan kering tanpa lemak (BKTL).
Kuaiitas susu lebih ditekankan kepada sifat susu antara lain seperti kebersihan,
warna, bau, rasa serta jumlah bakteri yang terdapat di dalamnya (Sudono, 1985). Menurut beberapa pengamat betapapun bersihnya susu tetap mengandung sejumlah
bakteri, karena susu merupakan media yang sangat baik untuk perkembangan mikroorganisme.
f(omposisi
S m
Komposisi susu relatif tetap kecuali kadar lemak. Beberapa komponen seperti asam lernak rantai pendek (C,-C,J, fraksi protein dan laktosa, disintesis di kelenjar ambing. Laktosa berasal dari glukosa, protein susu sebagian disintesis di ambing dari
asam amino dan sebagian lagi ditransfer langsung dari darah. Asam lemak rantai pendek berasal dari asetat dan beta-hidroksi butirat yang diproduksi di rumen sedangkan asam lemak rantai panjang, mineral dan vitamin ditransfer langsung dari
Secara fisik susu terdapat dalam tiga fase (Schalm ef ad., 1971; Foley m al,
1972) yaitu:
1. Fase larut yang terdiri atas air, sebagian protein (albumin dan globulin), karbohidrat
(laktosa), sebagian besar mineral dan vitamin yang larut.
2. Fase koloid, termasuk ke dalam fase ini yaitu kasein (bagian terbesar dari protein
susu), laktalbumin, laktoglobulin dan sebagian besar kalsiumfosfat. Ukuran koloid
sangat halus sehingga stabil dalam bentuk suspensi.
3. Fase emulsi merupakan butiran agak besar ymg tersebar dan tidak larut dalam air. Setiap partikel diselaputi oleh membran fosfolipid dan protein yang menjamin dalam
bentuk emulsi.
Lemak Susu. Sebagian besar lernak susu adalah lemak jenuh. Lebih dari 98%
lemak susu yang disintesis di ambing ada dalam bentuk trigliserida dan hanya sebagian
kecil berupa fosfolipid. Sekitar 50 persen lemak susu berasal dari asam lemak rantai
pendek (C4-CI4) yang disintesis di kelenjar ambing dari asam asem dan O-hidroksi butirat yang dihasilkan dari fermentasi selulosa di rumen. Sebagian lagi berasal dari
asam lemak rantai panjang (CI6-C18) dari rnakanan dan lemak cadangan tubuh (Holmes
dan Wilson, 1984). Asam lemak rantai panjang dari bahan rnahanan nabati umumnya tidak jenuh clan sebagian akan mengalami hidrogenasi di rumen. Lemak tubuh akan disintesis menjadi trigliserida di hati sebagai pul trigeliserida yang akan mensuplai
trigliserida ke ambing bila kurang dalam ransum. Penambahan lemak ransum meningkatkan asam lemak rantai panjang (C,,.d) tetapi menekan asan lemak rantai
Protein.
Kasein merupakan protein utama dan terbanyak (90% dari proteinsusu) dalam susu. ALfa-kasein, beta-kasein, kapa-kasein, a-laktalbumin clan 8-
taktoglobulin (90-95% dari protein susu) disintesis di dalam ambing dari asam amino
esensial dan asam amino non esensial yang berasal dari pool asam amino di &am darah. Immunoglobulin, serum albumin dan gama-kasein (5-10%) langsung ditransfer
dari darah (Foley et al., 1972; MacRae, 1988).
Protein susu dikelompokkan menjadi kasein dan serum susu (whey). Tennasuk
ke dalam whey adalah 6-laktoglobulin yang merupakan bagian terbesar dari whey,
a-laktalbumin dan serum albumin (Smith, 1969). Aifa-laktalbumin berperan sebagai
komponen dari laktosa sintetase yaitu sistem enzim dalam sintesis laktosa (Schalm et
al.. 1971; Ebner dan Schanbacher. 1974).
Immunoglobulin adalah globuiin yang sudah terbentuk dan langsung dialirkan
dari darah ke dalam ambing. Immunoglobulin rnengandung zat kekebalan, terdapat dalam kolostrum dan susu akhir laktasi clan terdiri atas dua fraksi yaitu euglobulin dan pseudoglobulin (Smith, 196% Schalm et al.. 1971). Kecuali itu terdapat bermacam-
macam enzim (satu persen), protein lain (0.06-0.17%) yang terdiri atas antitripsin,
antibakterial, delta-kasein dan lain-lain (Smith, 1969).
Kandungan protein di dalam susu sapi berkisar dari
2-7-4896
(Foley et al.,1972; Holmes dan Wilson, 1984) dan kadarnya relatif tetap sepanjang laktasi.
Prekursor protein adalah asam amino esensiai dan asam amino non esensial dalam
darah. Sintesis protein dipengaruhi oleh hormon dan ada di bawah kontrol genetik,
Lebih dari 60 persen asam amino esensial dalam darah, terutama asam amino yang mengandung sulfur, ditransfer dari darah ke dalam ambing. Pada umumnya
pengambilan (uotake) asam amino esensial sudah sesuai dengan asam amino yang
dikeluarkan dalam protein susu, tetapi kekurangan asam amino yang mengandung
sulfur diduga akan membatasi sintesis protein susu, bahkan sangat mempengaruhi
produksi susu (Holmes dan Wilson, 1984). Asam amino non esensial untuk sintesis protein di dalam ambing sebagian disintesis dari asam amino esensial yang diambil
dalam jumlah berlebihan (Holmes dan Wilson, 1984), sebagian lagi berasal dari darah
(Smith, 1969). Diduga 70 persen glutamin, asarn glutamat dan tirosin, 50 persen
asparagin clan prolin dan sebagian kecil serin ditransfer dari darah ke dalam ambing dan sebagian asam amino bebas bergabung ke dalam molekul protein di ambing. Hampir
separuh asam amino non esensial dalam kasein tidak disintesis di kelenjar ambing tetapi
ditransfer dari darah (Smith, 1969).
