PENGEMBANGAN VAKSIN DNA PENYANDI GLIKOPROTEIN
VIRUS KHV (KOI HERPESVIRUS) MENGGUNAKAN
ISOLAT LOKAL
SRI NURYATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi ‘Pengembangan Vaksin DNA Penyandi Glikoprotein Virus KHV (Koi Herpesvirus) Menggunakan Isolat Lokal’ adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, 6 Juni 2010
ABSTRACT
SRI NURYATI. Development of DNA Vaccine Encoding Glycoprotein of Koi Herpes Virus Using Local Isolate. Under direction of FACHRIYAN H.PASARIBU, RETNO D.SOEJOEDONO, ALIMUDDIN, and SUKENDA.
The aim of this research was to develop DNA vaccine of Koi Herpesvirus in carp. Glycoprotein gene originated from ORF 25 KHV virus that is 1.8 kbp in size, is a potential gene in DNA vaccine construction. The sequencing and similarity analysis of the gene using BLAST software showed that this gene is a GP25 gene and was found to be very similar (99%) to GP25 of KHV virus from the USA and Israel. When this GP25 gene was ligated with pGEMT Easy (pT, 3 kbp in size) vector, plasmid size was increasing up to 5 kb. Following this, a digestion of pT-GP25 plasmid using Sal I enzyme was applied resulted in a GP25 fragment (1.8 kbp). Subsequently, pAct-D6 with a size of 8.5 kbp was also cut using the same enzyme to eleminate a D6 gene with a size of 1.6 kbp. GP25 gene was then ligated with pAct resulted a pAct-GP25 plasmid with a size of 8.8 kbp. As there was only one restriction site applied, ligation can be occurred in two different directions. Thus, in order to determine the plasmid with the desired direction PCR analysis was carried out, and plasmid with the right direction was the one that sized approximately 2.1 kbp (no. 6, 17 and 20). High level of GFP gene expression was detected in all tissues analyzed, i.e. kidney, gil, spleen, and muscle 24 hours after injection which indicated that β-actin promoter of Japanese medaka could be active in common carp. Although not as high as on 24 hours, expression was still detected until day 7 and disappeared after day 28 post injection indicating that pAct-GFP can be remained and β-actin promoter of Japanese medaka can be active in muscle, gills and kidney at least a week after injection. The result of RT-PCR analysis on pAct-GP25 injected common carp showed that GP25 gene expression can be detected 14 days after injection in all tissues observed. The expression level differences between GFP and GP25 genes possibly related to the primer sensitivity on annealing process to cDNA template and the size of PCR targeted DNA fragment, as smaller PCR targeted product will be easier to be amplified than a bigger one. The length of PCR targeted DNA with GP25 primer was 1.8 kbp whereas GFP gene was 0.6 kbp.The challenge test on common carp results showed that all unvaccinated and negative control (not infected with KHV) was alive until the end of the experiment. Whereas, the survival of fish vaccinated with a dose of 12.5 was ranged from 60.9 and 96.7%. In relation to the activity of β-actin promoter that was used in this DNA vaccine, gene expression was occured 24 hours after vaccination and remained until 2 weeks. The control fish that was not injected by GP25 showed no GP25 expression either in the observed tissues or organs and resulted in a lower survival, i.e. 23.3%. The challenge test results has indicated that the fish vaccinated with GP25 at doses of 7.5 and 12.5 µg/100µL were able to produce KHV glycoprotein in their body which would be recognized as an antigen that further would activate the fish immune response. In the challenge test, the active immune response was then represented in a higher survival. This result also showed that GP25 is an immunogenic and its expression could enhance common carp immunity on KHV infection.
RINGKASAN
SRI NURYATI. Pengembangan Vaksin DNA Penyandi Glikoprotein Virus KHV (Koi Herpesvirus) Menggunakan Isolat Lokal. Dibimbing oleh FACHRIYAN H.PASARIBU, RETNO D.SOEJOEDONO, ALIMUDDIN, and SUKENDA.
Sekuen gen glikoprotein yang berukuran 1,8 kbp dan berasal dari ORF 25 virus KHV berpotensi untuk dijadikan gen target dalam konstruksi vaksin DNA. Berdasarkan hasil analisis sekuens seperti diungkapkan di atas, fragmen DNA tersebut merupakan gen GP25. Setelah GP25 diligasi ke vektor pGEMT Easy (pT, berukuran 3 kbp), maka ukuran plasmid menjadi sekitar 5 kbp. Digesti plasmid pT-GP25 menggunakan enzim Sal I menghasilkan fragmen GP25 yang berukuran sekitar 1,8 kbp. Berdasarkan hasil sekuensing dan analisis kemiripan (similarity) dengan menggunakan software BLAST diketahui bahwa GP25 dari virus KHV asal Indonesia memiliki kemiripan yang tinggi (99%) dengan GP25 KHV asal Jepang, Amerika Serikat dan Israel.
Selanjutnya, pAct-D6 yang berukuran sekitar 8,5 kbp dipotong dengan enzim Sal I untuk membuang gen D6 (berukuran sekitar 1,6 kbp). Setelah pAct diligasi dengan gen GP25 dihasilkan plasmid pAct-GP25 dengan ukuran sekitar 8,8 kbp. Karena situs restriksi yang digunakan hanya satu jenis, maka kemungkinan arah ligasi bisa 2 macam. Untuk menentukan plasmid dengan arah ligasi yang diinginkan, maka dilakukan analisis PCR. Produk PCR untuk plasmid dengan arah ligasi yang benar memiliki ukuran sekitar 2,1 kbp (nomor 6, 17 dan 20). Konstruksi pActGP25 ini merupakan plasmid DNA sirkuler yang digunakan sebagai vaksin, yang disingkat dengan nama GP25.
Untuk menguji aktivitas vaksin DNA GP25 yang telah dikonstruksi maka dilakukan uji ekspresi gen β-aktin yang digunakan sebagai promoter vaksin. Uji ekspresi gen dilakukan terhadap konstruksi pActGFP (representasi promoter β-aktin) dan pActGP25. Uji aktivitas promoter β-aktin dilakukan dengan menginjeksikan secara intramuscular pActGFP ke juvenil ikan mas ukuran 10-15 g/ekor. Konsentrasi DNA plasmid yang diinjeksikan adalah 12,5 µg/100µL fosfat buffer salin (PBS). Total RNA diekstraksi dari otot yang diinjeksi, insang, limpa dan ginjal ikan mas yang diinjeksi dengan pAct-GFP pada saat 24 jam pasca injeksi (hari pertama) dan 1 minggu pasca injeksi. Sementara itu untuk pAct-GP25, total RNA diekstraksi dari jaringan/organ yang sama dengan pada pAct-GFP, tetapi dilakukan pada 24 jam, 2 minggu dan hari 4 minggu setelah injeksi. Ekstraksi RNA dilakukan menggunakan Isogen (Nippon Gen, Japan). Sintesis cDNA dilakukan dengan menggunakan kit Ready-To-Go You-Prime First-Strand Beads (Amersham Pharmacia Biotech, USA).
Selanjutnya, hasil analisis RT-PCR pada ikan mas yang telah diinjeksi dengan pAct-GP25 menunjukkan bahwa ekspresi gen GP25 dapat terdeteksi setelah 14 hari injeksi pada semua jaringan yang diamati. Perbedaan tingkat ekspresi antara gen GFP dan GP25 diduga berhubungan dengan sensitivitas primer untuk melekat pada cDNA cetakan dan ukuran fragmen DNA target PCR. Secara umum, target produk PCR yang berukuran lebih kecil akan lebih mudah diamplifikasi dibandingkan dengan DNA yang lebih besar. Panjang DNA target PCR dengan primer GP25 sekitar 1,8 kbp, sementara gen GFP sekitar 0,6 kbp.
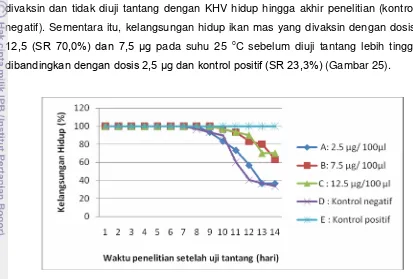
Setelah dilakukan uji tantang didapatkan hasil bahwa semua ikan mas yang tidak divaksin dan tidak diuji tantang dengan KHV hidup hingga akhir penelitian (kontrol negatif). Sementara itu, kelangsungan hidup relatif (sering disebut RPS=relative percent survival) ikan mas yang divaksin dengan dosis 12,5 µg sebesar 60,9% dan 96,7%. Apabila dikaitkan dengan aktivitas promoter β-aktin yang digunakan dalam vaksin DNA ini maka ekspresi gen terjadi 24 jam setelah vaksinasi dan masih terekspresi 2 minggu setelah vaksinasi. Kontrol ikan yang tidak diinjeksi dengan GP25 menunjukkan tidak adanya ekspresi GP25 pada jaringan yang diamati. Ikan kontrol yang tidak divaksinasi dengan GP25 juga menghasilkan kelangsungan hidup yang rendah yaitu sebesar 23,3%. Hal ini menunjukkan bahwa ikan yang divaksinasi dengan dosis 12,5 µg (dalam 100µl PBS) mampu memproduksi glikoprotein KHV di tubuh ikan. Protein ini dikenali tubuh sebagai antigen KHV sehingga ikan dapat mengaktifkan respons imun. Aktifnya respons imun direpresentasikan oleh tingginya kelangsungan hidup relatif ikan yang divaksinasi dengan dosis 12,5 µg dibandingkan dengan dosis lain yang lebih rendah. Setelah dilakukan analisis data dengan ANOVA dan uji lanjut dengan BNT (beda nyata terkecil) didapatkan hasil bahwa perlakuan dosis 2,5 µg memberikan pengaruh yang berbeda dibanding dosis 7,5 dan 12,5 µg, sedangkan perlakuan 7,5 µg memberikan pengaruh yang sama dengan perlakuan 12,5 µg.
