KARAKTERISTIK MORFOLOGI DAN FISIOLOGI BIBIT
RUMPUT LAUT
Kappaphycus alvarezii
HASIL SELEKSI
MASSA DI PERAIRAN GORONTALO
SITI FADILAH
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul “Karakteristik Morfologi
dan Fisiologi Bibit Rumput Laut Kappaphycus alvarezii Hasil Seleksi Massa di
Perairan Gorontalo” adalah benar karya saya dengan arahan dari komisi
pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan dan tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
SITI FADILAH. Karakteristik Morfologi dan Fisiologi Bibit Rumput Laut Kappaphycus alvarezii Hasil Seleksi Massa di Perairan Gorontalo. Dibimbing oleh ALIMUDDIN dan JOKO SANTOSO.
Rumput laut Kappaphycus alvarezii (Doty) merupakan salah satu jenis rumput laut komoditas unggulan. Penggunaan bibit yang baik dapat mendukung keberhasilan usaha budidaya rumput laut ini. Bibit rumput laut umumnya didapatkan dari pengembangbiakan vegetatif rumput laut hasil panen, sehingga kualitas bibit menurun seiring waktu. Untuk itu dibutuhkan upaya pengadaan bibit secara sistematis. Penggunaan metode seleksi massa untuk menghasilkan kualitas bibit K. alvarezii yang lebih baik telah dirintis, namun masih terbatas pengamatan pada hasil produksi bibit dan kandungan karaginan sebagai unsur penting dalam menentukan potensi hasil pada budidaya rumput laut. Potensi hasil tinggi dapat diamati pada karakteristik morfologi dan fisiologinya. Penelitian ini bertujuan untuk mengevaluasi karakteristik morfologi dan fisiologi bibit rumput laut Kappaphycus alvarezii hasil seleksi massa di perairan Gorontalo serta mempelajari hubungan antara karakteristik morfologi dan fisiologi.
Kegiatan seleksi massa dilakukan di Teluk Tomini (pesisir selatan Gorontalo) berdasarkan protokol seleksi massa bibit unggul rumput laut dengan modifikasi pada nilai cut-off menjadi 10% laju pertumbuhan harian tertinggi. Pemeliharaan bibit dilakukan menggunakan metode tali panjang (long line). Konstruksi pemeliharaan berukuran luas 50 x 35 m2, memuat 50 tali bentangan dengan panjang 35 m, dan jarak antar tali bentangan 1 m. Setiap tali bentangan memuat 230 titik rumpun bibit dengan jarak antar tali pengikat rumpun 15 cm. Bibit rumput laut berwarna coklat dengan bobot awal 50 g diikat pada tali rumpun. Penanaman bibit dilakukan pada kedalaman 30 cm dari permukaan perairan. Selanjutnya, rumput laut dipelihara selama 30 hari per siklus. Setiap siklus pemeliharaan terdapat kontrol internal dan kontrol eksternal. Seleksi massa ini dilakukan sampai generasi keempat. Bibit rumput laut hasil seleksi tersebut selanjutnya diproduksi massal untuk dilakukan evaluasi performa budidaya. Produksi massal dan evaluasi performa budidaya dilakukan di Laut Sulawesi (pesisir utara Gorontalo) selama tiga siklus. Metode pemeliharaan pada kegiatan produksi massal sama dengan kegiatan seleksi massa bibit. Analisis karakteristik morfologi dan fisiologi dilakukan pada setiap akhir siklus produksi massal. Karakteristik morfologi yang diamati adalah panjang talus utama, cabang I, II dan III; jarak ruas primer dan sekunder; diameter primer, sekunder dan tersier; jumlah cabang I, II dan III. Karakteristik fisiologi yang diukur adalah laju pertumbuhan harian, kadar gula total, kadar hormon kinetin dan indole acetic acid (IAA). Pengamatan kualitas perairan lokasi produksi massal secara in situ dilakukan pada variabel suhu, kadar oksigen terlarut (DO), dan salinitas. Pengamatan secara ex situ dilakukan pada variabel NO3 (nitrat), PO4 (fosfat) dan bahan organik total
(BOT). Data sekunder berupa curah hujan didapatkan dari Stasiun Meteorologi Gorontalo.
134,07% dibandingkan dengan kontrol eksternal dan 69,89% dibandingkan kontrol internal. Bibit hasil seleksi dan kontrol internal mempunyai jarak ruas primer dan sekunder yang lebih pendek daripada kontrol eksternal. Diameter talus primer, sekunder dan tersier pada ketiga jenis bibit rumput laut tidak menunjukkan perbedaan. Bibit kontrol internal, kontrol eksternal dan hasil seleksi mempunyai laju pertumbuhan harian di bawah 1%, namun rumput laut hasil seleksi mempunyai laju pertumbuhan harian yang lebih baik dibandingkan kedua kontrol. Kadar gula total pada bibit hasil seleksi adalah 15,52% lebih tinggi daripada kontrol internal dan 16,42% lebih tinggi daripada kontrol eksternal. Kadar hormon kinetin dan rasio kinetin:IAA cenderung lebih tinggi pada rumput laut hasil seleksi dibandingkan dengan kedua kontrol. Kadar hormon kinetin mempunyai korelasi tinggi dengan panjang total cabang, jumlah total cabang dan LPH daripada IAA. Warna talus berbanding lurus dengan LPH, kadar IAA, dan kadar kinetin. Kondisi lingkungan perairan saat produksi massal dari bulan Agustus sampai Oktober kurang produktif di perairan Gorontalo Utara. Kualitas lingkungan menurun dan biofouling berkembang pada periode tersebut.
SUMMARY
SITI FADILAH. Morphological and Physiological Characteristics of Seed Kappaphycus alvarezii Produced by Mass Selection in Gorontalo Waters. Supervised by ALIMUDDIN and JOKO SANTOSO.
Seaweed Kappaphycus alvarezii (Doty) is one of the leading commodity seaweed. The use of high quality seed can support the success of this seaweed cultivation. Seaweed seeds are generally obtained by some of the crop through vegetative propagation, so the quality of the seed decreases over time. Therefore, a systematic attempt to generate a high quality seed is necessary. The use of mass selection method to produce a high quality of K. alvarezii seed has been initiated, but it still limited observations on seed production and carrageenan content as an important element in determining the yield potential of seaweed farming. The high yield potential can be observed in morphological and physiological characteristics. This study aimed to evaluate the morphological and physiological characteristics of K. alvarezii seed seaweed produced by mass selection in Gorontalo waters and to study correlation between morphological and physiological characteristics.
Mass selection was carried out in Tomini Gulf (south coast of Gorontalo) based on seaweed mass selection protocol with slight modification in cut-off to 10% of the highest daily growth rate (DGR). Cultivation was performed for 30 days of each cycle, using long-line method. Cultivation construction was 50 x 35 m2, containing 50 long ropes and each rope was 35 m in length, and the distance between the ropes were 1 m. Each rope contained 230 seed clumps with distance between clumps were 15 cm. Seed seaweed with initial clump weight of 50 g was tied to the rope. The seeds were placed at 30 cm from water surface. Furthermore seaweed was maintained for 30 days per cycle. In each cultivation cycle, there were internal control and external control. Selection was carried out until 4th generation. Mass production and cultivation performance test were conducted in the Celebes Sea (north coast of Gorontalo) for three cycles of cultivation. Cultivation method was performed by the same procedures as applied in mass selection. Analysis of morphological and physiological characteristics were performed at the end of each mass production cycle. Morphological characteristics measured were thallus length (the main thallus, branches I, II and III), internodes (primary and secondary), thalli diameter (primary, secondary and tertiary), branches number (I, II and III). Physiological characteristics observed were DGR, total sugar content, and hormone level of kinetin and indole acetic acid (IAA). Waters qualities measured were in situ observation of temperature, dissolved oxygen, and salinity, while ex situ observation were NO3 (nitrate), PO4
(phosphate) and total organic matter (TOM). Rainfall as secondary data was obtained from the Gorontalo Meteorological Station.
seed showed no difference. Seeds of internal control, external control and selected had daily growth rate under 1%, but the selected seed had DGR better than both controls. Total sugar content of the selected seed increased 15.52% than internal control and 16.42% than external control. Hormone levels of kinetin and kinetin-IAA ratios tend to be higher in selected seed than both controls. Hormone kinetin level higher correlated with the total length of the branch, the total number of branches and DGR than IAA. Thalli color showed high correlation to DGR, levels of IAA and kinetin. Waters environmental conditions in August to October were less productive in the North Gorontalo waters. Environmental quality was declined and biofouling attached the seaweed in that period.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Akuakultur
KARAKTERISTIK MORFOLOGI DAN FISIOLOGI BIBIT
RUMPUT LAUT
Kappaphycus alvarezii
HASIL SELEKSI
MASSA DI PERAIRAN GORONTALO
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
Judul Tesis : Karakteristik Morfologi dan Fisiologi Bibit Rumput Laut
Kappaphycus alvarezii Hasil Seleksi Massa di Perairan Gorontalo Nama : Siti Fadilah
NIM : C151120581
Disetujui oleh Komisi Pembimbing
Dr Alimuddin, SPi MSc Ketua
Dr Ir Joko Santoso, MSi Anggota
Diketahui oleh
Ketua Program Studi Ilmu Akuakultur
Dr Ir Widanarni, MSi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
Tanggal Ujian: 25 Agustus 2014
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian ini adalah seleksi massa rumput laut, dengan judul Karakteristik Morfologi dan Fisiologi Bibit Rumput Laut Kappaphycus alvarezii Hasil Seleksi Massa di Perairan Gorontalo.