Laktosa S u . Laktosa m e ~ p a k a n karbohidrat dalarn susu yang disintesis di dalam ambing. Prekursor Iaktosa yang utama adalah glukosa darah. Delapan puluh
persen glukosa darah digunakan untuk sintesis laktosa dan harnpir seluruh laktosa
disintesis dari glukosa (Holmes dan Wiison, 1984). Pengambilan glukosa oleh kelenjar
ambing tidak dipengaruhi oleh insulin damh dan sebanding dengan glukosa
darah.
Glukosa
> glukosa-6P
glukosa-6-P
<
->
glukosa-1-PUTP
+
glukosa-1-P<
-
> UDP-glukosa
+
PPiUDP glukosa
<
-> UDP galaktosa
UDP galaktosa
+
glukosa-I-PO4 ->
laktosa-1-PO4+
UDPlaktosa- 1
PO^
>
laktosa+
PO4Reaksi ini dikatalisir oleh enzim laktosa sintetase yang sebenarnya melibatkan enzim
lain (galaktosil transferase) dan protein (a-laktalbumin) di lumen alat Golgi dalarn sel sekretori ambing. Dari glukosa yang masuk ke dalam glukosa-6P, 50-60%
dikonversikan menjadi galaktosa yang selanjutnya digunakan untuk sintesis laktosa susu
(Ebner dan Schanbacher, 1974).
Kadar laktosa susu relatif tetap (Preston clan Leng, 1987) tetapi produksi laktosa rneningkat sejdan dengan peningkatan produksi susu pada awal laktasi (Holmes dan
Wilson, 1984) dan berfluktuasi sesuai dengan dinamika produksi susu selama laktasi.
Kadar laktosa dalam susu sekitar 3.5-6.0 % (Foley et al., 1972) dan merupakan bagian terbesar (52%) dari bahan kering tanpa lemak (BKTL) susu (Schalm et al., 1971).
Kualltas Susu
Kualitas susu merupakan faktor penting dalam rangka penyediaan pangan yang
sehat untuk konsumen clan sangat mempengaruhi hasil olahannya. Salah satu penilaian kualitas susu yang terpenting adalah bilangan kuman (angka bakteri) karena merupakan
salah satu syarat penerimaan susu oleh Industri Pengolahan Susu dan mempengaruhi
harga susu. Menurut patokan "Milk Codex", susu yang sehat dan layak diminum
apabila mangandung kuman tidak lebih dari satu juta per mililiter susu segar (Sudono,
1985). sedangkan menurut Peraturan yang dikeluarkan oleh Direktorat Jendral
Peternakan (SK No. 17/KPTS/Dj.P/Dpta.n/83, pasal 7) ju~nlah kuman setinggi-
Jumlah kuman dalam susu juga sangat erat hubungannya dengan gejala mastitis
subklinis. Mastitis subklinis tidak memperlihatkan gejala fisik pada ambing maupun
pada komposisi susu, namun bila dibiarkan kemungkinan dapat menjadi mastitis klinis.
Mastitis merupakan problema utama bagi kesehatan sapi perah dan produknya.
Menurut Soewardi et al., (1990) sapi perah di pulau Jawa 63 persen menderita mastitis subklinis dan lima persen menderita mastitis klinis yang dapat menurunkan produksi
masing-masiing sekitar 10 dan 25 persen.
Susu yang dieksresikan ke luar ambing tidah pernah bebas dari kuman sebab
ketika melalui ductus alveoli, susu sudah mulai tercemar oleh kuman yang ada di sana,
walaupun susu itu berasal dari sapi yang sehat ( Foster et al., 1957). Jumlah bakteri dalam susu beragam tergantung dari faktor yang mempengaruhinya antara lain kondisi
waktu pemerahan dan penanganan susu setelah pemerahan. Jumlah kuman susu
meningkat sangat cepat sesuai dengan waktu tetapi juga bergantung kepada komposisi medium, organisme yang terlibat, kadar air, suhu, pasokan oksigen dan pH (Foster et
al., 1957).
Kontaminasi susu oleh bakteri dapat berasal dari dalam atau dari luar ambing.
Faktor luar sangat besar pengaruhnya terhadap jumlah kuman dalam susu seperti kondisi kandang dan tubuh sapi, kondisi alat untuk memerah susu dan kondisi pemerah.
Penggunaan mesin perah dilaporkan mengkibatkan jumlah kuman meningkat apabila
kebersihannya tidak diperhatikan, karena kuman yang berasal dari ambing sapi
penderita mastitis akan tertinggal di dalam mangkuk puting (Holmes dan Wilson,
1984). Foster et al., (1957) melaporkan jumlah kuman dalam susu b s i l pemerahan
oleh mesin perah (1616 sel/ml susu) berbeda sebesar 44.68 persen unit dengan hasil pemerahan oleh tangan (894 sel/ml susu)
.
Kebersihan ambing besar pengaruhnya terhadap jumlah kuman. Pencukuran
susu) (Foster et al, 1957). Ernawati (1990) melaporkan ha1 yang sama, yaitu kambing perah yang bulu ambingnya dicukur dapat menurunkan jumlah kuman sebesar 18.42 persen unit.
