KEMAMPUAN MATURASI DAN FERTILISASI OOSIT SAPI
YANG DISELEKSI MENGGUNAKAN TEKNIK PEWARNAAN
BRILLIANT CRESYL BLUE SECARA IN VITRO
ZULTINUR MUTTAQIN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Kemampuan Maturasi dan Fertilisasi Oosit Sapi yang Diseleksi Menggunakan Teknik Pewarnaan Brilliant Cresyl Blue secara in vitro adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
ZULTINUR MUTTAQIN. Kemampuan Maturasi dan Fertilisasi Oosit Sapi yang Diseleksi Menggunakan Teknik Pewarnaan Brilliant Cresyl Blue secara in vitro. Dibimbing oleh MOHAMAD AGUS SETIADI dan NI WAYAN KURNIANI KARJA
Tingkat keberhasilan produksi embrio sapi in vitro masih sangat rendah. Hal ini terutama dikaitkan dengan kualitas oosit yang digunakan saat maturasi. Seleksi rutin oosit berdasarkan kriteria morfologi masih menghasilkan oosit yang heterogen dalam kualitas dan kemampuan perkembangannya, dikarenakan aktivitas intraovarian yang terjadi pada masa hidupnya. Dilaporkan aktivitas enzim glukosa 6 fosfat dehidrogenase (G6PD) dalam sitoplasma oosit dapat dijadikan sebagai indikator kualitas oosit. Aktivitas enzim ini dilaporkan tinggi pada oosit yang sedang tumbuh dan akan bekurang aktivitasnya ketika oosit sudah tumbuh sempurna. Indikasi ini kemudian dijadikan acuan oleh para peneliti untuk mengembangkan sebuah metode non invasif seleksi oosit melalui pewarnaan dengan brilliant cresyl blue (BCB). Pewarna BCB yang ditambahkan pada sekumpulan oosit dapat bereaksi dan menyeleksi oosit berdasarkan aktivitas intraseluler G6PD di sitoplasma.
Penelitian ini bertujuan untuk mengkaji penggunaan pewarna BCB sebagai indikator aktivitas G6PD di sitoplasma oosit dalam upaya memilih oosit sapi yang lebih kompeten untuk berkembang secara in vitro. Sejumlah oosit dipaparkan pada 26 µM BCB selama 90 menit dalam inkubator 5% CO2 suhu 39ºC dan
diklasifikasikan berdasarkan tingkat penyerapan warna sitoplasmanya. Oosit dengan sitoplasma berwarna biru dikategorikan sebagai kelompok oosit BCB+ dan oosit dengan sitoplasma yang tidak berwarna dikategorikan sebagai kelompok oosit BCB-. Kelompok oosit kontrol dimaturasi secara langsung setelah terseleksi secara morfologi, tanpa dilakukan pewarnaan BCB. Masing-masing kelompok perlakuan (BCB+, BCB- dan kontrol) kemudian dilakukan maturasi dan fertilisasi secara in vitro. Oosit dikatakan matang jika dapat mencapai tahap metafase II (MII) setelah di maturasi selama 24 jam dan oosit dengan 2 atau lebih pronukleus (PN) setelah 14 jam inkubasi diklasifikasikan sebagai oosit yang telah terfertilisasi. Kedua parameter tersebut kemudian dievaluasi dan dibandingkan diantara perlakuan.
Persentase oosit yang mencapai tahap MII pada kelompok oosit BCB+ lebih tinggi (P< 0.05) dibandingkan kelompok oosit BCB- (78.7% vs 33.3%), namun tidak terdapat perbedaan yang signifikan (P> 0.05) antara kelompok oosit BCB+ dan kelompok oosit kontrol (78.7% vs 77.1%). Tingkat fertilisasi kelompok oosit BCB+ juga ditemukan lebih tinggi secara signifikan (P< 0.05) dibandingkan kelompok oosit BCB- dan kelompok oosit kontrol (30.5% vs 13.6, 23.6%). Hal ini mengindikasikan bahwa kelompok oosit BCB+ lebih kompeten untuk termaturasi dan terfertilisasi secara in vitro dibandingkan kelompok oosit BCB-. Dari hasil penelitian dapat disimpulkan bahwa seleksi oosit sapi dengan BCB sebelum maturasi dapat digunakan secara efektif dalam memilih oosit yang lebih kompeten untuk berkembang.
SUMMARY
ZULTINUR MUTTAQIN. Maturation and Fertilization Ability of Bovine Oocytes in vitro Selected Using Brilliant Cresyl Blue. Supervised by MOHAMAD AGUS SETIADI and NI WAYAN KURNIANI KARJA.
The success rate of bovine in vitro embryo production is still low. This is mainly attributed to the oocyte quality at the start of maturation. Routine selection of oocyte quality based on morphological criteria results heterogeneous in quality and developmental competence in vitro due to intraovarian activity. The activity of glucose 6 phospate dehydrogenase (G6PD) was reported to be an indicator of oocyte quality. This enzyme was synthesized in growing oocytes but with decreased activity in fully grown oocytes. This indication was used by some researchers as a basis to develop a non invasive method for oocytes selection by staining with brilliant cresyl blue (BCB). The BCB staining could react and determines the intracellular activity of G6PD.
The aim of this study was to evaluate the utility of BCB as an indicator for G6PD activity in order to select developmentally competent bovine oocytes to develop in vitro. The oocytes were exposed to 26 µM BCB for 90 minutes in 5% CO2 incubator 39ºC and were classified according to their cytoplasm coloration:
oocytes with a blue cytoplasm (BCB+) and colorless oocytes (BCB-). The oocytes of the control group were incubated directly after selection without exposure to BCB. Then, the oocytes of all groups (BCB+, BCB-, and control) were matured and fertilized in vitro. Matured oocytes were defined as those oocytes that reach metaphase II stage after being cultured for 24 hours. Oocytes showing two or more pronuclei at 14 hours incubation were classified as fertilized oocytes. Both parameters were evaluated and compared between treatments.
The nuclear maturation rate was higher (P< 0.05) in BCB+ oocytes group than BCB- oocytes group (78.7% vs 33.3%), however there is no significant difference (P> 0.05) between BCB+ oocytes group and control oocytes group (78.7% vs 77.1%). The fertilization rate of BCB+ oocytes group was higher (P< 0.05) than those of BCB- and control oocytes group (30.5% vs 13.6, 23.6%). This indicated that oocytes BCB+ more competence to be matured and fertilized in vitro than oocytes BCB-. In conclusion, staining of bovine oocytes with BCB before in vitro maturation could be used effectively to select developmentally competent bovine oocytes.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biologi Reproduksi
KEMAMPUAN MATURASI DAN FERTILISASI OOSIT SAPI
YANG DISELEKSI MENGGUNAKAN TEKNIK PEWARNAAN
BRILLIANT CRESYL BLUE SECARA IN VITRO
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
Judul Tesis : Kemampuan Maturasi dan Fertilisasi Oosit Sapi yang Diseleksi Menggunakan Teknik Pewarnaan Brilliant Cresyl Blue secara in vitro
Nama : Zultinur Muttaqin NIM : B352120041
Disetujui oleh Komisi Pembimbing
Prof Dr drh Mohamad Agus Setiadi drh Ni Wayan Kurniani Karja MP, PhD
Ketua Anggota
Diketahui oleh
Ketua Program Studi Dekan Sekolah Pascasarjana
Biologi Reproduksi
Prof Dr drh Mohamad Agus Setiadi Dr Ir Dahrul Syah MScAgr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Februari sampai dengan Mei 2014 ini ialah kualitas oosit sapi, dengan judul Kemampuan Maturasi dan Fertilisasi Oosit Sapi yang Diseleksi Menggunakan Teknik Pewarnaan Brilliant Cresyl Blue secara in vitro.
Terima kasih penulis ucapkan kepada Bapak Prof Dr drh Mohamad Agus Setiadi sebagai Ketua Komisi Pembimbing dan Ibu drh Ni Wayan Kurniani Karja MP, PhD selaku Anggota Komisi Pembimbing atas bimbingan, arahan, perhatian dan nasihatnya selama melakukan penelitian dan penulisan karya ilmiah ini sehingga penulis dapat menyelesaikan rangkaian penelitian ini hingga selesai. Terima kasih kepada Bapak Prof Dr drh Iman Supriatna selaku penguji luar komisi atas saran dan kritiknya sehingga dapat lebih memperkaya dan menyempurnakan substansi tesis. Terima kasih juga kepada Bakrie Center Foundation atas beasiswa yang telah diberikan dalam program Bakrie Graduate Fellowship 2013 sehingga dapat membantu penulis selama masa studi. Ungkapan terima kasih juga disampaikan kepada ayah, ibu, teman-teman, serta seluruh keluarga, atas segala doa dan kasih sayangnya. Kepada cut yasmin, I would like to thank you for your care, support, love, and every little effort you are doing for me is simply amazing. Terima kasih telah menjadi partner terbaik..