Terima kasih penulis ucapkan kepada :
1. Komisi pembimbing Bapak Dr Alimuddin, SPi MSc dan Bapak Dr Ir Joko Santoso atas bimbingan dan motivasi yang telah diberikan kepada penulis 2. Ibu Dr Ir Dinar Tri Soelistyowati, DEA selaku penguji luar komisi dan Bapak
Dr Ir Dedi Jusadi, MSc selaku penguji dari program studi atas saran dan masukan yang diberikan untuk tesis ini
3. Ibu Dr Ir Widanarni, MSi selaku ketua program studi
4. Kementerian Kelautan dan Perikanan yang telah memberikan beasiswa Program Pascasarjana
5. Bapak Dr Andi Parenrengi selaku Kepala Balai Penelitian dan Pengembangan Budidaya Air Payau (BPPBAP) Maros dan Bapak Petrus Rani Pong-Masak, SPi MSi selaku Kepala Loka Penelitian dan Pengembangan Budidaya Rumput Laut (LPPBRL) Gorontalo atas saran dan dukungan dalam penelitian ini
6. Segenap staf peneliti dan teknisi BPPBAP dan LPPBRL yang telah memberi dukungan teknis selama penelitian
7. Staf Stasiun Meteorologi Klas III Jalaluddin, Gorontalo yang telah membantu dalam dukungan data sekunder.
8. Ungkapan terima kasih juga disampaikan kepada kedua orang tua, serta seluruh keluarga, atas segala doa dan kasih sayangnya.
9. Teman-teman Ilmu Akuakultur 2012 atas bantuan dan dukungannya.
Tesis ini diharapkan dapat memberikan informasi bagi perkembangan ilmu pengetahuan. Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 2
Manfaat Penelitian 2
2 METODE 3
Waktu dan Lokasi Penelitian 3
Materi Uji 3
Prosedur Seleksi Massa 4
Produksi Massal dan Evaluasi Performa Budidaya 4
Variabel Uji 6
Analisis Data 8
3 HASIL DAN PEMBAHASAN 8
Karakteristik Morfologi Bibit Rumput Laut 8
Karakteristik Fisiologi Bibit Rumput Laut 10
Hubungan Antara Hormon dan Pertumbuhan Bibit Hasil Seleksi 13 Hubungan Antara Warna Talus dan Karakter Fisiologi Pertumbuhan
Bibit Hasil Seleksi 14
Kondisi Lingkungan Lokasi Penelitian 14
4 SIMPULAN 18
Simpulan 18
DAFTAR PUSTAKA 19
LAMPIRAN 23
DAFTAR TABEL
1 Panjang talus, jumlah cabang, jarak ruas dan diameter talus bibit rumput laut Kappaphycus alvarezii hasil seleksi massa, kontrol internal dan
eksternal 9
2 Kualitas perairan Gorontalo Utara dan kondisi bibit Kappaphycus alvarezii pada saat evaluasi performa budidaya 15 3 Variabel morfologi dan fisiologi tiga jenis bibit rumput laut
Kappaphycus alvarezii pada tiga siklus yang berbeda 17
DAFTAR GAMBAR
1 Lokasi penelitian seleksi massa di perairan Teluk Tomini dan produksi
massal di perairan Laut Sulawesi, Gorontalo 3
2 Skema tahapan kegiatan seleksi massa bibit rumput laut Kappaphycus alvarezii sampai dengan produksi massal bibit hasil seleksi (BPPBAP 2011) dengan modifikasi pada nilai cut-off menjadi 10% laju
pertumbuhan harian tertinggi (LPHt) 5
3 Pemeliharaan rumput laut dengan metode long line (Pong-Masak et al.
2013) 6
4 Morfologi bibit rumput laut : a) talus utama, b) cabang I, c) cabang II,
d) cabang III, e) ruas primer, f) ruas sekunder 7
5 Performa bibit rumput laut Kappaphycus alvarezii; a) kontrol internal,
b) kontrol eksternal, dan c) hasil seleksi 9
6 Rataan bobot rumpun dan laju pertumbuhan harian (LPH) pada jenis bibit rumput laut Kappaphycus alvarezii yang berbeda 11 7 Kadar gula total pada jenis bibit rumput laut Kappaphycus alvarezii
yang berbeda 11
8 Kadar hormon indole acetic acid (IAA) dan hormon kinetin pada jenis bibit rumput laut Kappaphycus alvarezii yang berbeda 12 9 Rasio kinetin:IAA pada jenis bibit rumput laut Kappaphycus alvarezii
yang berbeda 13
10 Kondisi curah hujan di lokasi seleksi massa (atas) dan evaluasi performa budidaya Kappaphycus alvarezii (bawah). Garis putus-putus menunjukkan waktu penelitian. Gambar diolah dari data curah hujan Stasiun Tilamuta (atas) dan Kwandang (bawah), Stasiun Meteorologi
(BMKG) Gorontalo 15
11 Talus yang ditutupi oleh alga epifit (A) dan teritip (B) di bulan Agustus 16
DAFTAR LAMPIRAN
1 Diagram alir proses pemurnian IAA, GA3, ABA dan zeatin 23 2 Kandungan pupuk organik cair Alam Hijau dan perendaman rumput
laut dengan pupuk organik cair 24
3 Hasil analisis ragam dan uji lanjut pada variabel panjang talus utama
4 Hasil analisis ragam dan uji lanjut pada variabel panjang cabang II dan
cabang III 26
5 Hasil analisis ragam dan uji lanjut pada variabel total panjang talus dan
jumlah cabang I 27
6 Hasil analisis ragam dan uji lanjut pada variabel jumlah cabang II dan
jumlah cabang III 28
7 Hasil analisis ragam dan uji lanjut pada variabel total jumlah cabang
dan jarak ruas primer 29
8 Hasil analisis ragam dan uji lanjut pada variabel jarak ruas sekunder
dan diameter primer 30
9 Hasil analisis ragam dan uji lanjut pada variabel diameter sekunder dan
diameter tersier 31
10 Hasil analisis ragam dan uji lanjut pada variabel kandungan gula total
dan rataan bobot 32
11 Hasil analisis ragam dan uji lanjut pada variabel LPH dan rasio
kinetin:IAA 33
1
1
PENDAHULUAN
Latar Belakang
Rumput laut Kappaphycus alvarezii (Doty) merupakan salah satu jenis rumput laut dari kelas Rhodophyceae (alga merah) yang menjadi komoditas unggulan. Rumput laut ini mengandung karaginan dari fraksi kappa yang secara luas dimanfaatkan dalam bidang industri makanan, farmasi, kosmetik dan lain-lain. Kebutuhan akan produk karaginan dan bahan mentah K. alvarezii diprediksi akan terus meningkat di masa depan.
Penggunaan bibit yang baik dapat mendukung keberhasilan usaha budidaya rumput laut. Bibit rumput laut umumnya diperoleh dari sebagian hasil panen melalui pengembangbiakan secara vegetatif. Penggunaan bibit secara terus-menerus dengan cara seperti ini dapat menurunkan keragaman genetik, laju pertumbuhan, kadar karaginan dan kekuatan gel serta meningkatkan kerentanan terhadap penyakit (Hurtado & Cheney 2003). Oleh karena itu, upaya pengadaan bibit yang sistematis diperlukan.
Keberadaan kebun bibit merupakan salah satu upaya yang dapat dilakukan dalam penyediaan bibit yang berkelanjutan, khususnya produksi bibit budidaya yang siap tebar (Parenrengi et al. 2011). Bibit rumput laut unggul dapat diperoleh dengan menggunakan beberapa metode, yaitu metode seleksi massa, teknik kultur jaringan dan rekayasa genetik. Teknik kultur jaringan dan rekayasa genetik adalah strategi jangka menengah dan panjang untuk mendapatkan bibit unggul. Penggunaan teknik kultur jaringan untuk mendapatkan bibit unggul telah banyak dilakukan (Reddy et al. 2003; Kumar et al. 2004; Mulyaningrum et al. 2012; Sulistiani et al. 2012; Suryati et al. 2013). Keberhasilan aklimatisasi dan budidaya lapang dari rumput laut K. alvarezii hasil kultur embrio somatik telah dilaporkan oleh Reddy et al. (2003) di India. Rumput laut tersebut dimonitoring selama 7 generasi dan menunjukkan laju pertumbuhan harian (LPH) 1,5-1,8 kali lebih tinggi daripada rumput laut pembudidaya. Di Indonesia, keberhasilan budidaya lapang dengan menggunakan bibit Gracilaria verrucosa hasil kultur jaringan telah dilaporkan oleh Suryati et al. (2013). Selama masa produksi massal diperoleh 1.200 kg bibit dalam 5 siklus pemeliharaan. Sementara itu, penggunaan rekayasa genetik difokuskan pada aplikasi transgenesis dengan tujuan peningkatan pertumbuhan, resistensi penyakit dan daya tahan terhadap kondisi lingkungan ekstrim (Rajamuddin 2010). Namun demikian, hingga kini bibit unggul hasil rekayasa genetik belum ada.