Uji keamanan vaksin dilakukan dengan menyuntik ikan mas ukuran 10-15 gram dengan vaksin DNA GP25 dengan dosis 12,5 µg. Ikan ini dipelihara dan dicatat kelangsungan hidupnya. Ikan yang diinjeksi dengan vaksin DNA GP25 selama dua bulan tidak mengalami kematian. Jaringan otot, insang, limpa dan ginjal yang diperiksa secara mikroskopis juga tidak menunjukkan adanya kelainan. Hasil ini menunjukkan bahwa vaksin GP25 bersifat imunogenik dan ekspresinya dapat meningkatkan kekebalan ikan mas terhadap infeksi KHV. Vaksin DNA ini juga aman karena tidak menimbulkan kematian pada ikan yang divaksinasi serta tidak menimbulkan gangguan pada jaringan.
© Hak Cipta milik IPB, tahun 2010
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau
menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau
tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang
wajar IPB
PENGEMBANGAN VAKSIN DNA PENYANDI GLIKOPROTEIN
VIRUS KHV (KOI HERPESVIRUS) MENGGUNAKAN
ISOLAT LOKAL
SRI NURYATI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Sains Veteriner
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
HALAMAN PENGESAHAN
Judul Disertasi : Pengembangan Vaksin DNA Penyandi Glikoprotein Virus KHV (Koi Herpesvirus) Menggunakan Isolat Lokal
Nama : SRI NURYATI
NRP : B 063050031
Program Studi : Sains Veteriner (SVT)
Disetujui,
Komisi Pembimbing,
Prof.Dr.Drh. Fachriyan H.Pasaribu Prof.Dr.Drh.Retno D.Soejoedono,M.S.
Ketua Anggota
Dr. Alimuddin, S.Pi., M.Sc. Dr.Ir. Sukenda, M.Sc.
Anggota Anggota
Diketahui,
Ketua Program Studi Dekan Sekolah Pascasarjana IPB
Penguji pada ujian tertutup: 1. Dr.Drh. Joko Pamungkas, M.Sc. 2. Dr.Ir. Munti Yuhana, M.Sc.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga karya ilmiah berupa disertasi ini berhasil diselesaikan. Judul dari disertasi ini adalah: Pengembangan Vaksin DNA Penyandi Glikoprotein Virus KHV (Koi Herpesvirus) Menggunakan Isolat Lokal.
Ucapan terima kasih dan penghargaan setinggi-tingginya penulis sampaikan kepada Prof.Dr.Drh. Fachriyan H.Pasaribu dan Prof.Dr.Drh. Retno D.Soejoedono, M.S. sebagai guru dan pembimbing, atas bimbingan dan arahan yang diberikan sejak kuliah, penulisan proposal, pelaksanaan penelitian dan penyusunan disertasi ini.
Penulis juga menyampaikan terima kasih dan penghargaan setinggi-tingginya kepada Dr.Alimuddin, S.Pi., M.Sc. dan Dr.Ir. Sukenda, M.Sc. sebagai guru dan pembimbing, atas bimbingan dan saran yang diberikan sejak kuliah, penulisan proposal, pelaksanaan penelitian dan penyusunan disertasi ini. Penulis juga menyampaikan terima kasih atas curahan waktu serta izin untuk memanfaatkan bahan-bahan penelitian di laboratorium selama penelitian berlangsung.
Penulis juga menyampaikan terima kasih dan penghargaan setinggi-tingginya kepada Dr.Drh. Djoko Pamungkas, M.Sc. dan Dr.Munti Yuhana, S.Pi., M.Si. yang berkenan menguji dalam ujian tertutup. Ucapan terima kasih dan penghargaan setinggi-tingginya juga penulis sampaikan kepada Dr.Ir.Murdjani, M.Sc. dan Drh.Agus Setiyono, M.S., PhD. yang bekenan menguji pada ujian terbuka.
untuk pelaksanaan uji tantang skala laboratorium. Ucapan terima kasih juga penulis sampaikan kepada Prof.Dr.Goro Yoshizaki atas kerjasamanya dalam sekuensing DNA glikoprotein yang digunakan dalam penelitian ini; serta Dinas Perikanan Propinsi Jawa Barat atas pendanaan untuk pembuatan vaksin DNA untuk virus KHV ini melalui peran Prof.Dr.Ir. Komar Sumantadinata, M.Sc.
Disertasi ini disusun sebagai syarat untuk kelulusan pada Program Studi Sains Veteriner, Sekolah Pascasarjana IPB. Sebagian materi disertasi ini telah dipresentasikan dalam Seminar Nasional VI Hasil Penelitian Perikanan dan Kelautan di Universitas Gajah Mada Yogyakarta pada tanggal 25 Juli 2009. Sebagian materi yang lain dipublikasi di: a) Jurnal Akuakultur Indonesia Vol 9, No.1, hal. 1-9 dengan judul Pengaruh Vaksin DNA terhadap Aktivitas Fagositosis Sel Darah Putih Ikan Mas Pasca Uji Tantang dengan Koi Herpesvirus (KHV), 2) Jurnal Natur Indonesia, Vol.13 No.1, Oktober 2010 dengan judul Construction of a DNA vaccine using glycoprotein-25 and its expression towards increasing survival rate of KHV- infected common carp
(Cyprinus carpio)(accepted).
Tersusunnya disertasi ini tak lepas dari peranan banyak orang. Kepada teman-teman dan mahasiswa yang banyak membantu: Anna Octavera, Bu Julie Ekasari, Bu Sutiastuti, Bu Novi, Bu Sri Wahyuni, Fuad, Bang Indra, Demin, Ade, Akbar, Iswi, Sofi, Tyas, serta teknisi P.Ranta dan Kang Dedi, dll, penulis mengucapkan terima kasih yang sebesar-besarnya atas bantuan dan kerjasamanya selama penelitian berlangsung. Ucapan terima kasih dan penghargaan yang setinggi-tingginya juga penulis sampaikan kepada Bapak-bapak dan Ibu-ibu di Departemen Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelauutan IPB yang telah memberikan support kepada penulis selama penulis menempuh studi.
Kepada semua pihak yang telah membantu dan tidak dapat penulis sebutkan namanya satu persatu, penulis mengucapkan terima kasih atas segala bantuannya, semoga Allah SWT berkenan memberikan balasan yang setimpal, amin.
RIWAYAT HIDUP
Penulis dilahirkan di Kediri pada tanggal 6 Juni tahun 1971 dari ayah Mulyono dan ibu Supriani. Pendidikan sarjana ditempuh di Program Studi Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan (FPIK) IPB, lulus pada tahun 1995. Pada akhir tahun 1995 penulis diterima sebagai staf pengajar di Departemen Budidaya Perairan, FPIK-IPB. Tahun 1997 penulis menempuh pendidikan S2 di Program Studi Ilmu Perairan, Program Pascasarjana IPB dan lulus pada tahun 2000. Beasiswa pendidikan pascasarjana diperoleh dari Departemen Pendidikan dan Kebudayaan.
Penulis menempuh pendidikan S3 di Program Studi Sains Veteriner, Sekolah pascasarjana IPB pada tahun 2005 dengan beasiswa dari Departemen Pendidikan Nasional. Penulis tercatat sebagai anggota Perhimpunan Mikrobiologi Indonesia. Selama penelitian untuk penyusunan disertasi, penulis telah memasukkan sebagian dari hasil penelitian dalam bentuk jurnal untuk dipublikasikan di jurnal nasional. Karya ilmiah yang dihasilkan oleh penulis adalah: 1) Pengaruh Vaksin DNA terhadap Aktivitas Fagositosis Sel Darah Putih Ikan Mas Pasca Uji Tantang dengan Koi Herpesvirus (KHV), diterbitkan di Jurnal Akuakultur Indonesia, Vol 9, No.1, hal.1-9; 2) Construction of a DNA vaccine using glycoprotein-25 and its expression towards
increasing survival rate of KHV- infected common carp (Cyprinus carpio), dimuat di
Jurnal Natur Indonesia, Vol.13 No.1, Oktober 2010 (accepted). Penulis juga sempat mempresentasikan sebagian hasil penelitian ini dalam Seminar Nasional VI Hasil Penelitian Perikanan dan Kelautan di Universitas Gajah Mada Yogyakarta pada tanggal 25 Juli 2009.
Karya ini kupersembahkan untuk orang tuaku Bapak Mulyono
dan Ibu Supriani,
i DAFTAR ISI DAFTAR ISI……… DAFTAR TABEL……….. DAFTAR GAMBAR………. DAFTAR LAMPIRAN………. PENDAHULUAN……….. Latar Belakang………. Tujuan ……… Manfaat Penelitian……… Aspek Kebaruan……… Hipotesis………. TINJAUAN PUSTAKA………. Penyakit Infeksi Koi Herpesvirus……… Virus KHV (Koi Herpesvirus)………. Vaksin DNA pada Ikan……… Vaksin DNA Penyandi Glikoprotein……… Konstruksi Vaksin………. Respons Imun pada Ikan……….………... Respons Imun Non-Spesifik……….. Respons Imun Spesifik….………..
BAHAN DAN METODE………. Tahap I: Pembuatan Konstruksi Vaksin………. DNA KHV dan Plasmid….………... Disain Primer………. Isolasi Gen Penyandi Glikoprotein………. Ligasi Fragmen Glikoprotein dan pGEMT-Easy……….. Transformasi pada E.coli Kompeten……….. Verifikasi Hasil Transformasi……….. Isolasi Plasmid……….. Pembuatan Konstruksi Vaksin DNA……….. Tahap II: Uji Ekspresi Vaksin………. a. Isolasi RNA………….……….………. b. Sintesis cDNA dan Amplifikasi PCR……… Tahap III: Uji Tantang Skala Laboratorium……….……….. Preparasi Virus……….. Uji Tantang Tahap …I………..……….. a. Pengamatan RPS (Relative Percent Survival)……….. b. Pengambilan Sampel Darah………...
ii
c. Penghitungan Jumlah Leukosit Total………….………. d. Aktivitas Fagositosis………. Uji Tantang Tahap II…..………….………. Uji Keamanan Vaksin……….. Analisis Data………..…………
HASIL DAN PEMBAHASAN……… Tahap I: Pembuatan Konstruksi Vaksin……… Isolasi DNA Glikoprotein Koi Herpesvirus………... Transformasi pada E.coli Kompeten……….. Pembuatan Konstruksi Vaksin DNA……….. Tahap II: Uji Ekspresi………..……… Tahap III: Uji Tantang Skala Laboratorium……….………. Uji Tantang I: RPS dan Aktivitas Sel Darah Putih………...………… Aktivitas Sel Darah Putih…..………. Leukosit Total……… Aktivitas Fagositosis……….… Uji Tantang II: RPS dan Keamanan Vaksin…………..…….……….. Keamanan Vaksin……….
iii DAFTAR TABEL
1. Penyebaran virus KHV di empat benua di dunia………. 2. Sensitivitas KHV terhadap kondisi fisika dan kimia.……….. 3. Perbedaan antara imunitas ikan dan mamalia………. 4. Kelangsungan hidup relatif ikan pasca uji tantang I dengan virus
KHV………. 5. Kelangsungan hidup relatif ikan pasca uji tantang II dengan virus
KHV………..
