Kajian Karakter Ketahanan terhadap Cekaman Kekeringan pada Beberapa Genotipe Bibit Kelapa Sawit (Elaeis guineensis Jacq.)
Teks penuh
Gambar

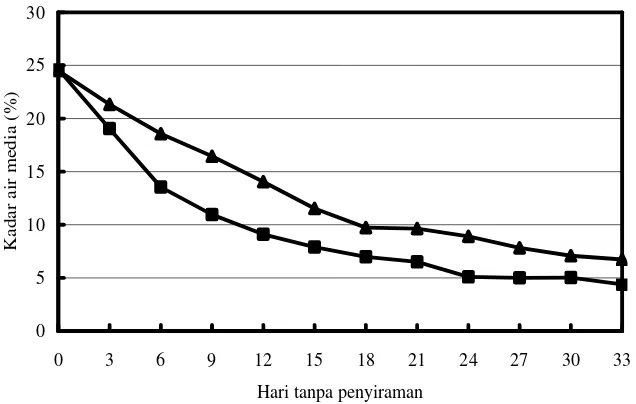
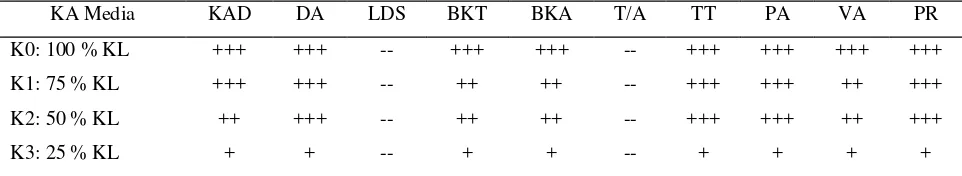
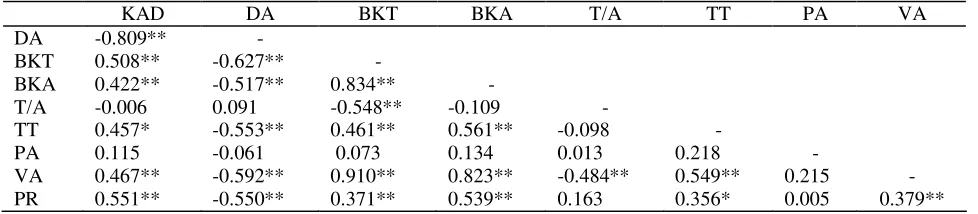
Dokumen terkait
Tesis Pondok pesantren dan perubahan ..... ADLN -
Antara yang jelas dapat diperhatikan adalah amalan-amalan berikut yang kini mula menjadi norma dalam kalangan masyarakat Islam di Malaysia iaitu, amalan menyalakan api
Pada proses validasi yang dilakukan oleh peneliti, baik validasi kepada ahli materi, ahli media dan ahli desain, peneliti mendapatkan penilain dari ahli materi,
model direct instruction berbantuan animasi energy2d , siswa dapat mengganti konsepsi awal yang salah menjadi konsepsi yang benar dengan bantuan animasi
Penawaran publik sekuritas yang dibuat di Amerika Serikat akan dilakukan melalui prospektus yang diperoleh dari Perusahaan dan berisi keterangan rinci mengenai
Studi Kesesuaian Jenis untuk Perencanaan Rehabilitasi Ekosistem Mangrove di Desa Wawatu Kecamatan Moramo Utara Kabupaten Konawe Selatan.. Analisis Kesesuaian Lahan Wilayah
Berdasarkan hal tersebut pada penelitian ini dirancang sebuah sistem informasi yang diberi nama GLoSha ( Grouping Location Sharing ) yang dapat membantu
Desain Sistem Prototype Akuarium yang dibuat pada penelitian ini dirancang dengan menggunakan sensor pH untuk mengetahui kualitas air serta sensor hcsr yang mengukur