Di sisi lain, metode seleksi massa telah banyak digunakan untuk meningkatkan hasil panen pada tumbuhan tingkat tinggi dan meningkatkan proporsi genotipe unggul dalam waktu singkat (Taran et al. 2004; Yang et al. 2013). Penggunaan metode seleksi massa untuk menghasilkan kualitas bibit K. alvarezii yang lebih baik telah dirintis oleh Pong-Masak et al. (2013).
2
laut terhadap penyakit dan stres lingkungan adalah bervariasi. Oleh karena itu, beberapa varietas dan sumber bibit awal bisa digunakan dalam program seleksi dengan penekanan pada varietas lokal yang telah teradaptasi lama di suatu kawasan. Berdasarkan hasil penelitian Pong-Masak et al. (2013) di perairan Takalar, Sulawesi Selatan, didapatkan bahwa bibit hasil seleksi massa memperlihatkan respons LPH yang lebih baik. Produksi bibit meningkat sebesar 15-25% dibandingkan dengan kontrol internal (rataan bobot dari masing-masing strain) dan kontrol eksternal (bibit yang berasal dari masyarakat pembudidaya lokal).
Penggunaan rumput laut varietas unggul sangat berperan dalam peningkatan produksi rumput laut nasional. Pada tumbuhan tingkat tinggi, seperti padi, banyak karakteristik morfologi dan fisiologi yang berperan terhadap potensi hasil tinggi. Karakteristik morfologi dan fisiologi antara varietas yang satu dengan lainnya berbeda dan dapat menjadi petunjuk kemampuan varietas tersebut (Wahyuti 2012).
Perumusan Masalah
Hasil penelitian Pong-Masak et al. (2013) masih terbatas pengamatan pada hasil produksi bibit dan kandungan karaginan sebagai unsur penting dalam menentukan potensi hasil pada budidaya rumput laut selama seleksi massa. Sementara itu, potensi hasil tinggi dapat dibedakan antara varietas yang satu dengan lainnya dengan didasarkan pada karakteristik morfologi dan fisiologinya. Pada penelitian ini dilakukan kajian yang lebih mendalam selama evaluasi performa budidaya terhadap bibit hasil seleksi massa. Kajian ini meliputi karakteristik morfologi dan fisiologi bibit. Karakteristik morfologi dievaluasi berdasarkan berbagai perubahan fenotipe, sedangkan karakteristik fisiologi adalah berdasarkan laju pertumbuhan, kadar fitohormon dan kadar gula. Selain bibit rumput laut, pengamatan juga dilakukan terhadap kondisi lingkungan sebagai pendukung pertumbuhan rumput laut.
Tujuan Penelitian
Tujuan penelitian ini ialah mengevaluasi karakteristik morfologi dan fisiologi bibit rumput laut Kappaphycus alvarezii hasil seleksi massa di perairan Gorontalo serta mempelajari hubungan antara karakteristik morfologi dan fisiologi.
Manfaat Penelitian
Hasil penelitian dapat memberikan:
1. Informasi karakteristik morfologi dan fisiologi yang berperan dalam menentukan hasil pada rumput laut hasil seleksi.
3
2 METODE
Waktu dan Lokasi Penelitian
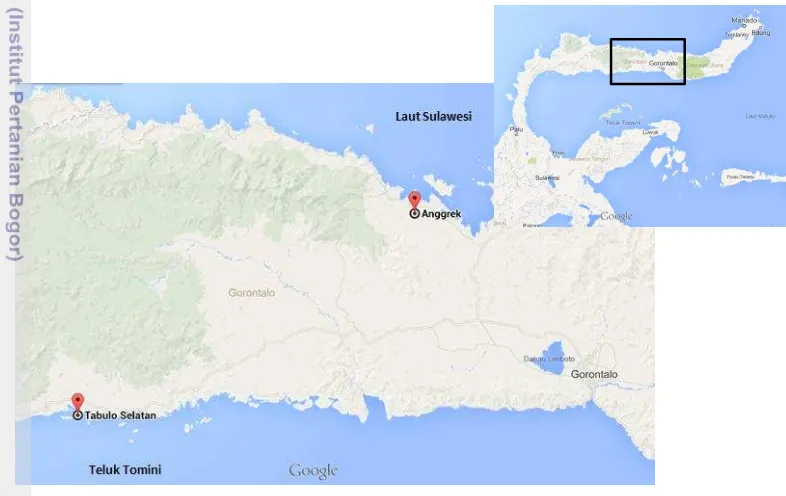
Penelitian ini dilaksanakan dari bulan Maret sampai Desember 2013. Penelitian diawali dengan kegiatan seleksi massa bibit rumput laut K. alvarezii yang dilakukan di perairan laut Teluk Tomini Loka Penelitian dan Pengembangan Budidaya Rumput Laut (LPPBRL), Desa Tabulo Selatan, Kecamatan Mananggu, Kabupaten Boalemo, Gorontalo (Gambar 1). Kegiatan seleksi massa ini dilaksanakan dari bulan Maret sampai Juli 2013. Bibit hasil seleksi selanjutnya diproduksi secara massal dan dilakukan evaluasi performa budidaya di perairan Laut Sulawesi, Kecamatan Anggrek, Kabupaten Gorontalo Utara. Kegiatan lanjutan ini dilakukan dari Agustus sampai Oktober 2013. Analisis sampel dilakukan di laboratorium sampai bulan Desember 2013.
Gambar 1 Lokasi penelitian seleksi massa di perairan Teluk Tomini dan produksi massal di perairan Laut Sulawesi, Gorontalo
Materi uji
4
dihasilkan dari seleksi massa digunakan untuk kegiatan produksi massal selama tiga siklus.
Prosedur Seleksi Massa
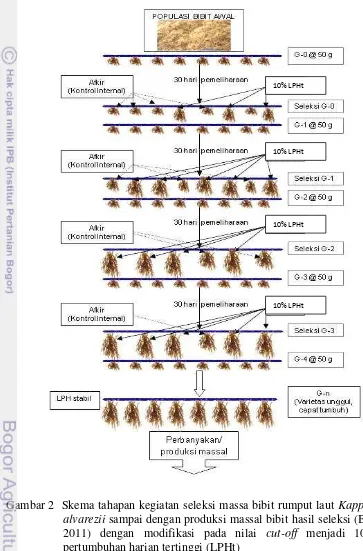
Kegiatan seleksi massa dilakukan berdasarkan protokol seleksi massa bibit unggul rumput laut (BPPBAP 2011) dengan modifikasi pada nilai cut-off seleksi menjadi 10% LPH tertinggi (Gambar 2). Menurut Pong-Masak et al. (2013), nilai cut-off seleksi 10% dari populasi bibit terbaik dalam setiap tali bentangan akan memberikan peningkatan produksi sebesar 25-32% dibandingkan dengan kontrol atau bibit yang dibudidayakan masyarakat. Pemeliharaan bibit menggunakan metode tali panjang (long line) (Gambar 3). Konstruksi pemeliharaan berukuran luas 50 x 35 m2, memuat 50 tali bentangan dengan panjang 35 m dan jarak antar tali bentangan 1 m. Setiap tali bentangan memuat 230 titik rumpun bibit dengan jarak antar tali pengikat rumpun 15 cm. Bibit rumput laut dengan bobot awal 50 g (G-0) diikat pada tali rumpun. Penanaman bibit dilakukan pada kedalaman 30 cm dari permukaan perairan. Selanjutnya rumput laut dipelihara selama 30 hari setiap siklus. Kegiatan pembersihan rumput laut dari organisme pengganggu dilakukan sesuai kebutuhan.
Bobot setiap rumpun ditimbang pada akhir pemeliharaan setiap siklus. Bibit yang memiliki LPH sampai dengan 10% nilai LPH tertinggi, dipisahkan dari populasi tali bentangan. Rumpun bibit yang terpilih ini dipotong menjadi rumpun baru (50 g) yang diikat pada tali bentangan baru, dan dipelihara dengan metode, proses dan periode yang sama dengan siklus sebelumnya. Bibit hasil seleksi dari setiap siklus disebut sebagai satu generasi. Setiap siklus pemeliharan ada kontrol internal dan kontrol eksternal. Kontrol internal adalah rumput laut yang memiliki rataan LPH populasi, sedangkan kontrol eksternal adalah bibit yang diperoleh dari pembudidaya sekitar lokasi penelitian.
Seleksi massa ini dilakukan sampai generasi keempat. Biasanya pada generasi keempat ini LPH telah stabil dan tingkat keseragaman mencapai 90%. Bibit rumput laut hasil seleksi tersebut selanjutnya digunakan untuk produksi massal dan evaluasi performa budidaya.
