BIOSISTEMATIKA MANGGA INDONESIA
FITMAWATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Biosistematika Mangga Indonesia adalah karya saya dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Agustus 2008
Fitmawati
ABSTRACT
FITMAWATI. Biosistematics of Indonesian Mangoes. Under the supervisions of ALEX HARTANA, MIEN A. RIFAI, and BAMBANG S. PURWOKO
Biosystematics of Indonesian mango were investigated. It consisted of reinstatement of Mangifera laurina and their related species, phylogenetic study based on cpDNA trnL-F intergenic spacer, genetic diversity and taxonomy of cultivated mango in practice. According to flower characters, M. rubropetala
Kosterm. considered as synonim of M. indica L. Otherwise M. laurina Bl., M. aplanata Kosterm, and M. lalijiwa Kosterm. is suggested as separate species from
M. indica L. Based on E-RAPD, specific bands distinguished M. aplanata from other related species. Phylogenetic analysis of the trnL-F intergenic spacer region placed the mangga Hiku (Mangifera sp) as the common ancestor or progenitor among the other M. laurina and supported well sister-group to monophyletic in
Mangifera laurina complex. Identification and characterization of germplasm of 76 cultivars from Cukurgondang-Pasuruan East Java based on 92 morphological characters formed three major groups and nine groups of cultivars with coefficient of genetic similarity at 0.38-0.85. Eighty two cultivars were analyzed with RAPD markers resulting in genetic similarity coefficient between 0.69-0.98. Combination analysis of morphological and RAPD markers of the 76 cultivars resulted in nineteen major clusters, with 0.69-0.88 similarity coefficient. This investigation will help breeders in mango improvement program. Available classifications on cultivated plants showed a merger two approaches; a botanical one, based on essential systematic plant characters, and a practical one, based on the analysis of commercial properties. A taxonomic study of cultivars grown in Indonesia based on morphology and agronomy characters resulted in 84 cultivars recognized. Eigth main cultivar-groups (i.e. Berem, Madu, Gedong, Golek, Bapang, Arumanis, Kepodang, and Kebo) and seventeen cultivars-groups were established from eighty four cultivars. The ‘Lalijiwo’ cultivar was the synonym of ‘Thaber’, ‘Tabar’, ‘Gurih’, while ‘Arumanis’ was the synonym of ‘Gadung’. Meanwhile, the cultivar ‘Kates277’ included in Golek main group and homonym with the cultivar ‘Kates’ in Arumanis main group. The cultivar ‘Nanas93’ one of Madu cultivar main group was different from the cultivar ‘Nanas71’ in cultivar main group Bapang.
RINGKASAN
FITMAWATI. Biosistematika Mangga Indonesia. Dibimbing oleh ALEX HARTANA, MIEN A. RIFAI, dan BAMBANG S. PURWOKO
Kajian Biosistematika mangga Indonesia meliputi tinjauan status dan kedudukan taksonomi Mangifera laurina Bl dan kerabat dekatnya yang terdiri dari M. aplanata Kosterm., M. rubropetala Kosterm., M. lalijiwa Kosterm., dan
M. indica, serta hubungan filogentiknya, keanekaragaman genetik kultivar mangga Indonesia dan pengelompokan kultivar mangga dalam praktek. Plastisitas ciri morfologi yang besar di antara mangga dan kerabat dekatnya menjadi penyebab batasan jenis yang sudah ada sering diperdebatkan. Penelitian ini mengurai kembali konsep jenis Kostermans & Bompard yang bertentangan dengan konsep Kochumen, keduanya membangun konsep jenisnya berdasarkan ciri morfologi. Spesimen yang diperiksa ditambah 609 yang berasal dari Sumatera, Jawa, Kalimantan, Sulawesi dan Maluku Analisis mengunakan data ciri morfologi, DNA sitoplasmik cpDNA trnL-F (intergenic spacer) dan DNA inti. M. laurina dan kerabat dekatnya dapat dibedakan menjadi empat jenis yaitu
M. indica, M. laurina, M. lalijiwa, dan M. aplanata, sedangkan M. rubropetala
menjadi sinonim M. indica. Analisis hubungan kekerabatan menghasilkan dua sister grup yaitu M. indica dan M. lalijiwa. serta M. laurina dan M. aplanata. Penanda E-RAPD dapat membedakan M. aplanata (Depeh) terhadap kerabat dekat lainnya. M. laurina ‘Dodol Ternate’, M. rubropetala (sinonim M. indica),
M.laurina ‘Betoel’, M. indica ‘Golek’,dan M. indica ‘Cengkir’.
Penelusuran nenek moyang M. laurina dan kerabatnya menggunakan cpDNA trnL-F intergenic spacer menunjukkan mangga Hiku sebagai kerabat liar dan diduga sebagai tetua bersama M. laurina dan kerabatnya. Selain itu, memperkuat Selawesi sebagai pusat keanekaragaman M. laurina dan kerabatnya, dan Indonesia adalah pusat keanekaragaman mangga dunia. Pengelompokan kultivar mangga berdasarkan 92 ciri morfologi dan penanda RAPD berbeda dengan pengelompokan yang selama dikenal oleh masyarakat mangga Indonesia. Keanekaragaman genetik mangga Indonesia tergolong luas yaitu 15-62% (morfologi), 2-31% (RAPD), dan 12-40% (kombinasi kedua penanda). Berdasarkan pengelompokan kultivar mangga koleksi KP Cukurgondang-Pasuruan Jawa Timur tidak didapatkan duplikasi aksesi, sehingga semua aksesi perlu dikonservasi untuk kelestariannya. Pengelompokan berdasarkan ciri agronomi ciri buah terhadap 84 kultivar mangga menghasilkan 8 kelompok utama yaitu Berem, Golek, Kepodang, Bapang, Arumanis, Gedong, Madu, dan Kebo. Tujuh belas kelompok kultivar dan 84 kultivar. Sinonim, kultivar ‘Lalijiwo’ sama dengan ‘Thaber’, ‘Tabar’, ‘Gurih’ dan ‘Arumanis’ sama dengan Gadung. Homonim yang dijumpai adalah pada ‘Kates277’ adalah anggota kelompok utama Golek, sedangkan kultivar ‘Kates’ adalah anggota kelompok utama Arumanis. Kultivar ‘Nanas93’ anggota kelompok utama Madu berbeda dengan ‘Nanas71’ anggota kelompok utama Bapang
Kata kunci: Keanekaragaman, genetik, mangga Indonesia, morfologi, cpDNA
© Hak cipta milik IPB, tahun 2008 Hak cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik ataupun tinjauan suatu masalah.
b. Pengutipan tidak merugikan kepentingan yang wajar IPB.
BIOSISTEMATIKA MANGGA INDONESIA
FITMAWATI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Biologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Biosistematika Mangga Indonesia
Nama : Fitmawati
NRP : G 361020061
Disetujui Komisi Pembimbing
Prof Dr Ir Alex Hartana, MSc Ketua
Prof Dr Mien A. Rifai Prof Dr Ir Bambang S Purwoko, MSc Anggota Anggota
Diketahui
Ketua Program Studi Biologi Dekan Sekolah Pascasarjana
Dr Ir Dedy Duryadi, DEA Prof Dr Ir Khairil A. Notodiputro, MS
Penguji pada Ujian Tertutup : Dr Ir Tatik Chikmawati MSi
Staf Pengajar Departemen Biologi, FMIPA IPB
Penguji pada Ujian Terbuka : Dr Ir Sobir, MS
Staf Pengajar Departemen Agronomi dan Hortikultura, Fakultas Pertanian IPB Dr Dra Titi Rugayah, MSc
PRAKATA
Puji syukur kehadirat Allah Yang Maha Sempurna atas segala limpahan rahmat dan karuniaNya tulisan ini dapat diselesaikan. Disertasi ini tentang Biosistematika mangga Indonesia yang meliputi tinjauan status dan kedudukan jenis mangga dan kerabat dekatnya serta hubungan filogentiknya, keanekaragaman genetik kultivar mangga Indonesia dan pengelompokan kultivar mangga dalam praktek.
Sejak penentuan topik sampai penyelesaian studi, penulis didukung oleh bebagai pihak secara perorangan maupun institusi. Maka pada kesempatan ini penulis menyampaikan terimakasih dan penghargaan yang tinggi kepada Bapak Prof Dr Ir Alex Hartana MSc, Prof Dr Mien A. Rifai, Prof Dr Ir Bambang S.Purwoko, selaku pembimbing yang telah banyak memberi saran, bimbingan, arahan, strategi, dan bahkan materi penelitian dari awal penelitian hingga selesai. Penulis juga menyampaikan penghargaan kepada Bapak Dr Ir Sobir MS, Ibu Prof. Dr Ir Syafrida Manuwoto MS, Bapak Prof Dr Sudarsono MSc (Staf Pengajar Agronomi). Ibu Dr Rugayah, Ibu Dr Yulita Kusuma Dewi, Bapak Dr Teguh, Dr Eko Baroto Waluyo, Bapak Dr Yohanis P. Mogea M.Sc (Staf LIPI Biologi), atas motivasi, penyegaran ide, dan arahan-arahannya.
Terima kasih juga disampaikan kepada Rektor Universitas Riau, Dekan FMIPA UNRI dan seluruh jajarannya yang telah banyak memberi kesempatan kepada saya melaksanakan studi S3. Kepada Dirjen Pendidikan Tinggi yang telah memberikan bantuan beasiswa BPPS selama 3.5 tahun. Kepada Pemimpin Riset Unggulan Nasional melalui Direktur Pusat Kajian Buah-buahan Tropika IPB atas bantuan biaya dan materi penelitian. Kepada Kepala LOKA-LIT Tanaman Buah dan Hortikultura di Tlekung Madang Jawa Timur, Kepala Kebun Percobaan Cukurgondang Pasuruan Jawa Timur berserta pegawai kebun yang banyak membantu dalam pengumpulan data dan sampel penelitian.
