KANDUNGAN ZAT PENGATUR TUMBUH DAUN DAN POLA
INFLORESEN BUNGA PADA JARAK PAGAR
(
Jatropha curcas
L.) ANDROMONOECIOUS
SITI KOMARIYAH
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
SITI KOMARIYAH. Kandungan Zat Pengatur Tumbuh Daun dan Pola Infloresen Bunga pada Jarak Pagar (Jatropha curcas L.) Andromonoecious. Dibimbing oleh TRIADIATI dan WULAN
TRI WAHYUNI.
Jarak pagar merupakan tanaman berumah satu (monoecious), dengan bunga jantan dan betina yang terpisah. Akan tetapi, pada Jarak pagar telah diketahui terdapat tanaman andromonoecious (yang memiliki bunga hermaprodit dan bunga jantan). Salah satu faktor internal yang mengendalikan fase reproduktif pada tanaman ialah Zat Pengatur Tumbuh (ZPT). Oleh karena itu, penambahan ZPT seringkali dilakukan untuk mengoptimalkan pertumbuhan vegetatif dan reproduktif tanaman. Penelitian ini bertujuan menganalisis kandungan auksin, giberelin, dan sitokinin daun serta pola infloresen bunga pada jarak pagar andromonocieous pada berbagai perlakuan ZPT. Zat pengatur tumbuh IAA, GA3, dan kinetin disemprotkan pada daun setiap dua minggu sekali sampai dua bulan sebelum tanaman berbunga. Analisis ZPT daun menggunakan metode ekstraksi dan spektrofotometri. Kandungan auksin dan giberelin pada tanaman jarak pagar monoecious lebih tinggi pada fase sebelum berbunga dibandingkan setelah proses pembungaan, sedangkan kandungan sitokinin pada tanaman jarak pagar monoecious sebelum berbunga lebih rendah dibandingkan setelah berbunga. Pada tanaman jarak pagar andromonoecious tanpa perlakuan ZPT kandungan auksin dan sitokinin mengalami kenaikan setelah proses pembungaan, tetapi tidak demikian dengan giberelin yang mengalami penurunan. Penyemprotan auksin dan giberelin 100 ppm terhadap tanaman andromonoecious mampu meningkatkan kandungan ZPT endogen. Penyemprotan kinetin pada tanaman Jarak pagar andromonoecious dapat meningkatkan jumlah percabangan infloresen bunga.
Kata kunci : monoecious, andromonoecious, zat pengatur tumbuh, infloresen
ABSTRACT
SITI KOMARIYAH. The Leaf Plant Growth Regulators Content and Flowers Inflorescense Pattern on Andromonoecious Jatropha curcas L. Supervised by TRIADIATI and WULAN TRI WAHYUNI.
Jatropha curcas is a monoecious plant with separated male and female flowers. However, there is also andromonoecious Jatropha plant that produce hermaphrodite and male flowers. One of the internal factors controlling reproductive phase is Plant Growth Regulators (PGRs). Addition of PGRs could optimize vegetative and reproductive phase. The aims of this study were to analysis the content of leaf auxins, gibberellins, and cytokinins, as well as flower inflorescense pattern on andromonoecious Jatropha curcas under PGR treatments. PGRs IAA, GA3, and kinetin was sprayed to leaves every two weeks until two months before flowering. PGR analysis was performed using extraction and spectrophotometric methods. The content of auxins and gibberellins in monoecious Jatropha werehigher when was analyzed before flowering than that of after flowering, whereas the content of sitokinin on monoecious plant was lower before flowering than that of after flowering. Auxin and sitokinin content in non treated andromonoecious increased after flowering. whereas gibberellins content decreased. Treatment of 100 ppm auxin and giberellin to the andromonoecious Jatropha could increase the content of PGR endogenous. Kinetin increased number of flower inflorescense on andromonoecious Jatropha.
Judul Skripsi : Kandungan Zat Pengatur Tumbuh Daun dan Pola Infloresen Bunga
pada Jarak pagar (
Jatropha curcas
L.) Andromonoecious
Nama
: Siti Komariyah
NRP
: G34070070
Menyetujui
Mengetahui
Kepala Departemen Biologi
Fakultas Matematika dan Ilmu Pengetahuan Alam
Tanggal lulus :
Pembimbing I,
Dr. Triadiati, M.Si
NIP 19600224 198603 2 001
Pembimbing II
,
Wulan Tri Wahyuni, M.Si
Dr. Ir. Ence Darmo Jaya Supena, M.Si
KANDUNGAN ZAT PENGATUR TUMBUH DAUN DAN POLA
INFLORESEN BUNGA PADA JARAK PAGAR (
Jatropha curcas
L.)