Hal.
11 12 19
46
iv DAFTAR GAMBAR
1. Penampang melintang kelompok herpesvirus………. 2. Koi herpesvirus yang diambil dari koi fin cell, diamati dengan
mikroskop elektron yang memperlihatkan adanya kapsid berbentuk simetri ikosahedral (tanda panah pada gambar A) dan beramplop (tanda panah pada gambar B), diwarnai dengan pewarnaan negative phosphotungstat 2 % (Hutoran 2005)... 3. Skema proteksi non-spesifik dan spesifik ikan rainbow trout yang
divaksinasi dengan vaksin DNA menggunakan gen glikoprotein VHS……… 4. Skema gambaran partikel Rhabdovirus (a), vaksin plasmid (b) dan
protein G virus ……….. 5. Fragmen DNA GP25 yang diisolasi dari insang yang diduga kuat
terinfeksi virus KHV (a); Fragmen GP25 KHV yang siap dipurifikasi (b); Fragmen DNA GP25 yang telah dipurifikasi (c) yang divisualisasikan di agarose dengan konsentrasi 0,7% dengan M adalah marker DNA (2 log ladder, Biolabs, England)………...……… 6. Hasil transformasi bakteri E.coli DH5α berupa koloni biru (b) dan
koloni putih (p)……….……… 7. Hasil verifikasi dengan metode cracking, dimana tanda panah
( ) menunjukkan hasil berupa plasmid yang berukuran lebih besar daripada marker yaitu koloni biru (tanda + )………. 8. Fragmen DNA hasil verifikasi dengan menggunakan PCR. Tanda (
) menunjukkan fragmen GP25 yang tersisip dip T-GP25………. 9. Hasil pengurutan sekuen GP25. Huruf miring di awal urutan
nukleotida adalah primer forward (F-GP), sedangkan huruf miring di akhir adalah urutan primer reverse (R-GP)... 10. Hasil penyejajaran (allignment) parsial GP25 asal Indonesia dengan GP25 KHV asal Israel, Jepang dan Amerika Serikat... 11. Hasil analisis kemiripan gen GP25 virus KHV asal Indonesia
dengan virus KHV asal Jepang (strain TUMST1), Amerika Serikat (strain KHV-U), dan virus KHV asal Israel (strain KHV-I). Persentase kemiripan ditunjukkan oleh kolom Identity……… 12. Hasil digesti pActD6 dengan enzim SalI menjadi pAct dan D6, pAct
v
13. Hasil digesti pT-GP25 dengan enzim SalI menjadi pT-Easy dan gen glikoprotein (GP25); dimana D adalah fragmen hasil digesti sedangkan K adalah kontrol plasmid yang tidak didigesti…………
14. Hasil verifikasi dengan metode cracking terhadap hasil transformasi tahap kedua yang menggunakan plasmid pAct. Tanda panah ( ) menunjukkan klon bakteri yang dipilih untuk diuji lanjut; tanda positif (+) menunjukkan marker yang berasal dari koloni biru………..
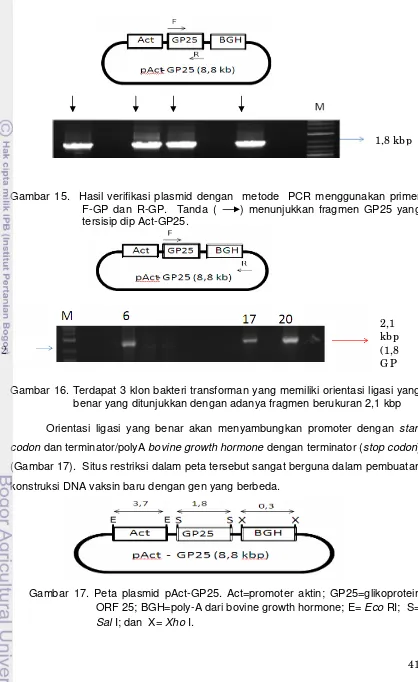
15. Hasil verifikasi plasmid dengan metode PCR menghasilkan produk sebesar 1,8 kbp. Tanda ( ) menunjukkan fragmen GP25 yang tersisip dip Act-GP25………. 16. Terdapat 3 klon bakteri transforman yang memiliki orientasi ligasi
yang benar yang ditunjukkan dengan adanya fragmen berukuran 2,1 kb………. 17. Peta plasmid pAct-GP25. Act=promoter aktin; GP25=glikoprotein
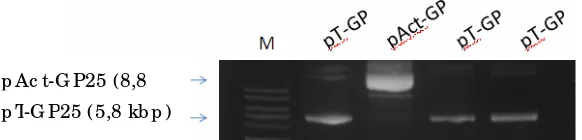
ORF 25; BGH=poly-A dari bovine growth hormone; E= Eco RI; S= Sal I; dan X= Xho I... 18. Perbandingan ukuran fragmen pT-GP yang berukuran 5,8 kb
(3 kb pT-Easy+1,8 kb GP25) dan pAct-GP25 yang berukuran 8,8 kb (3 kb pBlueScript+3,7mBA+1,8 GP+0.3). M adalah marker DNA……….. 19. Ekspresi gen GFP pada ginjal (G), insang (I), limpa (L), dan otot (O)
ikan mas pada jam ke-24 (A) dan seminggu (B) setelah injeksi dengan pAct-GFP. Ekspresi gen β-aktin sebagai kontrol internal terdeteksi pada semua jaringan (C). K+: kontrol positif plasmid pAct-GFP, K-: kontrol negatif tanpa cetakan DNA... 20. Ekspresi gen GP25 pada otot yang tidak diinjeksi (O-) dan otot
yang injeksi (O+), insang (I), limpa (L) dan ginjal (G) pada hari keempat belas (setelah dua minggu) setelah injeksi. Ekspresi gen β-aktin sebagai kontrol internal terdeteksi pada semua jaringan dengan level yang relatif sama (C). Angka di sebelah kanan gambar merupakan ukuran DNA produk PCR... 21. Grafik kelangsungan hidup ikan setelah diuji tantang dengan virus
KHV……… 22. Tren jumlah leukosit total ikan mas pada masing-masing perlakuan
vaksinasi dosis 2,5 µg/100µl (A); dosis 7,5 µg/100µl (B);dosis 12,5 µg/100µl (C)dantanpa vaksinasi (K)……….
vi
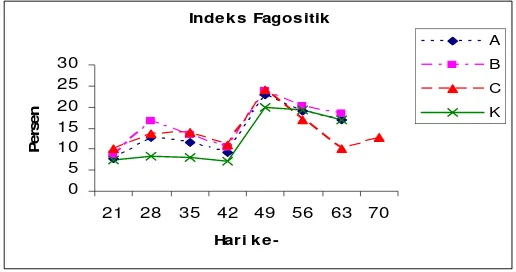
23. Aktivitas fagositosis pada perlakuan vaksinasi dengan dosis 12,5 µg/100µl (perlakuan A), 7,5 µg/100µl (perlakuan B), 12,5 µg/100µl (perlakuan C) dan tanpa vaksinasi (perlakuan K)…….……….. 24. Fagositosis yang dilakukan oleh limfosit (a), monosit (b), Neutrofil
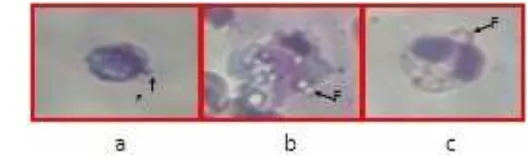
(c). Huruf F dan tanda panah menunjukkan tonjolan yang berisi benda asing yang telah difagositosis oleh sel-sel fagosit………….. 25. Grafik kelangsungan hidup ikan setelah diuji tantang dengan virus
KHV……….
Hal.
49
50
vii DAFTAR LAMPIRAN
1. Ilustrasi ligasi antara GP25 dan pT-Easy... 2. Skema pembuautan konstruksi vaksin... 3. Metode kultur cair perbanyakan bakteri dan isolasi plasmid………. 4. Peta plasmid pBluescript……… 5. Prosedur pembuatan preparat jaringan……… 6. Analisis data RPS (Relative Percent Survival) dengan ANOVA dan
uji lanjut dengan BNT (beda nyata terkecil)………. 7. Suhu Air setelah uji tantang dengan virus KHV………... 8. Jaringan otot dan insang yang tidak divaksinasi dan yang
divaksinasi (Pewarnaan hematoksilin dan eosin)……… 9. Jaringan limpa dan ginjal yang tidak divaksnasi dan yang
divaksinasi (Pewarnaan hematoksilin dan eosin)………
Hal. 64 65 66 67 68
69 70
72
PENDAHULUAN
Latar Belakang
Koi herpesvirus (KHV) adalah virus yang menginfeksi ikan mas dan koi dan bersosiasi dengan kematian massal (Hedrick et al. 2000). Virus ini pertama kali teridentifikasi pada tahun 1998 sebagai penyebab kematian massal ikan koi baik stadia juvenil maupun dewasa yang dibudidayakan di Israel, Amerika Serikat dan Jerman (Hedrick et al. 1999; Bretzinger et al.1999). Penyebaran virus ini sudah mencapai Eropa, Jepang, Indonesia, Afrika Selatan, Thailand, Taiwan, Cina dan Malaysia (Haenen et al. 2004; Sano et al. 2004; Tu et al. 2004). Virus KHV masuk ke Indonesia pada tahun 2002 melalui perdagangan ikan lintas negara (Sunarto et al. 2005). Penyakit akibat virus yang sangat menular ini telah menyebabkan kerugian finansial pada industri budidaya ikan mas dan koi (Hedrick 1996; Haenen et al. 2004).