Produksi Massal dan Evaluasi Performa Budidaya
Produksi massal dan evaluasi performa budidaya dilakukan selama tiga siklus dengan tiap siklusnya adalah 30 hari. Cara pemeliharaan pada kegiatan produksi massal ini sama dengan kegiatan seleksi massa bibit, yaitu menggunakan metode long line. Pada siklus pertama, budidaya bibit menggunakan bibit rumput laut hasil seleksi dan kontrol internal yang dibawa dari perairan Teluk Tomini. Performa keduanya dibandingkan dengan bibit kontrol eksternal yang berasal dari lokasi produksi massal (perairan Laut Sulawesi). Bibit rumput laut hasil seleksi, kontrol internal dan kontrol eksternal yang diuji masing-masing berjumlah 3 tali bentangan. Selanjutnya bibit hasil produksi massal dari siklus pertama dibudidayakan untuk siklus kedua, kemudian bibit hasil siklus kedua dibudidayakan untuk siklus ketiga.
5
eksternal secara acak sebanyak tiga titik pada beberapa bentangan dengan metode sampling-panen. Pertumbuhan diamati dengan menimbang bobot basah rumput laut dari setiap sampel uji.
Gambar 2 Skema tahapan kegiatan seleksi massa bibit rumput laut Kappaphycus alvarezii sampai dengan produksi massal bibit hasil seleksi (BPPBAP 2011) dengan modifikasi pada nilai cut-off menjadi 10% laju pertumbuhan harian tertinggi (LPHt)
10% LPHt
10% LPHt 10% LPHt
6
Variabel Uji
Pengukuran karakteristik morfologi dan fisiologi dilakukan pada setiap akhir siklus produksi massal. Variabel uji yang diamati pada setiap rumpun rumput laut K. alvarezii adalah :
1. Karakteristik morfologi (Gambar 4) berdasarkan Meneses (1996) :
a. Panjang talus yang terdiri atas talus utama, cabang I, II dan III. Cabang I adalah talus yang tumbuh dari talus utama. Cabang II adalah talus yang tumbuh dari cabang I, sedangkan cabang III adalah talus yang tumbuh dari cabang II. Panjang talus ini diukur dari pangkal hingga ujung talus.
b. Jarak ruas yang terdiri atas ruas primer dan ruas sekunder. Ruas primer adalah jarak antara tumbuhnya cabang-cabang I sedangkan ruas sekunder adalah jarak antara tumbuhnya cabang-cabang II. Jarak ruas diukur dari tempat tumbuhnya cabang sampai ke cabang terdekat.
c. Diameter talus yang terdiri atas diameter primer, sekunder dan tersier. Diameter primer adalah diameter pada talus utama. Diameter sekunder adalah diameter cabang I, sedangkan diameter tersier adalah diameter cabang II dan III.
d. Jumlah cabang pada cabang I, II dan III.
7
2. Karakteristik fisiologi :
a. LPH. LPH diperoleh dengan mengukur bobot basah bibit rumput laut setiap siklus (30 hari). Formulasi yang digunakan yaitu :
Keterangan :
LPH = Laju Pertumbuhan Harian (% /hari) Wt = Bobot pada waktu t (g)
W0 = Bobot pada waktu t = 0 (g) t = Jumlah hari pengamatan (hari)
b. Kadar gula total. Kadar gula total dianalisis secara titrimetri dengan menggunakan metode Luff Schoorl (AOAC 1995) di Laboratorium Terpadu, Institut Pertanian Bogor.
c. Kadar hormon kinetin dan indole acetic acid (IAA). Kadar hormon kinetin dan IAA dianalisis secara spektrofotometri (Unyayar et al. 1996; Lampiran 1) di Laboratorium Biologi, Institut Pertanian Bogor.
3. Pengamatan kualitas perairan di lokasi produksi massal dilakukan secara in situ dan ex situ. Pengamatan secara in situ dilakukan pada sekitar pukul 10.00 WITA. Variabel suhu dan DO diukur dengan menggunakan alat DOmeter, sedangkan salinitas diukur dengan menggunakan refraktometer. Pengamatan secara ex situ dilakukan pada variabel NO3 (nitrat), PO4 (fosfat) dan bahan
organik total (BOT) secara spektrofotometri. Analisis ini dilakukan di Laboratorium Kualitas Air, Balai Penelitian dan Pengembangan Budidaya Air Payau (BPPBAP), Maros, Sulawesi Selatan. Data sekunder berupa curah hujan didapatkan dari Stasiun Meteorologi Klas III Jalaluddin Gorontalo. Data curah hujan untuk kegiatan seleksi massa diperoleh dari stasiun Tilamuta, sedangkan untuk kegiatan produksi massal diperoleh dari stasiun Kwandang.
8
Analisis Data
Data karakteristik morfologi dan fisiologi dianalisis dengan analisis ragam (ANOVA) pada taraf uji 5% menggunakan program SPSS 17. Jika ada pengaruh nyata maka analisis dilanjutkan dengan menggunakan uji jarak berganda Duncan. Analisis korelasi Pearson digunakan untuk mengetahui hubungan antar variabel. Data kualitas air dan curah hujan dianalisis secara deskriptif dalam bentuk grafik.
3 HASIL DAN PEMBAHASAN
Karakteristik morfologis bibit rumput laut
Pertumbuhan dan perkembangan adalah proses berkelanjutan yang mengarah pada karakteristik morfologis dari suatu spesies. Karakteristik morfologis menjadi penciri untuk melihat perbedaan kondisi pertumbuhan dari tiga jenis bibit rumput laut yang diteliti. Dalam penelitian ini, karakteristik morfologis tersebut mencakup ciri-ciri yang dapat diamati visual secara langsung, yaitu panjang talus, jumlah cabang, jarak ruas dan diameter talus. Secara umum berbentuk silindris dengan permukaan licin dan bertekstur plastis.
Hasil penelitian menunjukkan bahwa panjang total talus rumput laut hasil seleksi adalah 66,94% lebih panjang daripada kontrol eksternal dan 48,35% lebih panjang daripada kontrol internal (p<0,05; Tabel 1). Panjang total talus merupakan jumlah keseluruhan dari panjang talus utama dan seluruh panjang cabang-cabangnya. Pertumbuhan total panjang talus rumput laut hasil seleksi ini didapatkan dari pertambahan nyata panjang talus utama, cabang II dan III (p<0,05) dibandingkan dengan kedua kontrol. Pada kontrol eksternal, pertumbuhan talus lebih didominasi oleh panjang talus utama, sedangkan pertumbuhan talus kontrol internal didominasi oleh panjang cabang III (p<0,05).
Jumlah total cabang pada rumput laut hasil seleksi (316 cabang) meningkat 134,07% dibandingkan dengan kontrol eksternal (135 cabang), dan 69,89% dibandingkan kontrol internal (186 cabang). Peningkatan jumlah total cabang rumput laut hasil seleksi ini tampak terjadi pada cabang I, II dan III, sedangkan pada kontrol internal terjadi pada cabang III (p<0,05; Tabel 1).
9
Tabel 1 Panjang talus, jumlah cabang, jarak ruas dan diameter talus bibit rumput laut Kappaphycus alvarezii hasil seleksi massa, kontrol internal dan eksternal
Gambar 5 Performa bibit rumput laut Kappaphycus alvarezii; a) kontrol internal, b) kontrol eksternal, dan c) hasil seleksi
Rumput laut hasil seleksi dan kontrol internal memperlihatkan ciri yang khas diakibatkan jarak ruasnya yang pendek. Pertumbuhan cabang-cabang talus menjadi bergerombol di sepanjang talus utama dan cabang I sehingga rumpun terlihat lebih rimbun (Gambar 5). Berbeda dengan rumput laut kontrol eksternal yang percabangannya terlihat tumbuh menyebar. Diduga hal ini disebabkan oleh kondisi lingkungan di perairan selatan Gorontalo telah memberi ciri yang khas pada rumput laut hasil seleksi dan kontrol internal selama kegiatan seleksi massa berlangsung. Sementara itu, ciri khas pada rumput laut kontrol eksternal dipengaruhi oleh lingkungan di perairan utara Gorontalo.
Variabel uji menunjukkan berbeda nyata pada taraf uji 5% (uji jarak berganda Duncan)
10
Menurut Gardner et al. (2008), pertumbuhan tumbuhan dapat dinyatakan sebagai fungsi dari faktor pertumbuhan internal (genotipe) dengan faktor pertumbuhan eksternal (lingkungan). Interaksi antara genotipe dan lingkungan memberikan penampakan (fenotipe) dari daya genetik tersebut. Tumbuhan memperoleh bentuk khasnya karena pertumbuhan bagian-bagian penyusunnya yang saling berhubungan. Bagian-bagian penyusunnya juga mempunyai bentuk yang khas yang dapat terulang menurut ruang dan waktu. Lingkungan yang menguntungkan dapat meningkatkan pertumbuhan secara kuantitatif (Gardner et al. 2008).