Teat dipp- (pencelupan puting ke dalarn larutan desinfektan) dilaporkan dapat menurunkan jumlah kuman susu. Dari beberapa peneliti Santosa (1986)
melaporkan teat dipping &pat menurunkan kejadian infeksi sekitar 53-64 %, tetapi
-g hanya dapat mengendalikan beberapa jenis bakteri terutama Staphylococcus aureus dan Streptococcus agalactiae, yang merupakan 90% penyebab mastitis.
.
.Penurunan jumlah maupun jenis kxman akibat W chppirg bergantung kepada jenis desinfektan yang digunakan. Chlorhexidine gluconat (0.55%) dapat menurunkan
kejadian infeksi oleh Staphylococcus aureus sekitar 92.5 persen, sedangkan Chlorhexidine digluconat menurunkan infeksi Streptococcus agahctiae sekitar 71
persen (Santosa, 1986). Staphylococcus aureus juga dapat dikendalikan oleh zeolit yang
berperan sebagai antibakteri dalarn balon catheter untuk mengontrol infeksi saluran urin pada manusia (Uchida et al., 1992).
Metabolisme Protein dan Energi
Metabolisme Protein
Metabolisme protein pada ternak ruminansia terjadi pada lokasi yaitu di rumen
dan di sel tubuh. Di rumen protein mengalami proteolisis oleh enzim asal mikroba menjadi oligopeptida dan asam amino. Sebagian asam amino didegradasikan menjadi
amonia (NH3) dengan menghasilkan asam keto d f a , asam lemak atsiri (VFA), CO, dan
CH, (Sutardi, 1977). Senyawa
N
bukan protein(NPN)
dirombak menjadi NH3.Amonia dibutuhkanoleh sebagian besar mikroba sebagai sumber N, asarn keto
sintesis aytm amino penyusun protein sel mikroba (Sutardi,l977; Maynard et al., 1979). Gsiensi sintesis protein mikroba bergantung pada sinkronispsi pelepasan NH3
I
i
dari protein dan VFA dari karbohidrat (Haresign dan Cole, 1984). Pelepasan amonia
yang diperlambatlbenahap misalnya dengan penggunaan biuret, zeolit beramonium
(Emanto, 1992) atau pelepasan VFA yang dipercepat misalnya dengan pemanasan -tk?
sumber pati (Sofyan) telah berhasil rneningkatkan sintesis protein mikroba rumen.
Kebutuhan amonia untuk sintesis protein mikroba yang maksimal terbatas pada
konsentrasi 50 mg/liter cairan rumen (Satter dan Slyter, 1974). Namun menurut Preston clan Leng (1987) kadar amonia yang optimum sebesar 150-200 mglliter Miran rumen dan tergantung
dari
jenis ransum sedangkan kadar amonia untuk efisiensi penggunaan protein dan energi ransum sebesar 7-8 mM (Erwanto et al., 1993).Arnonia di dalam rumen tidak selumhnya digunakan oleh mikroba, sebagian
amonia akan diserap melalui dinding rumen dan sebagian kecil akan masuk ke organ lain dari alat pencernaan. Jumlah amonia yang diserap melalui dinding rumen
bergantung kepada pH rumen, makin tinggi pH (basa) makin besar jumlah amonia yang diserap (Sutardi, 1977; Haresign dan Cole, 1984).
Amonia yang diserap akan dibawa darah ke hati kemudian dikonversikaan menjadi urea. Sebagian urea kembali ke rumen melalui saliva dan sebagian lagi dike- luarkan dari tub& melalui urin. Jumlah amonia yang melebihi kapasitas hati untuk
diubah menjadi urea memungkinkan tcxjadi keracunan karena meningkatnya kadar arnonia dalam darah.
Protein mikroba dan protein pakan yang 1010s degradasi akan dicerna dl abomasum clan usus halus menjadi asam amino dan selanjutnya diserap ke dalam darah melalui dinding usus. Asam amino kemudian dibawa ke hati dan bergabung ke dalam
untuk sintesis protein sel jaringan tubuh (Haresign dan Cole, 1984). Asam amino yang tidak digunakan akan kembali ke hati dan dideaminasikan menjadi amonia clan asam
keto alfa atau dikonversikan menjadi energi. Arnonia hasil katabolisme protein akan
diubah menjadi urea clan dikeluarkan melalui urin.
Metabolisme Enerei
Semua zat makanan kecuali air dan abu mempunyai potensi sebagai penghasil
energi. Di dalarn tubuh zat makanan dimetabolismekan menjadi energi untuk memenuhi
kebutuhan hidup dan produksi. Karbohidrat dan lemak dikatabolismekan menjadi
karbon dioksida dan air, sedangkan protein dirombak rnenjadi karbon dioksida, air dan senyawa nitrogen
(N)
yang keluar melalui urin. Pada mamalia sebagian besar N urindalam bentuk urea.
Sebagian energi asal makanan digunakan untuk metabolisrne basal yang besarnya sama dengan panas yang diproduksi ketika hewan dalam keadaan istirahat, pasca penyerapan (puasa) dan kondisi suhu lingkungan yang termone& (Brody, 1964).
Proses fermentasi dan metabolisme zat rnakanan mengakibatkan kenaikan produksi panas yang besarnya sekitar 50-40%, aktivitas t e d di kandang rnenaikkan produksi panas sekitar 5-7s dari panas metabolisme basal (Moen, 1973). Sapi perah pada puncak laktasi memproduksi panas dua kali lebih besar daripada sapi tidak laktas~ (Moen, 1973). Kenaikan produksi panas selaras dengan jumlah konsumsi pakan dan
produksi susu (Brody, 1964; Holmes dan Wilson, 1984).