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL x
DAFTAR GAMBAR x
DAFTAR LAMPIRAN x
1 PENDAHULUAN 1
Latar Belakang 1
Kerangka Pemikiran 2
Tujuan Penelitian 2
Manfaat Penelitian 2
Hipotesis Penelitian 2
2 TINJAUAN PUSTAKA 3
3 METODE 12
Waktu dan Tempat Penelitian 12
Metode Penelitian 12
Tahap I Kemampuan Maturasi Inti Oosit setelah Dipaparkan
Pewarna BCB 12
Koleksi dan Klasifikasi Oosit 12
Pewarnaan Brilliant Cresyl Blue (BCB) 12
Maturasi Oosit in vitro (IVM) 13
Evaluasi Kemampuan Maturasi Oosit in vitro 13
Tahap II Kemampuan Fertilisasi Oosit Setelah Dipaparkan
Pewarna BCB 14
Fertilisasi Oosit in vitro (IVF) 14
Evaluasi Kemampuan Fertilisasi Oosit in vitro 14
Analisis Data 14
4 HASIL DAN PEMBAHASAN 15
Hasil 15
Pembahasan 17
5 SIMPULAN DAN SARAN 21
Simpulan 21
Saran 21
DAFTAR PUSTAKA 22
LAMPIRAN 27
DAFTAR TABEL
1 Tingkat maturasi inti oosit sapi setelah pewarnaan BCB 16 2 Tingkat fertilisasi oosit sapi setelah pewarnaan BCB 17
DAFTAR GAMBAR
1 Gambaran skematis ultrastruktur oosit selama pertumbuhannya di
dalam folikel 4
2 Metabolisme karbohidrat melalui jalur fosfat pentosa 7 3 Skema distribusi organel selama pematangan dan fertilisasi 9 4 Mekanisme aktivasi oosit oleh spermatozoa dalam proses fertilisasi 10 5 Status inti oosit sapi setelah pematangan in vitro 15 6 Pembentukan pronukleus pada oosit sapi setelah fertilisasi in vitro 16
DAFTAR LAMPIRAN
1 Penghitungan konsentrasi BCB 26 µM 27
2 Komposisi medium transportasi ovarium 27
3 Komposisi medium koleksi oosit 28
4 Komposisi medium maturasi in vitro 28
1
1 PENDAHULUAN
Latar Belakang
Tingkat keberhasilan produksi embrio sapi in vitro masih sangat rendah karena sebagian besar oosit gagal untuk berkembang sampai ke tahap blastosis. Hal ini terutama dikaitkan dengan kualitas oosit yang digunakan saat maturasi. Oosit yang digunakan untuk produksi embrio in vitro umumnya diperoleh dari ovarium yang berasal dari rumah pemotongan hewan (RPH), dengan proses seleksi dilakukan secara morfologi berdasarkan jumlah dan kekompakan sel-sel kumulus serta homogenitas sitoplasmanya. Akan tetapi karena aktivitas intraovarian yang terjadi pada masa hidupnya, oosit yang terseleksi umumnya masih heterogen dalam kualitas dan kemampuan perkembangannya. Aktivitas intraovarian in vivo menghasilkan beberapa gelombang folikel yang memungkinkan oosit berada pada tahapan perkembangan yang berbeda-beda, seperti terdapatnya oosit yang sedang tumbuh, menuju ke arah atresia, dan atau sudah tumbuh sempurna (Ginther et al. 1999). Oleh karena itu, seleksi berdasarkan morfologi dirasa belum cukup memadai dan masih kurang efisien, karena ada kemungkinan oosit yang secara morfologi terseleksi dalam kriteria baik tetapi ternyata sudah berada dalam tahap mengalami degenerasi atau masih berada dalam tahap sedang tumbuh (Alm et al. 2005).
Dilaporkan bahwa pertumbuhan dan perkembangan oosit diindikasikan dengan adanya perubahan pada aktivitas enzim glukosa 6 fosfat dehidrogenase (G6PD) dalam sitoplasmanya. Aktivitas G6PD ditemukan tinggi dalam sitoplasma oosit yang sedang tumbuh (growing oocytes) dan aktivitasnya akan semakin menurun seiring dengan pertumbuhan oosit terutama pada oosit yang sudah tumbuh sempurna (fully grown oocytes) (Alm et al. 2005; Manjunatha et al. 2007). Indikasi ini kemudian dijadikan acuan oleh para peneliti dalam mengembangkan sebuah metode non invasif untuk seleksi oosit melalui pewarnaan dengan brilliant cresyl blue (BCB). Pewarna BCB dapat bereaksi dengan G6PD dan memunculkan perubahan intensitas zat warna BCB pada sitoplasma oosit sesuai dengan aktivitas intraseluler G6PD yang terjadi di dalamnya. Hal ini yang kemudian digunakan sebagai indikator untuk menyeleksi kualitas oosit.
2
Kerangka Pemikiran
Kriteria pemilihan oosit hanya berdasarkan aspek morfologi belum dapat dijadikan indikator tunggal dalam penentuan kualitas oosit sapi. Sementara itu faktor intrinsik sitoplasma yang dipercaya memiliki pengaruh signifikan terhadap kompetensi perkembangan oosit selanjutnya belum sepenuhnya teramati. Oleh karenanya dibutuhkan kombinasi antar metode yang bersifat non invasif dalam seleksi oosit guna mendapatkan oosit hasil seleksi yang lebih berkualitas. Dilaporkan selama pertumbuhannya di dalam folikel, oosit mensintesis berbagai macam protein spesifik penting dalam sitoplasmanya, termasuk enzim G6PD. Enzim ini aktif disintesis dalam sitoplasma oosit selama pertumbuhannya dan akan menurun aktivitasnya setelah oosit mencapai pertumbuhan yang sempurna. Pewarna BCB yang ditambahkan pada sekumpulan oosit dapat bereaksi dengan enzim G6PD dan memunculkan perubahan intensitas warna sejalan dengan aktivitas enzim tersebut dalam sitoplasma oosit. Oosit yang sedang tumbuh dengan aktivitas G6PD yang tinggi, akan menyebabkan pewarna BCB yang ditambahkan mudah untuk dimetabolisir, sehingga oosit menjadi tidak berwarna (BCB-). Sebaliknya, oosit yang sudah tumbuh sempurna, memiliki aktivitas G6PD yang rendah, sehingga pewarna BCB yang ditambahkan tidak mudah untuk dimetabolisir dan oosit akan menyerap zat warna biru (BCB+). Dengan melakukan teknik pewarnaan ini diharapkan kualitas oosit hasil seleksi menjadi lebih homogen dengan jumlah oosit tumbuh sempurna yang lebih banyak, sehingga diharapkan oosit tersebut memiliki kompetensi perkembangan selanjutnya yang lebih baik secara in vitro. Oleh karenanya oosit hasil seleksi dengan BCB diduga memiliki kualitas dan kompetensi perkembangan yang lebih baik dibandingkan dengan oosit yang tidak terseleksi (BCB-) atau tanpa seleksi BCB sebelumnya.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengkaji penggunaan pewarna BCB sebagai indikator aktivitas G6PD di sitoplasma oosit dalam upaya memilih oosit sapi yang lebih kompeten untuk berkembang secara in vitro.
Manfaat Penelitian
Hasil penelitian diharapkan dapat meningkatkan kualitas oosit sapi hasil seleksi sehingga produksi embrio in vitro dapat lebih efektif dan efisien.
Hipotesis Penelitian
3
2 TINJAUAN PUSTAKA
Ultrastruktur oosit sapi selama follikulogenesis
Folikulogenesis merupakan proses perkembangan folikel yang berawal dari terbentuknya folikel primordial, kemudian berkembang menjadi folikel primer, sekunder, tertier, de Graaf hingga pada akhirnya oosit tersebut akan diovulasikan. Dalam proses ini terjadi dinamika folikular yang di dalamnya termasuk pertumbuhan dan degenerasi folikel. Dinamika folikel antral atau gelombang folikel meliputi 4 proses, yaitu tahap rekruitmen, seleksi, dominan, dan atresia. Proses rekruitmen melibatkan sekelompok folikel antral yang memulai untuk tumbuh dengan sebagiannya mengalami atresia. Folikel yang bertahan dan tidak mengalami atresia merupakan folikel terseleksi yang kemudian akan terus tumbuh dan bersaing menjadi satu folikel dominan. Dalam upayanya menuju satu folikel dominan maka sebagian besar folikel terseleksi akan mengalami atresia. Pada sapi, setiap siklus estrus menghasilkan sebanyak 2-5 kali gelombang folikel. Studi yang dilakukan oleh Viana et al. (2000) terhadap 15 ekor sapi, teramati sebanyak 1 (6.67%) ekor sapi menunjukkan 2 kali gelombang folikel, 9 (60.0%) ekor sapi menunjukkan 3 kali gelombang folikel, 4 (26.67%) ekor sapi menunjukkan 4 kali gelombang folikel, dan 1 (6.67%) ekor sapi menunjukkan 5 kali gelombang folikel. Jika dalam satu kali siklus estrus menghasilkan 3 kali gelombang folikel, maka gelombang folikel pertama dan kedua terjadi saat fase luteal (metestrus dan diestrus), ketika konsentrasi hormon progesteron berada pada kadar yang tinggi di dalam tubuh. Setelah terjadi luteolisis dan gelombang folikel ketiga terjadi, maka folikulogenesis dapat berlangsung secara sempurna dan menghasilkan satu folikel dominan (Senger 2003). Oosit yang digunakan dalam proses produksi embrio in vitro diharapkan adalah oosit-oosit yang berada pada tahapan folikel terseleksi yang telah tumbuh sempurna, belum mengalami atresia dan berpeluang untuk tumbuh menjadi folikel dominan.