ANDROMONOECIOUS
SITI KOMARIYAH
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
PRAKATA
Puji syukur kehadirat Allah SWT karena atas rahmat dan hidayah-Nya penulis dapat menyelesaikan karya ilmiah yang berjudul “Kandungan Zat Pengatur Tumbuh Daun dan Pola Infloresen Bunga pada Jarak Pagar (Jatropha curcas L.) Andromonoecious.” Penelitian ini dilaksanakan mulai bulan Januari sampai Agustus 2011.
Penulis ucapkan terima kasih kepada Ibu Dr. Triadiati, M.Si dan Ibu Wulan Tri Wahyuni, M.Si selaku pembimbing atas pengarahan, teladan, kesabaran, waktu, serta nasehat yang diberikan kepada penulis salama penelitian hingga penulisan karya ilmiah ini. Ucapan terima kasih juga penulis sampaikan kepada bapak Dr.Ir Miftahuddin, M.Si selaku dosen penguji yang telah banyak memberikan saran dan masukan untuk perbaikan penulisan karya ilmiah ini. Penghargaan penulis sampaikan kepada Ibu Nung dan Pak Eman selaku teknisi di Laboratorium Kimia Analitik, Pak Eko selaku teknisi di Laboratorium Bersama Kimia, serta Pak Kus dan Ibu Gleny selaku teknisi di Laboratorium Fisiologi Tumbuhan, yang telah membantu selama pengumpulan data. Ungkapan terima kasih juga disampaikan kepada bapak, ibu, kakak, dan adik serta seluruh keluarga atas segala doa dan kasih sayangnya. Penulis juga mengucapkan terima kasih kepada mbak tri sundari, teman-teman seperjuanganku iip, danis, yuni, age, niswa, fina, dan ifit yang telah banyak memberikan dukungan, teman-teman wisma bintang dan teman-teman Biologi 44 yang telah memberikan dukungan dan semangat pada penulis serta semua pihak yang telah membantu penyusunan karya ilmiah ini.
Semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2012
RIWAYAT HIDUP
Penulis dilahirkan di Lampung pada tanggal 12 Mei 1989 dari pasangan bapak Sardi dan ibu Sumiyati. Penulis merupakan anak kedua dari tiga bersaudara.
Tahun 2007 penulis lulus dari SMAN 1Sukoharjo dan pada tahun yang sama penulis lulus seleksi masuk Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk IPB (USMI). Penulis memilih mayor Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam.
Selama mengikuti perkuliahan, penulis aktif di Unit Kegiatan Mahasiswa KSR Unit 1 IPB (2007-2008), KEMALA (Keluarga Mahasiswa Lampung, 2007), SERUM-G (2008-2010), dan Himpunan Mahasiswa Biologi (2008). Penulis mendapatkan beasiswa Program peningkatan Akademik (PPA) 2007-2008 dan Bantuan Belajar Mahasiswa (BBM) 2009-2011. Pada alih tahun 2008- 2009, penulis mengikuti kegiatan magang di Lembaga Ilmu Pengetahuan Indonesia (LIPI) dengan topik “Teknik Pembuatan Spesimen Herbarium”. Pada alih tahun 2009-2010, penulis mengikuti kegiatan magang di Sringanis mengenai tanam-tanaman obat. Pada tahun 2010, penulis melaksanakan Praktek Lapangan di Unit Peternakan Darul Fallah Ciampea-Bogor dengan judul “Proses Pemeliharaan Kambing Perah di Unit Peternakan Darul Fallah-Bogor”. Penulis juga pernah menjadi asisten mata kuliah Fisiologi tumbuhan (semester genap 2010-2011), Biologi Dasar (semester ganjil dan genap 2010/2011), dan Pendidikan Agama Islam (2010-2011).
DAFTAR ISI
Halaman
DAFTAR LAMPIRAN ... vii
PENDAHULUAN
Latar Belakang ... ...1
Tujuan ... 1
BAHAN DAN METODE
Tempat dan Waktu ... 1
Bahan dan Alat ... 1
Metode Penelitian... 1
HASIL ... 2
PEMBAHASAN ... 3
SIMPULAN ... 5
DAFTAR PUSTAKA ... 5
DAFTAR LAMPIRAN
Halaman1 Spektrum panjang
gelombang Giberelin………
...