Penggunaan energi zat makanan oleh ternak dapat diduga dengan mengukur produksi panas atau retensi energi dalam jaringan yang secara tidak langsung dapat diukur dengan neraca karbon - nitrogen (Church clan Pond, 1982; Bernejee, 1982; Lloyd er al., 1978). Neraca karbon-nitrogen (neraca
C-N)
dapat menduga jumlah energi yang tersimpan di dalam tubuh sebagai protein (pertumbuhan) dan lemakNeraca nitrogen dihitung dari selisih nitrogen yang dimakan dengan nitrogen
yang keluar melalui urin dan tinja (Munro dan Allison. 1964). Pada ternak Laktasi protein susu diperhitungkan sebagai N yang keluar tubuh melalui air susu (Maynard et
al., 1979; Holmes
dan
Wilson, 1984; Lloyd et al., 1978).Neraca karbon diperhitungkan dari selisih karbon yang masuk dengan karbon
yang keIuar melalui urin, tinja, gas dan susu (Holmes dan Wilson, 1984). Neraca nitrogen
dan
neraca karbon dapat bernilai positif, negatif dan nol. Neraca positifapabila jumlah pasokan nitrogen atau karbon lebih besar daripada jumlah nitrogen atau karbon yang keluar tubuh. Apabila keluaran N atau karbon (C) Iebih besar dari pasokannya neraca akan bernilai negatif
dan
nilai neraca no1 bila jumlah pasokan Natau C sarna dengan jumlah keluaran N atau C .
Neraca nitrogen dan neraca karbon yang positif dapat rnenduga N dan karbon
yang diretensi (Lloyd et a!., 1978; Barnejee, 1982; Holmes dan Wilson, 1984). Jumlah N yang diretensi dalam bentuk daging dapat dikonversikan sebagai karbon yang
diretensi. Dengan menggunakan hukum Hess (Brody, 1964) maka dari jumlah seluruh
karbon yang diretensi diperoleh gambaran produksi karbondioksida yang &pat
menduga jumlah panas yang diproduksi (Sastradipradja, 1995).
Zeolit dan Penggunaannya dalam Ransum
Zeolit terdapat berlimpah dan tersebar di sekitar 46 lokasi di Indonesia. Ekberapa lokasi terutama di pulau Jawa sudah dieksplorasi dan menghasilkan zeolit
alam yang terdapat di pasaran.
Zeolit ditemukan sekitar pertengahan abad ke 18, terdapat sekitar 40-50 spesies
penggunaan zeolit masih dalam taraf uji w b a (Suyartono dan Husaini, 1991). Walaupun penggunaan zeolit di Indonesia relatif masih baru, zeolit dilaporkan
memberikan respons positif terhadap produksi tanaman dan efisiensi pemupukan
(Astiana, 1992).
Zeolit yang terdiri atas beberapa spesies adalah kelompok mineral aluminosilikat
terhidrasi dengan alkali dan alkali tanah. Unit dasar zeolit adalah tetrahedra-tetrahedra yang berikatan satu sama lain dengan silika oksigen dan aluminium oksigen yang
rnenduduki atom pusat tetrahedra. Tetrahedra yang bergabung dalam bentuk polihedra
dalam susunan tekstosilikat membentuk kristal tiga dimensi yang berongga (Clarke,
1980; Mumpton
dan
Fishman, 1977; Tsitsishvili, 1988). Kedudukan aluminium yangbervalensi tiga dalarn tetrahedra mengakibatkan muatan listrik menjadi negatif. Kenetralan muatan listrik di dalam zeolit dipelihara oleh kation-kation bervalensi satu
clan valensi dua dari alkali dan alkali tanah. Kation alkali diantaranya Na',
K
'
,
Pb'
Cs' clan kation yang termasuk alkali tanah antara lainBe2+,
Mg',
Caz+, S?+ (Evans, 1989), tetapi kation yang paling sering dijumpai dalam zeolit adalah Na+, K+, CaZ+ dan M&+yang tersebar di permukaan maupun di dalam rongga zeolit
dan
dapat dipertukarkan dengan ion lain (Mumpton dan Fishman, 1977; Clarke, 1980; Tsitsishvili, 1988;Evans, 1989).
C
Struktur zeolit mempakan kerangka berongga yang dihubungkan ke segala arah oleh saluran-sduran b e ~ k ~ r a n 2-7 A". Rongga-rongga kosong tersebut menempati 50
persen &ri bangunan zeolit (Clarke, 1980). Struktur zeolit sangat terbuka, pori-pori
dalam celah-celah kristalnya memungkinkan zeolit dapat dilalui air dengan mudah.
Molekul air dapat ditahan dan dilepas secara reversible (Clarke, 1980 dan Evans,
1989).
Zeolit alam maupun zeolit sintetis selain berperan sebagai penyaring molekul,
dapat berfingsi sebagai adsorbent, dapat berperan sebagai katalis dan mempunyai
Tsitsishvili, 1988; Mumpton, 1988; Evans, 1989). Walaupun semua spesies zeolit
berkemampuan sebagai penukar kation tetapi kapasitasnya beragam dan kemampuan
menyeleksi kation yang akan dipertukarkan juga berbeda. Kapasitas tukar kation
klinoptilolit yang mernpunyai struktur (Na,K4)(Als Sia 0J.24H20, rendah (2.54 meqlg), sedangkan kapasitas tukar kation kabasit yang mempunyai struktur
(Na2Ca)6(All2 Si, 0 , ) . 4 0 H 2 0 Iebih tinggi (3.81 meq/g). Perbedaan kapasitas tukar
kation klinoptilolit dan kabasit dapat dilihat dari nisbah aluminium dengan silikanya, masing-masing sebesar 1:s dan 1:2. Menurut Mumpton dan Fishman (1977) serta Evans (1989), kapasitas tukar kation rnerupakan fungsi dari substitusi Si oieh A1 d d a m
kerangka zeol i t
.