4
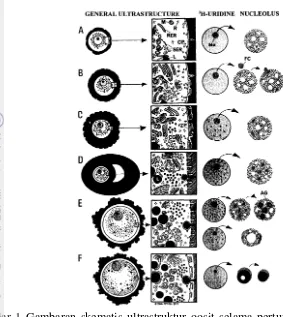
Gambar 1 Gambaran skematis ultrastruktur oosit selama pertumbuhannya di dalam folikel (Hyttel et al. 1996)
Folikel primordial memiliki diameter 34.6±3.7 µm. Oosit pada folikel ini memiliki diameter 27.9±3.3 µm yang dikelilingi oleh satu lapis sel granulosa berbentuk pipih. Secara ultrastruktur, bagian korteks oosit pada fase ini diisi oleh
coated pits dan vesikel dalam jumlah banyak yang berfungsi sebagai jalur komunikasi (endocytotic pathway) antara oosit dan sel granulosa, sedangkan bagian sitoplasma oosit ditandai oleh kehadiran beberapa organel sel seperti retikulum endoplasmik kasar (RER) dan halus (SER), badan golgi (G), serta mitokondria (M) berbentuk bulat dalam jumlah yang masih sangat sedikit di sekitar nukleus. Selain itu, bagian nukleolus (Nu) menunjukkan komponen granular yang diselingi oleh vakuol-vakuol (Gambar 1A).
5
Folikel sekunder memiliki diameter 101.7±41.8 µm. Oosit pada folikel ini memiliki diameter 45.6±14.0 µm yang dikelilingi oleh lebih dari satu lapis sel granulosa kuboid. Pada tahap ini zona pelusida (ZP) dan kortikal granula mulai terbentuk. Teramati pula adanya penurunan jumlah coated pits serta vesikel pada oolema, yang diikuti dengan pembentukan gap junction sebagai jalur komunikasi baru yang menghubungkan antara oosit dan sel granulosa. Mikrovili telah berubah bentuk menjadi tegak lurus dan FC telah secara sempurna tergabung pada bagian perifer dari nukleolus dan membentuk fibrilo-granular nukleolus. Transkripsi oosit termasuk fungsi nukleolus dan sintesis rRNA pada oosit mulai teraktivasi dan sintesis RNA tersebut akan terus dipertahankan sampai diameter oosit mencapai ±110 µm pada folikel tersier ukuran 3 mm (Gambar 1C).
Transisi dari folikel sekunder ke folikel tersier awal (pre-antral follicle) dicirikan oleh terbentuknya antrum dan peningkatan diameter folikel sampai 1 mm. Oosit pada folikel tersier awal memiliki diameter 80 µm. Pada tahap ini terjadi proliferasi sel granulosa, zona pelusida telah terbentuk dengan sempurna dan mikrovili telah tertanam di dalam zona pelusida. Selain itu, terjadi peningkatan jumlah mitokondria berbentuk panjang yang terdistribusi secara merata di seluruh bagian sitoplasma, diikuti dengan peningkatan jumlah lipid droplet, vesikel, badan golgi dan kortikal granula. Fibrillar center pada nukleolus jumlahnya meningkat dan terdistribusi secara merata (Gambar 1D). Ketika folikel tersier sampai pada diameter 3 mm, oosit yang berada di dalamnya umumnya memiliki diameter antara 80-110 µm. Jika diameter oosit <100 µm maka dicirikan oleh terjadinya pergerakan inti yang bergerak menyudut ke satu posisi, kortikal granula yang telah menduduki wilayah korteks dan susunan mikrotubul jelas terlihat di bagian sitoplasma. Oosit akan membentuk sebagian dari ruang perivitelin jika diameter oosit telah mencapai antara 100-110 µm (Gambar 1E).
Oosit dengan diameter <110 µm dilaporkan mempunyai kemampuan untuk melanjutkan proses meiosis, akan tetapi oosit ini belum memiliki kemampuan penuh untuk menyelesaikan proses meiosis sampai ke tahapan metafase II (MII). Persentase maturasi inti yang didapatkan dari oosit dengan diameter tersebut hanya mencapai 46.9% (Otoi et al. 1997). Hasil yang sama juga ditunjukkan oleh Fair et al. (1995) pada oosit berdiameter 100-110 µm dengan persentase angka maturasi inti sebesar 42%, begitupun dengan Hyttel et al (1996) yang menyatakan bahwa oosit dengan diameter ±100 µm baru memperoleh kemampuan untuk melanjutkan meiosis. Lebih lanjut dikatakan oleh Fair et al. (1995), bahwa oosit dengan diameter <110 µm mengandung tingkat 3H-uridin yang lebih tinggi jika dibandingkan dengan oosit yang memiliki diameter ≥110 µm. Kadar 3H-uridin yang tinggi mengindikasikan bahwa oosit tersebut masih berada dalam tahap pertumbuhan, karena 3H-uridin merupakan indikator yang terkait dengan aktivitas sintesis RNA pada saat oosit masih tumbuh.
6
Oosit yang terus bertumbuh dan mencapai diameter antara ≥110-120 µm dilaporkan memiliki kemampuan penuh untuk dapat menyelesaikan maturasi inti sampai ke tahap MII, dengan persentase maturasi inti yang dihasilkan adalah sebesar 75.9% (Fair et al. 1995), 76.0% (Hyttel et al. 1996) dan 75.1% (Otoi et al. 1997). Secara lebih spesifik dikatakan oleh Otoi et al. (1997), bahwa interval diameter tersebut jika dibagi lagi ke dalam dua bagian maka akan lebih jelas terlihat kompetensi perkembangannya masing-masing. Pada oosit dengan diameter antara 115-<120 µm teramati menghasilkan persentase maturasi inti yang lebih tinggi secara signifikan dibandingkan oosit dengan diameter antara 110-<115 µm (84.8% vs 64.6%). Hal ini menegaskan bahwa kemampuan untuk menyelesaikan meiosis dan mencapai tahapan MII dapat diperoleh ketika oosit berada pada diameter 115 µm. Oosit akan memiliki kompetensi penuh untuk berkembang lebih lanjut sampai ke tahap blastosis jika diameter oosit telah mencapai ≥120 µm.
Oosit yang telah tumbuh sempurna memiliki diameter ≥120 µm. Secara ultrastruktur kandungan dalam sitoplasma oosit pada diameter ini dicirikan oleh adanya mitokondria dalam jumlah banyak yang terletak di bagian perifer dan terdistribusi di seluruh bagian oosit, jumlah lipid droplet, vesikel dan badan golgi juga mengalami peningkatan. Selain itu pembentukan ruang perivitellin telah berlangsung dengan sempurna. Secara keseluruhan selama proses pertumbuhannya di dalam folikel, oosit sapi mengalami berbagai perubahan penting pada sitoplasma oositnya, yang ditunjukkan dengan terjadinya relokasi dan modulasi dari sejumlah organel sel dan pengembangan struktur tertentu dari oosit seperti zona pelusida serta kortikal granul. Selain itu adanya proses transkripsi yang aktif juga terjadi di dalamnya. Hal tersebut dibutuhkan agar oosit dapat memiliki kompetensi perkembangan meiotik. Dalam proses produksi embrio in vitro, seleksi ukuran folikel pada saat aspirasi oosit sebaiknya dilakukan pada folikel berdiameter >3mm, karena diharapkan ukuran oosit yang berada di dalamnya telah mencapai >110 µm (Fair et al. 1996; Hyttel et al. 1996; Gordon 2003).
Enzim glukosa 6 fosfat dehidrogenase, kompetensi perkembangan oosit, dan aplikasi pewarna brilliant cresyl blue
Seiring pertumbuhannya di dalam folikel, oosit mensintesis berbagai macam protein spesifik penting di dalam sitoplasmanya, termasuk enzim glukosa 6 fosfat dehidrogenase (G6PD). Enzim ini berperan sangat penting dalam proses metabolisme karbohidrat melalui jalur fosfat pentosa. Pada jalur ini tidak dihasilkan adenosine triphosphate (ATP) seperti halnya pada proses glikolisis, melainkan berfungsi dalam menghasilkan ribosa 5 fosfat dan nikotinamida adenin dinukleotida fosfat (NADPH) (Marks et al. 1996).