Ucapan terima kasih juga disampaikan kepada Ibu Dra. Eko Sri Sulasmi SU, Ibu Dra. Hermin Msc, di Malang Jawa Timur, dan Lely F. Djafar SPi MSi di Kendari. Kepada rekan-rekan seperjuangan Dr Soaloon Sinaga, Dr Nunik S. Ariyanti, Ir Amin Retnoningsih SU, Ir Donata Pandin MSi, Dr. Ir Mohammad Arief Nasoetion MSi, Mutmainna Ridwan SPi MSi, Drs Pudji Widodo, MSc, Sulassih SP, Dewi Indriyani Roslim SSi MSi, atas diskusi dan sumbangan waktu dan tenaga. Kepada Bapak Sutiyo, Bapak Heri Jumhair, dan Saudari Endang di Pusat Penelitian Sumberdaya Hayati dan Bioteknologi. Juga disampaikan terimakasih secara khusus dan penghargaan yang tinggi kepada ayahanda, ibunda, kakak, dan adik-adik atas doa dan pengorbanan yang tulus demi kesuksesan penulis.
Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2008
RIWAYAT HIDUP
Penulis dilahirkan di Bukittinggi pada tanggal 20 April 1973 dari ayahanda Sofyan Salim dan Ibunda Akmal Alwi. Penulis merupakan anak pertama dari empat bersaudara.
Pada tahun 1991 diterima sebagai mahasiswa Jurusan Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Andalas Padang, lulus Juli tahun 1996. Pada Agustus tahun 1998 dengan beasiswa TMPD, penulis mendapat kesempatan melanjutkan pendidikan sebagai mahasiswa S2 di Program Studi Biologi Sub-Prodi Taksonomi Institut Pertanian Bogor lulus Juli 2001. Kesempatan berikutnya tahun 2002 dengan beasiswa BPPS, pada program studi dan bidang ilmu yang sama di Institut Pertanian Bogor, penulis melanjutkan pendidikan Program S3
Penulis sejak tahun 1997 menjadi staf pengajar di Jurusan Biologi FMIPA Universitas Riau-Pekanbaru sampai sekarang. Bidang ilmu yang diampu adalah Biologi Umum, Taksonomi Tumbuhan, dan Metode Penelitian Taksonomi. Aktif sebagai anggota Perhimpunan Taksonomi Tumbuhan Indonesia. Seminar yang pernah diikuti diantaranya Seminar Internasional Bioteknologi Indonesia tahun 1997 dan Seminar Nasional Penggalang Taksonomi Tumbuhan Indonesia 2006.
DAFTAR TABEL……… x
DAFTAR GAMBAR... xi
I. PENDAHULUAN ... 1
II. TINJAUAN STATUS Mangifera laurina Bl. DAN KERABAT DEKATNYA Tinjauan Status Mangifera laurina Bl. dan Kerabat Dekatnya... 5
Abstract... 5
Pendahuluan... 5
Bahan dan Metode... 7
Hasil dan Pembahasan... 8
Simpulan... 17
Studi Filogenetik Mangifera laurina dan Kerabat Dekatnya Menggunakan Penanda cpDNA trnL-F Intergenik Spacer... 18
Abstract... 18
Pendahuluan... 18
Bahan dan Metode... 19
Hasil dan Pembahasan... 21
Simpulan... 25
III. SISTEMATIKA KULTIVAR MANGGA INDONESIA Keanekaragaman Kultivar Mangga Indonesia Berdasarkan Penanda Morfologi dan RAPD... 26
Abstract... 26
Pendahuluan... 26
Bahan dan Metode... 28
Hasil dan Pembahasan... 30
Simpulan... 40
Taksonomi Budidaya Mangga Indonesia Dalam Praktek... 41
Abstract... 41
Pendahuluan... 41
Bahan dan Metode... 42
Hasil dan Pembahasan... 43
Simpulan... 58
PEMAHASAN UMUM... 59
SIMPULAN UMUM... 67
DAFTAR TABEL
Halaman
1 Sifat ciri yang digunakan untuk analisis filogeni mangga ... 10 2 Delapan puluh tiga kultivar mangga asal kebun koleksi KP
Cukurgondang-Pasuruan Jawa Timur yang dijadikan bahan
Penelitian... 29 3 Koefisien kemiripan dan koefisien kopenetik mangga Indonesia
Berdasarkan penanda morfologi, RAPD, dan kombinasi kedua
Penanda... 39 4 Ciri diagnosis kelompok utama kultivar mangga... 44 5 Kelompok utama dan kelompok kultivar mangga asal KP Cukur
DAFTAR GAMBAR
Halaman
1 Kladogram paling parsimoni berdasarkan ciri morfologi... 12
2 Profil pita DNA M. aplanata dan M. indica masing-masing dengan primer E2 dan E1E3... 14
3 Dendrogram M. laurina dan kerabat dekatnya dengan penanda E-RAPD... 15
4 Dendrogram M. laurina dan kerabat dekatnya dengan kombinasi penanda E-RAPD dan morfologi... 16
5 DNA produk PCR yang diperiksa dalam agarose 1.2%... 21
6 Kladogram 6 aksesi mangga dan grup luarnya berdasarkan Penanda trnL-F... 24
7 Kladogram 6 aksesi mangga berdasarkan penanda trnL-F dengan Metode Neighbour Joining (Saitou dan Nei 1987)... 25
8 Dendrogram mangga Indonesia berdasarkan penanda morfologi... 32
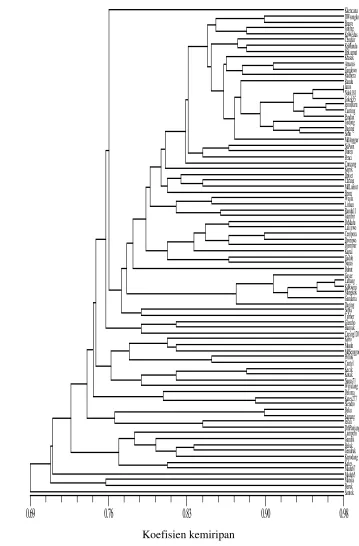
9 Dendrogram mangga Indonesia berdasarkan penanda RAPD…... 36
10 Dendrogram mangga Indonesia berdasarkan penanda morfologi Dan RAPD……….. 38
11 Dendrogram kelompok utama mangga berdasarkan penanda morfologi... 46
12 Kelompok utama Berem... 47
13 Kelompok utama Golek... 48
14 Kelompok utama Kepodang... 49
15 Kelompok utama Gedong... 50
16 Kelompok utama Madu... 51
17 Kelompok utama Bapang... 52
18 Kelompok utama Arumanis... 53
19 Kelompok utama Kebo... 53
Mangga termasuk marga Mangifera L. dari keluarga Anacardiaceae. Marga ini setidaknya mempunyai 14 jenis mangga yang biasa dimakan dari 68 jenis mangga yang ada (Kostermans & Bompard 1993). Secara morfologi ke-14 jenis mangga tersebut mirip dan sulit dibedakan. Pada umumnya yang dikenal sebagai mangga adalah anggota Mangifera indica. Mangifera lainnya yang dapat dimakan mempunyai kualitas buah yang lebih rendah dan umumnya dikenal sebagai mangga liar (kerabat mangga). Marga Mangifera berasal dari Asia tropika, sebagian besar jenisnya ditemukan di Semenanjung Malaysia, Sumatera, Jawa, Kalimantan, dan Sulawesi seperti Mangifera laurina Bl., M. aplanata
Kosterm., M. lalijiwa Kosterm., dan M. indica L yang bersifat poliembrioni, sedangkan M. indica yang berasal dari India dan Myanmar bersifat monoembrioni.
Secara taksonomi, sejumlah ahli botani bersilang pendapat mengenai kedudukan jenis-jenis liar yang berkerabat dekat dengan M. indica meliputi M. laurina, M. aplanata, M. rubropetala Kosterm., M. orophila Kosterm., M. bompardii Kosterm., M. lalijiwa, dan M. longipes Griff. Kostermans dan Bompard (1993) membedakan jenis M. aplanata dari M. indica. M. laurina dari.
M. indica, M. aplanata dan M. rubropetala. Selain itu Kostermans dan Bompard (1993) juga membagi sebaran biogeografi Mangifera menjadi dua wilayah yaitu India-Myanmar dan Indo-Maleisia.
Kochummen (1996) tidak menyetujui pemisahan kerabat liar M. indica
tersebut menjadi jenis-jenis baru, melainkan menganggapnya sebagai sinonim M. indica. Pertimbangan Kochummen (1996) lainnya, jenis-jenis yang dikenali berbeda dan digunakan oleh Kostermans dan Bompard (1993) sebagai tipe, oleh penduduk di daerah asalnya dinamai nama daerah yang sama untuk semua jenis tersebut yakni empelam atau mempelam. M. aplanata, M. laurina, M. rubropetala, M. orophila, M. Bompardii, dan M. longipes hanyalah varian M. indica.
yang belum dikenali oleh Kochummen (1996), karena Kochummen hanya bekerja dengan Mangifera asal Borneo, sedangkan M. lalijiwa endemik di Pulau Jawa.
Mangga mudah beradaptasi pada lingkungan budidayanya dan merupakan salah satu komoditas buah tropis paling populer. Mangga telah dibudidayakan selama ribuan tahun dan menjadi bagian dari budaya di banyak tempat, sehingga penyebutan mangga berbeda-beda sesuai dengan kultur dan bahasa yang ada. Penamaan mangga yang berbeda tersebut mencerminkan asal-usul dan penyebarannya. Nama-nama mangga lebih mengikuti pola penamaan yang berkembang di kawasan Asia-Pasifik sesuai dengan daerah dan negara asalnya.