Kemampuan setiap jenis zeolit untuk menyeleksi kation yang dapat dipertukar-
kan penting diketahui sehingga penggunaan jenis zeolit akan dapat disesuaikan dengan
fungsi dan tujuannya. Kation yang dapat dipertukarkan dalam struktur klinoptilolit
beru~tZUI sebagai berikut: Cs
>
Rb>
K>
N Y>
Ba
>
S r>
Na>
Ca>
Fe>
A1 > Mg>
Li (Mumpton dan Fishman, 1977). Walaupun kemampuan tukar kation klinoptilolit rendahtetapi daya ikatnya terhadap ion amonium lebih kuat daripada daya ikatnya terhadap
kation alkali penting lainnya seperti Na, Ca, Mg, Sr. Kemampuannya untuk mengikat
ion amonium menyebabkan klinoptilolit banyak digunakan untuk membersihkan bahan buangan dan di bidang pertanian dipakai sebagai "slow release reservoir N" untuk pemupukan tanaman (Mumpton dan Fishman, 1977).
ZeoIit alam walaupun berlimpah dan lebih murah namun manfaamya dibatasi
oleh kemurniannya sebagai mineral campuran. Zeolit alam sering tercarnpur bahan
yang tidak diperlukan seperti kuwarsa, felspar, illit, kalsit, kristobalit. Zeolit yang baik
kualitasnya mengandung 90-95 % mineral tunggal zeolit (Mumpton, 1990).
Zeolit d a m di Indonesia pa& umurnnya terdiri atas dua spesies yaitu
klinoptilolit dan mordenit serta mengandung beberapa senyawa lain seperti kwarsa,
mengandung Si, Al, Na, Mg, Ca, K dan Ti (Anwar et al., 1985; Husaini, 1992; Astiana, 1992). Na, Mg, Ca dan K diduga merupakan kation yang terdapat dalam
saluran kristal zeolit yang dapat dipertukarkan dengan kation lain walaupun menurut
Anwar er al. (1985) ada kemungkinan berasal dari mineral lain clan Ti bukan kation yang dapat dipertukarkan. Zeolit alam Indonesia dianggap bermutu baik apabila
kandungan zeolitnya lebih dari 60 persen, mengandung lempung tidak lebih dari 10 persen dan tidak mengandung bahan karsinogenik (asbes, erionit). KTK = 100-200
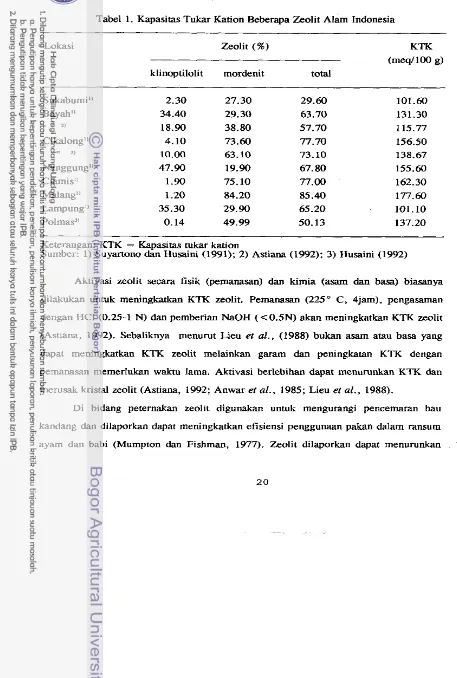
meq/100g (Suyartono dan Husaini, 1991). Nilai
KTK
zeolit d a m dari beberapa lokasi di Indonesia disajikan pada Tabel 1.Pada Tabel 1 dapat dilihat hubungan positif antara total zeolit dengan nilai KTK.
Menurut Anwar et al. (1985) zeolit dari MaIang (Jatim), Cikalong-Tasikmalaya dan Ciamis (Jabar) yang proporsi mordenitnya lebih tinggi dari IclinoptiIolit, mutunya ter-
baik, sebaliknya zeolit dari Lampung, Nanggung-Bogor clan Bayah-Lebak (Jabar) yang
proporsi klinoptilolitnya lebih tinggi dari mordenit, mutunya lebih rendah. Klinoptilolit mempunyai KTK yang refatif rendah (Mumpton dan Fishman, 1977; Evans, 1989). Kapasitas tukar kation juga tergantung kepada keragaman struktur kristalnya yang juga
menentukan kemurniannya (Lieu et al. 1988).
Menurut Astiana (1992) zeolit d a m di Indonesia mempunyai kemampuan mem-
pertukarkan kation yang ada di dalam rongga zeolit dengan kation dalarn larutan
Tabel 1. Kapasitas Tukar Kation Beberapa Zeolit Alam Indonesia
Zeolit (%) KTK
(meq/lOO g)
klinoptilolit mordenit total
Keterangan: KTK = Kapasitas tukar kation
Sumber: 1) Suyartono dan Husaini (1991); 2) Astiana (1992); 3) Husaini (1992)
Aktivasi zeolit secara fisik (pemanasan) dan kirnia (asam dan basa) biasanya
dilakukan untuk rneningkatkan KTK zeolit. Pernanasan (225" C, 4jam), pengasaman dengan HCI (0.25-1
N)
dan pemberian NaOH (<0.5N)akan
meningkatkan KTK zeolit (Astiana, 1992). Sebaliknya rnenurut Lieu et al., (1988) bukan asam atau basa yang dapat meningkatkan KTK zeolit melainkan garam dan peningkatan KTK dengan pemanasan memerlukan waktu lama. Aktivasi berlebihan dapat menumnkan KTK danmerusak kristal zeolit (Astiana, 1992; Anwar et al., 1985; Lieu et al., 1988).