7
mereduksi glutation teroksidasi (GSSG) menjadi glutation tereduksi (GSH) melalui kerja dari enzim glutation reduktase. Komponen GSH yang dihasilkan dari reaksi ini kemudian berfungsi untuk menghilangkan senyawa H2O2 dari sel melalui reaksi yang
dikatalisis oleh enzim glutation peroksidase. Glutation peroksidase akan mempercepat reduksi hidrogen peroksidase dan lipid peroksida dengan kehadiran GSH, yang selanjutnya akan dikonversi menjadi GSSG. Reaksi ini terus berlanjut dan berulang selama proses pertumbuhan sel dengan tujuan untuk mengurangi terjadinya stres oksidatif dan mencegah penurunan fungsi normal suatu sel sampai sel tersebut telah tumbuh sempurna. Penurunan aktivitas metabolisme pada sel yang telah tumbuh sempurna dicirikan oleh penurunan aktivitas G6PD yang menandakan sintesis RNA telah cukup memadai di dalam sitoplasma sel dan kandungan antioksidan intraseluler yang diproduksi setelahnya berfungsi untuk membantu perkembangan oosit selanjutnya (Luberda 2005). Secara skematis jalur fosfat pentosa dapat dilihat pada Gambar 2.
Gambar 2 Metabolisme karbohidrat melalui jalur fosfat pentosa (Marks et al. 1996)
8
antioksidan intraseluler. Aktivitas G6PD yang ditemukan tinggi pada oosit yang sedang tumbuh berkaitan dengan dihasilkannya NADPH dalam jumlah banyak, sehingga dapat menghasilkan senyawa glutation yang berperan sebagai antioksidan intraseluler guna menekan terjadinya stres oksidatif pada oosit. Jika suatu sel mengalami penurunan level dari G6PD pada saat proses pertumbuhannya, maka sel tersebut akan sangat sensitif terhadap terjadinya stres oksidatif (Berg et al. 2002). Selain itu, aktivitas G6PD yang tinggi pada oosit yang sedang tumbuh juga berkaitan dengan dihasilkannya ribosa 5 fosfat dalam jumlah banyak. Senyawa ini dibutuhkan untuk sintesis nukleotida dan asam nukleat yang merupakan komponen penyusun utama dari DNA dan RNA. Selama pertumbuhannya di dalam folikel dibutuhkan sintesis RNA yang cukup pada sitoplasma oosit agar oosit memiliki kompetensi perkembangan meiotik. Torner et al. (2008) mengatakan bahwa oosit yang sedang tumbuh cenderung lebih aktif dalam melakukan proses transkripsi dan translasi, yang menghasilkan RNA dan protein penting untuk proses perkembangan selanjutnya. Sintesis RNA terjadi di nuklelolus pada saat oosit berada di fase folikel sekunder. Proses ini terus berlanjut hingga oosit mencapai diameter 110 µ m dan setelahnya fungsi nukleolus mengalami inaktivasi (Fair et al. 1995; Hyttel et al. 1996). Aktivitas G6PD yang cenderung menurun pada oosit yang telah menyelesaikan fase pertumbuhannya mengindikasikan bahwa sintesis RNA dan aktivitas metabolisme pada oosit tersebut telah mencukupi untuk perkembangan awal oosit dan embrio.
Aktivitas G6PD sebagai indikator kualitas oosit dapat ditentukan dengan aplikasi pewarna brilliant cresyl blue (BCB). Pewarna ini memiliki komponen zat biokimia tentang mekanisme yang terjadi pada metabolisme BCB oleh G6PD belum sepenuhnya dapat dipahami. Alm et al. (2005) dalam studinya menyatakan bahwa BCB memiliki kemampuan untuk berperan sebagai akseptor elektron, dengan cara ini sitoplasma oosit menjadi tidak berwarna selama aliran elektron terus diinduksi oleh G6PD. Elektron penginduksi tersebut dihasilkan dari kerja G6PD sebagai katalisator dalam proses oksidasi glukosa 6 fosfat dan reduksi NADP+.
Maturasi oosit in vitro dan perolehan kompetensinya
9
Gambar 3 Skema distribusi organel sitoplasma selama pematangan dan fertilisasi. A Pematangan inti dan pergerakan organel sitoplasma dari tahap germinal vesicle (GV) ke tahap metafase II dan pembentukan zigot. B Distribusi organel dan mekanisme pelepasan isi kortikal granul dan Ca2+ intraseluler, setelah penetrasi spermatozoa ke dalam oosit (Ferreira et al. 2009)
10
Pematangan sitoplasma yang terjadi selama proses maturasi meliputi organisasi sitoskeletal dari oosit seperti migrasi kortikal granula ke oolemma, peningkatan mitokondria dan lipid droplet, akumulasi protein dan mRNA, perubahan susunan aparatus golgi, keberadaan retikulum endoplasmik granular, perubahan aktivitas maturation promoting factor (MPF) dan mitogen activated protein kinase (MAPK) (Hyttel et al. 1996; Sirard et al. 2006).
Fertilisasi oosit in vitro
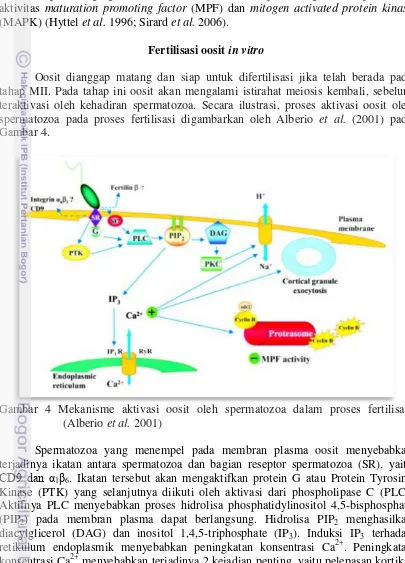
Oosit dianggap matang dan siap untuk difertilisasi jika telah berada pada tahap MII. Pada tahap ini oosit akan mengalami istirahat meiosis kembali, sebelum teraktivasi oleh kehadiran spermatozoa. Secara ilustrasi, proses aktivasi oosit oleh spermatozoa pada proses fertilisasi digambarkan oleh Alberio et al. (2001) pada Gambar 4.
Gambar 4 Mekanisme aktivasi oosit oleh spermatozoa dalam proses fertilisasi (Alberio et al. 2001)
Spermatozoa yang menempel pada membran plasma oosit menyebabkan terjadinya ikatan antara spermatozoa dan bagian reseptor spermatozoa (SR), yaitu
CD9 dan α1β6. Ikatan tersebut akan mengaktifkan protein G atau Protein Tyrosine
Kinase (PTK) yang selanjutnya diikuti oleh aktivasi dari phospholipase C (PLC). Aktifnya PLC menyebabkan proses hidrolisa phosphatidylinositol 4,5-bisphosphate (PIP2) pada membran plasma dapat berlangsung. Hidrolisa PIP2 menghasilkan
diacylglicerol (DAG) dan inositol 1,4,5-triphosphate (IP3). Induksi IP3 terhadap
11
granula dan inisiasi berlanjutnya siklus pembelahan meiosis sel. Pelepasan kortikal granula ke dalam ruang perivitellin menyebabkan pengerasan pada zona pelusida untuk mencegah terjadinya polispermia dan inisiasi berlanjutnya siklus pembelahan meiosis sel yang ditandai dengan menurunnya aktivitas MPF (Alberio et al. 2001; Jones 2007). Peningkatan Ca2+ intraseluler juga akan diikuti dengan meningkatnya pH, menyebabkan terjadinya pertukaran ion Na+ dari luar dan H+ dari dalam oosit. Peningkatan pH penting untuk menstimulasi replikasi DNA, perkembangan konduktan K+ dan perubahan metabolisme seluler lainnya (Machaty dan Prather 1998).
Fertilisasi melibatkan aktivasi oosit oleh spermatozoa, sehingga terjadi pembentukan pronukleus dan menjadi zigot. Transformasi kepala spermatozoa (inti), yaitu dari morfologi dan aktivitas sintetisnya, melibatkan pecahnya selubung inti dan dekondensasi kromatin dalam sitoplasma. Dekompaksi inti spermatozoa melibatkan reduksi rantai disulfida dari protamin. Protamin digantikan oleh histon dan non-histon selama pembentukan pronukleus jantan sebagai persiapan untuk sintesis DNA. Perubahan morfologi dari spermatozoa setelah penetrasi kedalam oosit dan bertransformasi menjadi pronkleus jantan adalah: dekondensasi inti spermatozoa (1), rekondensasi inti spermatozoa (2), redekondensasi inti spermatozoa (3), prepronukleus (4), dan pronukleus jantan (5). Dalam proses in vitro, penetrasi spermatozoa ke dalam oosit pertama kali teramati pada 4 jam post inseminasi dan mencapai puncaknya pada 9 jam post inseminasi. Pembentukan pronukleus jantan teramati pada 9 jam post inseminasi dan mencapai angka maksimumnya pada 14 jam
12
3 METODE
Waktu dan Tempat Penelitian
Ovarium diperoleh dari Rumah Potong Hewan Cibinong, Kabupaten Bogor. Penelitian dilaksanakan di Laboratorium Fertilisasi in vitro, Bagian Reproduksi dan Kebidanan, Departemen Klinik Reproduksi dan Patologi, Fakultas Kedokteran Hewan, Institut Pertanian Bogor. Penelitian ini berlangsung dari bulan Februari sampai dengan Mei 2014.