8
2 Spektrum
panjang gelombang kinetin………...
...
8
3 Spektrum panjang gelombang Auksin
………..
...
9
4 Hasil pengukuran kurva standar giberelin...
5 Hasil pengukuran kurva standar auksin...
6 Hasil pengukuran kurva standar kinetin...
10
10
1
PENDAHULUAN
Latar Belakang
Pada umumnya, jarak pagar (Jatropha curcas) merupakan tanaman berumah satu (monoecious), dengan bunga jantan dan betina terpisah tetapi masih dalam satu tanaman yang sama (Hartati 2009). Saat ini, diketahui terdapat tanaman jarak pagar yang mempunyai bunga hermaprodit dari tanaman monoecious, namun jumlahnya sangat terbatas. Tanaman jarak pagar berbunga hermaprodit, termasuk tipe tanaman andromonoecious (hanya mempunyai bunga hermaprodit dan bunga jantan). Pada satu karangan bunga, jumlah bunga jantan lebih banyak dibandingkan bunga hermaprodit. Bunga hermaprodit memiliki ukuran yang lebih besar dibandingkan bunga jantan (Delita et al. 2008). Bunga hermaprodit dapat memberikan peluang terjadinya penyerbukan sendiri. Tanaman yang menyerbuk sendiri akan menghasilkan biji yang bersifat homogen, sedangkan tanaman menyerbuk silang umumnya menghasilkan biji yang bersifat heterogen.
Zat pengatur tumbuh ialah senyawa organik yang disintesis di salah satu bagian tumbuhan dan dipindahkan ke bagian lain, dan pada konsentrasi yang sangat rendah mampu menimbulkan respon fisiologis (Salisbury & Ross 1995). Tanaman secara alami sudah mengandung hormon pertumbuhan yang disebut hormon endogen. Namun, hormon ini kurang optimum mempengaruhi proses pertumbuhan vegetatif dan reproduktif tanaman. Penambahan Zat Pengatur Tumbuh (ZPT) sering kali dilakukan untuk mengoptimalkan pertumbuhan vegetatif dan reproduktif tanaman, misalnya giberelin (GA3) yang mampu mempercepat pertumbuhan dan pembungaan (Abidin 1985). Giberelin mampu mempercepat pembungaan tanaman melalui pengaktifan gen meristem bunga dengan menghasilkan protein yang akan menginduksi ekspresi gen-gen pembentuk organ bunga (seperti korolla, kalix, stamen, dan pistillum) (Arika et al. 2009). Auksin merupakan salah satu hormon tanaman yang dapat meregulasi banyak proses fisiologi, seperti pertumbuhan, pembelahan dan diferensiasi sel serta sintesa protein (Darnell
et al. 1986). Peningkatan konsentrasi sitokinin pada daun merupakan salah satu fenomena hormonal yang penting bagi
induksi bunga. Ryugo (1990) menyatakan bahwa aplikasi sitokinin pada permukaan petiol yang dipotong dapat meningkatkan persentase tunas bunga. Berdasarkan hal tersebut diduga ZPT memiliki peranan yang penting dalam memacu munculnya bunga khususnya hermaprodit pada tanaman jarak pagar.
Bunga jarak pagar tersusun secara majemuk (infloresen). Menurut Heller (1996) infloresen pada jarak pagar terbentuk pada cabang terminal. Infloresen bunga pada jarak pagar bertipe dichasial
cyme. Tipe infloresen bunga tersebut ialah tipe bunga yang memiliki ciri antara lain pada ujung tangkai utama terdapat sebuah kuncup bunga, bunga pada ujung tangkai utama akan mekar lebih dahulu, tangkai utama biasanya lebih pendek dari anak tangkai, tangkai utama hanya membentuk anak tangkai dengan jumlah sedikit dan percabangan pada anak tangkai tidak berbeda dari tangkai utama (Raju & Ezradanam 2002).
Tujuan
Penelitian ini bertujuan menganalisis kandungan auksin, giberelin, dan kinetin dari daun serta pembentukan pola infloresen bunga pada tanaman jarak pagar (J. curcas) andromonoecious pada berbagai perlakuan zat pengatur tumbuh.