Di bidang peternakan zeolit digunakan untuk mengurangi pencemaran bau
kandang dan dilaporkan dapat meningkatkan efisiensi penggunaan pakan dalam ransum
[image:45.557.44.501.48.726.2]kasus diare pada anak sapi (Verzgula et al., 1988). Di dalam rumen, zeolit dapat menangkap NH4+ sarnpa 15 persen dan menurunkan kadar amonia rumen serta menurunkan kadar amonia dan urea darah (Mumpton, 1984).
Urea biasanya digunakan sebagai sumber N yang murah bagi mikroba rumen, tetapi karena sangat mudah dan sangat cepat didegradasikan menjadi amonia di dalam
rumen, maka risiko keracunannya tinggi. Penggunaan urea juga harus diIengkapi
dengan sumber karbohidrat yang mudah difermentasikan yang dalam kondisi tertentu
sulit diperoleh karena penggunaannya bersaing dengan kepentingan lain. Hasil
fermentasi karbohidrat di dalam rumen berupa volatile fatty acid (VFA) akan digunakan
sebagai sumber energi oleh mikroba untuk mensintesis seinya. Sinkronisasi pelepasan
amonia dengan VFA akan meningkatkm efisiensi penggunaan amonia, mengurangi
akumulasi amonia di dalam rumen dan risiko keracunan dapat dicegah. Upaya memper-
lambat pelepasan N dengan penggunaan biuret memerlukan masa adaptasi yang lama
dan mahal.
Melalui proses tukar kation, sebagian kation dalam zeolit dapat diganti oleh ion
amonium (Lieu et al., 1988) menjadi zeolit beramonium (Z-NH,'). Zeolit yang sebagian besar kationnya sudah diganti oleh ion amonium dapat diregenerasikan baik
oleh larutan garam yang kationnya dapat menggantikan posisi amonium maupun oleh
pemanasan bahkan oleh bakten yang berpenetrasi ke dalam rongga zeol~t dan menggunakan NH,' untuk nitrifikasi yang memproduksi NO2 (Lieu et a l . , 1988). Pengisian zeolit dengan ion amonium dari garam amonium asetat telah dilakukan oleh
Erwanto (1992) dalam skala Iaboratorium. Dengan menggunakan larutan amonium
asetat satu N dan nisbah amonium asetat : zeolit = 10 : 1 diperoIeh zeolit beramonium yang mengandung 34.44 mg ion amonium dalam setiap gram zeolit. Untuk
mendapatkan zeolit beramonium skala besar Lieu et al., (1988) menyarankan perendaman zeolit dengan larutan garam amonium asetat selama satu minggu.
'.
"
,
\Melalui teknologi amoniasi seperti yang dilakukan oleh Erwanto (1990) dan disarankan oleh Lieu et aC., (1988) zeolit dapat dirnanfaatkan sebagai "slow release
reservoir N" bagi mikroba di ddam rumen. Melalui pertukaran kation posisi ion
amonium di dalam zeolit kemungkinan besar dapat digantikan oleh kation-kation ~ a +
dan
K +
yang be& dari saliva. Laju aliran saliva yang tents menems memungkinkanpelepasan ion arnonium dari zeolit terjadi secara bertahap sehingga akumulasi arnonia
yang dapat rnengakibatkan keracunan dapat dicegah. Nitrogen dari zeolit ~ ~ ~ U I I O N U ~
akan di-tkan oleh mikroba mmen untuk mensintesis protein selnya. Efek zeolit berZtUI0~~m dalam m s u m tinggi konsentrat pada beberapa peubah di cairan rumen in
vitro disajikan pada Tabel 2.
Tabel 2. Efek Zeolit Beramonium terhadap Kadar N-NH3, pH dan Sintesis Mikroba Rurnen
Parameter Kadar zeolit beramonium ( % I
Amonia, mM 3 . 6 3 " 3.98b 4 . 5 3 ' 4.78=" 4 . 8 3 * PH 6.40' 6.43"' 6
.
441b 6 . 47bC 6.
44ab Laju sintesa protein 102. I l l a b C 117= 107"' 114bc mikroba (mg/l/ j am)La j u pertumbuhan 209" 228"" 240' 220ab 234bC mikroba (mg/l/ j am)
Sumber : Erwanto, 19W
Di dalarn rumen zeolit juga berperan sebagai pengendali pH (Clarke, 1980;
Kalachnyuk, 1989; Erwanto, 1992). Efek positif zeolit terhadap sistem fermentasi
[image:47.561.48.501.21.727.2]lemak susu
(X),
Z = kadar zeolit beramonium dalarn konsentrat (g/kg) (Sutardi, 1990).Murni (1993) melaporkan bahwa penggunaan zeolit 5-7.5 % dapat menurunkan
kadar air ransum yang disimpan selama 10-52 hari. Pertumbuhan jamur dapat ditekan
dengan pemberian zeolit 10 persen. Penggunaan zeolit 2.5 persen dapat menekan produksi aflatoksin sebesar 80 persen. Zeolit sangat nyata menyerap produksi amonia
manur ayam (Azhari, 1995).
Hasil percobaan beberapa peneliti yang dilaporkan oleh Mumpton (1984)
menunjukkan pemberian zeolit 2-5% pa& sapi perah mu& dapat memperbaiki
performans dan mengurangi kejadian diare serta mengurangi bau kotoran. Pada sapi laktasi penggunaan dua persen zeolit m e ~ l ~ e g a h diare dan meningkatkan produksi. Penggunaan zeolit 2-3% dari konsentrat &pat meningkatkan kadar lemak susu,
menurunkan bilangan bakteri dan mengurangi kejadian mastitis (Sutardi, 1990).