Di Indonesia dikenal serangkaian nama untuk merujuk lebih kurang 270 kultivar mangga diantaranya ‘Pari’, ‘Gadung’, ‘Kopyor’, ‘Pelem’, ‘Bapang’, ‘Kates’, ‘Dodol’, ‘Gedong’, ‘Golek’, ‘Cengkir’, ‘Sengir’, ‘Endok’, ‘Wangi’, ‘Kelapa’, ‘Kidang’, ‘Madu’, ‘Thaber’, ‘Tabar’, ‘Lalijiwa’, ‘Gurih’, ‘Nenas’, dan ‘Daging’ (Heyne 1927; Mukharejee 1949; dan Hou 1978). Selain itu, mangga ‘Arumanis’ di sebagian tempat juga dikenal sebagai mangga ’Gadung’, sedangkan mangga yang dikenal dengan nama ‘Lalijiwa’ di Solo, di Madura dikenali sebagai ‘Tabar’, dan ‘Manalagi Probolinggo’ di Probolinggo.
Kerancuan penamaan mangga dimungkinkan oleh plastisitas morfologi yang besar antar jenis-jenis mangga yang ada terutama M. indica kompleks yang disebabkan tingginya tingkat kesesuaian silang antar jenis mangga sehingga menghasilkan bentuk-bentuk antara yang sulit dibuat batasan jenisnya. Di sisi lain, untuk tujuan pendayagunaan, pengelolaan, dan konservasi plasmanutfah mangga memerlukan kejelasan nama dan batasan jenis maupun kultivar.
Identifikasi, karakterisasi, dan evaluasi kultivar mangga baik budidaya maupun kerabat dekatnya sampai saat ini belum tuntas dilakukan, terutama untuk kultivar mangga di luar Pulau Jawa. Sebagian kultivar mangga di Jawa telah diidentifikasi berdasarkan ciri morfologi oleh Kusumo et al. (1975) dan Efendy et al. (2003). Pertautan ciri antar kultivar mangga dan besarnya plastisitas ciri morfologi, menyulitkan dalam membuat batasan kultivar, sehingga perlu didukung sumber data dengan pendekatan lain yang lebih komprehensif.
Analisis ini, dapat dilakukan dengan menggunakan penanda molekuler baik pada DNA sitoplasma maupun pada DNA inti. Pada DNA sitoplasma, analisis genetik dapat dilakukan dengan perunutan (sekuensing) DNA kloroplas (cpDNA) yang relatif lebih konservatif dibanding DNA inti. Ciri molekuler juga dapat digunakan untuk mengidentifikasi kultivar dan menduga kekerabatan antar plasmanutfah, sehingga variasi genotipe antar kultivar dapat dibedakan dengan jelas dan dapat dihindari adanya duplikasi aksesi.
Analisis genetik pada DNA inti dapat dilakukan dengan mempelajari pola pemisahan pita DNA hasil amplifikasi teknik Random Amplified Polymorphic DNA (RAPD) atau dengan teknik Enhanced-RAPD (E-RAPD). Teknik RAPD dan E-RAPD merupakan penanda molekuler yang mengamplifikasi pita DNA secara acak dengan menggunakan primer acak dan bersifat dominan (Curley & Jung 2004). Teknik ini sederhana karena mudah dalam persiapan dan memberikan hasil relatif cepat, menghasilkan ciri relatif banyak. Selain itu, teknik ini sangat berguna dalam mengungkap dan menganalisis keanekaragaman genetik tanaman yang tidak diketahui latar belakang genomnya (Tingey et al. 1992).
Penanda RAPD bermanfaat untuk memisahkan antar spesies dan menentukan kemurnian kultivar (Curley & Jung 2004), dan kemungkinan untuk memperlihatkan hubungan kekerabatan antar kultivar yang mirip secara morfologi (Kusch & Heckmannn 1996), analisis variabilitas genetik tanaman Desmodium sumichrastii (Bedolla-Garcia & Lara-Cabrera 2006). Analisis RAPD telah dilakukan pada kultivar mangga di Australia (Bally et al. 1996), di Florida (Shcnell et al. 1995), dan di Venezuela (Lo'pez-Valezuela et al. 1997), di India (Kumar & Narayanaswamy 2001; Karihaloo & Dwivedy 2003). Analisis E-RAPD telah dilakukan terhadap tanaman teh (Tanaka & Tanighuci 2002), pada tanaman manggis (Sobir et al. 2008). Penanda cpDNA telah digunakan pada analisis filogeni Pinus lamberiana (Liston et al. 2008), dan filogeni Anisophylleaceae (Zang et al. 2007).
Tujuan penelitian ini adalah untuk meninjau status jenis M. laurina dan kerabat dekatnya dengan penanda morfologi dan E-RAPD, menganalisis hubungan filogenetik M. laurina dan kerabat dekatnya dengan penanda cpDNA
ABSTRACT
Mangifera laurina Bl. and the related species, e.g. M. rubropetala Kosterm., M. lalijiwa Kosterm. were treated as synonym of M. indica L. This study is aimed at investigating the taxonomical status of those taxa. Current discussion focuses on whether they represent natural species or should be placed in several different species. The result showed that morphological phylogenetic analysis supported for separating four monophyletic species (M. laurina, M. aplanata, M. lalijiwa), considering that M. rubropetala and M. indica are synonym, and defining two independent lineage of sister group (i.e. the clade of M. lalijiwa and the clade consist of M. indica, M. laurina, M. aplanata, and M. rubropetala. Morphologically, M. lalijiwa is characterized by having glomerulate and glabrous in inflorescence and tips of petal ridge free of the surface. In contrast, the other species usually posseses non-glomerulate and puberulous in inflorescence and tips of petal at free of the surface. Phylogenetic analysis of E-RAPD was incongruent with that of morphological analysis. However, the molecular phylogeny support that M. aplanata is separated from other species and cultivars of M. lalijiwa are a monophyletic group.
Keywords: Mangifera indica, M. laurina, M. aplanata, M. rubropetala, M. lalijiwa,morphology, Enhanced-RAPD.
PENDAHULUAN
Mangifera laurina Bl. adalah salah satu anggota marga Mangifera yang berkerabat dekat dengan M. indica L. Sebagian besar masih tumbuh liar dan tersebar dari Pulau Sumatera, Jawa, Kalimantan, Sulawesi sampai Maluku. Kerabat lainnya adalah M. aplanata Kosterm., M. rubropetala Kosterm., M. orophila Kosterm., M. bompardii Kosterm., M. lalijiwa Kosterm., dan M. longipes Griff. Ciri bersama yang dimiliki oleh M. laurina dan kerabat dekatnya adalah ciri organ bunga kelipatan 4-5 merous, jumlah stamen fertil 1-2, dan biji tidak berupa labirin. Ciri yang membedakan jenis-jenis di atas adalah ciri perbungaan tidak kompak (non-glomerulate), permukaan bawah daun memiliki retikulasi rapat dan perhiasan bunga linear-oblong 4-5.5 mm yang dimiliki oleh
aplanata menjadi pembeda antara M. laurina dan M. aplanata. Pada M. rubropetala perubahan warna perhiasan bunga menjadi merah setelah antesis dipakai sebagai penciri untuk membedakannya dari M. indica. Kochummen (1996) tidak mengakui batasan jenis-jenis yang dibuat oleh Kosterman & Bompard (1993). Menurut Kochummen (1996), ciri morfologi yang digunakan Kosterman & Bompard (1993) dalam membuat batasan jenis tidak cukup untuk membedakan antara M. laurina, M. aplanata, M. rubropetala, M. orophila, M. bompardii, dan M. longipes dari M. indica. Selain itu, berdasarkan nama daerah spesimen empelam dan mempelam yang diperiksa oleh Kosterman & Bompard (1993) adalah nama yang sama untuk merujuk M. indica, M. laurina, dan M. aplanata. Kochummen (1996) juga menyatakan M. aplanata, M. laurina., M. rubropetala, M. orophila, M. bompardii., dan M. longipes hanyalah varian M. indica, sehingga nama-nama jenis tersebut adalah sinonim M. indica.
M. lalijiwa dibedakan dari M. laurina dan M. indica berdasarkan ciri daun menjangat (corioceus), guratan (ridge) menyatu pada pangkal perhiasan bunga (Kostermans & Bompard 1993). Jenis ini tidak diverifikasi oleh Kochummen (1996), karena Kochummen hanya bekerja dengan Mangifera asal Borneo, sedangkan M. lalijiwa endemik di Pulau Jawa. Dalam menyusun konsep jenisnya Kostermans & Bompard (1993) menguji spesimen asal Pulau Sumatera, Jawa, dan Borneo, sedangkan spesimen asal Sulawesi yang dieksplorasi tahun 1993 oleh Wirawan dan kawan-kawan belum diperiksa oleh Kostermans & Bompard (1993). Status jenis M. laurina dan kerabat dekatnya perlu ditinjau ulang berdasarkan spesimen dan ciri yang lebih lengkap serta berasal dari daerah lain, seperti Pulau Sulawesi yang mempunyai keanekaragaman mangga tinggi, tetapi belum diperiksa oleh peneliti terdahulu.
Gambaran menyeluruh suatu jenis dapat diperoleh dengan menggunakan berbagai pendekatan seperti ciri morfologi yang mudah diamati maupun ciri molekuler yang lebih banyak dan polimorf serta tidak dipengaruhi oleh lingkungan. Penelitian molekuler terdahulu pada Mangifera telah dilakukan oleh Ediathong et al. (2000) yang mempelajari hubungan filogenetik antar 14 jenis
Analisis kluster UPGMA sub-marga Mangifera berdasarkan pola pita RAPD dan kesahihan terhadap pengelompokan berdasarkan morfologi bunga dilakukan oleh Bompard & Schnell (1998).