Sejak terjangkit pertama kali di Blitar, Jawa Timur, penyakit ini telah menyebar ke hampir semua daerah di Indonesia. Virus ini mengakibatkan kematian massal, yaitu kematian mencapai 80-95 % populasi sehingga berdampak pada kerugian ekonomi dan sosial. Kerugian secara materi akibat penyakit ini mencapai 15 milyar rupiah dalam tiga bulan pertama sejak kejadian penyakit ditemukan, yaitu bulan Maret sampai September 2002 (Sunarto 2004). Dinas Perikanan Jawa Barat menemukan data bahwa sepanjang tahun 2002 kerugian yang ditimbulkan oleh penyakit akibat KHV ini mencapai 100 milyar rupiah (Sunarto & Kusrini 2006). Kerugian total sampai tahun 2006 mencapai 250 milyar rupiah. Namun jumlah ini merupakan perhitungan berdasarkan kasus yang berhasil diketahui maupun dari laporan masyarakat. Jumlah kerugian yang ditimbulkan seperti fenomena gunung es yaitu kemungkinan kerugian yang ditimbulkan bisa lebih besar dari jumlah tersebut (Taukhid 2010; komunikasi pribadi).
2
sampai saat ini terutama pada musim hujan, meskipun kematian yang ditimbulkan tidak ekstrim seperti tahun-tahun sebelumnya (Sunarto & Kusrini 2006). Untuk daerah endemik seperti Cirata maka kematian massal terjadi sekitar 40-60% dan biasanya terjadi pada pertengahan dan akhir tahun. Mortalitas ikan mas yang dibudidayakan di kolam air deras lebih tinggi yaitu mencapai 70% dan terjadi sepanjang musim, sedangkan di daerah yang jarang ada kasus atau baru pertama terjadi kasus maka kematian bisa lebih tinggi dari 60%. Mortalitas ikan mas yang dibudidayakan di keramba di Sungai Mahakam mencapai 70-80% (Taukhid 2010; komunikasi pribadi).
Meskipun muncul pertama kali pada tahun 2002, namun belum ada langkah strategis yang dapat menanggulangi wabah penyakit ini. Hal ini menyebabkan Indonesia belum aman dari ancaman wabah KHV. Oleh karena itu Komisi Kesehatan Ikan dan Lingkungan - Kementerian Kelautan dan Perikanan menetapkan penyakit yang diakibatkan oleh infeksi KHV termasuk dalam lima penyakit utama pada budidaya ikan di Indonesia. Kelima penyakit tersebut adalah penyakit yang diakibatkan oleh parasit Ichtyophtirius multifilis pada ikan air tawar, penyakit MAS (motile aeromonad septicaemia) oleh infeksi bakteri Aeromonas hydrophila, penyakit akibat infeksi virus KHV (Koi Herpesvirus) pada ikan mas dan koi, penyakit infeksi virus WSSV (White Spot Syndrome Virus) pada udang dan penyakit VNN (viral nervous necrosis) akibat infeksi virus VNN pada ikan kerapu. Kelima penyakit utama
ini memerlukan perhatian yang sungguh-sungguh karena langkah untuk menanggulangi penyakit pada komoditas strategis tersebut belum dapat menuntaskan masalah penyakit tersebut.
Upaya penanggulangan wabah KHV di daerah telah dilakukan dengan menggunakan bahan-bahan kimia. Penanggulangan dengan metode tersebut terbukti tidak efektif karena hanya mambantu mengatasi infeksi sekunder oleh bakteri, fungi atau parasit. Ketidakefektifan tersebut disebabkan karena penyebaran KHV dalam tubuh ikan berlangsung antar sel sehingga virus tidak perlu keluar sel dan masuk ke dalam sistem sirkulasi tubuh inang untuk penyebarannya. Oleh karena itu sifat dari golongan virus herpes tersebut adalah berasosiasi kuat dengan sel (highly cell associated) dan bersifat laten yaitu seumur hidup berada dalam tubuh inangnya (Arvin
1996).
3
ekstrak mengkudu (pace) dan cacahan buah maja. Kendala dalam metode ini adalah terbatasnya pengkajian secara ilmiah terhadap penggunaan bahan-bahan alami baik mengenai kandungan senyawa aktif maupun mekanisme kerja bahan-bahan ini pada tubuh ikan. Mekanisme bahan-bahan alami ini terbatas pada respon non spesifik sehingga tidak spesifik dalam menghadapi infeksi KHV.
Penanggulangan yang dilakukan oleh pihak-pihak terkait adalah penerapan biosecurity yang bertujuan mengamankan wilayah budidaya dari sebaran atau
tersebarnya penyakit dari wilayah tersebut. Penanggulangan ini mudah dilakukan pada sistem budidaya dalam ekosistem tertutup semisal kolam, artinya tidak ada kaitan satu unit kolam dengan kolam lainnya. Konsep biosecurity ini tidak dapat diterapkan pada budidaya dengan sistem terbuka pada danau/waduk. Hal ini disebabkan karena unit jaring apung pada danau/waduk tersebut saling berkaitan antara satu dengan yang lain sementara air yang digunakan sebagai media budidaya adalah air yang sama dan menyatu dalam satu wadah yaitu danau/waduk. Dampak dari suhu media budidaya yang rendah terhadap aktivitas virus KHV maupun menurunnya daya tahan tubuh ikan tidak cukup diantisipasi dengan penerapan biosecurity ini.
Mengingat sifat virus herpes yang berasosiasi kuat dengan sel dan sulit untuk ditanggulangi maka langkah pencegahan mutlak perlu dilakukan. Salah satu langkah pencegahan yang bisa dilakukan adalah dengan vaksinasi. Vaksin yang diberikan dapat berupa vaksin konvensional dan vaksin rekombinan. Vaksin konvensional yang diberikan berupa virus dilemahkan maupun dimatikan. Vaksin rekombinan merupakan hasil rekayasa genetika dimana sekuen gen virus yang bersifat imunogenik disisipkan ke plasmid dan plasmid ini selanjutnya dipropagasi di bakteri E.coli. Vaksin rekombinan ini berupa protein rekombinan yang bersifat imunogenik.
Vaksin DNA merupakan hasil terobosan teknik eksperimental untuk melindungi organisme melawan penyakit dengan cara menginjeksikan DNA murni (naked DNA) untuk membangkitkan respon kekebalan. Vaksin revolusioner yang muncul tahun 1992 ini sudah dikembangkan untuk meningkatkan kekebalan tubuh terhadap penyakit baik pada manusia maupun hewan, termasuk ikan.
4
atenuasi/pelemahan yang dilakukan tidak sempurna. Oleh karena itu Rakus (2008) tidak menganjurkan digunakannya vaksin KV3 ini pada ikan. Rakus juga menyebutkan bahwa Undang-Undang Uni Eropa juga tidak mengizinkan digunakannya vaksin ini. Vaksin yang dilemahkan memang memiliki keterbatasan yang dapat disempurnakan oleh vaksin DNA.
Vaksin DNA dapat dijadikan sebagai alternatif penanggulangan penyakit akibat KHV pada ikan mas dan koi untuk mengurangi cadangan devisa yang dibelanjakan untuk pengadaan vaksin KHV dari luar negeri. Direktorat Kesehatan Ikan dan Lingkungan, Direktorat Jenderal Budidaya, Kementrian Kelautan dan Perikanan menganggap keberadaan vaksin untuk mencegah infeksi KHV menjadi sesuatu yang sangat penting dan mendesak. Pengambilan keputusan untuk mengimpor vaksin dari luar negeri yang sekarang sedang diuji efikasi harus terintegrasi dengan kebijakan pengembangan vaksin KHV dengan menggunakan isolat lokal. Keunggulan vaksin isolat lokal adalah dapat membangkitkan sistem imun secara efektif karena vaksin yang digunakan berasal dari virus yang sama dengan virus yang menyebabkan infeksi pada ikan mas dan koi. Kehomologan antara antibodi yang dihasilkan setelah vaksinasi dengan antigen sebagai sumber vaksin maupun penyebab penyakit merupakan syarat penting untuk mencapai keberhasilan vaksinasi. Dikembangkannya vaksin DNA di dalam negeri dengan menggunakan isolat lokal akan mencegah ketergantungan komponen budidaya ikan dari luar negeri. Ketergantungan ini dalam jangka panjang dapat mengancam kedaulatan pangan Indonesia, khususnya sumber protein asal ikan.
Keunggulan vaksin DNA (Lorenzen & LaPatra 2005) yang dapat dijadikan sebagai alasan untuk mengembangkannya adalah:
1. Bersifat generik dan sederhana
2. Aman dan tidak menimbulkan resiko terinfeksi penyakit
3. Kombinasi keuntungan dari vaksin tradisional (inactivated vaccine) dan yang dilemahkan (attenuated vaccine)
4. Dapat mencapai keberhasilan tujuan vaksinasi ketika vaksinasi konvensional gagal
5
7. Memungkinkan vaksinasi multivalen yaitu dengan mencampur vaksin DNA untuk lebih dari satu jenis penyakit melalui vaksinasi yang dilakukan secara bersamaan
8. Memberikan proteksi yang baik apabila diberikan pada stadia awal
9. Proteksi dapat diinduksi dalam waktu singkat dan memberikan efek proteksi dalam jangka waktu lama
10. Dapat memberikan proteksi baik dalam suhu rendah maupun tinggi 11. Dapat memberikan proteksi pada heterologous strain pathogen
12. Dapat menyediakan vaksin untuk patogen baru dalam waktu cepat dan biaya relatif lebih murah
13. Produk murni memiliki stabilitas yang tinggi
14. Biaya produksi relatif murah dan mudah diproduksi
Hirono (2005) menjelaskan bahwa proteksi yang diberikan oleh vaksin yang dilemahkan cukup tinggi baik dalam membangkitkan kekebalan seluler maupun humoral, akan tetapi berpotensi untuk terjadinya infeksi. Kelemahan ini dapat diperbaiki oleh vaksin DNA yang mampu membangkitkan respon kekebalan seluler maupun humoral akan tetapi tidak menimbulkan terjadinya infeksi karena yang dimasukkan hanya bagian tertentu saja dari virus, dalam hal ini gen glikoprotein.