Metode Penelitian
Tahap I. Kemampuan maturasi inti oosit setelah dipaparkan pewarna BCB
Koleksi dan klasifikasi oosit
Oosit diperoleh dari ovarium sapi yang dipotong di rumah potong hewan (RPH) dan dibawa ke laboratorium dalam kantung plastik yang berisi larutan NaCl fisiologis (0.9 w/v) suhu 27-28°C. Dikarenakan lokasi RPH yang cukup jauh dan waktu pemotongan hewan, oosit tiba di laboratorium ±6-8 jam kemudian. Koleksi oosit dilakukan dengan teknik aspirasi menggunakan syringe dan jarum ukuran 18G. Cairan hasil aspirasi kemudian dimasukkan ke dalam tabung sentrifus steril 15 mL (Corning Centristar®) dan dibiarkan mengendap selama 3-5 menit. Setelah terbentuk endapan, supernatan kemudian dibuang dan sisa endapan dituang pada cawan petri steril yang mengandung larutan mPBS (PBS plus 10% fetal bovine serum) (Sigma, USA).
Seleksi oosit dilakukan di bawah mikroskop stereo berdasarkan homogenitas sitoplasma dan kekompakan sel-sel kumulus. Oosit hasil seleksi kemudian dibagi menjadi 2 kelompok, yaitu kelompok kontrol dan kelompok pewarnaan BCB. Oosit kelompok kontrol dilakukan maturasi secara langsung tanpa dilakukan pewarnaan BCB sebelumnya.
Pewarnaan brilliant cresyl blue (BCB)
Oosit kelompok pewarnaan BCB diinkubasi dalam pewarna BCB 26 µM (B-5388, Sigma) selama 90 menit pada inkubator 5% CO2 dan suhu 39°C (Pujol et al.
13
Maturasi oosit in vitro (IVM)
Masing-masing oosit dari kelompok kontrol, BCB+, dan BCB- dimatangkan secara terpisah pada media maturasi dalam bentuk drop 100 µL yang berisi 10-20 oosit dan ditutup dengan mineral oil (Sigma, USA). Medium maturasi terdiri dari: Tissue Culture Medium-199/TCM-199 (Gibco, USA) yang disuplementasi dengan 10 IU/mL Pregnant Mare Serum Gonadothropine/PMSG (Kyoritsu Seiyaku, Japan), 10 IU/mL Human Chorionic Gonadotropin/hCG (Kyoritsu Seiyaku, Japan), 50 µg/mL Gentamycin (Sigma, USA), dan 10% fetal bovine serum/FBS (Sigma, USA). Maturasi oosit dilakukan dalam inkubator 5% CO2 pada suhu 39°C selama 24 jam.
Evaluasi kemampuan maturasi oosit in vitro
Oosit didenudasi sel-sel kumulusnya menggunakan enzim hyaluronidase 0.25% (Sigma, USA) dengan cara dipipet berulang-ulang menggunakan ujung pipet yang sesuai dengan ukuran oosit. Oosit hasil denudasi kemudian diletakkan pada drop mPBS diatas gelas objek yang memiliki bantalan campuran paraffin dan vaseline di kedua sisinya. Drop ditutup cover glass dan ditekan secara perlahan menggunakan pensil pada kedua sisi bantalan paraffin dan vaseline. Setelah cover glass menutup dengan sempurna, keempat sisinya direkatkan dengan bantuan lem untuk menghindari terlepas pada saat fiksasi. Preparat difiksasi selama 48-72 jam pada kombinasi larutan Etanol : asam asetat (3:1 v/v) dan setelahnya diwarnai dengan aceto orcein 2% selama 2-3 menit melalui celah diantara kedua sisinya. Zat pewarna kemudian dibilas dengan larutan asam asetat 25%. Pengerjaan dilakukan di bawah mikroskop untuk menghindari adanya oosit yang ikut terbawa larutan pada saat pewarnaan dan pembilasan. Selanjutnya setiap sisi cover glass diberi larutan kuteks bening dan dilakukan pengamatan morfologi di bawah mikroskop menggunakan lensa dengan fitur bright field (Olympus IX 70, Japan).
14
dengan jumlah total oosit yang dimaturasi. Persentase jumlah oosit yang mampu mencapai tahap metafase II dan tahapan meiosis lainnya (germinal vesicle, germinal vecicle breakdown (diakinesis), metafase I, anafase I, dan telofase I) dibandingkan diantara perlakuan dan kontrol.
Tahap II. Kemampuan fertilisasi oosit setelah dipaparkan pewarna BCB
Fertilisasi oosit in vitro (IVF)
Oosit dimaturasi seperti prosedur sebelumnya yang dilakukan pada penelitian tahap I. Fertilisasi oosit dilakukan menggunakan semen beku sapi yang berasal dari Balai Besar Inseminasi Buatan (BBIB) Lembang, Bandung. Proses thawing dilakukan dengan menempatkan straw dalam air bersuhu 37°C selama 30 detik. Selanjutnya semen dimasukkan ke dalam tabung sentrifus steril 15 mL (Corning Centristar®) yang berisi 4 mL medium fertilisasi (Suzuki et al. 2000) dan disentrifugasi pada kecepatan 700 g suhu 28°C selama 8 menit. Setelah sentrifugasi, bagian supernatan dibuang dan sisa spermatozoa sebagai endapannya dilakukan pengenceran menggunakan medium fertilisasi hingga mencapai konsentrasi akhir 2x106 spermatozoa/mL.
Spermatozoa yang telah disiapkan dibuat dalam bentuk drop pada petri dish (Nunclon, Denmark) masing-masing sebanyak 100 µL untuk 10-20 oosit dan ditutup dengan mineral oil (Sigma, USA). Masing-masing kelompok oosit (BCB+, BCB-, dan kontrol) yang telah dimaturasi dicuci sebanyak 2 kali dalam medium fertilisasi dan kemudian dipindahkan ke dalam drop spermatozoa untuk proses fertilisasi. Inkubasi oosit dilakukan selama 14 jam pada inkubator 5% CO2 suhu 39°C.
Evaluasi kemampuan fertilisasi oosit in vitro
Tingkat fertilisasi dievaluasi berdasarkan pada pembentukan dan jumLah pronukleus setelah dilakukan pewarnaan dengan aceto orcein 2%. Metode pewarnaan menggunakan metode yang sama seperti yang dilakukan pada evaluasi tahap maturasi oosit. Oosit terfertilisasi ditandai dengan terbentuknya 2 atau lebih pronukleus (PN) (Wongsrikeao 2006) dan tingkat fertilisasi merupakan perbandingan antara jumlah oosit yang terfertilisasi dibandingkan jumlah keseluruhan oosit yang difertilisasi. Analisis data
16
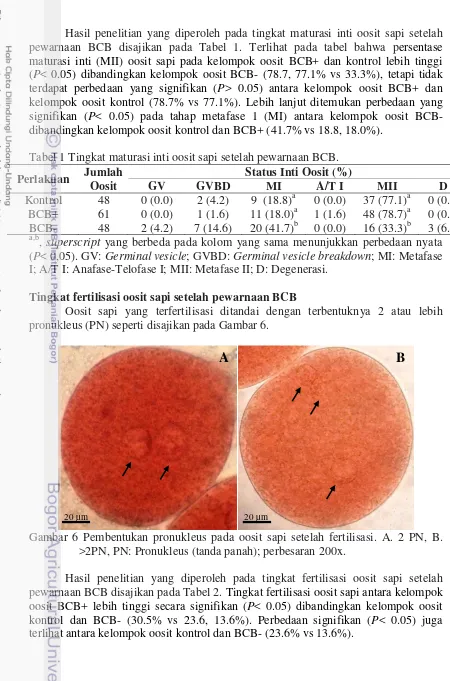
Hasil penelitian yang diperoleh pada tingkat maturasi inti oosit sapi setelah pewarnaan BCB disajikan pada Tabel 1. Terlihat pada tabel bahwa persentase maturasi inti (MII) oosit sapi pada kelompok oosit BCB+ dan kontrol lebih tinggi (P< 0.05) dibandingkan kelompok oosit BCB- (78.7, 77.1% vs 33.3%), tetapi tidak terdapat perbedaan yang signifikan (P> 0.05) antara kelompok oosit BCB+ dan kelompok oosit kontrol (78.7% vs 77.1%). Lebih lanjut ditemukan perbedaan yang signifikan (P< 0.05) pada tahap metafase 1 (MI) antara kelompok oosit BCB- dibandingkan kelompok oosit kontrol dan BCB+ (41.7% vs 18.8, 18.0%).