BAHAN DAN METODE
Tempat dan Waktu
Penelitian ini dilaksanakan pada bulan Januari sampai Agustus 2011 di lahan pertanian Cibeureum, lahan pertanian Cikabayan, Laboratorium Kimia Analitik dan Laboratorium Bersama Kimia Departemen Kimia dan Laboratorium Fisiologi Tumbuhan Departemen Biologi FMIPA, Institut Pertanian Bogor.
Bahan dan Alat
Bahan yang digunakan ialah daun tanaman jarak pagar andromonoecious dan daun tanaman jarak pagar monoecious. Zat pengatur tumbuh yang digunakan antara lain auksin (IAA), giberelin (GA3), dan kinetin (K), bahan kimia yang digunakan ialah metanol, NH4OH, kloroform, etilasetat, dan isopropanol.
2
lain yang biasa digunakan di dalam laboratorium.
Metode Penelitian
Perlakuan. Tanaman jarak pagar andromonoecious disemprot pada permukaan bawah daun pada pagi hari dengan menggunakan GA3 100 ppm, IAA 100 ppm, kinetin 100 ppm, GA3:IAA (50:50 ppm), GA3:K (50:50 ppm), IAA:K (50:50 ppm) masing-masing sebanyak 300 ml. Perlakuan diberikan 2 minggu sekali pada tanaman saat jumlah daun minimal 5 helai sampai dua bulan sebelum berbunga. Analisis kandungan ZPT menggunakan tanaman dengan perlakuan ZPT tunggal, sedangkan pengamatan pola infloresen bunga untuk semua perlakuan ZPT.
Pengambilan Sampel. Sampel daun diambil sebelum munculnya bunga dan saat bunga mulai muncul. Sampel yang diambil adalah daun dengan lebar penuh yang memiliki warna hijau tua dan rata-rata terletak pada duduk daun ketiga dari pucuk.
Analisis ZPT pada daun. Analisis ZPT pada daun mengikuti metode yang digunakan Unyayar et al. (1996). Sampel daun yang diambil secara komposit dari 3 ulangan seberat ± 5 gr diekstrak dengan cara digerus, kemudian dilarutkan dengan metanol, kloroform, dan 2N ammonium hidroksida (12:5:3 v/v/v) sebanyak 100 ml. Ekstrak yang diperoleh kemudian disaring. Air destilata sebanyak 22,4 ml ditambahkan ke dalam ekstrak yang diperoleh, selanjutnya ekstrak dimasukkan ke dalam corong pemisah dan didiamkan selama 24 jam. Fase cair diambil sedangkan fase kloroform dibuang. Selanjutnya, pH pada fase air diatur menjadi 2,5 dengan menambahkan HCL 5N, kemudian diekstraksi sebanyak 3 kali dengan menggunakan 15 ml etilasetat. Ekstrak didiamkan beberapa menit agar fase air terpisah dari fase etilasetat. Fase etilasetat diambil dan fase air dilanjutkan untuk perlakuan berikutnya. Fase etilasetat yang diperoleh diharapkan mengandung IAA dan GA3. Fase air berikutnya diatur pH nya menjadi 7 dengan menambahkan 4N NaOH kemudian diekstraksi kembali sebanyak 3 kali menggunakan 15 ml etilasetat. Perlakuan ini akan menghasilkan kinetin bebas. Ekstrak IAA, GA3, dan kinetin yang diperoleh selanjutnya dievaporasi. Ekstrak kering yang diperoleh kemudian ditimbang ± 0,1-0,5 mg dan dilarutkan menggunakan 10 ml metanol kemudian diukur
menggunakan spektrofotometri UV-VIS pada panjang gelombang 253 untuk GA3, 279,6 untuk IAA, dan 267,8 untuk kinetin. Panjang gelombang tersebut diperoleh dari hasil scanning ZPT standar (Lampiran 1-3). Konsentrasi ZPT pada sampel yang diperoleh dihitung dengan persamaan kurva standar (Lampiran 4-6).
HASIL
Hasil analisis ZPT pada tanaman jarak pagar monoecious dan kontrol andromonoecious menunjukkan bahwa kandungan giberelin mengalami penurunan dari fase sebelum berbunga ke fase pembungaan sebesar 4,47 mg/g ekstrak dan 0,22 mg/g ekstrak, sedangkan pada tanaman jarak pagar andromonoecious dengan penyemprotan giberelin 100 ppm mengalami peningkatan kandungan giberelin sebesar 2,42 mg/g ekstrak (Tabel 1).