Ketiga jenis rumput laut yang diteliti saat evaluasi performa budidaya, dibudidayakan di lokasi yang sama yang berarti bahwa kondisi lingkungan perairan juga sama. Namun demikian, bibit rumput laut hasil seleksi memperlihatkan pertumbuhan yang lebih baik dibandingkan kedua kontrol. Hal ini dapat dilihat dari ciri morfologi bibit hasil seleksi yang mempunyai talus lebih panjang, jumlah cabang lebih banyak, namun jarak ruas yang lebih pendek daripada kedua kontrol. Morfologi seperti ini dapat memberikan luas permukaan yang lebih besar dalam penyerapan nutrien serta cahaya matahari untuk proses fotosintesis. Hasil penelitian Stewart & Carpenter (2003) menunjukkan bahwa laju fotosintesis makroalga meningkat dengan semakin meningkatnya rasio luas permukaan:volume.
Pertumbuhan talus merupakan akibat dari pembelahan dan pembesaran sel yang terjadi di jaringan meristem. Jaringan meristem adalah jaringan yang bersifat embrionik, yaitu sel-selnya mempunyai kemampuan membelah diri yang tinggi secara terus-menerus. Menurut Gardner et al. (2008) jaringan meristem terdapat pada beberapa bagian tumbuhan, yaitu meristem ujung, interkalar dan lateral. Meristem ujung terletak di ujung akar atau batang, sedangkan meristem lateral terletak di organ yang mengalami pelebaran. Sementara itu, meristem interkalar terletak diantara jaringan yang terdiferensiasi. Pada penelitian ini, jaringan meristem ujung menghasilkan sel-sel baru di ujung talus rumput laut yang mengakibatkan talus bertambah panjang. Meristem lateral menghasilkan sel-sel baru yang memperluas diameter talus. Meristem interkalar memberi pemanjangan sel-sel pada ruas-ruas cabang.
Karakteristik fisiologis bibit rumput laut
Karakteristik morfologis didukung oleh karakteristik fisiologis. Karakteristik fisiologis pertumbuhan yang diukur pada penelitian ini mencakup LPH, kadar gula total dan kadar hormon yang terkandung dalam tiga jenis bibit.
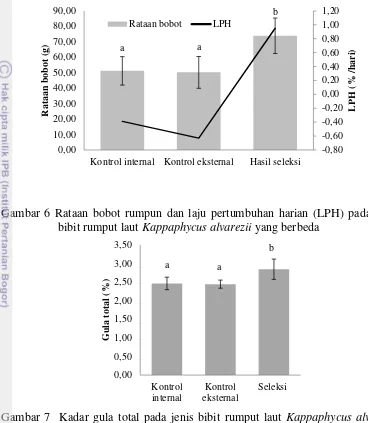
LPH merupakan salah satu karakteristik fisiologis yang telah dijadikan patokan pertumbuhan rumput laut. Kegiatan budidaya rumput laut dikategorikan baik jika rataan LPH minimal 3%. Selama evaluasi performa budidaya, rumput laut kontrol internal, kontrol eksternal dan hasil seleksi ternyata hanya mempunyai LPH di bawah 1% (Gambar 6). Bahkan rumput laut kontrol internal dan eksternal tidak mengalami pertumbuhan karena terjadi penurunan rataan bobot rumpun, sehingga LPH di bawah 0%.
11
diserang oleh biofouling dan mengalami bleaching (pemutihan warna). Kondisi ini terjadi karena lingkungan kurang optimal untuk pertumbuhan rumput laut (Tabel 2).
Gambar 6 Rataan bobot rumpun dan laju pertumbuhan harian (LPH) pada jenis bibit rumput laut Kappaphycus alvarezii yang berbeda
Gambar 7 Kadar gula total pada jenis bibit rumput laut Kappaphycus alvarezii yang berbeda
Pengukuran kadar gula total bibit rumput laut (30 hari) dalam penelitian ini digunakan untuk menduga potensi kadar karaginan rumput laut pada umur panen (45 atau 60 hari). Hasil penelitian menunjukkan bahwa kadar gula total pada bibit rumput laut hasil seleksi lebih tinggi daripada kedua kontrol (p<0,05) (Gambar 7). Kadar gula total bibit hasil seleksi adalah 15,52% lebih tinggi daripada kontrol internal dan 16,42% lebih tinggi daripada kontrol eksternal. Hasil ini sejalan dengan hasil penelitian Pong-Masak et al. (2013) yang menyatakan bahwa ada indikasi bibit hasil seleksi memiliki kadar karaginan yang lebih baik, yaitu dengan peningkatan sebesar 6,07-12,72% dibandingkan dengan kontrol internal dan kontrol eksternal.
Kontrol internal Kontrol eksternal Hasil seleksi
12
Selama pertumbuhan bibit, dinding sel terisi dengan karaginan. Karaginan adalah polisakarida linier sulfat dari D-galaktosa dan 3,6 anhidro-D-galaktosa yang diekstraksi dari rumput laut kelas Rhodophyceae yang merupakan bahan pengisi dinding sel (Campo et al. 2009). Lebih dari 60% dari bahan dinding sel rumput laut ini adalah karaginan (Atmadja 2001). Karaginan dan biomassa sangat berhubungan dengan waktu panen optimum. Waktu panen optimum untuk spesies Kappaphycus adalah 8-9 minggu budidaya (Villanueva et al. 2011). Umumnya, rumput laut dengan LPH yang lebih tinggi mempunyai karaginan yang lebih tinggi pula (Naguit et al. 2009). Dengan demikian rumput laut hasil seleksi berpotensi tinggi meningkatkan biomassa panen dan produksi karaginan.
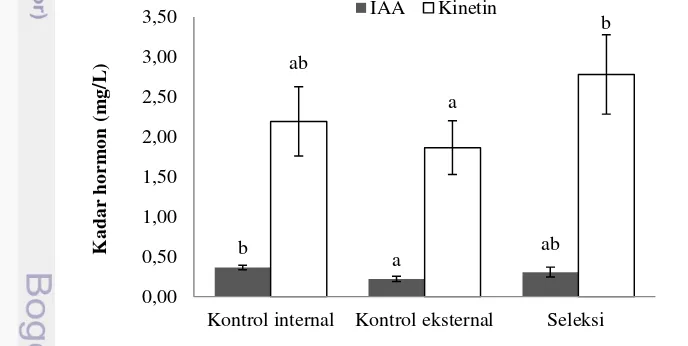
Pertumbuhan dan perkembangan tumbuhan dipengaruhi oleh hormon sebagai pengatur pertumbuhan. Sejalan dengan LPH yang lebih tinggi pada rumput laut hasil seleksi, kadar hormon kinetin dan rasio kinetin-IAA cenderung juga lebih tinggi (p<0,05) pada rumput laut hasil seleksi dibanding kedua kontrol (Gambar 8 dan 9). Kadar hormon kinetin dan rasio kinetin-IAA yang lebih tinggi mengindikasikan bahwa kedua hormon tersebut berperan penting dalam pertumbuhan rumput laut. Kinetin merupakan salah satu jenis hormon golongan sitokinin dan IAA termasuk golongan auksin.
Pada tumbuhan tingkat tinggi, pertumbuhan akar dan batang tergantung pada rasio sitokinin-auksin. Kadar sitokinin tinggi mendukung pembentukan batang dan kadar IAA yang tinggi mendorong pertumbuhan akar (Brault & Maldiney 1999; Moubayidin et al. 2009). Pada penelitian ini tampak bahwa pertumbuhan cabang-cabang talus rumput laut sejalan dengan pertumbuhan batang pada tumbuhan tingkat tinggi.
Gambar 8 Kadar hormon indole acetic acid (IAA) dan hormon kinetin pada jenis bibit rumput laut Kappaphycus alvarezii yang berbeda
13
Gambar 9 Rasio kinetin:IAA pada jenis bibit rumput laut Kappaphycus alvarezii yang berbeda
Yokoya (2010) telah mengidentifikasi keberadaan hormon endogen berupa auksin, sitokinin dan abscisic acid (ABA) pada 11 spesies alga merah dari pantai Brazil. Secara in vitro, auksin dan sitokinin mempunyai peran pengaturan dalam pertumbuhan dan morfogenesis pada beberapa spesies alga merah seperti Gracilaria tenuistipitata, G. perplexa (Yokoya et al. 2004) dan Solieria filiformis (Yokoya & Handro 2002).
Aktivitas fisiologis dari hormon sitokinin adalah mengendalikan sitokinesis (pembelahan sel) dan perkembangan tunas; sedangkan auksin menginduksi pemanjangan (Tarakhovskaya et al. 2007). Sitokinin juga menengahi respons terhadap faktor variabel ekstrinsik seperti kondisi cahaya dan ketersediaan nutrisi dan air, dan memiliki peran dalam respons terhadap stres biotik dan abiotik. Secara bersama-sama, kedua hormon tersebut berkontribusi pada regulasi pertumbuhan kuantitatif pada tanaman (Werner & Schmulling 2009).