Tabel 1 Tingkat maturasi inti oosit sapi setelah pewarnaan BCB.
Perlakuan Jumlah Oosit
Status Inti Oosit (%)
GV GVBD MI A/T I MII D
Kontrol 48 0 (0.0) 2 (4.2) 9 (18.8)a 0 (0.0) 37 (77.1)a 0 (0.0) BCB+ 61 0 (0.0) 1 (1.6) 11 (18.0)a 1 (1.6) 48 (78.7)a 0 (0.0) BCB- 48 2 (4.2) 7 (14.6) 20 (41.7)b 0 (0.0) 16 (33.3)b 3 (6.3) a,b
, superscript yang berbeda pada kolom yang sama menunjukkan perbedaan nyata (P< 0,05). GV: Germinal vesicle; GVBD: Germinal vesicle breakdown; MI: Metafase I; A/T I: Anafase-Telofase I; MII: Metafase II; D: Degenerasi.
Tingkat fertilisasi oosit sapi setelah pewarnaan BCB
Oosit sapi yang terfertilisasi ditandai dengan terbentuknya 2 atau lebih pronukleus (PN) seperti disajikan pada Gambar 6.
Gambar 6 Pembentukan pronukleus pada oosit sapi setelah fertilisasi. A. 2 PN, B. >2PN, PN: Pronukleus (tanda panah); perbesaran 200x.
Hasil penelitian yang diperoleh pada tingkat fertilisasi oosit sapi setelah pewarnaan BCB disajikan pada Tabel 2. Tingkat fertilisasi oosit sapi antara kelompok oosit BCB+ lebih tinggi secara signifikan (P< 0.05) dibandingkan kelompok oosit kontrol dan BCB- (30.5% vs 23.6, 13.6%). Perbedaan signifikan (P< 0.05) juga terlihat antara kelompok oosit kontrol dan BCB- (23.6% vs 13.6%).
A
B
17
Tabel 2 Tingkat fertilisasi oosit sapi setelah pewarnaan BCB.
Perlakuan Jumlah (P< 0.05). Normal: membentuk 2 Pronukleus (PN); Polispermia: membentuk >2 PN;
Tingkat fertilisasi: jumlah oosit yang dapat membentuk 2 atau lebih PN dari keseluruhan jumlah oosit yang difertilisasi.
PEMBAHASAN
Tingkat maturasi inti oosit
Tingkat pematangan inti oosit sangat dipengaruhi oleh kualitas oosit yang digunakan. Pada penelitian ini oosit sapi dipilih secara morfologi berdasarkan homogenitas sitoplasma dan kekompakan sel-sel kumulus, sehingga oosit yang digunakan diasumsikan seragam dan mempunyai kompetensi perkembangan yang sama. Untuk mendapatkan kualitas oosit yang lebih homogen, seleksi dilakukan dengan teknik pewarnaan BCB berdasarkan aktivitas enzim G6PD dalam sitoplasma oosit. Dikarenakan hanya oosit yang telah matang (MII) yang memiliki kemampuan untuk dibuahi oleh spermatozoa dan berkembang lebih lanjut membentuk embrio, maka pada produksi embrio in vitro, oosit perlu dimatangkan terlebih dahulu. Oleh karenanya tingkat maturasi inti oosit sering dijadikan sebagai salah satu parameter dan indikator kuat untuk mengamati tahap awal kompetensi perkembangan oosit.
18
sehingga terjadi ketidakcukupan kondisi sitoplasmik yang akan mempengaruhi kemampuan perkembangan oosit selanjutnya. Defisiensi sitoplasmik menurut Salamone et al. (2001), dicirikan oleh aktivitas yang rendah dari 2 kinase, yaitu
maturation promoting factor (MPF) dan mitogen activated protein kinase (MAPK). Selain itu, jumlah relatif dari IP3R (reseptor dari inositol 1,4,5-triphospate) sebagai mediator dari osilasi kalsium secara substansial juga lebih rendah. Penyebab lainnya, kemungkinan oosit tersebut berada pada tahap awal terjadinya degenerasi, karena oosit yang telah terseleksi secara morfologi dalam kriteria baik tetapi dikategorikan sebagai kelompok oosit BCB-, teramati 6.3% mengalami degenerasi. Hal ini bertentangan dengan yang telah dilaporkan oleh Rodriguez-Gonzalez et al. (2002) dan Tiffin et al. (1991) bahwa pewarnaan BCB tidak berpengaruh terhadap viabilitas oosit dan tidak menimbulkan efek lethal pada embrio. Namun, dikatakan oleh Katska-Ksiazkiewic et al. (2007) bahwa beberapa oosit dengan perubahan apoptotik yang mungkin telah terjadi, dapat menjadi lebih sensitif terhadap pewarnaan dengan BCB. Dalam laporan lainnya oleh Opiela et al. (2008) dinyatakan bahwa oosit yang terpapar BCB memiliki kecenderungan untuk terjadinya apoptosis, meskipun secara statistika dari hasil analisa protein tidak meneguhkan kejadian apoptosis tersebut. Diketahui pula dalam proses pertumbuhan folikel pada sapi yang dimulai dari tahap rekruitmen, seleksi, dan dominan dengan hanya didapatkan satu folikel dominan dan sebagian besar folikel lainnya akan mengalami atresia (Senger 2003).
Hasil penelitian lainnya menunjukkan terdapat perbedaan signifikan (P< 0.05) dalam persentase oosit yang mencapai tahap MII antara kelompok oosit BCB+ dan kontrol terhadap kelompok oosit BCB- (78.7%, 77.1% vs 33.3%). Data ini sejalan dengan hasil penelitian yang dilakukan pada kambing (Rodriguez-Gonzalez et al.
2002), sapi (Alm et al. 2005), kerbau (Manjunantha et al. 2007), babi (Isizhaki et al.
2009) dan domba (Setiadi dan Supriatna 2010) bahwa prosedur seleksi oosit dengan menggunakan teknik pewarnaan BCB sebelum pematangan mampu menyeleksi oosit yang kompeten (BCB+) dan tidak kompeten (BCB-) untuk berkembang. Lebih lanjut dinyatakan bahwa sebagian besar oosit kelompok BCB+ telah menyelesaikan fase pertumbuhannya di dalam folikel yang ditandai oleh kemampuannya dalam mencapai maturasi inti (MII).
19
Lebih lanjut Karami-Shabankareh et al. (2012) menemukan adanya hubungan positif antara oosit yang terseleksi sebagai BCB+ dalam hubungannya dengan diameter oosit dan diameter folikel oosit berasal yang lebih besar. Diameter oosit akan sangat berpengaruh dan menentukan terhadap kompetensi perkembangan oosit selanjutnya. Studi lainnya oleh Castaneda et al. (2013) dilaporkan bahwa oosit BCB+ mengandung lebih banyak sitoplasmik lipid dibandingkan oosit BCB-. Lipid yang tersimpan merupakan sumber energi bagi oosit selama serangkaian proses in vitro
yang dimulai dari maturasi, fertilisasi, dan permulaan pembelahan, sehingga oosit BCB+ memiliki keuntungan dalam penyimpanan energi yang lebih banyak dibandingkan oosit BCB-. Hal ini dikatakan menjadi salah satu sebab yang menjadikan oosit BCB+ memiliki kemampuan perkembangan yang lebih baik dibandingkan oosit BCB-. Sejalan dengan studi ini, Sierard et al. (2006) melaporkan bahwa akumulasi lipid dalam sitoplasma oosit memiliki peranan yang sama pentingnya dengan akumulasi protein dan mRNA untuk mendukung perkembangan awal oosit. Dikatakan oleh Torner et al. (2008), oosit yang bertumbuh cenderung lebih aktif dalam melakukan proses transkripsi dan translasi, yang menghasilkan RNA dan protein penting untuk proses perkembangan selanjutnya. Oosit BCB+ telah menyelesaikan sintesis RNA dan akumulasi komponen sitoplasma yang cukup yang dibutuhkan untuk perkembangan awal oosit dan embrio. Sedangkan oosit BCB-, dengan aktivitas enzim G6PD yang tinggi mengindikasikan adanya ketidakcukupan maturasi sitoplasmik.
Tingkat Fertilisasi Oosit
Oosit yang telah terfertilisasi dicirikan dengan terbentuknya 2 atau lebih pronukleus. Hasil penelitian ini menunjukkan tingkat fertilisasi yang lebih tinggi secara signifkan (P< 0,05) antara kelompok oosit BCB+ dibandingkan kelompok oosit kontrol dan BCB- (30.5% vs 23.6%, 13.6%). Data ini sejalan dengan hasil penelitian yang dilakukan pada kambing prepuber (Rodriguez-Gonzalez et al. 2003) dan kerbau (Manjunatha et al. 2007).