Banyaknya kelebihan yang dimiliki oleh vaksin DNA memungkinkan vaksin ini punya peluang untuk diterapkan di bidang perikanan. Vaksin DNA dengan menggunakan gen glikoprotein virus telah dikembangkan untuk ikan rainbow trout oleh Corbeil et al. (1999); pada ikan salmon oleh Lapatra et al. (2001). Zheng et al. (2006) juga sedang mengembangkan vaksin DNA untuk ikan sebelah / flounder Paralichthys olivaceus di Cina. TUMST (Tokyo University of Marine Science and Technology) juga telah mengembangkan vaksin DNA KHV untuk ikan mas dan koi di Jepang dengan menggunakan isolat asal Jepang (Hirono 2008; komunikasi pribadi). Pengembangan vaksin DNA penyandi glikoprotein KHV dengan menggunakan isolat virus asal Indonesia mendesak untuk dilakukan. Penggunaan teknologi molekuler dalam pengendalian penyakit dalam budidaya ikan diharapkan dapat meminimalkan resiko kerugian akibat kematian dengan cara yang lebih mudah dan biaya yang relatif murah dibandingkan vaksin konvensional maupun protein rekombinan.
6
DNA; tahap kedua adalah uji ekspresi yaitu uji aktivitas promoter vaksin; dan tahap ketiga adalah uji tantang skala laboratorium.
Tujuan
Penelitian ini bertujuan untuk:
a. Membuat konstruksi vaksin DNA dengan menggunakan gen glikoprotein KHV b. Menguji ekspresi plasmid vaksin
c. Menguji vaksin DNA KHV dan peranannya dalam meningkatkan kelangsungan hidup relatif.
Manfaat Penelitian
Penelitian ini diharapkan dapat menambah khasanah metode dalam penanggulangan penyakit pada ikan pada umumnya karena penelitian tentang vaksin DNA untuk penyakit pada ikan baru pertama dilakukan di Indonesia. Manfaat lain yang didapatkan adalah diperolehnya alternatif pencegahan penyakit khususnya akibat infeksi KHV. Seperti diketahui bahwa panyakit karena virus ini relatif sulit untuk ditanggulangi sehingga memerlukan penanganan yang mampu meningkatkan respons kekebalan ikan yang bersifat spesifik mencakup seluler maupun humoral. Penanganan yang tepat akan meningkatkan produktivitas ikan budidaya khususnya ikan mas dan koi.
Aspek Kebaruan
Aspek kebaruan dari penelitian ini adalah:
a. Vaksin DNA ini menggunakan gen virus KHV isolat lokal sebagai sumber DNA yang disisipkan ke plasmid vaksin. Perbedaan strain virus mengandung konsekuensi adanya perbedaan sekuen gen penyandi protein tertentu misalnya glikoprotein. Gen penyandi glikoprotein tertama ORF 25 (yang merupakan bagian virus yang bersifat imunogenik) dari KHV asal Indonesia memiliki perbedaan susunan asam amino dengan KHV asal Jepang, Amerika Serikat maupun Israel.
7
dengan menggunakan promoter dari ikan lebih kuat dibandingkan dengan menggunakan promoter dari non-ikan.
Hipotesis
TINJAUAN PUSTAKA
Penyakit Infeksi Koi Herpesvirus (KHV)
Ikan mas (Cyprinus carpio) adalah ikan yang dibudidayakan secara luas sebagai bahan makanan. Sebanyak 1.5 juta metrik ton diproduksi terutama di Asia maupun Eropa (www.fao.org). Budidaya ikan mas memberikan sumbangan protein untuk penduduk yang berdiam di Asia, Eropa dan Timur Tengah (Aoki et al. 2007).
Koi herpesvirus diidentifikasi pertama kali tahun 1998 yang menyebabkan kematian massal pada ikan mas budidaya di Israel (Gilad et al. 2002) dan Amerika Serikat (Gray et al. 2002). Carp nephritis and gill necrosis virus (CNGV) adalah nama awal virus yang berasal dari virus DNA yang morfologinya mirip dengan anggota kelompok Herpesviridae yang nama lainnya adalah koi herpesvirus dan Cyprinid herpesvirus (Dishon et al. 2005). Nama lain dari virus KHV adalah Cyprinid Herpesvirus 3 atau CyHV-3 (Aoki et al. 2007). Virus ini masuk ke Indonesia pada tahun 2002 melalui perdagangan ikan koi (Sunarto et al. 2004).
Dari percobaan kohabitasi antara ikan sehat dan ikan terinfeksi KHV yang dilakukan oleh Hutoran et al. (2005) diperoleh hasil bahwa ikan yang sakit mengalami ganggunan berupa gerakan yang tidak terkoordinasi dan berenang tidak beraturan yang merupakan tanda-tanda adanya gangguan saraf (neurological disorder). Gangguan ini diperjelas dengan berkurangnya frekuensi gerakan ekor dan kehilangan keseimbangan pada beberapa ikan. Penyebaran penyakit ini melalui air dan bersifat sangat menular.
Menilik dari nama gejala penyakit yang ditimbulkan, virus ini memang menginfeksi terutama pada bagian insang dan ginjal ikan. Dari kajian histopatologi pada insang, tampak jelas bahwa virus ini mengakibatkan inflamasi pada renal tubul ginjal dan mengakibatkan sel-sel yang terinfeksi mengalami pembentukan badan inklusi pada inti selnya. Kajian histopatologi insang ikan yang sakit menunjukkan bahwa terdapat sel-sel inflamasi di insang dan epitel insang mengalami hiperplasia. Kajian dengan menggunakan indirect immunofluorescen microscopy terhadap insang, ginjal, otak dan hati menunjukkan bahwa virus KHV terakumulasi pada insang dan ginjal (Pikarsky et al. 2005).
9
Metode LAMP tidak memerlukan mesin PCR dalam mengamplifikasi DNA, akan tetapi memerlukan Bst DNA polymerase, dua primer inner, dua primer outer dan dua primer loop untuk mengamplifikasi DNA. Reaksi dilakukan pada suhu 65 oC selama 60 menit. Dari 50 ekor ikan sampel yang diperiksa dengan PCR, 37 ekor dinyatakan positif KHV sedang 13 ekor negatif. Melalui metode LAMP semua sampel dinyatakan positif.
Virus KHV (Koi Herpesvirus)
Nama herpes berasal dari bahasa Yunani yaitu herpein yang berarti kronis / laten / infeksi yang selalu terjadi. Sebanyak 100 macam herpesvirus telah diisolasi. Virus ini memiliki ukuran DNA genom yang besar mencapai 235 kbp dan tersusun dari 35 polipeptida. Secara umum kelompok herpesvirus ini berukuran 180-200 nm, memiliki amplop yang tersusun dari glikoprotein. Pada bagian antara amplop dan kapsid terdapat tegument yang tersusun dari protein. Kapsid berbentuk ikosahedral dengan diameter 95-105 nm. Pada bagian pusat terdapat DNA yang dikelilingi oleh nukleokapsid. Bentuk DNA genom kelompok herpesvirus adalah berutas ganda yang berukuran 130-230 kbp (www.herpes.org).
Struktur virus herpes dapat dilihat pada gambar berikut ini:
Gambar 1. Penampang melintang kelompok herpesvirus (pathmicro.med.sc.edu/virol/herpes)
10
(channel catfish virus). KHV memiliki kapsid simetri ikosahedral dengan diameter 100-110 nm, sedangkan virion matang memiliki amplop yang longgar sehingga ukuran diameternya menjadi 170-230 nm. Selain itu juga terdapat benang-benang penyangga seperti struktur tegument pada permukaan inti yang mirip dengan kelompok Herpesvirus (Pokorova et al. 2005).
Hutoran et al. (2005) melaporkan bahwa virus yang diisolasi dari ikan mas yang mengalami kematian massal di Israel memiliki ukuran DNA genom besar yaitu 277 kbp, lebih besar dari ukuran DNA genom kelompok Herpesviridae. Penemuan yang lain adalah hanya sebagian kecil fragmen yaitu 16-45 bp yang mirip dengan beberapa DNA genom virus yang lain. Karakter koi herpesvirus yang dilaporakan dari penelitian Hutoran adalah:
1. Virus sangat menular 2. Transmisi melalui air
3. Terinduksi menjadi penyakit apabila temperatur tertekan sehingga berada pada level 18-24 oC, baik pada skala penelitian di laboratorium (indoor) maupun di kolam (outdoor)
4. Memiliki inang yang terbatas yaitu ikan mas dan koi
5. Tidak dapat dipropagasi pada epitelioma pappilosum cell (EPC), tetapi dapat dipropagasi pada koi fin cell (KFC)
6. Morfologi dan diameter konsisten sesuai dengan herpesvirus meskipun kadang-kadang pada bagian inti virus (core) berisi massa non-simetri berukuran kecil
7. Analisis terhadap sekuen fragmen DNA tidak menunjukkan adanya kesamaan dengan genom virus yang sudah diketahui
8. Virus memiliki molekul DNA utas ganda yang sangat besar yaitu 277 kbp, lebih besar dibanding dengan genom herpesvirus yang sudah ada.
11
Tabel 1. Penyebaran virus KHV di empat benua di dunia
Sumber: Crane et al. (2004)
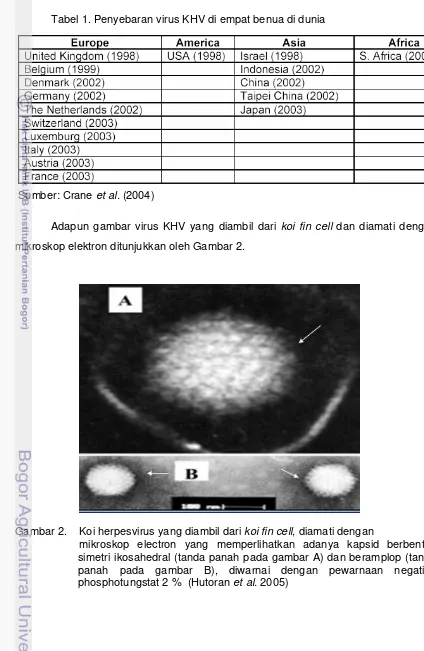
Adapun gambar virus KHV yang diambil dari koi fin cell dan diamati dengan mikroskop elektron ditunjukkan oleh Gambar 2.