Penanda Enhanced (Emphasized)-Random Amplified Polymorphic DNA (E-RAPD) dapat digunakan untuk mengklarifikasi status taxa dan untuk mempelajari hubungan kekerabatan pada tingkat jenis dan taksa di bawahnya. Penanda ini mempunyai kesamaan dengan RAPD pada umumnya, tetapi pada E-RAPD jumlah basa (mer) primer yang digunakan ditambah 1-2 basa sehingga berjumlah 11-12 basa. Penambahan basa tersebut untuk meningkatkan kejelasan pita dan spesifikasi penempelan primer pada DNA cetakan (templat) (Tanaka & Taniguchi 2002).
Penelitian ini bertujuan meninjau ulang status jenis M. laurina dan kerabatnya yang meliputi M. indica, M. aplanata,M. rubroptala, dan M. lalijiwa
berdasarkan ciri morfologi dan E-RAPD, dan menganalisis keanekaragaman dan kekerabatannya berdasarkan kemiripan genetik dengan penanda morfologi dan E-RAPD.
BAHAN DAN METODE
Bahan
Spesimen tanaman mangga yang diperiksa dikumpulkan dari seluruh Indonesia sebanyak 150 spesimen dan 459 spesimen Herbarium Bogoriense, termasuk spesimen voucer dari Herbarium Leiden (L), Kepong (Kep.) dan Sarawak (Sar.) sedangkan yang dianalisis filogeni berdasarkan ciri morfologinya sebanyak 16 sampel mewakili variasi yang ada. Analisis molekuler mengggunakan DNA yang diekstraksi dari spesimen segar seperti pada pengamatan morfologi dan mewakili variasi di dalam jenis sebanyak 14 spesimen mangga (4 jenis mangga) dan 2 jenis grup luar (outgroup) M. kemanga dan
Metode
Pengamatan morfologi mengacu pada kriteria yang digunakan oleh Rifai (1976) dan Vogel (1987). Analisis molekuler, ekstraksi DNA dari daun muda tanaman mangga mengikuti prosedur CTAB (Doyle & Doyle 1987) dengan beberapa modifikasi. Teknik E-RAPD mengikuti metode (Tanaka & Taniguchi 2002). Primer RAPD yang digunakan merupakan hasil seleksi dari 27 primer acak. Primer SBH 13 merupakan primer yang dipilih dan dimodifikasi dengan menambah satu basa, sehingga tingkat keterulangannya menjadi lebih baik dibanding primer awalnya. Primer SBH 13 ((5’-GACGCCACAC-3’) setelah ditambah 1 basa diperoleh primer E1 GACGCCACACT-3’), E2 (5’-GACGCCACACG-3’), E3 (5’-GACGCCACACA -3’), dan E4 (5’-GACGCCACACC-3’). Selain ke-4 primer tersebut, digunakan juga primer kombinasi E1E2 dan E1E4.
Kemunculan pita yang diperoleh dari teknik E-RAPD diterjemahkan menjadi data biner. Setiap pita mewakili satu karakter dan diberi nilai 1 bila ada pita dan 0 bila tidak ada pita. Analisis data berdasarkan koefisien Jaccard dengan metode UPGMA (Unweighted Pair-Group Method Arithmetic Avarage) menggunakan program NTSYS (Numerical Taxonomy and Multivariate System) versi 2.02 (Rolf 1998). Analisis kladistik data morfologi berdasarkan langkah paling parsimoni (maximum parsimony) menggunakan PAUP versi 4.0b8 (Swofford 2002) dengan bootstrap 1000 kali.
HASIL DAN PEMBAHASAN
Kajian tinjauan status Mangifera laurina Bl. dan kerabatnya dilakukan terhadap 609 spesimen hasil eksplorasi dan spesimen herbarium, terdiri dari 8 jenis Mangifera berdasarkan batasan jenis Kosterman & Bompard (1993) yang terdiri atas M. indica L., M. aplanata Kosterm., M. rubropetala Kosterm., M. lalijiwa Kosterm., M. orophila Kosterm., M. bompardii Kosterm., M. longipes
Ciri kunci yang digunakan penelitian ini sama seperti yang dipakai Kostermans & Bompard (1993), yaitu bentuk dan retikulasi daun, kelipatan organ bunga, kekompakan perbungaan, dan warna petal setelah anthesis. Tiga ciri kunci baru yang tidak digunakan Kostermans & Bompard (1993) adalah bentuk tajuk, kerapatan dan percabangan areola pada permukaan daun bagaian bawah, dan ada tidaknya rambut pada perbungaan maupun bagian-bagian bunga dan hasil analisis ini diperoleh 4 jenis Mangifera berbeda yaitu M. indica, M. aplanata, M. lalijiwa
dan M. laurina, sedangkan M. rubropetala memiliki sifat ciri warna petal merah setelah antesis yang merupakan ciri kontinyu dan berada dalam batasan ciri yang dimiliki oleh M. indica. Sifat ciri kontinyu ini tidak dapat digunakan sebagai ciri pembeda antar jenis, sehingga diusulkan menjadi sinonim M. indica.
Secara morfologi mangga M. laurina dan kerabat dekatnya memiliki keanekaragaman pada ciri fenologi pohon, bentuk daun, pola retikulasi mesofil daun, struktur bunga, rambut pada bunga dan perbungaan, guratan (ridge) pada perhiasan bunga, warna perhiasan bunga sebelum dan setelah antesis, bentuk, dan warna buah (Tabel 1). Berdasarkan bentuk tajuk pohon, mangga dibedakan atas tajuk membulat yang dimiliki oleh M. lalijiwa dan M. laurina, sedangkan 2 jenis lainnya memiliki bentuk tajuk jorong ke atas dan ke samping. Warna daun hijau tua dimiliki oleh M. lalijiwa sedangkan jenis lainnya memiliki warna daun hijau muda sampai hijau. Warna daun muda juga beraneka yaitu hijau muda sampai kuning kehijauan, kuning muda kecoklatan, merah kecoklatan, dan coklat kehitaman. Perbedaan warna pucuk ini dapat dipakai sebagai penciri kultivar pada
M. indica. Pola retikulasi pada M. lalijiwa rapat dengan dua percabangan, M. aplanata memiliki retikulasi rapat dengan percabangan lebih dari dua, M. indica
dan M. rubropetala retikulasi bercabang dua dan jarang, sedangkan M. laurina
memiliki retikulasi rapat dan bercabang banyak
Perhiasan bunga berwarna putih sampai putih kotor dimiliki oleh M. laurina, sedangkan jenis lainnya memiliki warna bunga kuning muda. Warna perhiasan bunga setelah antesis berkisar antara kuning, merah muda sampai merah. M. rubropetala dan sebagian anggota M. indica memiliki warna perhiasan bunga merah muda sampai merah setelah antesis, jenis lainnya memiliki perhiasan bunga berwarna kuning setelah antesis.
Adanya guratan (ridge) pada perhiasan bunga juga memperlihatkan perbedaan. M. indica dan M. rubropetala memiliki 3 guratan (ridge) yang tidak menebal dan menyatu pada sepertiga bagian perhiasan bunga. M. lalijiwa
Bentuk, warna, rasa, dan aroma buah merupakan bagian yang paling bervariasi yang dimiliki oleh M. laurina dan kerabatnya, tidak kurang dari 200 variasi bentuk buah dijumpai selama pengamatan. Ciri bentuk buah ini dapat dipakai untuk membedakan M. laurina dan kerabatnya.
Kunci berdasarkan ciri morfologi
1. a. Retikulasi mesofil bawah daun jarang, areola bercabang 2, bunga glomerulate, puberolous, guratan menyatu pada bagian tengah perhiasan bunga,3, tidak menebal ... M. indica
b. Bunga non-glomerolous, glabrous...2 2. a. Warna daun hijau tua, guratan 5 menebal, menyatu pada pangkal
perhiasan bunga...M. lalijiwa
b. Warna daun hijau Guratan 3, menebal, menyatu pada bagian tengah perhiasan bunga. ...3
3 a. Tajuk pohon membulat, perhiasan bunga kelipatan 5...M. laurina
b. Tajuk pohon jorong ke atas Perhiasan bunga kelipatan 4....M. aplanata
Analisis kladistik menggunakan PAUP * 4.0b4a (Swofford 2002), pada 14 taksa M. lalijiwa ‘Madu’, M. lalijiwa ‘Lalijiwa’, M. lalijiwa ‘Gurih Panjang’, M. indica ‘Golek’, M. indica ‘Arumanis’, M. indica ‘Kiyal’, M. indica ‘Kepodang’,
M. indica ‘Kates’, M. laurina ‘Betoel’, M. laurina ‘Hiku’, M. laurina ‘Tengguni’,
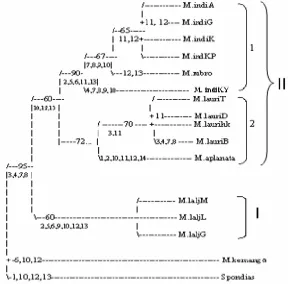
M. laurina ‘Dodol Ternate’, M. aplanata dan dua grup luar (outgroup) yaitu M. kemanga dan Spondias sp. Berdasarkan 14 ciri menghasilkan kladogram dengan 34 langkah, nilai CI (indeks consistensi) sebesar 0.818, HI (indeks homoplasi) 0.182, RI (indeks retensi) 0.866, berarti ciri apomorf tinggi dan homoplasi ciri rendah (Gambar 1). Pada kladogram ini tampak dua garis keturunan dari 2 kelompok bersaudara (sister group).
filogeni berdasarkan ciri morfologi terhadap M. lalijiwa mendukung pendapat Kostermans & Bompard (1993) yang memisahkannya dari M. indica.
Kelompok M. lalijiwa memiliki ciri warna perhiasan bunga sama dengan kelompok M. indica, tetapi berdasarkan ciri bentuk tajuk membulat, daun menjangat dan guratan mengalami penebalan juga merupakan ciri yang dimiliki oleh kelompok M. laurina.
Gambar 1. Kladogram paling parsimoni berdasarkan ciri morfologi Ket.: angka-angka pada percabangan adalah ciri diagnostik (merujuk pada Tabel 1).