Tabel 1 Kandungan zat pengatur tumbuh endogen pada tanaman jarak pagar andromonoecious dengan perlakuan berbagai ZPT
Sampel Kandungan ZPT daun
mg/gr ekstrak
GA3 K IAA
Monoecious
Sebelum berbunga 5,52 0,001 5,47
Setelah berbunga 0,75 0,37 0,94
Kontrol Andromonoecious
Sebelum berbunga 0,56 0,04 0,24
Setelah berbunga 0,34 0,06 0,52
Andromonoecious + IAA 100 ppm
Sebelum berbunga 0,08
Setelah berbunga 1,32
Andromonoecious + GA3 100 ppm
Sebelum berbunga 0,06
Setelah berbunga 2,48
Andromonoecious + Kinetin 100 ppm
Sebelum berbunga 0,04
Setelah berbunga -
Keterangan : GA3: giberelin, IAA: auksin, K: kinetin
3
karena sampai batas waktu yang ditentukan tanaman belum berbunga.
Kandungan auksin pada tanaman jarak pagar monoecious mengalami penurunan 4,53 mg/g ekstrak dari fase sebelum berbunga ke fase pembungaan. Pada tanaman jarak pagar kontrol andromonoecious dan andromonoecious dengan penyemprotan IAA 100 ppm keduanya mengalami peningkatan kandungan auksin dari fase sebelum berbunga ke fase setelah berbunga. Kandungan auksin pada tanaman jarak pagar kontrol andromonoecious meningkat 0,28
mg/g ekstrak, sedangkan pada tanaman jarak pagar andromonoecious dengan penyemprotan IAA 100 ppm meningkat 1,24 mg/g ekstrak.
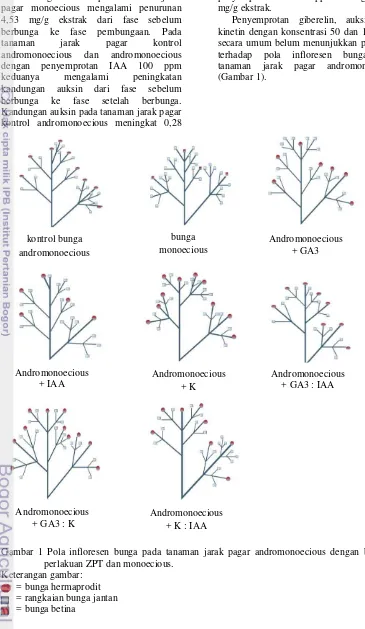
Penyemprotan giberelin, auksin, dan kinetin dengan konsentrasi 50 dan 100 ppm secara umum belum menunjukkan pengaruh terhadap pola infloresen bunga pada tanaman jarak pagar andromonoecious (Gambar 1).
Gambar 1 Pola infloresen bunga pada tanaman jarak pagar andromonoecious dengan berbagai perlakuan ZPT dan monoecious.
Keterangan gambar: = bunga hermaprodit = rangkaian bunga jantan = bunga betina
kontrol bunga andromonoecious
Andromonoecious + GA3 : IAA
Andromonoecious + GA3 : K
Andromonoecious + K : IAA Andromonoecious
+ K bunga monoecious
Andromonoecious + GA3
[image:10.595.112.477.117.746.2]4
PEMBAHASAN
Jarak pagar merupakan anggota famili Euphorbiaceae dengan jumlah bunga jantan lebih banyak daripada bunga betina (monoecious). Perbandingan bunga jantan dan bunga betina pada jarak pagar monoecious sebesar 29:1 (Raju & Ezradanam 2002).
Saat dimulainya proses pembungaan, terjadi peralihan dari proses produksi struktur daun yang relatif sederhana menjadi struktur pembungaan yang lebih kompleks. Saat pucuk terinduksi untuk berbunga, meristem berhenti membentuk daun dan mulai menghasilkan organ bunga. Pada saat itu terjadi perubahan-perubahan metabolisme pada pucuk yang disertai kenaikan laju pembelahan sel. Keberhasilan proses pembungaan dimulai setelah terjadi induksi bunga, diikuti proses diferensiasi, pendewasaan organ-organ bunga, antesis, dan polinasi (Bernier et al. 1985).