Gambar 2. Koi herpesvirus yang diambil dari koi fin cell, diamati dengan
12
Crane et al. (2004) melaporkan bahwa virus KHV memiliki sensitifitas terhadap kondisi fisika-kimia. Namun penelitian berikutnya secara detil masih perlu dilakukan untuk mengungkap kondisi fisika-kimia yang berpengaruh terhadap sensitifitas virus.
Tabel 2. Sensitivitas KHV terhadap kondisi fisika dan kimia Sensitifitas/Kerentanan Terhadap Kondisi Fisika-Kimia
Temperatur Infektifitas virus hilang setelah berada pada suhu 35 oC selama dua hari atau pada suhu 60 oC selama 30 menit
pH Infektifitas hilang pada pH <3 atau pH>11
Bahan kimia Sensitif terhadap kloroform (diasumsikan sensitive juga terhadap bahan pelarut lemak yang lain)
Desinfektan Desinfektan yang dianjurkan merujuk pada
ketetapan Office International des Epizooties, OIE (2003), perlu diteliti lebijh lanjut
Kelangsungan hidup Virus dapat bertahan hidup di air selama 20 jam dan lebih lama pada kolam dengan kondisi buruk, ada yang menyebutkan juga bahwa virus hanya mampu bertahan paling lama empat jam.
Sumber: Crane et al. (2004)
KHV merupakan virus yang baru dikenal keberadaannya ketika menyebabkan kematian massal pada budidaya ikan mas dan Koi di Israel tahun 1998. Sebelum disekuensing, KHV masih diberi nama CNGV (carp nephritis and gill necrosis virus) (Dishon et al. 2005) dan masih diduga merupakan anggota family Herpesviridae karena kemiripannya dengan family Herpesviridae yaitu berupa DNA berutas ganda yang dibungkus oleh kapsid ikosahedral , lapisan protein tegument dan amplop virus yang tersusun oleh glikoprotein. Berdasarkan data yang ada di GeneBank, dari seluruh KHV yang ada sekarang baru terdapat tiga genom virus yang sudah disekuensing secara lengkap (genom). Tiga genom KHV tersebut berasal dari Jepang, Israel dan Amerika Serikat (Aoki et al. 2007). Genom KHV memiliki ukuran sekitar 295 kbp dengan perincian sebesar 295.271 bp untuk KHV dari Jepang, 295.146 bp untuk KHV dari Amerika Serikat dan 295.138 bp untuk KHV yang berasal dari Israel. Dari ukuran tersebut tampak ada perbedaan jumlah basa nitrogen dan bervariasi untuk masing-masing strain/serotype virus dimana KHV dari Jepang memiliki ukuran genom yang paling besar diikuti Amerika Serikat dan Israel.
13
diduga sebesar 277 kb ketika diketahui pertama kali, lebih besar dari ukuran molekul anggota family Herpesviridae yang sudah ada yaitu 125-245 kb. Di samping itu kegagalan dalam membuktikan adanya hubungan kekerabatan (secara genetik) antara KHV dengan family Herpesviridae juga menyebabkan keraguan uuntuk memasukkan KHV ke dalam family ini. Setelah disekuensing oleh Aoki et al. (2007) baru dapat dipastikan bahwa KHV memang bagian dari family Herpesviridae. KHV memiliki hubungan erat dengan Cyprinid Herpesvirus 1 dan 2 (CyHV-1 dan CyHV-2) yang menyebabkan penyakit carp pox dan hematopoietic necrosis pada ikan mas koki (gold fish). Selain itu KHV juga memiliki kekerabatan dengan Ictalurid herpesviridae (IcHV-1) dan ranid HV-1 (penyebab tumor pada katak). Selanjutnya KHV telah diusulkan secara formal ke dalam anggota Alloherpesviridae dengan nama spesies Cyprinid Herpesvirus 3 atau disingkat menjadi CyHV-3 (Aoki et al. 2007).
Vaksin DNA pada Ikan
Ikan adalah organisme yang mudah terifeksi penyakit yang diakibatkan oleh parasit, bakteri, cendawan dan virus apabila dibudidayakan dalam sistem terkontrol. Penanggulangan penyakit dengan menggunakan bahan kimia termasuk antibiotik memberikan dampak yang tidak baik bagi lingkungan maupun manusia yang mengonsumsinya. Penyakit yang disebabkan oleh virus relatif lebih ditangani karena tidak ada treatmen komersial maupun kemoterapetan yang ekonomis yang bermanfaat dalam penanggulangan penyakit infeksi KHV. Oleh karena itu langkah-langkah yang perlu dilakukan adalah langkah-langkah yang bersifat profilaksis misalnya vaksinasi dan diagnosis penyakit dalam rangka pencegahan terjadinya wabah penyakit (Leong et al. dalam www.nps.ars.usda.gov).
Vaksinasi mampu meningkatkan produktifitas ikan salmon secara signifikan di Norwegia. Produksi ikan salmon pada tahun 1987 sebesar 65,000 metrik ton dan meningkat menjadi 700,000 metrik ton pada tahun 2007. Penggunaan vaksin juga mereduksi penggunaan antibiotik dari 48,500 kg menjadi 649 kg (Gravningen & Berntsen 2008).
14
terhadap lingkungan serta peningkatan mutu bahan pangan dari ikan karena adanya minimalisasi dalam penggunaan antibotik (Lorenzen & LaPatra 2005).
Vaksin virus untuk ikan jarang dijual secara komersial. Di Amerika Serikat sendiri agak sulit untuk mendapatkan lisensi peredaran karena prosesnya panjang dan biayanya mahal serta efikasi vaksin yang tidak konsisten. Kendala yang lain adalah masalah keamanan vaksin virus yang diatenuasi masih dipertanyakan karena memiliki potensi untuk bangkit kembali dan menginfeksi inang yang divaksinasi (Leong et al. dalam www.nps.ars.usda.gov). Berkembangnya penyediaan vaksin untuk menanggulangi penyakit yang diakibatkan oleh viral haemorrhagic septicaemia virus (VHSV), infectious haematopoietic necrosis virus (IHNV), infectious pancreatic
necrosis virus (IPNV) dan infectious salmon anemia virus (ISAV) cukup memberikan perlindungan bagi budidaya ikan salmon. Di sisi lain, penumbuhan virus bakal vaksin di sel kultur ikan memerlukan biaya yang tidak sedikit. Untuk efisiensi biaya budidaya maka vaksin DNA perlu dikembangkan lebih lanjut. Pada level eksperimen vaksin ini dapat melawan virus dengan tingkat paling efisien. Vaksin ini berbasis pada plasmid DNA yang membawa sisipan gen misalnya glikoprotein dan disertai dengan promoter dan terminator/polyA untuk keperluan ekspresi di ikan (Lorenzen & LaPatra 2005).
Hirono (2005) mengelompokkan perkembangan vaksin pada ikan menjadi tiga generasi. Generasi pertama adalah vaksin konvensional yang dibagi menjadi dua kelompok yaitu vaksin yang diinaktivasi/dimatikan (inactivated vaccine) dan vaksin hidup yang dilemahkan (live attenuated vaccine). Vaksin generasi kedua adalah vaksin protein rekombinan (recombinant protein vaccine) dan vaksin generasi ketiga adalah vaksin DNA (DNA vaccine).
15
mampu mengaktivasi sistem kekebalan baik humoral maupun seluler, sedang kelemahannya adalah terbatasnya protein yang bersifat imunogenik.
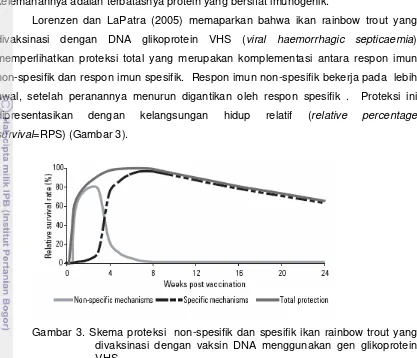
[image:36.612.105.522.96.454.2]Lorenzen dan LaPatra (2005) memaparkan bahwa ikan rainbow trout yang divaksinasi dengan DNA glikoprotein VHS (viral haemorrhagic septicaemia) memperlihatkan proteksi total yang merupakan komplementasi antara respon imun non-spesifik dan respon imun spesifik. Respon imun non-spesifik bekerja pada lebih awal, setelah peranannya menurun digantikan oleh respon spesifik . Proteksi ini dipresentasikan dengan kelangsungan hidup relatif (relative percentage survival=RPS) (Gambar 3).
Gambar 3. Skema proteksi non-spesifik dan spesifik ikan rainbow trout yang divaksinasi dengan vaksin DNA menggunakan gen glikoprotein VHS
Vaksin DNA Penyandi Glikoprotein
16
Aplikasi vaksin DNA yang mengandung sisipan gen glikoprotein dapat menginduksi terbentuknya alpha/beta interferon pada ikan rainbow trout yang diuji tantang setelah 30 atau 70 hari setelah vaksinasi. Proteksi yang diberikan cukup lama karena respon imun yang terbentuk bersifat spesifik yaitu terhadap gen G. Vaksin DNA menginduksi perlindungan antiviral yang bersifat non-spesifik pada mulanya yang dimediasi oleh alpha/beta interferon, berikutnya baru terbentuk respon imun yang bersifat spesifik. Vaksin DNA dengan menggunakan gen glikoprotein untuk mencegah penyakit IHHNV pada ikan rainbow tout mampu memberikan proteksi secara signifikan empat hari setelah vaksinasi. Uji tantang setelah 28 hari vaksinasi memberikan rentang yang paling tinggi antara kematian ikan kontrol dan ikan perlakuan, dibandingkan dengan uji tantang pada 1, 7, 14 dan 21 hari (Kim et al. 2000).
Vaksin DNA untuk penyakit infeksi SVCV (spring viremia carp virus) yang mengandung gen glikoprotein lengkap (full length) menghasilkan nilai relative percent survival (RPS) sebesar 48% (primer tidak disebutkan). Perlakuan vaksinasi 10 µg
pada penelitian ini dapat menginduksi respon kekebalan berperantara sel (CMI=cell mediated immunity), namun antibody tidak terdeteksi. Penelitian tentang vaksin DNA
untuk SVCV pada ikan mas merupakan penelitian yang pertama kali dilakukan (Kanellos et al. 2006).