Rendahnya tingkat fertilisasi pada kelompok oosit BCB- sebagai salah satu sebab dari belum sempurnanya maturasi sitoplasmik sejalan dengan rendahnya angka maturasi inti yang dihasilkan jika dibandingkan dengan kelompok oosit BCB+. Pada penelitian sebelumnya yang telah dilakukan pada babi (Roca et al. 1998), sapi dara (Pujol et al. 2000; Pujol et al. 2004), kambing pre-puber (Rodriguez-Gonzalez et al.
20
yang terjadi pada bagian inti dan sitoplasma selama proses maturasi oosit diduga juga sangat berpengaruh terhadap kejadian fertilisasi. Oosit yang berhasil mencapai maturasi inti (MII), namun tidak diiringi dengan sempurnanya maturasi sitoplasmik dapat menyebabkan kegagalan dalam fertilisasi (Kishida et al. 2004). Isizhaki et al.
(2009) mengatakan bahwa seleksi oosit dengan BCB sebelum maturasi dapat digunakan sebagai penanda terhadap kemampuan maturasi inti dan sitoplasmiknya. Oosit BCB+ dengan aktivitas G6PD yang rendah cenderung lebih dapat menyelesaikan maturasi inti dan sitoplasmik dibandingkan oosit BCB-.
Spikings et al. (2007) dalam studinya menambahkan bahwa oosit BCB- mengalami keterlambatan dalam onset ekspresi protein dibandingkan oosit BCB+. Perbedaan signifikan dalam ekspresi inti yang mengkode replikasi protein antara oosit BCB+ dan BCB- mengakibatkan terjadinya keterlambatan pada replikasi mitokondrial DNA. Adanya keterlambatan pada replikasi mitokondrial DNA berkorelasi dengan menurunnya kejadian fertilisasi dan perkembangan embrionik pada oosit BCB-. Lebih lanjut Castaneda et al. (2013), menyatakan bahwa akumulasi lipid yang rendah pada oosit BCB- berkaitan dengan sumber energi yang tersimpan tidak mencukupi untuk proses awal perkembangannya, terutama kaitannya dengan fertilisasi. Fertilisasi merupakan sebuah proses dengan konsumsi energi yang tinggi sehingga cadangan lipid yang ada sangat memberikan keuntungan dalam terjadinya proses ini. Selain itu, oosit BCB- dengan defisiensi sitoplasmik yang terjadi menyebabkan kemampuan untuk berkembang setelah fertilisasi dan membentuk pronukleus juga menjadi berkurang. Hal berbeda ditemukan pada kelompok oosit BCB+ dengan tingkat fertilisasi yang lebih baik.
21
5 SIMPULAN DAN SARAN
Simpulan
Dari hasil penelitian yang diperoleh dapat disimpulkan bahwa seleksi kompetensi oosit menggunakan pewarna BCB efektif untuk dapat menyeleksi oosit dengan kemampuan maturasi dan fertilisasi yang lebih baik.
Saran
22
DAFTAR PUSTAKA
Alberio R, Zakhartchenko V, Motlik J, Wolf E. 2001. Mammalian oocyte activation: lessons from the sperm and implication for nuclear transfer. Int J Dev Biol 45:797-809
Alm H, Torner H, Lohrke B, Viequtz T, Ghoneim IM, Kanitz W. 2005. Bovine blastocyst development rate in vitro is influenced by selection of oocytes by brillant cresyl blue staining before IVM as an indicator for glucose 6 phosphate dehydrogenase activity. Theriogenology 63(8): 2194-2205
Bhojwani S, Alm H, Torner H, Kanitz W, Poehland R. 2007. Selection developmentally competent oocytes through brilliant cresyl blue stain enhances blastocyst development rate after bovine nuclear transfer. Theriogenology 67: 341-345
Castaneda CA, Kaye P, Pantaleon M, Philips N, Norman S, Fry R, D’Occhio MJ.
2013. Lipid content, active mitochondria and brilliant cresyl blue staining in bovine oocytes. Theriogenology 79: 417-422
Covarrubias L, Hernandez-Garcia Dm Schnabel D, Salas-Vidal E, Castro-Obregon S. 2008. Function of reactive oxygen species during animal development: passive or active. Dev Biol 320: 1-11
Ducibella T. 2002. Egg to embryo transition is driven by differential responses to Ca2+ oscillation number. Reprod Fertil Dev 8: 485-489
Ericsson SA, Boice ML, Funahashi H, Day BN. 1993. Assesment of porcine oocytes using brilliant cresyl blue. Theriogenology 39: 214
Fair T, Hyttel P, Greve T. 1995. Bovine oocyte diameter in relation to maturational competence and transcriptional activity. Mol Reprod Dev 42: 437-442
Fair T, Hulshof SC, Hyttel P, Greve T, Boland M. 1997. Oocytes ultrastructure in bovine primordial to early tertiary follicles. Anat embryo 195(4): 327-336 Gordon I. 2003. Laboratory of Cattle Production: 2nd edition. London (GB): CABI
publishing.
Ginther OJ, Bergfelt DR, Kulick LJ, Kot K. 1999. Selection of the dominant follicle in cattle: establishment of follicle deviation in less than 8h through depression of FSH concentrations. Theriogenology 52: 1079-1093
Greer JP, Foerster J, Rodgers GM, Paraskevas F, Glader B, Arber DA, Means RT. 2009. Wintrobes’s Clinical Hematology: Volume 1. Philadelphia (USA): Lippincot Williams & Wilkins
Gupta S, Malhotra N, Sharma D, Chandra A, Agarwal A. 2009. Oxidative stress and its role in female infertility and assisted reproduction: Clinical implication. Int J Fertil Steril 2(4): 147-164
Hafez B, Hafez ESE. 2000. Reproduction in Farm Animals: 7th edition. USA (US): Lippincott Williams & Wilkins
Hyttel P, Fair T, Callesen H, Greve T. 1996. Oocyte growth capacitation and final maturation in cattle. Theriogenology 47(1): 23-32
23
Jortzik E, Mailu BM, Preuss J, Fischer M, Bode L, Rahlfs S, Becker K. 2011. Glucose 6 phospate dehydrogenase 6 phosphogluconalactonase: a unique bifunctional enzyme from plasmodium falciparum. J Biochem (436): 641-650
Jones KT. 2007. Intracellular calcium in the fertilization and development of mammalian eggs. Proc Aust Phys Soc. 38:35-41
Karami-Shabankareh H, Mirshamsi SM. 2012. Selection developmentally competent sheep oocytes using the brilliant cresyl blue test and the relationship to follicle size and oocyte diameter. Small Rum Res 105: 244-249
Katska-Ksiazkiewicz L, Opiela J, Rynska B. 2007. Effects of oocyte quality, semen donor and embryo co-culture system on the efficiency of blastocyst prodction in goats. Theriogenology 68: 736-744
Kishida R, Lee ES, Fukui Y. 2004. In vitro maturation of porcine oocytes using a defined medium and developmental capacity after intracytoplasmic sperm injection. Theriogenology 62: 1663-1676
Lehninger AL, Nelson DL, Cox MM. 1993. Principles of biochemistry: second edition. New York (US): Worth Public Inc.
Lequarre SA. 2005. Influence of antral follicle size on oocyte characteristics and embryo development in the bovine. Theriogenology 63: 841-859
Luberda Z. 2005. The role of glutathione in mammalian gametes. Biol Reprod 5(1): 5-17
Machaty Z, Prather RS. 1998. Strategies for activating nuclear transfer oocytes. Reprod Fertil Dev 10: 599-613
Manjunatha BM, Gupta PS, Devaraj M, Ravindra JP, Nadi S. 2007. Selection of developmentally competent buffalo oocytes by brilliant cresyl blue staining before IVM. Theriogenology 68(9): 1299-1304
Marks DB, Marks AS, Smith CM. 1996. Biokimia Kedokteran Dasar: Sebuah Pendekatan Klinis. EGC (US): Williams and Wilkins
Opiela J, Katska-Ksiazkiewizt L, Lipinski D, Slomski R, Bzowska M, Rynska B. 2008. Interaction among activity of glucose 6 phosphat dehydrogenase in immature oocytes, expression of apoptosis related genes bcl-2 and bax and developmental competence following ivp in cattle. Theriogenology 69(5): 546–555
Pereira GR, Lorenzo PL, Carneiro GF, Bilodeau-Goeseels S, Kastelic JP, Esteller-Vico A, Liu IKM. 2010. Selection of developmentally competent immature equine oocytes by brilliant cresyl blue staining prior to maturation with equine growth hormone in vitro. Anim Reprod Sci 121: 248-249
Pujol M, Lopez-Bejar M, Mertens MJ, Rodriguez Gonzales E, Velila E, Paramio MT. 2000. Selection of immature oocytes using the brilliant cresyl blue test. Theriogenology 53: 466
Pujol M, Lopez-Bejar M, Paramio MT. 2004. Developmental competence of heifer oocytes selected using the brilliant cresyl blue test. Theriogenology 61: 735-744
24
Rodriguez-Gonzalez E, Lopez-Bejar M, Velilla E, Paramio MT. 2002. Selection of prepubertal goat oocytes using the brilliant cresyl blue test. Theriogenology 57: 1397-1409
Rodriguez-Gonzalez E, Lopez-Bejar M, Izquierdo D, Paramio MT. 2003. Developmental competence of prepubertal goat oocyte selected with brilliant cresyl blue and matured with cyteamine supplementation. Reprod Nutr Dev 43: 179-187
Salamone DF, Damiani P, Fissore RA, Robl JM, Duby RT. 2001. Biochemical and developmental evidence that ooplasmic maturation of prepubertal bovine oocytes is comprimised. Biol Reprod 64: 1761-1768
Senger PL. 2003. Pathways to Pregnancy and Parturition. 2nd edition. Washington (US): Current Conceptions Inc.