Salah satu peran hormon tanaman adalah mempengaruhi inisiasi pembungaan dengan mengatur keseimbangan antara karbohidrat dan nitrogen (rasio C/N). Kelebihan suplai zat makanan (karbohidrat) dalam tanaman akan merangsang terjadinya pembungaan. Arika et al. (2009) menyatakan bahwa giberelin mampu meningkatkan rasio C/N. Semakin tinggi rasio C/N, tanaman akan mengalami peralihan dari masa vegetatif ke reproduktif. Hal tersebut menyebabkan waktu inisiasi bunga lebih cepat.
Kandungan giberelin dan auksin tanaman jarak pagar monoecious pada penelitian ini lebih rendah pada fase sebelum berbunga dibandingkan fase setelah berbunga, sedangkan kandungan sitokininnya meningkat. Hal ini sesuai dengan pernyataan Rai et al. (2006), bahwa kandungan giberelin pada pucuk yang berbunga tetap rendah setelah stadium induksi terlewati, sedangkan kandungan giberelin pada pucuk yang tidak berbunga relatif tinggi. Menurut Tumbull et al. (1996) bahwa kandungan giberelin yang tinggi akan memacu pembelahan dan pemanjangan sel di apeks pucuk, terutama di bagian sel meristematik, sehingga memacu pertumbuhan vegetatif dan menghambat pembungaan.
Pada tanaman jarak pagar kontrol andromonoecious, kandungan giberelin menurun setelah melewati proses pembungaan, sedangkan kandungan auksin
dan kinetin meningkat setelah melewati proses pembungaan. Tanaman jarak pagar monoecious dan kontrol andromonoecious menunjukkan bahwa kandungan kinetin yang dihasilkan sebelum berbunga lebih rendah dibandingkan setelah berbunga dengan besar peningkatan yang berbeda. Kinetin merupakan ZPT yang mendorong pembelahan (sitokinesis), umumnya banyak terdapat pada daerah ujung akar dan daerah meristem yang mengalami pembelahan sel cepat, serta pada daerah yang sedang berkembang (Prawitasari 2002).
Produksi bunga berkorelasi tinggi dengan aktivitas kinetin pada meristem apikal. Pucuk yang terinduksi untuk menghasilkan bunga memiliki kandungan sitokinin yang lebih tinggi dibandingkan pucuk yang tidak menghasilkan bunga (Kinet et al. 1985). Berdasarkan penelitian yang dilakukan oleh Prawitasari et al.
(2002), bahwa kandungan sitokinin terus berfluktuasi sesuai perkembangan fase pembungaan. Kandungan sitokinin yang dihasilkan tanaman lengkeng saat H-30 hari sebelum bunga mekar cukup rendah dan terus mengalami peningkatan sampai H-15 hari sebelum bunga mekar. Fluktuasi kandungan sitokinin sangat tajam pada fase perkembangan organ bunga, karena pada saat itu, terjadi perkembangan primordia-primordia calon organ bunga yang menandakan aktivitas daerah perkembangan sangat tinggi.
Kandungan sitokinin mengalami penurunan sebelum bunga mekar. Hal ini disebabkan adanya aktivitas daerah meristematik yang mengalami pembelahan sel sudah menurun dan ditandai dengan membukanya petal secara sempurna. Beberapa hari setelah bunga mekar sempurna terjadi peningkatan sitokinin kembali, yang disebabkan adanya perkembangan organ calon buah yang berasosiasi dengan kebutuhan sitokinin bagi pembelahan sel-sel baru di daerah perkembangan hingga buah mencapai ukuran maksimum. Hal ini menandakan bahwa sitokinin berperan dalam induksi pembungaan dan diferensiasi sel.
5
melalui peningkatan laju sintesis protein. Beberapa di antara protein ini dapat berperan sebagai enzim yang dibutuhkan untuk terjadinya mitosis.
Penyemprotan kinetin 100 ppm ternyata menghambat pembungaan dan munculnya bunga hermaprodit. Hal ini menguatkan hasil penelitian Palupi et al. (2009) yang menyatakan bahwa perlakuan BAP 35 ppm pada tanaman jarak pagar mampu menghasilkan 24 bunga hermaprodit dari 18 infloresen yang diamati, sedangkan perlakuan BAP 50 ppm tidak menghasilkan bunga hermaprodit. BAP dan kinetin merupakan jenis sitokinin sintetik.