Kelompok ke-II terdiri atas dua sub kelompok, yaitu sub kelompok M. indica (1) dan sub kelompok M. laurina (2) yang disatukan oleh ciri lembaran daun mengertas dengan nilai bootstrap 60. Sub kelompok M. indica terdiri atas
M. indica ‘Arumanis’, M. indica ‘Golek’, M. indica ‘Kates’, M. indica
bagian pangkal perhiasan bunga. Berdasarkan analisis filogeni terhadap ciri morfologi yang diamati M. rubropetala mengelompok bersama dengan kultivar-kultivar M. indica lainnya dan tidak dapat dipisahkan menjadi jenis yang berbeda. Nilai bootstrap yang mendukung pengelompokan ini adalah 90. Artinya, secara statistik kelompok ini dapat dipisahkan dengan tegas (kuat) dari sub kelompok M. laurina sebagai jenis yang berbeda.
Sub-kelompok M. laurina didukung oleh nilai bootstrap 72, terdiri dari M.
laurina ‘Tengguni’, M. laurina ‘Hiku’, M. laurina ‘Dodol Ternate’ dan M. laurina ‘Betoel’. Ke-4 kultivar M. laurina berasal dari tiga pulau berbeda yaitu dua dari Sulawesi Tenggara (M. laurina ‘Tengguni’ dan M. laurina ‘Hiku’), Ternate (M. laurina ‘Dodol Ternate’)danKalimantan Barat (M. laurina ‘Betoel’). Jumlah bagian-bagian bunganya kelipatan 4 pada M. aplanata dan berkelipatan 5 pada M. laurina dapat digunakan untuk membedakan M. aplanata dari M. laurina
Analisis Pengelompokan berdasarkan Penanda Enhanced-RAPD (E-RAPD)
Analisis E-RAPD menggunakan 16 spesimen segar mewakili variasi jenis yang diamati secara morfologi, yaitu M. indica ‘Arumanis’ (M. indicaA), M. indica ‘Kepodang’ (M. indicaKP), M. indica ‘Kiyal’ (M. indicaKY), M. indica
‘Kates’ (M. indicaK), M. indica ‘Golek’ (M. indicaG), M. laurina ‘Hiku’ (M. laurinaHK), M. laurina ‘Dodol Ternate’ (M. laurinaD), M. laurina ‘Betoel’ (M. laurinaB), M. laurina ‘Tengguni’(M. laurinaT), M. rubropetala (M. rubro), M. aplanata, M. lalijiwa ‘Madu’ (M. lalijiwaM), M. lalijiwa ‘Gurih Panjang’ (M. lalijiwaGP), dan M. lalijiwa ‘Lalijiwa’(M. lalijiwaL), dan grup luar M. kemanga
dan Spondias sp.
Pada primer E1E2 pita ukuran 500 pb tidak dimiliki oleh M. laurina
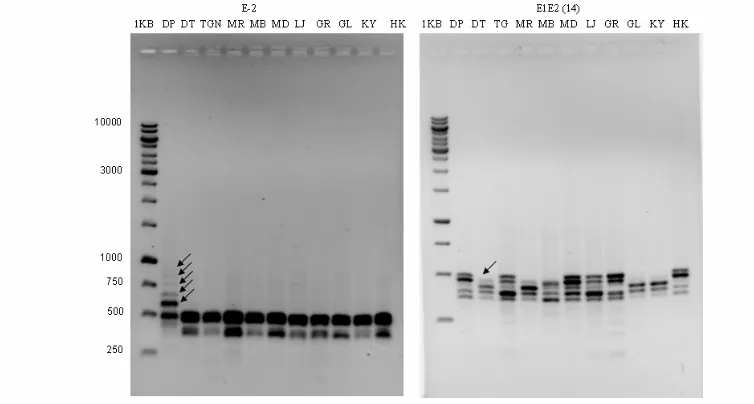
Analisis filogeni menghasilkan dendrogram yang memisahkan M. aplanata dari Mangifera lainnya. Berdasarkan pita yang dihasilkan primer E2 dengan ukuran 583 pb, 667 pb 750, 830 pb, dan 916 pb dan tidak terdapatnya pita dengan ukuran 333 pb pada primer E3 (Gambar 2), M. aplanata dapat dibedakan dari Mangifera pada tingkat ketidakmiripan 40% (Fitmawati et al. 2006).
Gambar 2. Profil pita DNA M. aplanata dan M. indica. masing-masing dengan primer E2 dan E1E3.
Ket.: DP (Depeh, M. aplanata), DT (Dodol Ternate), TGN (Tengguni, M. laurina), MR (M. rubropetala), MB (Mangga Betoel
M. laurina), MD (Madu), LJ (Lalijiwo), GR (Gurih, M. lalijiwa), KY (Kiyal/Cengkir) dan HK (Hiku M. laurina ).
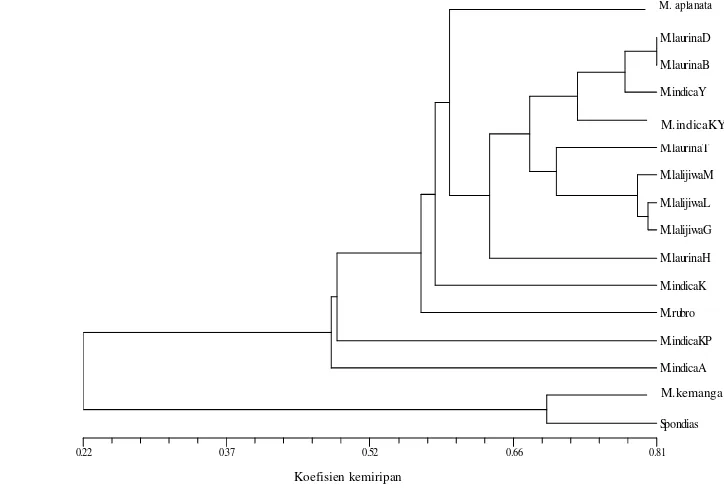
Selanjutnya, M. lalijiwa yang terdiri dari M. lalijiwa ‘Madu’, M. lalijiwa
‘Lalijiwa’ dan M. laljiwa ‘Gurih Panjang’ mengelompok bersama-sama (pada tingkat kemiripan 81%). Kelompok yang dihasilkan berdasarkan DNA secara konsisten sejalan dengan pengelompokan berdasarkan ciri morfologi. Hasil ini mendukung pengelompokan yang dihasilkan oleh Kosterman & Bompard (1993) yang memisahkan M. lalijiwa dari M. indica.
Berdasarkan penanda E-RAPD pengelompokan M. laurina dan M. indica
tidak membentuk pengelompokan secara terpisah antar jenisnya tetapi bersifat parafiletik karena anggota kedua jenis tersebut tersebar di antara anggota jenis
penciri yang dapat mengelompokkan M. indica dan M. laurina ke dalam satu kelompok. Oleh karena itu, pengelompokan M. indica dengan penanda E-RAPD harus didukung oleh ciri yang dihasilkan oleh penanda morfologi.
Gambar 3. Dendrogram M. laurina dan kerabat dekatnya dengan penanda E-RAPD.
Pengelompokan Mangifera Berdasarkan Penanda Morfologi dan E-RAPD
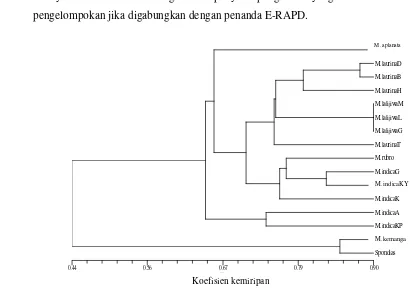
Spesimen yang digunakan dalam analisis kombinasi penanda morfologi dan E-RAPD sama dengan spesimen yang digunakan dengan analisis E-RAPD. Dilakukan analisis kombinasi kedua penanda untuk melihat pengelompokan yang lebih baik. Dendrogram yang dihasilkan dari kombinasi penanda morfologi dan E-RAPD (Gambar 4) memperlihatkan pola pengelompokan yang berbeda dengan dendrogram yang dihasilkan dari analisis filogeni berdasarkan ciri morfologi dan E-RAPD secara terpisah. Seperti pengelompokan menggunakan penanda morfologi, M. laurina dan kerabatnya pada kombinasi kedua penanda terpisah secara tegas menurut jenisnya. M. aplanata juga secara tegas dapat dipisahkan dari jenis lainnya. M. laurina mengelompok bersama seluruh anggota kultivarnya dan membentuk kelompok yang lebih besar bersama anggota kultivar M. lalijiwa.
Pengelompokan mangga berdasarkan kombinasi penanda morfologi dan DNA memberikan gambaran yang berbeda dengan pengelompokan berdasarkan DNA. M. indica dan kultivarnya membentuk kelompok bersama dengan M. rubropetala atau bentuk kelompok yang parafiletik. Pada Gambar 4 terdapat dua sub-kelompok pada M. indica dan kultivarnya yang dihasilkan berdasarkan kombinasi kedua penanda yaitu kelompok M. indica ‘Kepodang’ dan M. indica ‘ Arumanis’ yang terpisah pada pangkal percabangan dengan anggota M. indica
lainnya yaitu M. indica ‘Golek’, M. indica ‘Kiyal’ M. indica ‘Kates’, dan M. rubropetala yang lebih mengelompok dengan M. laurina, M. lalijiwa dan M. aplanata.
M. laurina membentuk kelompok tersendiri kecuali M. laurina ‘Tengguni’ yang membentuk kelompok dengan anggota lebih besar bersama anggota M. lalijiwa lainnya. Berdasarkan kombinasi kedua penanda ini kelompok M. lalijiwa
juga mampu memisah secara tegas dari kelompok lainnya pada kemiripan 90%. Pengelompokan ini mendukung pemisahan M. lalijiwa dari jenis M. laurina
lainnya. Penanda morfologi mempunyai pengaruh yang kuat dalam pengelompokan jika digabungkan dengan penanda E-RAPD.