Setiadi MA, Supriatna I. 2010. Seleksi kemampuan pematangan oosit domba menggunakan teknik brilliant cresyl blue. J Vet 11(4): 251-256
Sierard, MA, Richard F, Blondin P, Robert C. 2006. Contribution of the oocytes to embryo quality. Theriogenology 65: 126-136
Smith LC, Alciver AA. 1993. Cytoplasmic inheritance and its effects on development and performance. J Reprod Fert 48: 31-43.
Spikings EC, Alderson J, S.t. John JC. 2007. Regulated mitochondrial DNA replication during oocyte maturation is essential for successful porcine embryonic development. Biol Reprod 76: 327-335
Steel RGD, Torrie JH. 1991. Prinsip dan Prosedur Statistika: Suatu Pendekatan Biometrik. Jakarta (ID): PT. Gramedia Pustaka Utama
Suzuki K, Eriksson B, Shimizu H, Nagai T, Rodriguez-Martinez H. 2000. Effect of hyaluronan on monospermic penetration of porcine oocytes fertilized in vitro. Int J Androl 23: 13-21
Tabandeh MR, Golestani N, Kafi M, Hosseini A, Saeb M, Sarkoohi P. 2012. Gene expression of adinopectin and adinopectin receptors in dominant and atretic follicels and oocytes screened based on brilliant cresyl blue staining. Anim Reprod Sci 131: 30-40
Tiffin GJ, Rieger D, Betteridge KJ, Yadav BR, King WA. 1991. Glucose and glutamine metabolism in pre-attachment cattle embryos in relation to sex and stage development. J Reprod Fertil 93: 125-132
Torner H, Ghanem N, Ambros C, Holker M, Tornek W, Phatsara C, Alm H, Sirard MA, Kanitz W, Schellander K, Tesfaye D. 2008. Molecular and subcellular characterisation of oocytes screened for their developmental competence based on glucose-6-phospate dehydrogenase activity. Reproduction 135: 197-212
Viana JHM, Ferreira ADM, Ferreira W, Camargo LSA. 2000. Follicular dynamics in zebu cattle. Pesq Agropec Bras 35(12): 10-20
Watson AJ. 2007. Oocyte cytoplasmic maturation: a key mediator of oocyte and embryo developmental competence. J Anim Sci 85: E1-E3
25
27
Lampiran 1 Penghitungan konsentrasi BCB 26 µM
Definisi 1 Molar (M) : 1 M BCB berisi 1 mol BCB (=385.96 g) per liter PBS. Untuk membuat 26 µM BCB maka 26 x (385.96 x10-6 g) per liter PBS.
Lampiran 2 Komposisi medium transportasi ovarium
Bahan Jumlah
Sodium Chloride (Sigma-Aldrich, USA) 9.0 g
Milli Q water 1000 mL
Penicillin (100 IU/mL) dan streptomycin 1000 µL
Total 1000 mL
Larutan disterilisasi menggunakan auto clave pada suhu 121ºC selama 30 menit. Penambahan antibiotik penicillin-streptomycin dilakukan pada suhu 27-28ºC setelah sterilisasi.
1 M = massa molekul dalam g/L
28
Lampiran 3 Komposisi medium koleksi oosit (modified Phosphate Buffered Saline, mPBS)
Bahan Jumlah
PBS (Dulbecco’s Nissui, Japan) 90 mL
Fetal Bovine Serum (FBS) 10% (v/v) 10 mL
Penicillin (100 IU/mL) dan Streptomycin 100 µL
Total 100 mL
Stok PBS:
Untuk membuat stok PBS, maka 9.6 g PBS (Dulbecco’s Nissui, Japan) dilarutkan dalam 1000 mL Milli-Q water dan setelahnya difiltrasi menggunakan filter (Sartorius®) ukuran 0.22 µm.
Stok penicillin dan streptomycin:
Dosis penicillin G: 0.06 g/l ~ 100.000 IU/l Dosis streptomycin sulfate: 0.1 g/l
Untuk membuat stok penicillin-streptomycin, maka 0.0006 g penicillin (Sigma-Aldrich. Inc, P-4687) dan 0.001 g streptomycin (Sigma-Aldrich. Inc, S-9137) dilarutkan dalam 10 mL PBS.
Lampiran 4 Komposisi medium maturasi in vitro
Bahan Jumlah
Tissue Culture Medium/TCM-199 2700 µL
Fetal Bovine Serum (FBS) 10% 300 µL
Pregnant Mare Serum Gonadotropin (PMSG) 30 µL
Human Chorionic Gonadotropin (hCG) 30 µL
Gentamycin (50 µg/mL) 3 µL
Total 3 mL
Stok PMSG (Kyoritsu Seiyaku®, Japan) Dosis: 10 IU/mL
Untuk membuat stok, maka 1000 IU PMSG dilarutkan dalam 1 mL TCM-199 (1 IU/µL)
Stok hCG (Kyoritsu Seiyaku®, Japan) Dosis: 10 IU/mL
29
Lampiran 5 Komposisi medium fertilisasi in vitro
Reagents mM g/100mL
NaCl 90 0.52596
KCl 12 0.08946
NaHCO3 25 0.21003
NaH2PO4 anhydrous 0.5 0.006
MgSO4 7H2O 0.5 0.01233
Sodium Laktat
(60% sirup) 10
0.1121 0.19 mL
HEPES 10 0.2383
CaCl2 2H2O 8 0.1176
Sodium piruvat 2 0.0220
Kafein anhydrous 2 0.03884
BSA (fatty acid free) 5 0.5
Larutan stok dapat dibuat terlebih dahulu secara terpisah (A: NaCl, KCl, NaH2PO4
anhydrous, MgSO4 7H2O, Sodium Laktat, HEPES, CaCl2 2H2O; B: NaHCO3).
30
RIWAYAT HIDUP
Zultinur muttaqin lahir di Bogor pada tanggal 2 Juli 1987 dari pasangan bapak Prof Dr Nurheni Wijayanto dan ibu Kiswati S.Hut. Penulis merupakan anak ke-2 dari 4 bersaudara, yaitu Hanifa Akrom S.T., Aisyah Iadha Nuraini S.T, M.T., dan Muhammad Insan Rabbani. Pada Tahun 2005 penulis lulus dari SMA Negeri 2 Bogor dan pada tahun yang sama penulis diterima di Institut Pertanian Bogor (IPB) melalui jalur Seleksi Penerimaan Mahasiswa baru (SPMB). Pergantian kurikulum di IPB menjadi kurikulum baru (Mayor-Minor) menyebabkan pemilihan jurusan baru dilakukan pada tahun kedua. Setelah melewati tahapan tingkat persiapan bersama (TPB) pada tahun pertama, akhirnya pada tahun kedua penulis diterima di Fakultas Kedokteran Hewan (FKH).
Gelar sarjana kedokteran hewan (S.KH) diperoleh penulis pada tahun 2009, diikuti dengan gelar dokter hewan pada tahun 2011 melalui program pendidikan profesi dokter hewan (PPDH) IPB. Dalam rentang 2011 sampai 2012 penulis bekerja sebagai technical sales representative (TSR) wilayah Bandung, Jawa Barat di PT. Indovetraco Makmur Abadi (IMA) yang merupakan perusahaan obat hewan dari grup Charoen Pokphand. Setelah 1 tahun bekerja, penulis memutuskan untuk resign dan melanjutkan studi pascasarjana pada program Biologi Reproduksi (BRP) FKH-IPB dengan beasiswa dari Bakrie Graduate Fellowship. Selama mengikuti program master, penulis menulis tesis dengan judul
“Kemampuan Maturasi dan Fertilisasi Oosit Sapi yang Diseleksi Menggunakan Teknik Pewarnaan Brilliant Cresyl Blue secara in vitro” yang diterbitkan pada Jurnal Veteriner, Fakultas Kedokteran Hewan Universitas Udayana, Bali. Karya ilmiah tersebut merupakan bagian dari program master penulis yang diharapkan dapat bermanfaat khususnya bagi diri penulis sendiri, para pencari ilmu dan bagi masyarakat sekalian, sehingga karya ilmiah ini dapat menjadi amal jariyah yang pahalanya tidak ada putusnya dan terus mengalir hingga hari kiamat. Aamiin Ya