Penyemprotan giberelin dan auksin 100 ppm pada tanaman andromonoecious mampu meningkatkan kandungan ZPT endogen. Menurut Arika et al. (2009) bahwa pemberian giberelin pada tanaman akan meningkatkan kandungan auksin melalui pembentukan enzim proteolitik yang akan membebaskan senyawa triptophan sebagai prekursor auksin. Menurut Kaminek et al.
(1997) bahwa penambahan kinetin mampu meningkatkan kadar kinetin endogen baik melalui pengambilan ataupun peningkatan biosintesis kinetin. Peningkatan jumlah kinetin endogen ternyata justru menghambat perkembangan reproduktif tanaman. Salisbury & Ross (1995) menyatakan bahwa pemberian ZPT secara eksogen pada konsentrasi tinggi akan mengganggu metabolisme sel, akibatnya menghambat proses pembentukan bunga.
Sitokinin adalah salah satu faktor penting yang mengatur pembentukan percabangan bunga dan perkembangan buah. Pada tanaman leci, penyemprotan kinetin lebih efektif dalam menginduksi pembentukan cabang bunga dibandingkan jenis sitokinin yang lain ( Chen 1990).
Pada tanaman anggur, sitokinin mendorong differensiasi primordia menjadi inflorensen. Dalam hal ini sitokinin meningkatkan kemampuan inflorensen anggur untuk menarik assimilat karena inflorensen yang sedang berkembang memiliki daya saing yang rendah dibandingkan daun-daun muda tanpa adanya sitokinin (Kinet et al. 1985).
Pada tanaman jarak pagar andromonoecious, menunjukkan bahwa bunga hermaprodit selalu muncul dari pucuk apikal infloresen. Raju dan Ezradanam (2002) menyatakan bahwa tanaman jarak pagar menghasilkan infloresen dengan tipe
dichasial cyme. Perlakuan ZPT secara
umum tidak mempengaruhi pola infloresen bunga, tetapi mempengaruhi pola percabangan pada cabang sekunder. Tanaman jarak pagar dengan penyemprotan GA3:K (50:50 ppm) menghasilkan percabangan infloresen yang paling banyak. Menurut Palupi et al. (2009) bahwa aplikasi sitokinin menghasilkan percabangan infloresen yang lebih banyak sehingga bunga yang terbentuk juga lebih banyak. Dari hasil pengamatan dapat terlihat bahwa pola infloresen bunga pada tanaman jarak pagar monoecious berbeda dengan tanaman andromonoecious. Pucuk apikal tanaman jarak pagar monoecious lebih didominasi oleh bunga jantan, sedangkan pucuk apikal pada tanaman jarak pagar andromonoecious berpotensi menghasilkan bunga hermaprodit.
SIMPULAN
Kandungan giberelin dan auksin tanaman jarak pagar monoecious lebih tinggi pada fase sebelum berbunga dibandingkan setelah proses pembungaan, sedangkan kandungan kinetin pada tanaman jarak pagar monoecious lebih rendah saat sebelum berbunga dibandingkan setelah berbunga. Pada tanaman jarak pagar kontrol andromonoecious kandungan auksin dan kinetin mengalami kenaikan setelah proses pembungaan, sedangkan giberelin mengalami penurunan. Penyemprotan auksin dan giberelin terhadap tanaman andromonoecious mampu meningkatkan kandungan auksin dan giberelin endogen. Penyemprotan kinetin pada tanaman jarak pagar andromonoecious dapat meningkatkan jumlah percabangan infloresen bunga.
DAFTAR PUSTAKA
Abidin Z. 1985. Dasar-Dasar Pengetahuan Tentang Zat Pengatur Tumbuh. Bandung: Angkasa.
Arika KA, Hastuti ED, Setiari N. 2009. Pertumbuhan dan pembungaan tanaman jarak pagar setelah penyemprotan GA3 dengan konsentrasi yang berbeda. Jurnal Penelitian Sains & Teknologi. 10 (1) : 18 – 29.
6
reproductive growth. Vol II. Florida: CRC Press.
Chen, WS. 1990. Changes in citokinins before and during early flower bud differentiation in Lychee (Litchi chinensis Sonn.) Plant Physiol. 96: 1203-1206.
Darnell J, Lodish H, Baltimore H. 1986.
Molecular Cell Biology. New York: Scientific American Books, Inc. Delita K, Mareza E, Kalsum U. 2008.