SIMPULAN
Berdasarkan ciri bentuk tajuk, bentuk daun, bunga, dan buah diperoleh 4 jenis monofiletik yang berbeda yaitu M. laurina, M. indica, M. lalijiwa, dan M. aplanata. Hasil ini mendukung pendapat Kostermans & Bompard (1993), tetapi
M. rubropetala menjadi sinonim M. indica karena ciri kunci perhiasan bunga nyata, berdaging, kaku dan warna perhiasan bunga setelah antesis yang digunakan Kostermans & Bompard (1993) tidak dapat digunakan sebagai ciri diagnostik karena bersifat kontinyu.
Berdasarkan penanda E-RAPD, M. aplanata berbeda dengan kerabat dekat lainnya dan M. lalijiwa merupakan jenis monofiletik yang mengelompok bersama tiga kultivarnya. Analisis berdasarkan kombinasi penanda E-RAPD dan morfologi mendukung pemisahan M. laurina terhadap kerabat dekatnya yaitu M. indica, M. lalijiwa,dan M. aplanata seperti pengelompokan berdasarkan penanda morfologi. Pengelompokan kultivar M. laurina masih belum tegas karena masih tercampur dengan anggota M. indica lainnya, sedangkan hubungan antar kultivar
STUDI FILOGENETIK Mangifera laurina dan KERABAT DEKATNYA MENGGUNAKAN PENANDA cpDNA trnL-FINTERGENIK SPACER
(Phylogenetic study of M. laurina and related species based on cpDNA trnL-F intergenic spacer)
ABSTRACT
The phylogeny of Mangifera laurina and the related species were investigated using cpDNA intergenic spacer trnL-F sequence of the accession from Indonesia. Rutaceae was chosen as the outgroup. The objective of this study were 1) to reconstruct the phylogenetic relationships of these complex species within
Mangifera, and 2) to understand the infraspecific relationships within Mangifera
phylogenies with cpDNA trnL-F intergenic spacer sequences using maximum parsimony and neighbour joining as the optimal criteria. The result showed that
M. laurina from Celebes (mangga Hiku) was placed at the base of the phylogenetic tree of monophyletic group of M. laurina complex. It is suggested that mangga Hiku is the oldest cultivar among M. laurina complex and perhaps it is the progenitor of the complex species.
Key word; Mangifera laurina, phylogenetic, cpDNA trnL-F intergenic spacer, progenitor, Hiku
PENDAHULUAN
Klasifikasi mangga berbasis molekuler yang meliputi seluruh jenis dan wilayah di Indonesia belum banyak dilakukan, sehingga informasi filogeni molekuler masih sangat terbatas. Informasi filogeni molekuler sangat penting dalam rangka memperjelas kedudukan sistematika (klasifikasi), konservasi, dan menjadi data dasar keanekaragaman genetik untuk penangkar tanaman mangga dalam rangka perakitan mangga unggul Indonesia.
Penggunaan penanda berbasiskan DNA inti seperti E-RAPD menghasilkan pengelompokan antar kultivar mangga M. laurina dan kerabatnya bersifat parafiletik, kultivar-kultivar M. indica dan M. laurina tidak mengelompok pada cabang (klade) yang terpisah. Oleh karena itu perlu dicari penanda yang mengalami perubahan lebih lambat seperti DNA sitoplasmik (DNA kloroplas dan DNA mitokondria).
Penanda kloroplas (cpDNA) yang banyak digunakan adalah trnL-F
rendah (Bayer et al. 2000). Beberapa studi pada daerah non koding kloroplas memperlihatkan variasi yang lebih tinggi dan sering mengalami mutasi (Baldwin
et al. 1995) dalam bentuk transversi, transisi, insersi, dan delesi.
Daerah intergenic spacer antara trnL (UAA) 3’ exon dan gen trnF (GAA) juga berpotensi untuk studi filogenetik (Soltis et al. 1998). Daerah DNA ini mudah diamplifikasi dan disekuen, ukurannya relatif kecil 120-350 bp dan gen kopi tunggal (single copy), sehingga relatif mudah untuk menguji keseluruhan genom. Sekuen daerah trnL-F lebih informatif pada tingkatan marga dan jenis (Alejandro et al. 2005, Barfuss et al. 2005, Shaw et al. 2005).
Penggunaan penanda molekuler kloroplas (cpDNA) untuk mengungkap keanekaragaman, menelusuri hubungan kekerabatan berdasarkan evolusinya dan memperjelas kedudukan taksa mangga Indonesia belum pernah dilakukan. Penanda ini sangat bermanfaat untuk mendukung data molekuler mangga yang sudah ada sebelumnya, sekaligus untuk memahami evolusi mangga berdasarkan sekuen DNA kloroplas. Informasi evolusi mangga bermakna untuk memprediksi tetua bersama dari mangga yang ada di Indonesia saat ini. Penanda cpDNA telah banyak digunakan untuk studi filogeni tanaman lainnya. Misalnya Morus oleh Weiguo et al. (2005), Cucumis spp oleh Chung et al. (2006, 2007). cpDNA sering digunakan sebagai penanda karena mudah diisolasi dan dipurifikasi, dikarakterisasi, dan dikloning, dan sangat konservatif dengan laju evolusi yang rendah, sehingga dapat digunakan untuk rekonstruksi filogeni antar taksa pada tingkat famili tumbuhan berbunga (Clegg & Durbin 1990, Kajita et al. 1998).
Tujuan dari penelitian ini adalah untuk meninjau ulang hubungan antar kultivar M. laurina dan M. indica berdasarkan cpDNA trnL-F transgenic spacer, memata-matai evolusi yang terjadi pada mangga dan melengkapi data klasifikasi
M. laurina dan kerabatnya yang telah disusun sebelumnya.
BAHAN DAN METODE
Bahan
(M. indica) dan ‘Kiyal’ (M. indica)dari Jawa Timur, ‘Dodol Ternate’ (M. laurina)
dari Maluku Utara, dan ‘Hiku’ (M. laurina) dari Sulawesi. M. lalijiwa tidak dilakukan analisis karena panjangnya sekuen data lebih pendek 100 basa dari jenis lain, sehingga menyulitkan dalam perunutan (aligment). Semua spesimen contoh yang digunakan disimpan di Herbarium Bogoriense. Digunakan 11 grup luar (outgrup) anggota famili Rutaceae yaitu; Chisocheton macrophyllus, Guarea macrophylla subsp, Chisocheton divergens, Chisocheton tomentosus, Citrus aurantium, Citrus limon, Guarea guidonia, Guarea glabra, Citrus sinensis, Murraya paniculada, dan Citrus medica.
Metode
Isolasi DNA
Ekstraksi DNA mengikuti prosedur CTAB (Doyle & Doyle 1987) dengan beberapa modifikasi. Sekuen intergenic spacer trnL-F diamplifikasi dengan pasangan primer E dengan urutan basa (GGTTCAAGTCCCTCTATCCC) dan F (ATTTGAACTGGTGACACGAG) (Small et al. 2005). Amplifikasi daerah DNA
trnL-F intergenic spacer menggunakan mesin PCR (GeneAmp PCR sistem 2400 Perkin Elmer), sebanyak 35 siklus setelah pra PCR selama 4 menit 950C. Setiap siklus terdiri atas 940C selama 30 detik untuk denaturasi, 520C 30 detik annealing, dan 720C 1 menit untuk ekstensi dan selanjutnya diakhiri post PCR 720C 7 menit.
Sekuensing DNA analisis Filogenetik
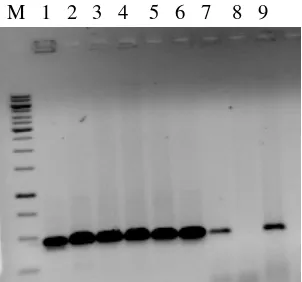
DNA produk PCR diperiksa dalam agarose 1.2% (Gambar 5) setelah itu dipurifikasi, kemudian disekuen dalam reaksi 10µl menggunakan primer trnL-F
bootstrap diulang 1000 kali. Kladogram dihasilkan dari analisis Neighbour Joining (Saitou & Nei 1987) menggunakan program Phylip 3.67.
HASIL DAN PEMBAHASAN
Sebanyak 433 pasang basa (bp) hasil sekuen trnL-F intergenic spacer
yang dihasilkan dari 6 aksesi mangga dan 11 grup luar (outgroup). Semua karakter diberi bobot yang sama, terdapat 357 karakter konstan, 7 karakter parsimoni tidak informatif, dan 69 karakter parsimoni informatif. Khusus untuk mangga, rata-rata komposisi basa nukleotidanya adalah: A (0.3014), T (0.3316), C (0.206) dan G (0.1608). Nilai tengah kandungan G+C adalah 0.3668 yang menunjukkan bahwa sekuen daerah antara (spacer) merupakan daerah yang kaya nukleotida AT. DNA hasil amplifikasi dengan cpDNA disajikan pada Gambar 5.