Hubungan antara hormon dan pertumbuhan bibit hasil seleksi
Kandungan hormon dalam penelitian ini didapatkan dari bagian keseluruhan percabangan. Berdasarkan analisis korelasi antara kadar homon dengan karakteristik morfologi pada bibit hasil seleksi didapatkan bahwa hormon IAA berkorelasi positif dengan panjang total cabang (R2= 0,66), jumlah total cabang (R2= 0,69) dan LPH (R2= 0,66). Sementara itu, hormon kinetin berkorelasi positif dengan panjang total cabang (R2= 0,83), jumlah total cabang (R2= 0,70) dan LPH (R2= 0,94). Hasil ini mengindikasikan bahwa keberadaan hormon kinetin memacu LPH, meningkatkan panjang total cabang dan jumlah total cabang lebih tinggi dibandingkan dengan IAA. Kinetin banyak terakumulasi di bagian percabangan, sedangkan IAA cenderung terakumulasi lebih sedikit. Diduga hal ini disebabkan kinetin dan IAA dihasilkan di bagian talus yang berbeda.
14
sedangkan pengangkutan secara akropetal adalah sebaliknya. Hal ini dikarenakan auksin dihasilkan bagian ujung batang, sedangkan sitokinin dihasilkan bagian akar (Salisbury & Ross 1995). Dalam penelitian ini diduga IAA dihasilkan di bagian ujung percabangan rumput laut kemudian diangkut menuju ke arah talus utama, sedangkan kinetin dihasilkan di bagian talus utama lalu diangkut ke arah ujung-ujung percabangan.
Hubungan antara warna talus dan karakter fisiologis pertumbuhan bibit hasil seleksi
Analisis korelasi antara warna talus dan karakter fisiologis pertumbuhan bibit hasil seleksi didapatkan bahwa warna talus berkorelasi positif dengan LPH (R2= 0,99), kadar IAA (R2= 0,73) dan kadar kinetin (R2= 0,94). Hasil ini menginformasikan bahwa warna talus berbanding lurus dengan beberapa kondisi fisiologis rumput laut. Semakin pucat warna talus maka LPH semakin lambat dan kadar hormon semakin rendah.
Berdasarkan hasil penelitian, warna talus mengalami perubahan dari bulan Agustus ke Oktober. Talus berwarna coklat gelap di bulan Agustus, lalu kuning pucat di bulan September dan coklat muda di bulan Oktober (Tabel 2). Warna kuning pucat di bulan September mengindikasikan adanya penurunan kadar pigmen dalam talus. Kemungkinan hal ini disebabkan tidak adanya hujan di bulan September yang memberi dugaan adanya pengaruh tingginya intensitas cahaya matahari. Dugaan lainnya yaitu kandungan nutrien dalam perairan tidak mencukupi untuk pembentukan pigmen.
Cahaya matahari mengandung radiasi sinar UV. Jika K. alvarezii terpapar radiasi sinar UV-B terlalu lama maka dapat menyebabkan penurunan kandungan pigmen klorofil a, fikoeritrin dan fikosianin. Selanjutnya, penurunan kandungan pigmen ini akan mempengaruhi proses fotosintesis dan laju pertumbuhan (Eswaran et al. 2001; Schmidt et al. 2010).
Hasil penelitian Mizuta et al. (2002) menyatakan bahwa perubahan musim telah mempengaruhi komposisi pigmen pada dua spesies alga merah yaitu Gloiopeltis furcata and Porphyra yezoensis. Konsentrasi klorofil a dan fikoeritrin tinggi selama musim dingin tapi menurun di akhir musim semi atau musim panas. Perubahan ini terjadi dengan perubahan warna dari merah gelap menjadi hijau atau kuning. Hal ini terkait erat dengan fluktuasi kandungan fikoeritrin. Kandungan fikoeritrin berkorelasi dengan kandungan nitrogen pada kedua spesies (Mizuta et al. 2002).
Kondisi lingkungan lokasi penelitian
15
performa budidaya pada bulan Agustus sampai Oktober 2013. Bahkan pada bulan September hingga akhir Oktober tidak ada hujan.
Gambar 10 Kondisi curah hujan di lokasi seleksi massa (atas) dan evaluasi performa budidaya Kappaphycus alvarezii (bawah). Garis putus-putus menunjukkan waktu penelitian. Gambar diolah dari data curah hujan Stasiun Tilamuta (atas) dan Kwandang (bawah), Stasiun Meteorologi (BMKG) Gorontalo
Beberapa variabel kualitas perairan berada di luar kondisi optimum pertumbuhan rumput laut (Tabel 2). Penurunan kualitas lingkungan yang dipicu salinitas tinggi, fosfat rendah dan nitrat tinggi di bulan Agustus menyebabkan berkembangnya biofouling berupa teritip dan alga epifit berfilamen yang menempel dan menutupi permukaan talus (Gambar 11). Alga epifit berfilamen dan teritip yang menutupi talus dapat menghalangi penyerapan cahaya matahari dan nutrien. Jika terjadi dalam jangka waktu lama maka jaringan talus akan mati. Menurut Tisera & Naguit (2009), permukaan talus K. alvarezii yang kasar memudahkan epifit, parasit dan mikroorganisme menempel. Semakin kuat biofouling ini menempel maka bisa menjadi media bagi patogen untuk
Bulan
Tabel 2 Kualitas perairan Gorontalo Utara dan kondisi bibit Kappaphycus alvarezii pada saat evaluasi performa budidaya
Variabel Unit Bulan Kondisi optimuma
Agustus September Oktober
Kualitas perairan
Suhu °C 28 29 30 24-30
Oksigen terlarut mg/L 5,62 6,31 5,93 >5b
Salinitas g/L 36 33 34 28-34
Total Fosfat mg/L 0,03 <0,002 0,15 0,10-0,20
Nitrat mg/L 0,40 0,19 0,78 0,01-0,07
BOT mg/L 47,03 36,07 38,65
Kondisi bibit
Warna talus Coklat gelap Kuning pucat Coklat muda
Biofouling Teritip; alga epifit - -
16
berkembang. Patogen kemudian memasuki lapisan korteks dan medula talus sehingga talus menjadi lemah. Talus yang lemah cenderung mengalami infeksi bakteri (Tisera & Naguit 2009).
A B
Gambar 11 Talus yang ditutupi oleh alga epifit (A) dan teritip (B) di bulan Agustus
Biofouling yang menempel pada talus diduga disebabkan oleh kecepatan arus laut yang lemah. Dalam penelitian ini kecepatan arus tidak diukur. Kecepatan arus yang lemah diduga pula menjadi penyebab pertumbuhan talus lambat karena pengambilan nutrien dari lingkungan perairan juga lambat. Menurut Largo (2002), arus yang kuat selain meningkatkan pertukaran nutrien juga mencegah patogen potensial untuk menempel pada permukaan talus rumput laut. Jika arus lemah maka patogen khususnya bakteri menjadi sangat motil dan cepat menyerang permukaan talus. Menurut BPPBAP (2011), kecepatan arus untuk budidaya rumput laut adalah 25-40 cm/detik. Kecepatan arus merupakan faktor ekologi primer yang memungkinkan terjadinya aerasi, suplai nutrien secara tetap, terhindar dari bahan-bahan tersuspensi dalam air dan epifit serta menyebabkan fluktuasi salinitas dan suhu yang kecil (BPPBAP 2011).
Pang et al. (2011) menyatakan bahwa alga epifit berfilamen tercatat menjadi penyebab menurunnya produksi dan kualitas K. alvarezii. Hasil studi secara morfologi didapatkan bahwa epifit utama K. alvarezii di China adalah Neosiphonia savatieri. Epifit ini mempengaruhi aktivitas fotosintesis K. alvarezii. (Pang et al., 2011). Sementara itu di Filipina, Indonesia, Malaysia dan Tanzania, alga epifit berfilamen yang teridentifikasi menginfeksi karaginofit adalah N. apiculata (Vairappan et al. 2009).
17
dan kandungan karaginan. Karaginan maksimum diperoleh selama periode suplai nutrien tertinggi (Orbita 2013).
Tabel 3 Variabel morfologi dan fisiologi tiga jenis bibit rumput laut Kappaphycus alvarezii pada tiga siklus yang berbeda
Variabel uji Siklus
Angka-angka pada baris yang sama yang diikuti oleh huruf superskrip berbeda menunjukkan berbeda nyata pada taraf uji 5% (uji jarak berganda Duncan)
Fried et al. (2003) menyatakan bahwa jika hanya satu nutrien yang meningkat maka nutrien lain akan membatasi pertumbuhan. Jika nitrogen berada pada konsentrasi yang sangat tinggi dan fosfor berada pada konsentrasi rendah, maka fosfor akan menjadi faktor pembatas pertumbuhan pada alga (Fried et al. 2003). Menurut Correll (1998), fosfor adalah komponen esensial asam nukleat dan banyak metabolit perantara, seperti gula fosfat dan adenosin fosfat. Sementara itu nitrogen adalah komponen protein dan ditemukan di banyak senyawa biologi (Rezael & Samimi 2013).
18
(Lampiran 2). Cara ini memberikan hasil yang cukup baik bagi talus rumput laut. Di bulan Oktober, warna talus berubah menjadi coklat muda yang mengindikasikan telah terjadi proses pemulihan. Pemulihan kondisi ini kemungkinan juga disebabkan kandungan fosfor dalam perairan yang sudah berada dalam jumlah optimal.