Korelasi aktivitas enzim nitrat reduktase dan pertumbuhan beberapa genotipe tanaman jarak pagar (Jatropha Curcas Linn.) yang diperlakukan dengan zat pengatur tumbuh 2,4 D. Acta 11: 80-86.
Hartati S. 2009. Info tek perkebunan media bahan bakar nabati dan perkebunan. Bogor: Badan Penelitian dan Pengembangan Pertanian. Vol.1. No. 1. 2009.
Heller, J. 1996. Physisc nut Jatropha curcas
L. promoting the conservation and used undertilized and neglected crops 1. Institut of Plant Genetic and Crop Plant Research . Gatersleben/IPGRI. Kaminek M, Motyka V, Vankova R. 1997.
Regulation of cytokinin content in plant cell. PlantPhysiol. 101: 689-700. Kinet JM, Bernier G, Sachs RM. 1985. The
Initiation of Flowers: The physiology of flowering. Vol I. Florida: CRC Press.
Lakitan, B. 1995. Fisiologi Pertumbuhan dan Perkembangan Tanaman. Jakarta: PT Raja Grafindo Persada.
Palupi ER, Surahman M, Warid K. 2009. Aplikasi ZPT untuk keserempakan pemasakan buah jarak pagar (Jatropha curcas L.). di dalam: Prosiding seminar hasil-hasil penelitian IPB 2009. Bogor.
Prawitasari T, Guhardja E, Darusman LK, Harran S, Poerwanto R. 2002.
Perkembangan struktur meristem reproduktif pada proses pembungaan tanaman lengkeng. Hayati 9(4):119-124.
Rai Y, Poerwanto R, Darusman LK, Purwoko BS. 2006. Perubahan kandungan giberelin dan gula total pada fase-fase perkembangan bunga manggis. Hayati 13(3): 101-106. Raju S, Ezradanam V. 2002. Pollination
ecology and fruiting behaviour in a monoecious spesies, Jatropha curcas
L. (Euphorbiaceae). Curr Sci 83: 1395-1398.
Ryugo K. 1990. Flowering and fruit set in temperate fruit trees. di dalam:
Proceedings of the lnternasional Seminar "Off Season Production of Horticultural Crops ". Taipei, 27 Nov-3 Des 1989.
Salisbury FB, Ross CW. 1995. Fisiologi Tumbuhan Jilid 3. Diah RL, Sumaryono, penerjemah. Bandung: ITB Press. Terjemahan dari: Plant Physiology.
Tumbull CGN, Anderson KL, Winston EC. 1996. Influence of gibberellins treatment on flowering and fruiting patterns in mango. Aust J Agric 36: 603-611.
Unyayar S, Topcuoglu SF, Unyayar A. 1996. A modified method for extraction and identification of indole-3- acetic acid (IAA), giberelin acid (GA3), abcisic acid (ABA) and zeatin produced by phanerochaete chrysosporium ME446. Bulg.J. Plant physiol . 22 (3-4): 105-110
7
8
Lampiran 1 Spektrum panjang gelombang giberelin
Lampiran 2 Spektrum panjang gelombang kinetin
No. Wavelength nm. Abs. 1 365,4 -0,001 2 361,8 -0,001 3 354,6 -0,001 4 346,2 -0,001 5 344,2 -0,001 6 338 -0,001
7 333 0
8 320,4 0 9 303,2 0
10 253 0,302
11 206,2 3,329 12 202,8 4
No.
Wavelength nm. Abs. 1 365,4 0,005 2 361,6 0,005 3 350,2 0,004 4 337,6 0,006 5 331,6 0,006 6 315,2 0,005
7 267,8 1,836
8 205,8 4
9
Lampiran 3 Spektrum panjang gelombang auksin
No. Wavelength nm. Abs. 1 369,6 0,002 2 363,2 0,002 3 354,6 0,002 4 348,4 0,002 5 337,6 0,002 6 330,8 0,002 7 325,2 0,002 8 289,4 0,69
9 279,6 0,808
10 273,8 0,768 11 223,2 3,311 12 217,6 3,161 13 215 3,101 14 206 2,376
10
Lampiran 4 Hasil pengukuran kurva standar giberelin
Lampiran 5 Hasil pengukuran kurva standar auksin
Lampiran 6 Hasil pengukuran kurva standar kinetin
ppm ppm