Gambar 5. DNA produk PCR yang diperiksa dalam agarose 1,2%
Ket.: M= marker1kb, 1=Hiku(M. laurinaHK), 2=Depeh(M. aplanata), 3=Golek(M. indicaG), 4=Betoel(M. laurinaB), 5=Madu(M. lalijiwaM), 6=Dodol ternate(M. laurinaD), 7=Kiyal(M.indicaKY), 8=Lalijiwa(M. lalijiwaL), dan 9=Kates(M. indicaK)
Perunutan berulang (Multiple alignment) dilakukan untuk menentukan nilai kesamaan dan tingkat homologinya. Perunutan sekuen nukleotida cpDNA
trnL-F pada semua aksesi mangga menunjukkan homologi yang sangat tinggi (99%). Nilai ini jauh lebih tinggi dibanding tingkat homologi 14 spesies famili Anacardiaceae yaitu 75% pada daerah ITS-1 genom inti (Hidayat & Pancoro 2001). Tingginya homologi sekuen trnL-F intergenic spacer pada M. laurina dan kerabat dekatnya disebabkan karena dekatnya kekerabatan aksesi mangga yang
diamplifikasi dan cpDNA yang lebih konservatif dibandingkan DNA inti dan diwariskan secara uniparental. Namun demikian, perubahan dapat digunakan untuk memperkirakan pola homologi fragmen cpDNA (Raubeson & Jansen 2005). Perubahan ini dapat juga digunakan untuk merujuk hubungan kekerabatan antara keturunan dan mendukung tipe dan pola proses mutasi yang mempengaruhi gen dalam genom kloroplas. Meskipun cpDNA umumnya konservatif, diversitas cpDNA telah dilaporkan terjadi pada spesies tanaman yang berbeda seperti
Fagopyrum cymosum, Astragalus sp, Conifers dan jenis yang berbeda pada
Dipterocarpaceae (Yamane et al. 2003, Liston 2008, Tsumura et al. 1996). Variasi yang terjadi pada cpDNA biasanya disebabkan oleh mutasi nukleotida tunggal yang merepresentasikan mutasi yang terjadi dalam jangka waktu lama pada waktu silam. Kecepatan mutasi lokus cpDNA antara 3.2 x 10-5 dan 7.9 x 10-5 (Provan et at. 1999). Perubahan basa pada cpDNA meskipun dalam jumlah yang sangat kecil dibanding genom inti, tetap bermakna penting dalam menyediakan sejumlah informasi untuk menjelaskan proses evolusi.
Gap terjadi karena adanya insersi dan delesi (Baldwin 1993). Pada grup dalam (ingroup) terjadi delesi pada basa no. 2 dan 59 pada aksesi ’Hiku’, insersi terjadi pada basa ke-5 dan 431 (A → T). Pada aksesi ‘Depeh’ (M. aplanata), insersi terjadi pada basa no.421 (C→ G) dan ke-431 (A → T). Pada aksesi ‘Golek’ (M. indica), insersi terjadi pada basa ke-431 (A → T). Insersi dan delesi (indel) merupakan kode untuk merujuk posisi homologi, selangnya berkisar antara 1-19 basa. Perubahan sekuen cpDNA pada tingkat taksa yang lebih rendah seperti spesies dan intra spesies terjadi dengan laju sangat rendah (<1%). Perubahan pada level nukleotida ini dapat digunakan untuk merekonstruksi pohon filogeni. Terjadinya indel pada ‘Hiku’ (M. Laurina). mendukung pembentukan percabangan pada grup dalam.
Konfirmasi identitas sekuen trnL-F intergenic spacer dilakukan dengan BLAST di GenBank. Berdasarkan pencarian BLAST, diidentifikasi bahwa daerah sekuen cpDNA trnL-F M. laurina dan kerabatnya diturunkan dari sekuen
menunjukkan kemungkinan bahwa sekuen basa trnL-FM. laurina dan kerabatnya diturunkan dari cpDNA trnL-F moyang umum (common ancestor) Anacardiaceae. Rekonstruksi pohon filogeni berdasarkan daerah trnL-F menggunakan PAUP maksimum parsimoni menghasilkan pohon yang disajikan pada Gambar 6. Nilai konsistensi indek (CI) 0.9625 atau apomorfi ciri sebesar 96.25%, nilai retensi indeks (RI) 0.99, sedangkan homoplasi indeks (HI) sebesar 0,0375. Nilai ini menunjukkan bahwa homoplasi terjadi hanya 3.75%. Konfirmasi pengelompokan
M. laurina dan kerabatnya dilakukan dengan BLAST terhadap 11 anggota famili
Rutaceae dan dijadikan sebagai outgrup. Dibanding grup luar (outgroup), ukuran daerah trnL-F mangga menunjukkan kesesuaian dengan famili Rutaceae. Enam taksa grup dalam (ingroup) membentuk kelompok yang monofiletik, terpisah dari 11 taksa group luar dari famili Rutaceae yang digunakan dalam analisis filogeni.
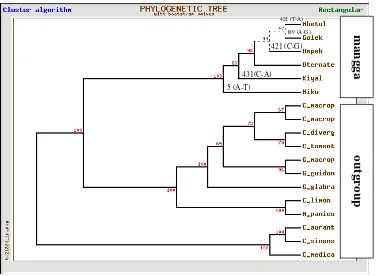
Pohon filogeni hasil analisis parsimoni (PAUP) membentuk tiga cabang. Cabang pertama ditempati oleh M. laurina dan kerabatnya yang mengelompok terpisah dari grup luar dengan nilai bootstrap 100% dan merupakan kelompok yang monofiletik atau berasal dari moyang yang sama. Kelompok ini mangga ‘Hiku’ (M. laurinaHK) yang berasal dari Sulawesi Tenggara berada pada pangkal percabangan dan berpisah dari anggota kelompok lainnya dengan perubahan basa Adenin menjadi Timin pada posisi basa ke lima, sedangkan mangga ‘Betoel’ (M.
Gambar 6. Kladogram 6 aksesi mangga dan grup luarnya berdasarkan penanda
trnL-F
Berdasarkan analisis Neighbour Joining (Saitou & Nei 1987) pada Gambar 7, Hiku (M. laurinaHK) mempunyai ruas (node) terpanjang dan muncul lebih awal dibanding kerabatnya, sehingga ‘Hiku’ diduga sebagai tetua bersama dari M. laurina dan kerabatnya. Panjang tangkai menggambarkan jarak sekuen dan kosiderasi umur molekuler (molecular clock). Dengan demikian, mangga ‘Hiku’ merupakan aksesi dengan umur molekuler yang lebih kuno yang tumbuh liar di Sulawesi Tenggara. Morfologi buah mangga ‘Hiku’ mirip dengan M. indica tetapi memiliki rasa sangat asam, berserat kasar, daging buah berwarna kuning muda. Gambaran morfologi mangga ‘Hiku’ ini lebih primitif dibanding mangga lainnya. Pola pengelompokan M. laurina dan kerabatnya berdasarkan penanda morfologi, E-RAPD, dan kombinasinya memperlihatkan pengelompokan yang berbeda dengan penanda trnL-F. Mangga Hiku berada dalam kelompok M. laurina. Fenomena cpDNA tidak harus berhubungan dengan morfologi (misalnya bentuk buah) yang sama, dan demikian sebaliknya. Pola keragaman yang ditunjukkan oleh penanda kloroplas dapat berbeda dengan pola keragaman yang
m
a
n
g
ga
ou
tg
r
ou
p
5 (A-T) 431(C-A)
421 (C-G)
ditunjukkan oleh penanda morfologi. Kloroplas diwariskan hanya dari tetua betina, sedangkan morfologi selain diwariskan kedua tetua juga dipengaruhi lingkungan.
Gambar 7. Kladogram 6 aksesi mangga berdasarkan penanda trnL-F dengan metode Neighbour Joining (Saitou & Nei 1987)
Analisis penanda cpDNA trnL-F intergenic spacer terhadap enam aksesi mangga ‘Betoel’ (M. laurinaB), ‘Depeh’ (M. aplanata), ‘Golek’ (M. indicaG), ‘Hiku’ (M. laurinaHK), ‘Dodol Ternate’ (M. laurinaD), ‘Kiyal’ (M. indicaKY), tidak sejalan dengan pengelompokan berdasarkan penanda morfologi seperti yang dikemukakan oleh Kostermans & Bompard (1993).
SIMPULAN
Analisis filogeni berdasarkan cpDNA trnL-F intergenic spacer
(Diversity of Indonesian mango cultivars based on morphological and RAPD markers)
ABSTRACT
Identification and characterization of germplasm is an important link between the conservation and utilization of plant genetic resources. In the present research, we identified and characterized genetical and phenotypical variation within 76 cultivars from Cukurgondang-Pasuruan East Java. We used RAPD marker for obtaining genetical characters of the accessions. Morphological clustering analysis grouped the cultivars into three major group. The first is ‘Kidang Kencono’. The second group consist of cultivars of Kopek and Carang and the third group was devided into 7 sub-grup of cultivars i.e. Arumanis, Madu, Gedong, Kepodang, Bapang, Golek, and Berem. The coefficient of genetic similarity within the cultivars by using morphological characters are 0.38-0.85. The genetic markers with 25 (10-mer) random primers of the 25 primer screened, 8 not reproducibile, and 10 not amplified. Only 8 primers gave reproducible polymorphic DNA amplification pattern. The number of bands generated was primer and genotype dependent and ranged from 1-13. A total of 65 distinct DNA fragments ranging from 0.20–2.0 Kb were amplified by using seven selected primers. The coefficient of genetic similarity within mango cultivars was 0.69-0.98. The cultivar Santok has the smallest similarity compared to the other cultivars, whereas the cultivars Jenis Baru and Ndok181 have the highest similarity coefficient. There is 2-31% range of genetic variability in 82 accession mango cultivars from Cukur-Gondang germplasm. The cluster analysis indicated that the eighty three cultivars of mangoes formed nine major clusters with 75% similarity coefficient. The first major cluster had nine accessions. Meanwhile, 76 cultivars from combined markers had nineteen major clusters with 0.69-0.88 of similarity coefficient. According to this result, both molecular and morpho-agronomical data sets were not equally effective to quantify and organize the genetic diversity of mango cultivars. It is better to use both markers separately. This investigation will help breeders of mangoes for the improvement program.
Keyword: Mango cultivars, identification, characterization, germplasm, morphology, RAPD
PENDAHULUAN
sebagai rujukan bagi pemulia, petani, pengusaha terkait, badan sertifikasi, dan hak kekayaan intelektual (HAKI). HAKI dan perjanjian perdagangan (Anand 2000), melindungi kultivar mangga Indonesia dari pembajakan dan menjamin keaslian tanaman yang diperjualbelikan (true to type). Pemilihan tetua berdasarkan jarak genetik antar kultivar merupakan hal penting dalam program pemuliaan tanaman. Selain itu, pemahaman pola variasi genetik infraspesies sangat penting dalam pengelolaan sumberdaya genetik dan konservasi.