Vaksin DNA untuk penyakit SVCV asal Amerika Utara pada ikan koi menggunakan gen glikoprotein menghasilkan nilai RPS sebesar 50-88%. Penelitian ini merupakan penelitian pertama yang dilaporkan berhasil dilakukan pada ikan koi. Penelitian ini membuktikan bahwa vaksin DNA untuk penyakit SVCV (pSGnc) dapat menginduksi terbentuknya proteksi yang bersifat spesifik. Hasil penelitian tersebut sekaligus memvalidasi potensi pSGnc yang dapat digunanakan dalam pencegahan penyakit SVCV (Emmenegger & Kurath 2008).
Konstruksi Vaksin
17
maka tubuh memberikan reaksi dengan terbentuknya antibodi yang homolog. Skema tentang konstruksi singkat plasmid untuk vaksin dapat dilihat dalam ilustrasi yang diwakili oleh vaksin DNA dari kelompok Rhabdovirus berikut ini (Lorenzen & LaPatra 2005).
Gambar. 4. Skema gambaran partikel Rhabdovirus (a), vaksin plasmid (b) dan protein G virus
18
promoter yang bersifat house-keeping yaitu akan selalu aktif dalam siklus hidupnya. Selain bersifat house-keeping, β-actin juga mempunyai sifat ubiquitous (Hacket 1993), dimana promoter ini akan aktif dimana-mana dan constitutive (Volckaert et al. 1994) yang berarti bahwa promoter ini bisa aktif tanpa diberikan rangsangan dari luar seperti suhu dan hormon.
Promoter β-actin dari ikan medaka dilaporkan dapat aktif pada spesies yang sama, sekerabat atau berbeda jenis dengan asal promoter, seperti pada ikan rainbow trout (Yoshizaki 2001), ikan zebra (Alimuddin et al. 2005), ikan nila (Kobayashi et al. 2007), ikan lele (Ath-thar 2007), dan ikan mas (Purwanti 2007). Penggunaan promoter β-actin pada konstruksi vaksin DNA KHV memiliki peluang besar untuk mengaktivasi gen glikoprotein pada ikan mas.
Metode transfer vaksin DNA adalah hal yang penting ketika dikaitkan dengan aplikasi di lapangan (Leong et al. dalam www.nps.ars.usda.gov). Transfer vaksin pada mamalia menggunakan strategi injeksi intramuscular (IM) dengan gene-gun. Teknik ini sebenarnya efektif dalam menginduksi terbentuknya respon imun pada ikan, akan tetapi teknologi ini terlalu mahal apabila diaplikasikan pada budidaya ikan. Injeksi intramuscular (IM) sederhana plasmid DNA yang telah dimurnikan dalam buffer netral lebih efisien diterapkan di ikan daripada di hewan tipe yang lain (Lorenzen & LaPatra 2005). Meskipun aplikasi melalui injeksi IM merupakan metode yang dapat dipertimbangkan dalam vaksinasi, akan tetapi pengembangan aplikasi dengan metode yang lain perlu terus dikembangkan misalnya melalui perendaman atau melalui pencampuran dengan pakan (edible vaccine) dengan mempertimbangkan keamanan bagi lingkungan (Leong et al. dalam www.nps.ars.usda.gov.)
Kajian yang dilakukan terhadap distribusi dan ekspresi vaksin DNA terhadap lymphocystis disease virus (LCDV) pada ikan sebelah menunjukkan hasil bahwa
plasmid yang mengandung vaksin terdistribusi pada otot bekas penyuntikan, otot yang berseberangan dengan lokasi penyuntikan, usus, insang, limpa, ginjal depan, hati dan gonad setelah vaksinasi selama tujuh hari. Hasil tersebut diperoleh melalui kajian PCR maupun RT-PCR. Konstruksi vaksin tersebut membawa gen gfp (gene fluorescent protein) dimana fluoresensi dapat diamati pada otot bekas penyuntikan,
19
Terdistribusinya vaksin DNA pada beberapa organ yang lain dapat memproduksi antigen yang dapat menginduksi kekebalan spesifik ikan (Zheng et al. 2006).
Respons Imun pada Ikan
Ikan memiliki respon imun humoral maupun seluler sebagaimana vertebrata yang lain. Ikan dan mamalia memiliki kesamaan dan perbedaan imunitas (Tabel 3).
Tabel 3. Perbedaan antara imunitas ikan dan mamalia
Parameter Ikan Mamalia
Keadaan biotik Rentang suhu Lingkungan primer Metabolisme
-2 – 35 oC Air
Poikiloterm
36.5 – 37.5 oC Udara
Homoioterm Keragaman humoral
Ig isotope
Ig gene rearrangement
Keragaman non spesifik
IgM, IgD? (teleostei) IgM, IgX/IgR, IgW, NAR(C) (condrichthyes), IgM redoks Multiseluler (condrichthyes dan beberapa teleostei) C3 (teleostei)
IgM, IgA, IgD, IgE, IgG
Translocon
Tidak ada bentuk C3 Performa keseluruhan Afinitas antibodi Respon antibodi Respon memori Pematangan afinitas Suhu rendah Rendah Perlahan Lemah
Rendah atau tidak ada Ketergantungan tinggi, respon immunosuppressive (hanya pada ikan poikiloterm)
Tinggi Cepat Kuat Tinggi Ketergantungan rendah Organ Limfoid Jaringan hematopoietic Timus Kelenjar limfoid
Jaringan limfoid yang
berasosoasi dengan usus Pusat Germinal
Ginjal depan (teleostei), epigonal, organ Leydig, jaringan meningeal, orbital,
jaringan hematopoietic
subcranial (condrichthyes) Involusi tergantung spesies, dipengaruhi musim
Tidak ada
Tidak terorganisir, kumpulan limfoid
Tidak ada (pusat
melanomakrofag?), sel
dendrite kemungkinan ada
Sumsum tulang belakang
Involusi
Ada
Terorganisir
Ada
Sumber: Tort et al. (2003)
Respons Imun Non-Spesifik
20
harus berhadapan terlebih dahulu dengan barier pertahanan tubuh yang bersifat fisik dan kimiawi. Patogen harus menembus barikade lendir/mucus yang ada di bagian tubuh paling luar. Lendir ini memiliki kemampuan untuk menggumpalkan antigen secara kimiawi. Setelah itu patogen harus mampu menerobos kulit maupun melewati sisik terlebih dahulu untuk ikan yang bersisik. Setelah bagian ini lolos maka pathogen harus berhadapan dengan sistem pertahanan non-spesifik lainnya dalam tubuh (Tort et al. 2003).
Sistem pertahanan yang bersifat non-spesifik terdiri atas pertahanan seluler dan humoral. Pertahanan non spesifik seluler melibatkan makrofag, granulosit, non-specific cytotoxic cells (NCC).dan cell-line. Makrofag dan granulosit merupakan sel fagositik yang bersifat motil yaitu dapat bergerak ke seluruh bagian-bagian tubuh. Makrofag dan granulosit dapat ditemukan atau diisolasi dari darah, organ limfoid yaitu ginjal depan dan rongga peritoneal (peritoneal cavity) Granulosit terdiri atas neutrofil dan eosinofil, sedangkan basofil jarang ditemukan. Non-specific cytotoxic cells (NCC) pada ikan identik dengan natural killer (NK) cells pada mamalia. Sel ini bertugas untuk melisis sel kanker pada mamalia, sedangkan di ikan berfungsi untuk menghadapi parasit. Sel ini dapat ditemukan di darah, jaringan limfoid, usus dan ginjal depan. Cell line leukosit ini jarang ditemukan. Cell line ini berperanan dalam melakukan fagositosis. Pertahanan non spesifik humoral diperankan oleh lisozim, komplemen, interferon, protein C-reaktif, transferin dan lektin (Iwama & Nakanishi 1996).
Lisozim dapat ditemukan di lendir/mucus, serum dan telur. Zat ini berfungsi untuk membantu mendegradasi lapisan peptidoglikan pada dinding sel bakteri baik gram positif maupun gram negatif. Lisozim juga mendorong aktifitas fagositosis yaitu sebagai opsonin atau secara langsung mengaktifkan leukosit polimorfonuklear (neutrofil) dan makrofag. Neutrofil juga berisi sebagian besar myeloperoksidase yang terlibat dalam aktifitas bakterisidal (Mohanty et al. 2007). Lisozim merupakan salah satu respon alamiah (innate) yang dapat terinduksi dengan cepat (Tort et al. 2003).
21
mamalia berbeda dengan ikan. Mamalia hanya punya satu isoform molekul C3 yang dikode oleh satu loci, sementara ikan mengekspresikan beberapa isoform C3 aktif. Ikan trout (Onchoryncus mykiss) dan medaka (Oryzias latipes) mengekspresikan tiga isoform C3, ikan seabrem (Sparus aurata) dan mas (Cyprinus carpio) mengekspresikan lima isoform C3 serta ikan zebra (Danio rerio) memiliki tiga loci yang mengkode tiga isoform C3 (Tort et al. 2003).
Interferon adalah protein atau glikoprotein yang dapat menghambat replikasi virus. Pada mamalia terdapat tiga tipe interferon yaitu interferon α, β, dan Ә. Interferon α dan β terdapat pada ikan bertulang sejati dan tidak terdapat pada kelompok lainnya. Interferon Ә disekresikan oleh leukosit yang berasal dari ginjal depan ikan. Protein C-reaktif adalah protein pertama yang tampak pada plasma manusia dan hampir semua hewan (termasuk invertebrata dan moluska) ketika terjadi kerusakan jaringan, infeksi atau inflamasi. Protein ini berperanan dalam melakukan presipitasi polisakarida C (CPS=C-polysaccharides) yang terdapat pada dinding sel bakteri (Iwama & Nakanishi 1996). Protein C-reaktif berukuran 118 kDa dengan koefisien sediimentasi 6.5 S dan memperlihatkan mobilitas tipe β pada medan elektroforesis (Ingram 1980). Faktor ini akan meningkat jumlahnya pada fase akut suatu penyakit infeksi mikrobial dan akan mengikat fosforil-kolin dari glikopeptida dinding sel bakteri, cendawan dan parasit (Rijkers 1982).