Kondisi lingkungan perairan pada saat evaluasi performa budidaya dalam penelitian ini sejalan dengan yang dilaporkan oleh Pong-Masak et al. (2009), yang menyatakan bahwa musim tanam rumput laut K. alvarezii yang produktif di perairan Gorontalo Utara adalah pada bulan November sampai April, sedangkan pada bulan Mei sampai Oktober perairan kurang produktif. Hal ini disebabkan terjadi penurunan kualitas lingkungan, berkembangnya biofouling dan serangan penyakit ice-ice sehingga laju pertumbuhan rumput laut kurang dari 3% (Pong-Masak et al. 2009). Pada penelitian ini didapatkan bahwa kondisi musim peralihan dan kemarau kurang sesuai untuk budidaya rumput laut K. alvarezii.
4 SIMPULAN
Simpulan
1. Bibit Kappaphycus alvarezii hasil seleksi mempunyai talus yang lebih panjang, jumlah cabang lebih banyak dan jarak ruas lebih pendek; serta rataan bobot dan LPH lebih tinggi, kadar gula total, kadar hormon kinetin dan rasio kinetin:IAA lebih tinggi daripada kontrol.
19
DAFTAR PUSTAKA
[AOAC] Association of The Official Analytical Chemist. 1995. Official Method and Analysis of the Association of The Official Analytical Chemist. Washington (US): Association of The Official Analytical Chemist.
Aslan LM. 1998. Budidaya Rumput Laut. Yogyakarta (ID): Kanisius.
Atmadja W. 2001. Kappaphycus alvarezii (Doty) Doty ex P.C Silva. Di dalam:
Prud’homme van Reine WF dan Trono Jr GC, editor. Plant Resources of South-East Asia (Cryptogams:Algae). Leiden (AN): Blackhuys Publishers. hlm 240-246.
Brault M, Maldiney R. 1999. Mechanisms of cytokinin action. Plant Physiol Biochem. 37(5):403−412.
[BPPBAP] Balai Penelitian dan Pengembangan Budidaya Air Payau. 2011. Protokol Seleksi Varietas Bibit Unggul Rumput Laut. Maros (ID) : BPPBAP.
Campo VL, Kawano DF, Da Silva Jr DB, Carvalho I. 2009. Carrageenans: Biological properties, chemical modifications and structural analysis [ulasan]. Carbohydrate Polymers. 77: 167-180.
Correll DL. 1998. The role of phosphorus in the eutrophication of receiving waters: a review. Journal of Environmental Quality. 27(2):261-6.
Eswaran K, Rao PVS, Mairh OP. 2001. Impact of ultraviolet-B radiation on a marine red alga Kappaphycus alvarezii (Solieriaceae, Rhodophyta). Indian Journal of Marine Sciences. 30:105-107.
Fried S, Mackie B, Nothwehr E. 2003. Nitrate and phosphate levels positively affect the growth of algae species found in Perry Pond. Tillers 4:21-24. Gardner FP, Pearce RB, Mitchell. 2008. Fisiologi Tanaman Budidaya. Susilowati
H, penerjemah. Jakarta (ID): UI Press.
Glen EP, Doty MS. 1990. Growth of the seaweeds Kappaphycus alvarezii, K. striatum and Eucheuma denticulatum as affected by environment in Hawaii. Aquaculture. 84:245-255.
Hurtado AQ, Cheney DP. 2003. Propagule production of Eucheuma denticulatum (Burman) Collins et Harvey by tissue culture. Bot Mar. 46: 338-341.
Kumar GJ, Reddy CRK, Ganesan M, Thiruppathi S, Dipakkore S, Eswaran K, Rao PVS, Jha B. 2004. Tissue culture and regeneration of thallus from callus of Geladiella acerosa (Geladiales, Rhodophyta). Phycologia. 43(5): 596-602.
Largo DB. 2002. Recent developments in seaweed diseases. Di dalam: Hurtado AQ, Guanzon NG, de Castro-Mallare TR, Luhan MRJ, editor. Proceedings of the National Seaweed Planning Workshop; 2001Agustus 2-3, Tigbauan, Philipina. Tigbauan (PH): SEAFDEC Aquaculture Department. hlm 35-42. [MENLH] Kementerian Lingkungan Hidup. 2004. Keputusan Menteri Negara
Lingkungan Hidup Nomor 51Tahun 2004 tentang Baku Mutu Air Laut untuk Biota Laut. Jakarta (ID): MENLH.
20
Mizuta H, Shirakura Y, Yasui H. 2002. Relationship between phycoerythrin and nitrogen content in Gloiopeltis furcata and Porphyra yezoensis. Algae. 17(2):89-93.
Moubayidin L, Di Mambro R, Sabatini S. 2009. Cytokinin-auxin crosstalk [ulasan]. Trends in Plant Science. 14 (10): 557-562.
Mulyaningrum SRH, Nursyam H, Risjani Y, Parenrengi A. 2012. Regenerasi filamen kalus rumput laut Kappaphycus alvarezii dengan formulasi zat pengatur tumbuh yang berbeda. Jurnal Penelitian Perikanan. 1(1): 52-60. Naguit MRA, Tisera WL, Lanioso A. 2009. Growth performance and carrageenan
yield of Kappaphycus alvarezii (Doty) and Eucheuma denticulatum (Burman) Collins et Harvey, farmed in Bais Bay, Negros Oriental and Olingan, Dipolog City. The Threshold. 4: 38-51.
Ongaro V, Leyser O. 2008. Hormonal control of shoot branching. Journal of Experimental Botany. 59(1):67–74.
Orbita MLS. 2013. Growth rate and carrageenan yield of Kappaphycus alvarezii (Rhodophyta, Gigartinales) cultivated in Kolambugan, Lanao del Norte, Mindanao, Philippines. AAB Bioflux. 5(3): 128-139.
Pang T, Liu J, Liu Q, Lin W. 2011. Changes of photosynthetic behaviors in Kappaphycus alvarezii infected by epiphyte. Evidence-Based Complementary and Alternative Medicine. doi: 10.1155/658906.
Parenrengi A, Rachmansyah, Suryati E. 2011. Budidaya Rumput Laut Penghasil Karaginan (Karaginofit) : Seri Teknologi untuk Minapolitan. Jakarta (ID) : Badan Penelitian dan Pengembangan Kelautan dan Perikanan.
Pong-Masak PR, Pantjara B, Rachmansyah. 2009. Musim Tanam Rumput Laut di Perairan Anggrek, Pantai Utara Gorontalo. Prosiding Seminar Nasional VI. Hasil Penelitian Perikanan dan Kelautan Tahun 2009 UGM_BRKP. RB-11. Hal 1-10.
Pong-Masak PR, Parenrengi A, Tjaronge M. 2013. Produksi Bibit Unggul Rumput Laut Kappaphycus alvarezii. Di dalam: Sukadi F, Djunaidah IS, Nurkhakim S, Sugama K, Heruwati ES, Purba M, Hanggono A, editor. Rekomendasi Teknologi Kelautan dan Perikanan. Jakarta (ID): Badan Litbang Kelautan dan Perikanan. hlm 161-175.
Rajamuddin MAL. 2010. Introduksi dan Ekspresi Gen GFP (Green Fluorescent Protein) dengan Promoter Berbeda pada Sel Embrio Somatik Rumput Laut Kappaphycus alvarezii [tesis]. Bogor (ID): Institut Pertanian Bogor.
Reddy CRK, Kumar GRK, Shiddhanta AK, Tewari A. 2003. In vitro somatic embryogenesis and regeneration of somatic embryos from pigmented callus of Kappaphycus alvarezii (Doty) Doty (Rhodophyta, Gigartinales). J Phycol. 39: 610-616.
Rezael R, Samimi A. 2013. Effects of phosphorus and nitrate in wastewater Whahinshahr City use for oil refinery. International Journal of Innovation and Applied Studies. 2(3):250-258.
21
Schmidt EC, Maraschin M, Bouzon ZL. 2010. Effects of UVB radiation on the carragenophyte Kappaphycus alvarezii (Rhodophyta, Gigartinales): changes in ultrastructure, growth, and photosynthetic pigments. Hydrobiologia. 649(1):171-182.
Stewart HL, Carpenter RC. 2003. The effects of morphology and water flow on photosynthesis of marine macroalgae. Ecology. 84(11):2999-3012.
Sulistiani E, Soelistyowati DT, Alimuddin, Yani SA. 2012. Callus induction and filaments regeneration from callus of cottonii seaweed (Kappaphycus alvarezii (Doty)) collected from Natuna Islands, Riau Islands Province. Biotropia.19(2):103-114.
Suryati E, Fadilah S, Mujayana. 2012. Regenerasi mikroplanlet Kappaphycus alvarezii pada media PES 1/20 yang diperkaya indole acetic acid (IAA), zeatin dan kinetin. Prosiding Forum Inovasi Teknologi Akuakultur, Makassar.