Nampaknya dosis zeolit yang optimum adalah &tar 2-38 dari konsentrat, seperti
yang direkomendasikan oleh Sutardi (1990) sebesar 2.7-346 untuk sapi laktasi, 2.5
persen untuk sintesis protein mikroba (Erwanto,l992) clan 0.5 g/kg bobot badan untuk sapi pedaging (Kalachnyuk, 1989).
Kebutuhan Asam Amino S a ~ i s i
Semula diduga bahwa kualitas protein tidak $enting bagi ruminansia.
Ruminansia dianggap tidak membutuhkan kelengkapan asam amino karena kebutuhan
proteinnya sudah dipenuhi oleh mikroba rumen. Mikroba rurnen dapat mensintesis
asam amino esensial maupun asam amino non esensial (Bryant, 1977). Mikroba rumen
mengandung sekitar 65 persen protein yang terdiri atas asam amino esensial (Hungate,
Dugaan ini diperjelas oleh beberapa pedoman pemberian ransum yang tidak
merekomendasikan kebutuhan asam amino untuk sapi laktasi. Ternyata beberapa
pendapat melaporkan bahwa terdapat perbedaan antara komposisi dan proporsi asam
amino antara pakan, protein mikroba dan air susu.
Pada Tabel 3 dapat dibandingkan antara kandungan asam amino dalam mikroba
rumen dengan rumput. Pada umumnya kandungan asam amino mikroba lebih tinggi
daripada asam amino rumput kecuali metionin dan leusin. Hal ini menunjukkan telah
terjadi perbaikan mutu protein rumput setelah disintesis dalam bentuk protein mikroba
(Shirley, 1986).
Tabel 3. Komposisi Asam Amino Protein Pakan, Mikroba Rumen, Digesta Abomasum dan Susu
Asam amino rumput" mikroba rumen') digest2 susu2'
Metionin (MET) Arginin (ARG) Triptofan (TRP) Threonin (THR) Histidin (HIS) Isoleusin (ILE) Leusin (LEU) Lisin (LYS) Valin (VAL) Fenilalanin (PHE) Sistin (CYS)
Sumber : 1) Shirley, 1986 2) Hogan, 1974
Kebutuhan asam amino untuk sapi laktasi sulit diketahui karena fermentas; telah
mengubah komposisi asam amino yang diserap di usus sehingga terjadi "imbalance"
antara asarn amino yang tersedia dengan asam amino yang dibutuhkan (MacRae clan
[image:49.546.41.493.7.719.2]dinding sel bakteri yang lebih tahan terhadap enzim proteolitik ternak sehingga bakteri lebih sulit dicerna daripada kasein (Hogan, 1974). Kecernaan bakteri tergantung dari
kemarnpuan enzim untuk menembus (penetrasi) dinding sel.
Proporsi asam amino dalam digesta abomasum dengan asam amino dalam susu
nampak berbeda (Tabel 3). Asam amino lisin dan asarn amino yang mengandung sulfur
(metionin) di dalam susu lebih tinggi daripada dalam digesta. Leusin diduga merupakan
asam amino yang membatasi produksi susu (Hogan, 1974) sedangkan pada awal laktasi
kecuali leusin, pasokan metionin, isoleusin, lisin dan histidin juga sering tidak dapat
memenuhi kebutuhan tetapi metionin nampaknya merupakan asam amino pembatas pertarna. Digesta, mikroba maupun rumput hanya dapat menyediakan leusin masing-
masing 71.82. 73.64 dan 90.90% dari leusin yang dibutuhkan untuk susu.
Menurut Shirley (1986) kebutuhan asam amino pada sapi perah bergantung
kepada jumlah produksi susu. Metionin, valin, isoleusin, triptofan dan lisin dari pakan
dan mikroba sudah cukup untuk memenuhi kebutuhan produksi susu 10 kglhari, tetapi
untuk produksi 15 kglhari dibatasi oleh kecukupan asam amino MET dan
VAL.
Isoleusin menjadi pembatas produksi susu 20 kg/hari, sedangkan produksi lebih dari 20
kg/hari dibatasi oleh triptofan dan lisin (Shirley, 1986). Kaufmann dan Lupping (1982) menyebutkan MET, LYS, HIS, PHE dan asam amino berantai cabang sebagai asam amino pembatas pada sapi laktasi.
Walaupun beberapa asam amino membatasi produksi susu tetapi menurut Hogan
(1974) hanya pada dua status fisiologis yaitu sapi awal laktasi dan pertumbuhan wol
pada domba yang terbukti terjadi defisiensi asam amino. Menurut Broderick et al.,
(1974) asam amino esensial yang dikategorikan sebagai asam amino pembatas ialah
apabila konsentrasinya dalam plasma darah meningkat sesuai dengan peningkatan taraf protein dalarn ransum. Oleh sebab itu kecukupan asam amino dapat diukur dengan
melihat h d a r asam amino tersebut dalarn plasma darah tepat pada saat kadarnya
meningkat. Broderick et al. (1974) menernukan valin, rnetionin dan lisin sebagai asam
amino pembatas untuk produksi susu berdasarkan pola perubahan konsentrasi asarn
amino dalam vena leher dan vena ambing sebagai respons terhadap perubahan konsumsi protein.