Transferin adalah glikoprotein pengikat besi (Fe) yang berperanan dalam transport Fe mulai dari absorbsi, penyimpanan, dan pemanfaatannya pada semua vertebrata (Iwama & Nakanishi 1996). Berat molekul transferin tergantung spesies ikan. Berat molekul transferin pada ikan mas 58-70 kDa, sedang pada ikan dogfish 75-80 kDa. Transferin mempunyai aktifitas antimikrobial yang menghambat pemanfaatan metal oleh bakteri, mengangkut Fe dari situs penyerapannya di usus dan perombakan Hb ke berbagai lokasi penyimpanan (Ingram 1980).
22
meningkatkan aktifitas bakterisidal dan aktifitas makrofag dalam memfagositosis bakteri A.salmonicida (Ewart et al. 2001).
Respons Imun Spesifik
Sistem pertahanan spesifik dibedakan menjadi dua macam yaitu yang bersifat seluler dan humoral. Sistem pertahanan spesifik seluler diperankan oleh kekebalan berperantara sel (cell mediated immunity/CMI) dalam hal ini limfosit Tc, sedangkan pertahanan spesifik humoral diperankan oleh antibodi (Tort et al. 2003).
Kekebalan berperantara sel pada mamalia diperankan oleh sel Tcytotoxic (CTL=cytotoxic lymphocyte). Sel T ini memiliki molekul CD8 pada permukaannya untuk berinteraksi dengan molekul major histocompatibility complex kelas I (MHC I) pada permukaan sel APC (antigen presenting cell) yang menyajikan antigen (Hirono 2005). Sel Tc ini akan mencari sel-sel yang mengalami kelainan fisiologis untuk dihancurkan. Tujuan pembungihangusan ini adalah untuk menghindari penyebaran penyakit (Kuby 1997).
Roitt (2003) memaparkan bahwa kekebalan berperantara sel (seluler) melibatkan limfosit Tc yang dihasilkan di kelenjar timus yang bekerjasama dengan NK (natural killer= non specific cytitoxic cells/NCC) dalam menghancurkan sel yang mengalami kelainan. Ruang lingkup sel NK ini terbatas sehingga perlu bantuan antibodi untuk mengahancurkan sel yang terinfeksi parasit intraseluler yaitu virus. Antibodi menyelimuti sel yang terinfeksi virus sementara NK yang memiliki reseptor khusus terhadap antibodi berikatan dengan antibodi. Antibodi akan membawa sel NK mendekat ke sel sasaran dengan membentuk suatu jembatan. Sel NK yang diaktifkan oleh kompleks antigen-antibodi mampu membunuh sel terinfeksi melalui mekanisme ekstraseluler. Pembunuhan sel ini bersifat non-spesifik. Pembunuhan sel terinfeksi virus yang bersifat spesifik dilakukan oleh sel Tc. Sel ini berhubungan dengan sel target dengan bantuan molekul major histocompatibility complex (MHC) kelas I. Melalui pengenalan terhadap antigen permukaan ini maka sel-sel sitotoksik datang untuk membuat kontak yang lebih intim dengan sel target. Sel sitotoksik juga melepaskan Ә-interferon yang membantu memperkecil peluang penyebaran virus ke sel-sel yang lainnya yang berdekatan. Sel sitotoksik melakukan pembumihangusan sel target sehingga penyebaran virus dapat dihentikan.
23
terdeteksi oleh sel Thelper dan sel T ini memberikan sinyal melalui sitokin/interleukin kepada sel B untuk melakukan prolifareasi. Sel B yang telah mengalami proliferasi menghasilkan antibodi yang sesuai dengan antibodi yang menginduksinya (Roitt 2003).
Mamalia memiliki beberapa kelas antibodi/immunoglobulin (Ig yaitu IgA, IgD, IgE IgG dan IgM. Ikan bertulang sejati memiliki kelas Ig yang terbatas yaitu IgM. IgM ini berukuran 800 kD (Tort et al. 2003), dengan koefisien sedimentasi 16 S dan berbentuk monomer, tetramer dan pentamer (Walczak 1985). Semua kelas Ig mengandung karbohidrat yang terikat pada atom C dari rantai H. Dalam proses pengikatan karbohidrat seperti manosa, galaktosa, fukosa pada situs asparagin, serin atau treonin diperlukan enzim N-asetil-glukosamin-asparagin transglikosilase. Ikatan karbohidrat ini diperlukan untuk meningkatkan kelarutan Ig, mencegah degradasi katabolik dan mempermudah sekresi antibodi dari sel pembentuknya. Ig M merupakan makroglobulin dimana kestabilan struktur molekulnya dilakukan oleh rantai J. Klasifikasi Ig tersebut didasarkan atas sifat fisiko-kimia, kandungan karbohidrat dan komposisi asam amino molekul Ig (Rosenshein et al. 1985).
BAHAN DAN METODE
Tahap I: Pembuatan Konstruksi Vaksin
DNA KHV dan Plasmid
DNA KHV yang digunakan sebagai sumber isolasi gen adalah DNA yang berasal dari virus tipe liar. DNA ini diperoleh dari Balai Riset Perikanan Budidaya Air Tawar (BRPBAT Bogor). Sampel DNA dari virus tipe liar tersebut berkode 2, 3, 1A, 2A, 3A, B1, B2, B3, U2B, U2C.
Plasmid yang digunakan adalah pGEMT-Easy (Promega) yang memiliki gen lacZ dan penanda resisten ampisilin, sedangkan plasmid yang berfungsi untuk mengekspresikan gen adalah pActD6 (Alimuddin et al. 2005) yang
mengandung promoter β-aktin (Act) dari ikan medaka (Jepang) yang menyisip pada situs EcoRI dan poly-A (terminator) yang yang berasal dari bovine growth hormone dan menyisip pada situs XhoI. Plasmid ini masih mengandung gen
masu salmon Δ6-desaturase- like di antara Act dan BGH-polyA.
Disain Primer
Disain primer dilakukan terhadap ORF 25 sekuens lengkap koi herpesvirus (Aoki et al. 2007) yang dimulai pada basa ke-45587 sampai 47393. Primer forward disusun dari start codon ATG sampai basa ke-19, sedangkan primer reverse disusun dari stop codon TAA dan ditentukan basa komplemennya sampai sebanyak 22 basa. Untuk keperluan konstruksi sehingga hasil amplifikasi sesuai dengan situs yang ada di vektor ekspresi maka ke dalam primer forward disisipkan situs SalI.
Isolasi Gen Penyandi Glicoprotein
Isolasi DNA dilakukan dengan cara mengamplifikasi DNA KHV dengan menggunakan primer yang didisain khusus (primer spesifik) yaitu primer forward
F: TTGTCGACATGACGGGTTGTGGGGTTTG dan primer reverse R:
TTAGGGCCTCCGGGAAACCTGG. Kondisi reaksi polimerisasi adalah prakondisi pada suhu 95oC selama tujuh menit, denaturasi pada suhu 95oC selama 30 detik, annealing pada suhu 64oC selama 30 detik, ekstensi pada suhu 72 oC selama dua menit dan finally pada suhu 72oC selama tujuh menit.
25
menggunakan skalpel dan hasil potongan dimasukkan ke dalam tabung mikro 1.5 ml. Larutan binding buffer sebanyak 400 ul ditambahkan ke dalam tabung mikro yang berisi potongan DNA dalam agar, diinkubasi pada suhu 55oC selama 10 menit dan selanjutnya dikocok dengan kuat sampai agar terlarut.
Isi tabung mikro dituang ke dalam EZ-10 column dan dibiarkan selama dua menit, selanjutnya disentrifugasi pada 10000 rpm selama dua menit dan supernatan dibuang. Cairan pencuci (wash solution) ditambahkan ke dalam column sebanyak 500 ul, disentrifugasi pada 10000 rpm selama satu menit.
Cairan dalam tabung dibuang. Cairan pencuci (wash solution) ditambahkan lagi ke dalam column sebanyak 500 ul dan disentrifugasi pada 10000 rpm selama satu menit. Column ditempatkan pada tabung Eppendorf, ditambahkan sebanyak 30-50 ul elution buffer pada bagian tengah column dan diinkubasi pada suhu ruang selama dua menit. Column dan tabung eppendorf tersebut disentrifugasi pada 10000 rpm selama dua menit untuk mengelusi DNA.
Ligasi Fragmen Glikoprotein dan pGEMT-Easy
Gen penyandi glicoprotein pada KHV diligasikan dengan plasmid pGEMT-Easy (Promega) dengan cara mencampurkan antara 1 ul pGEMT-Easy dengan 4 ul gen glicoprotein KHV. Enzim ligase yang digunakan untuk proses ligasi tersebut adalah enzim T4 DNA ligase sebanyak 1 ul dan buffer T4 DNA ligase sebanyak 7 ul. Pencampuran dilakukan pada suhu ruang, dibiarkan selama 2 jam, setelah itu disimpan di refrigerator.
Transformasi pada E.coli Kompeten
Setelah plasmid pGEMT-Easy tersisipi gen glicoprotein KHV maka plasmid ini siap diintroduksi ke dalam sel kompeten yang berasal dari bakteri E.coli DH5α. Plasmid ini dispindown sebentar, sebanyak 6 ul plasmid
26
media Luria Bertani yang diberi ampisilin ( 100 ug/ml atau 20 ul/cawan), IPTG (20 mg/ml) sebanyak 100 ul/cawan dan X-gal (40 mg/ml) sebanyak 100 ul/cawan. Cawan diinkubasi overnight selama 16-24 jam pada 37 oC.
Verifikasi Hasil Transformasi
Verifikasi terhadap hasil transformasi dilakukan dengan cara mengamati adanya koloni putih-biru. Plasmid pGEMT-Easy yang digunakan memang mamiliki penanda khusus koloni putih-biru ini. Koloni putih adalah koloni yang tersisipi plasmid yang mengandung sisipan gen glikoprotein. Plasmid pGEMT-Easy ini memiliki gen lacZ yang menyandi β-galactosidasse yang akan mengubah molekul X-gal dari tidak berwarna menjadi melekul yang berwarna biru. Ekspresi gen lacZ diinduksi oleh IPTG (isopropil thiogalactosida). Bila gen lacZ tersisipi oleh molekul DNA lain, maka lacZ ini tidak dapat diekspresikan, sehingga sel yang mengandung plasmid demikian tidak dapat mengubah X-gal menjadi b