Suryati E, Mulyaningrum SRH, Parenrengi A. 2013. Budidaya Rumput Laut Gracilaria verrucosa menggunakan Bibit Hasil Kultur Jaringan. Di dalam: Sukadi F, Djunaidah IS, Nurkhakim S, Sugama K, Heruwati ES, Purba M, Hanggono A, editor. Rekomendasi Teknologi Kelautan dan Perikanan. Jakarta (ID): Badan Litbang Kelautan dan Perikanan. hlm 207-213.
Tarakhovskaya ER, Maslov YI, Shishova MF. 2007. Phytohormones in algae [ulasan]. Russian Journal of Plant Physiology.54(2): 163-170.
Taran SA, Nasibullah S, Mian MA, Shah H. 2004. Genetic improvement through mass selection in maize (Zea mays L.). J App Em Sc. 1(1):152-157.
Tisera WL, Naguit MRA. 2009. Ice-ice disease occurrence in seaweed farms in Bais Bay, Negros Oriental and Zamboanga Del Norte. The Threshold. 4: 1-16.
Unyayar S, Topcuoglu SF, Unyayar A. 1996. A modified method for extraction and modification of indole-3-acetic acid (IAA), gibberellic acid (GA3), abscisic acid (ABA) and zeatin produced by Phanerochate chrysosporium ME446. Bulg J Plant Physiol. 22 (3-4): 105-110.
Vairappan CS, Chung CS, Hurtado AQ, Soya FE, Lhonneur GB, Critchley A. 2009. Distribution and symptoms of epiphyte infection in major carrageenophyte-producing farms. Nineteenth International Seaweed Symposium Developments in Applied Phycology Volume 2 : 27-33
Villanueva RD, Romero JB, Montano MNE, de la Pena PO. 2011. Harvest optimization of four Kappaphycus species from the Philippines. Biomass and Bioenergy. 35(3):1311-1316.
Wahyuti TB. 2012. Hubungan Karakter Morfologi dan Fisiologi dengan Hasil dan Upaya Meningkatkan Hasil Padi Varietas Unggul [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Werner T, Schmulling T. 2009. Cytokinin action in plant development. Current Opinion in Plant Biology. 12: 527-538.
Yang Y, Chen J, Zhao M, Li C, Meng Z, Wang J, Chen Z, Zhang G, Yang S. 2013. Phenotypic and genetic uniformity in three populations of Panax notoginseng by mass selection. Molecular Plant Breeding. 4(21): 169-176. Yokoya NS, Handro W. 2002. Effects of plant growth regulators and culture
22
Yokoya NS, West JA, Luchi AE. 2004. Effects of plant growth regulators on callus formation, growth and regeneration in axenic tissue cultures of Gracilaria tenuistipitata and Gracilaria perplexa (Gracilariales, Rhodophyta). Phycological Research. 52:244–254.
23
Lampiran 1 Diagram alir proses pemurnian IAA, GA3 , ABA dan zeatin (Unyayar
et al. 1996)
ditambah aquades 22,4 ml
kloroform (dibuang)
pH diatur menjadi 2,5 kombinasi ekstrak (100 ml)
dengan HCl 7N atau NaOH 7 N pH diatur menjadi 2,5
diekstrak 3 kali dengan dengan HCl 7N atau NaOH 7 N
etilasetat (15 ml) diekstrak 3 kali dengan
etilasetat (15 ml)
pH diatur menjadi 7 pH diatur menjadi 7
dengan HCl 7N atau NaOH 7 N dengan HCl 7N atau NaOH 7 N
diekstrak 3 kali dengan diekstrak 3 kali dengan
etilasetat (15 ml) etilasetat (15 ml)
pH diatur menjadi 11 pH diatur menjadi 11
dengan HCl 7N atau NaOH 7 N dengan HCl 7N atau NaOH 7 N
dihidrolisis selama 1 jam dihidrolisis selama 1 jam
pada suhu 70°C pada suhu 70°C
pH diatur menjadi 7 pH diatur menjadi 7
dengan HCl 7N atau NaOH 7 N dengan HCl 7N atau NaOH 7 N
diekstrak 3 kali dengan diekstrak 3 kali dengan
etilasetat (15 ml) etilasetat (15 ml)
pH diatur menjadi 2,5 pH diatur menjadi 2,5
dengan HCl 7N atau NaOH 7 N dengan HCl 7N atau NaOH 7 N
diekstrak 3 kali dengan diekstrak 3 kali dengan
etilasetat (15 ml) etilasetat (15 ml)
24
Lampiran 2 Kandungan pupuk organik cair Alam Hijau dan perendaman rumput laut dengan pupuk organik cair
Senyawa persentase
C organik ≥ 4
N < 2
PO < 2
KO < 2
pH 4-8
PT. Disantya prestasi
25
Lampiran 3 Hasil analisis ragam dan uji lanjut pada variabel panjang talus utama dan panjang cabang I
Corrected Total 6681.605 26
a. R Squared = .664 (Adjusted R Squared = .603)
Corrected Model 81548.461a 4 20387.115 3.285 .030
Intercept 489152.552 1 489152.552 78.820 .000
Jenis_bibit 27072.041 2 13536.020 2.181 .137
Bulan 54476.421 2 27238.210 4.389 .025
Error 136531.397 22 6205.973
Total 707232.410 27
Corrected Total 218079.858 26
26
Lampiran 4 Hasil analisis ragam dan uji lanjut pada variabel panjang cabang II dan cabang III
Corrected Model 114851.397a 4 28712.849 11.673 .000
Intercept 754613.740 1 754613.740 306.789 .000
Jenis_bibit 58260.683 2 29130.342 11.843 .000
Bulan 56590.713 2 28295.357 11.504 .000
Error 54113.659 22 2459.712
Total 923578.797 27
Corrected Total 168965.056 26
a. R Squared = .680 (Adjusted R Squared = .622)
Intercept 132902.685 1 132902.685 123.190 .000
Jenis_bibit 18918.822 2 9459.411 8.768 .002
Bulan 9775.874 2 4887.937 4.531 .023
Error 23734.582 22 1078.845
Total 185331.963 27
Corrected Total 52429.278 26
27
Lampiran 5 Hasil analisis ragam dan uji lanjut pada variabel total panjang talus dan jumlah cabang I
Corrected Model 542048.632a 4 135512.158 9.031 .000
Intercept 4337481.650 1 4337481.650 289.057 .000
Jenis_bibit 228891.573 2 114445.787 7.627 .003
Bulan 313157.059 2 156578.529 10.435 .001
Error 330123.926 22 15005.633
Total 5209654.208 27
Corrected Total 872172.558 26
a. R Squared = .621 (Adjusted R Squared = .553)
Corrected Total 18108.296 26
28
Corrected Model 94251.556a 4 23562.889 4.820 .006
Intercept 239701.333 1 239701.333 49.034 .000
Jenis_bibit 48942.889 2 24471.444 5.006 .016
Bulan 45308.667 2 22654.333 4.634 .021
Error 107547.111 22 4888.505
Total 441500.000 27
Corrected Total 201798.667 26
a. R Squared = .467 (Adjusted R Squared = .370)
Corrected Model 47505.704a 4 11876.426 7.659 .001
Intercept 145934.259 1 145934.259 94.110 .000
Jenis_bibit 24371.185 2 12185.593 7.858 .003
Bulan 23134.519 2 11567.259 7.459 .003
Error 34115.037 22 1550.684
Total 227555.000 27
Corrected Total 81620.741 26
29
Lampiran 7 Hasil analisis ragam dan uji lanjut pada variabel total jumlah cabang dan jarak ruas primer
Corrected Model 344196.370a 4 86049.093 5.931 .002
Intercept 1217731.704 1 1217731.704 83.937 .000
Jenis_bibit 157312.519 2 78656.259 5.422 .012
Bulan 186883.852 2 93441.926 6.441 .006
Error 319167.926 22 14507.633
Total 1881096.000 27
Corrected Total 663364.296 26
30
31
32
Corrected Model 25320.230a 4 6330.058 120.369 .000
Intercept 92186.764 1 92186.764 1752.969 .000
Jenis_bibit 3233.920 2 1616.960 30.747 .000
Bulan 22086.310 2 11043.155 209.990 .000
Error 1156.957 22 52.589
Total 118663.951 27
Corrected Total 26477.187 26
33
Corrected Total 107.613 26
a. R Squared = .955 (Adjusted R Squared = .947)
Corrected Total 128.252 16
34
Lampiran 12 Hasil analisis ragam dan uji lanjut pada variabel kadar Kinetin dan IAA
Corrected Total 25.532 16
35 RIWAYAT HIDUP
Penulis dilahirkan di Bangkalan pada tanggal 22 September 1982. Penulis adalah putri pertama dari pasangan Achmad Farihin dan Suriyah. Penulis telah menempuh pendidikan sarjana di jurusan Biologi, Fakultas MIPA, Universitas Brawijaya dan lulus tahun 2005. Tahun 2012, penulis mendapat kesempatan untuk melanjutkan ke program magister pada Sekolah Pascasarjana di program studi Ilmu Akuakultur, Fakultas Perikanan dan Kelautan, Institut Pertanian Bogor. Beasiswa pendidikan Pascasarjana didapatkan dari Kementerian Kelautan dan Perikanan Republik Indonesia.