Di sisi lain, keanekaragaman kultivar mangga Indonesia terancam punah seiring dengan musnahnya hutan sebagai habitat alaminya. Dalam waktu kurang dari seperempat abad diduga telah hilang puluhan sampai ratusan mangga liar yang belum dieksplorasi dan diidentifikasi. Identifikasi, karakterisasi, dan evaluasi kultivar mangga baik budidaya maupun kerabat dekatnya belum tuntas dilakukan, terutama untuk kultivar mangga di luar Pulau Jawa. Sebagian kultivar mangga di Jawa telah diidentifikasi berdasarkan ciri morfologi oleh Kusumo et al. (1975) dan Efendy et al. (2003).
Pertautan ciri antar kultivar mangga dan besarnya plastisitas ciri morfologi, cukup menyulitkan dalam membuat batasan kultivar yang ada, sehingga perlu didukung oleh sumber data dengan pendekatan lain. Pemanfaatan ciri molekuler dapat digunakan untuk mengidentifikasi kultivar dan menduga kekerabatan antar kultivar, sehingga variasi genotipe antar kultivar dapat dibedakan dengan jelas dan dapat menghindari adanya duplikasi aksesi.
Penelitian ini bertujuan untuk mengklasifikasi kultivar mangga Indonesia dengan penanda morfologi dan RAPD guna menyediakan sistem rujukan yang efektif bagi pengelompokan kultivar mangga, dan dapat menghindari duplikasi aksesi serta menjamin kemurnian kultivar, menyediakan informasi yang akurat tentang keanekaragaman genetik mangga dalam rangka pengelolaan dan konservasi plasma nutfah mangga, menyediakan informasi kemiripan kultivar yang dapat digunakan untuk membantu menyeleksi tetua dalam program pemuliaan tanaman mangga.
BAHAN DAN METODE
Bahan
Bahan penelitian berasal dari Jawa Barat, Jawa Tengah, Daerah Istimewa Yogyakarta, dan Jawa Timur yang terdapat di Kebun Koleksi Cukur Gondang Pasuruan Jawa Timur meliputi 82 kultivar mangga dianalisis RAPD, dan hanya 76 kultivar mangga yang berhasil dianalisis morfologinya (Tabel 2). Kultivar yang tidak dianalisis morfologinya adalah ‘Gandewo25’, ‘Kapuk Randu’, Glembo 361’, ‘Gambir 263’, ‘Madu Lumut’, dan ‘Slendro’.
Metode
buah, kadar air buah, rasa dan aroma buah, tebal kulit buah, bintik pada buah, dan lilin kulit.
Tabel 2. Delapan puluh dua kultivar mangga asal kebun koleksi KP
Cukurgondang-Pasuruan Jawa Timur yang dijadikan bahan penelitian
No. Nama kultivar Asal No. Nama kultivar Asal
13 ‘Dodol Wirosongko’ Cirebon Jabar 54 ‘Lalijiwo 91’ Semarang Jateng
14 ‘Gandariya ‘ Cirebon Jabar 55 ‘Kopek Mundu 329’ Cirebon Jabar
15 ‘Manila 337’ Pohjontrek Jatim 56 ‘Kecik 47’ Probolinggo Jatim
16 ‘Beku 279’ Jati Roto Jateng 57 ‘Carang’ Cirebon Jabar
32 ‘Pandan147’ Probolinggo Jatim 73 ‘Randu 411’ Pasuruan Jatim
33 ‘Canting 137’ Probolinggo Jatim 74 ‘Soho 199’ Pasuruan Jatim
34 ‘Delima 209’ Pasuruan Jatim 75 ‘Janis 17’ Bangil Jatim
acak 10 basa (OPA 14, OPA 16, OPA 17, OPA 18, SBH 12, SBH 13, SBH 14, dan SBH 19) mengikuti metode William et al. (1990). Fragmen DNA hasil amplifikasi dielektroforesis bersama DNA standar 1 KB DNA ladder (Promega) pada gel agarose 1.2% dalam larutan penyangga TBE 1X. Elektroforesis dilakukan selama 150 menit pada tegangan 60 volt, suhu ruang. Pencirian dilakukan berdasarkan pengamatan pita DNA pada setiap aksesi. Setiap pita merupakan fragmen DNA yang diukur berdasarkan 1 KB ladder.
Pengelompokan disusun berdasarkan ciri morfologi dan fragmen DNA yang dapat diamplifikasi pada tiap aksesi. Sinonim, homonim dan hubungan kekerabatan antar aksesi diketahui dari analisis similaritas menggunakan program NTSys PC versi 2.02. Kemiripan morfologi dan genetika mangga berdasarkan penanda morfologi dan RAPD dianalisis menggunakan pengelompokan (SAHN clustering) dengan metode Unweighted Pair Group Method with Arithmatic Average (UPGMA) dan hasil pengelompokannya ditampilkan dalam bentuk dendrogram. Berdasarkan ciri morfologi dan ada atau tidaknya pita DNA hasil amplifikasi menggunakan 8 primer acak RAPD dibuat matriks rata-rata kemiripan. Individu-individu yang memiliki kemiripan genetika berdekatan akan mengelompok bersama-sama atau berdekatan. Makin besar ketidakmiripannya, maka pengelompokannya akan makin jauh. Pola pengelompokan individu berdasarkan matriks kemiripan genetika tercermin dalam bentuk dendrogram, dengan jarak kemiripan genetik 0.00 (0%) sampai dengan 1.00 (100%).
HASIL DAN PEMBAHASAN
Pengelompokan dan Keanekaragaman Kultivar Mangga Berdasarkan Penanda Morfologi
Keanekaragaman tanaman mangga dapat diidentifikasi menggunakan ciri morfologi. Penanda ini banyak digunakan dalam diterminasi dan identifikasi keanekaragaman tanaman karena mudah dilakukan dan dapat diamati secara langsung.
yang tinggi pada bentuk buah, warna kulit masak, warna daging masak, aroma, ukuran buah. Mangga ‘Golek’, ‘Kepodang’ dan ‘Bapang’, dicirikan dengan bentuk buahnya memanjang. Mangga ‘Madu’, ‘Kebo’, ‘Gedong’ mempunyai bentuk buah membulat, mangga ‘Arumanis’, ‘Kopyor’, ‘Gendruk’, ‘Delima’ bentuk buahnya bulat memanjang. Warna kulit masak kuning jingga tua sampai merah pada pangkal buah dimiliki oleh mangga ‘Kepodang’, ‘Delima’, ‘Sengir’, ‘Madu Nangka’, ‘Mangkok’, ‘Urang’ dan ‘Beruk’, sedangkan warna kulit buah masak hijau sampai hijau kekuningan pada bagian pangkal buah dimiliki oleh mangga ‘Berem’, ‘Golek’ dan ‘Madu’. Mangga ‘Thaber’, ‘Gurih Panjang’, ‘Manalagi Probolinggo’ memiliki warna masak hijau tua. Warna daging buah masak kuning muda dimiliki oleh kultivar ‘Berem’, ‘Madu’ dan ‘Nanas’. Mangga ‘Arumanis’, ‘Beruk’, ‘Gedong’ memiliki daging buah kuning sampai jingga tua.
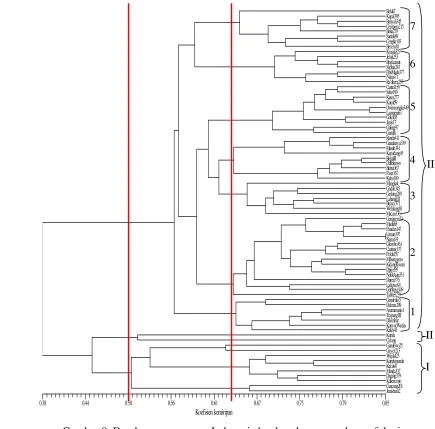
Hasil analisis pengelompokan yang diturunkan dari matriks kemiripan morfologi tanaman mangga tidak memberikan pengelompokan berdasarkan daerah asal ataupun bentuk buah seperti yang dikenali selama ini. Pengelompokan dibentuk berdasarkan kemiripan 92 ciri morfologi tertentu. Seluruh individu menjadi satu kelompok pada kemiripan morfologi 38%. Pengelompokan berdasarkan ciri morfologi menunjukkan bahwa 76 kultivar mangga berada pada kisaran kemiripan 0.38-0.85. Kultivar ‘Limun197’ dan ‘Nanas93’ dengan nilai kemiripan tertinggi (0.85). Kultivar ‘Lalijiwo’ dan kultivar ‘Trapang’ memiliki nilai kemiripan paling rendah (0.38) kedua kultivar ini merupakan jenis berbeda.
Kelompok utama II terdiri 2 kultivar yaitu ‘Kopek’ dan ‘Carang’, kelompok ini disatukan oleh persamaan ciri bentuk daun, tonjolan areola, lebar perbungaan, rambut pada perbungaan, posisi rambut pada organ bunga, lebar sepal, ujung petal, arah lipatan petal, posisi ovari, ukuran dan warna kulit buah.
Gambar 8. Dendrogram mangga Indonesia berdasarkan penanda morfologi
Kelompok utama III merupakan kelompok terbesar yang terdiri dari 64 kultivar. Pada nilai kemiripan 61% terbentuk 7 kelompok kultivar. Penamaan kelompok kultivar mangga didasarkan pada kultivar-kultivar yang umum dikenal oleh petani dan konsumen dengan ciri-ciri yang menonjol untuk mewakili
Koefisien kemiripan
0.38 0.44 0.50 0.56 0.61 0.67 0.73 0.79 0.85