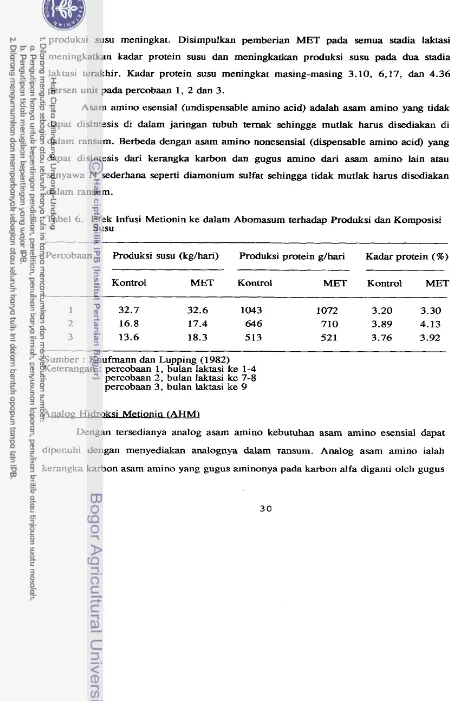
Dari beberapa pendapat ternyata MET selalu menjadi pernbatas untuk produksi susu (Shirley, 1986; Hogan, 1974 dan Broderick et al., 1974) bahkan pa& awal laktasi
MET rnenjadi asam amino pembatas pertama (Hogan, 1974; Broderick et al., 1974; Taminga, 1982; Kaufmann dan Lupping, 1982). Metionin sebagai asarn amino pembatas pertama diperkuat oleh data pada Tabel 4 yang rnemperlihatkan efek
penarnbahan asam amino langsung ke dalam abomasum (intra abomasal infusions) yang
sudah diseirnbangkan terhadap kandungan protein susu ("amino acid balance").
Tabel 4. Efek Carnpuran Asam Amino terhadap Kandungan Protein Susu
Substrat yang diinfusikan Perubahan kandungan
protein susu (g/kg)
Kasein atau campuran AAE lengkap
+
1.7MET - 0.5
LYS
+
0.1MET LYS
+
1.1MET LYS VAL
+
1.4MET LYS THR
+
0.4MET LYS VAL ILE
+
1.6MET LYS VAL ILE PHE
+
1.9MET LYS VAL ILE LEU
+
1.5MET LYS VAL ILE LEU HIS
+
0.9MET LYS VAL THR ILE PHE LEU HIS
+
2.1Sumber : Thomas dan Martin dulm Garnsworthy (1988)
Keterangan : AAE = asam amino esensial; MET = metionin; LYS = lisin; VAL = valin; THR = threonin; ILE = isoleusin; PHE =
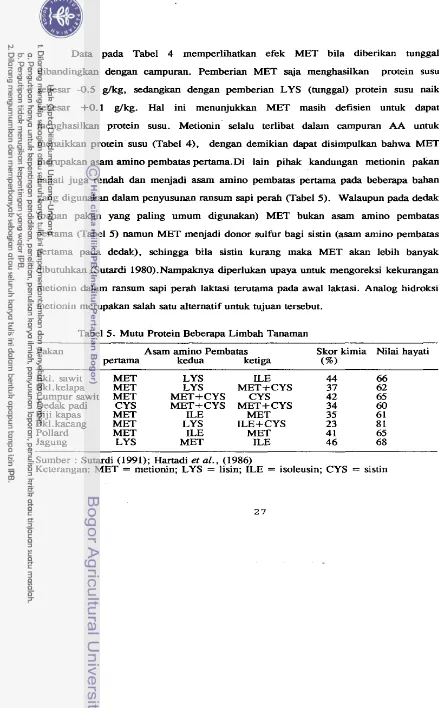
[image:51.559.49.489.10.726.2]Data pada Tabel 4 memperlihatkan efek MET bila diberikan tunggal
dibandingkan dengan campuran. Pemberian MET saja menghasilkan protein susu
sebesar -0.5 glkg, sedangkan dengan pemberian LYS (tunggal) protein susu naik
sebesar +0.1 glkg. Hal ini menunjukkan MET masih defisien untuk dapat
menghasilkan protein susu. Metionin selalu terlibat dalam campuran AA untuk menaikkan protein susu (Tabel 4), dengan demikian dapat disimpulkan bahwa MET merupakan asarn amino pembatas pertama.Di lain pihak kandungan metionin pakan
nabati juga rendah dan menjadi asarn amino pembatas pertarna pada beberapa bahan yang digunakan dalam penyusunan ransum sapi perah (Tabel 5). Walaupun pada dedak (bahan pakan yang paling m u m digunakan) MET bukan asam amino pembatas
pertama (Tabel 5) namun MET menjadi donor sulfur bagi sistin (asam amino pembatas
pertama pada dedak), sehingga bila sistin kurang maka MET akan lebih banyak dibutuhkan (Sutardi 1980).Nampaknya diperlukan upaya untuk mengoreksi kekurangan
metionin dalam ransum sapi perah laktasi terutama pa& awal laktasi. Analog hidroksi metionin merupakan salah satu alternatif untuk tujuan tersebut.
Tabel 5. Mutu Protein Beberapa Limbah Tanaman
Pakan Asam amino Pembatas Skor kimia Nilai hayati
Pertama kedua ketiga (%)
Bkl. sawit MET LYS ILE 44 66
Bkl. kelapa MET LYS MET+CYS 37 62
Lumpur sawit MET MET+CYS CYS 42 65
Dedak padi CYS MET+CYS MET+CYS 34 60
Biji kapas MET ILE MET 35 6 1
Bkl
.
kacang MET LYS ILE+CYS 23 8 1Pollard MET ILE MET 4 1 65
Jagung LYS MET ILE 46 68
Sumber : Sutardi (1991); Hartadi et al., (1986)
[image:52.550.46.485.14.722.2]Metionin (MET) clan Analog Hidroksi Metionin (AHM) dalam Ransum Sapi Perah Laktasi
.
.
etlomnMetionin (MET) addah asam amino yang mengandung sulfur d m esensial
(undispensable) bagi manusia dan ternak monogastrik. Metionin bersifat glukogenik dan lipotropi