ISOLASI DAN IDENTIFIKASI MOLEKULER BAKTERI
ENDOFIT TUMBUHAN
Lantana camara
L. SERTA
IDENTIFIKASI SENYAWA ANTIBAKTERINYA
DINA DYAH SAPUTRI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Isolasi dan Karakterisasi dengan Analisis 16S rRNA Bakteri Endofit dari Tumbuhan Tembelekan (Lantana camara L.) sebagai Penghasil Senyawa Antibakteri adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2015
Dina Dyah Saputri
RINGKASAN
DINA DYAH SAPUTRI. Isolasi dan Karakterisasi dengan Analisis 16S rRNA Bakteri Endofit dari Tumbuhan Tembelekan (Lantana camara L.) sebagai Penghasil Senyawa Antibakteri. Dibimbing oleh MARIA BINTANG dan FACHRIYAN HASMI PASARIBU.
Peningkatan kasus penyakit infeksi yang disebabkan oleh bakteri patogen, berdampak pada meningkatnya kejadian resistensi bakteri patogen terhadap antibiotik komersil yang saat ini banyak digunakan. Pemanfaatan tumbuhan akan menjadi sumber terbaik untuk mendapatkan berbagai obat yang alami dan aman. Salah satu tumbuhan yang telah diketahui khasiatnya sebagai obat adalah tumbuhan tembelekan (Lantana camara L.). Lantana camara L. dapat mengobati berbagai jenis penyakit. Bagian akar digunakan untuk mengobati penyakit influenza disertai demam tinggi, TBC kelenjar (skrofulodeurm), rematik, dan penyakit kulit. Bagian daunnya digunakan sebagai obat sakit batuk, kulit, bisul, bengkak, gatal-gatal, panas tinggi, rematik, dan memar. Permasalahannya adalah pengambilan senyawa bioaktif memerlukan biomassa yang cukup banyak. Alternatif terbaru dari permasalahan tersebut adalah dengan pemanfaatan bakteri endofit di dalam jaringan tumbuhan. Bakteri endofit merupakan mikroba yang hidup di dalam jaringan internal tumbuhan hidup tanpa menyebabkan efek negatif langsung yang nyata. Kemampuan bakteri endofit memproduksi senyawa metabolit sekunder sesuai dengan tumbuhan inangnya merupakan peluang yang sangat besar dan dapat diandalkan untuk memproduksi senyawa bioaktif yang berpotensi sebagai antibiotik.
Isolasi bakteri dari tumbuhan Lantana camara L. menghasilkan sebanyak 21 isolat bakteri endofit. Semua isolat dilakukan uji aktivitas antibakteri terhadap bakteri patogen Escherichia coli, Salmonella enteritidis, Bacillus cereus, Staphylococcus aureus. Isolat potensial BT22 mampu menghambat keempat bakteri patogen. Selanjutnya dilakukan karakterisasi isolat BT22 dengan analisis sekuen 16S rRNA. Hasil analisis sekuen 16S rRNA menunjukkan bahwa isolat BT22 memiliki kemiripan sebanyak 99% dengan Bacillus amyloliquefaciens YB-1402. Ekstraksi senyawa antibakteri isolat BT22 dengan menggunakan pelarut kloroform. Identifikasi senyawa antibakteri dengan analisis GC-MS menunjukkan beberapa senyawa antibakteri di antaranya : sikloheksanon, tetrakosan dan oktadekan.
SUMMARY
DINA DYAH SAPUTRI. Isolation and Characterization of the 16S rRNA Analysis Endophytic Bacteria from Tembelekan (Lantana camara L.) can Produce Antibacterial Compounds. Supervised by MARIA BINTANG and FACHRIYAN HASMI PASARIBU.
The cases of infection diseases which was caused by pathogenic bacteria was increased, so the effect was increased incidence of pathogenic bacterial resistance to antibiotics commercial. Utilization of the plant will be the best source to get various drugs that are natural and safety. One of the plants that have known as medicinal plants is tembelekan (Lantana camara L.). Lantana camara
L. can treat many diseases. The roots are used to treat influenza disease, tuberculosis gland (skrofulodeurm), rheumatism, and skin diseases. The leaves are used as a cough medicine, skin, boils, swelling, itching, fever, rheumatism, and bruises. The problem is making bioactive compounds requires considerable biomass. The latest alternative to these problems is the use of endophytic bacteria in plant tissue. Endophytic bacteria are microbes that live in the internal tissues of plant life without causing significant negative effect. Endophytic bacteria can produce secondary metabolites which is same with the host plant is a huge opportunity and can be relied upon to produce bioactive compounds that have the potential as an antibiotic.
Isolation of endophytic bacteria from Lantana camara L. produce 21 isolates of endophytic bacteria. All isolates were tested antibacterial activity against pathogenic bacteria Escherichia coli, Salmonella enteritidis, Bacillus cereus, Staphylococcus aureus. Potential isolates of BT22 could inhibit fourth pathogenic bacteria. Characterization of isolates BT22 with 16S rRNA sequence analysis. The results of 16S rRNA sequence analysis showed that BT22 isolate have similarities as much as 99% by Bacillus amyloliquefaciens YB-1402. Extraction of antibacterial compounds BT22 isolate using chloroform. The identification of antibacterial compounds with GC-MS analysis showed some antibacterial compounds include: Cyclohexanone, Tetracosane and Octadecane
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biokimia
ISOLASI DAN IDENTIFIKASI MOLEKULER BAKTERI
ENDOFIT TUMBUHAN
Lantana camara
L. SERTA
IDENTIFIKASI SENYAWA ANTIBAKTERINYA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah SWT atas segala rahmat dan karunia-Nya, sehingga penulis dapat menyelesaikan tesis yang berjudul Isolasi dan Karakterisasi dengan Analisis 16S rRNA Bakteri Endofit dari Tumbuhan Tembelekan (Lantana camara L.) sebagai Penghasil Senyawa Antibakteri yang telah dilaksanakan sejak bulan September 2014 sampai Februari 2015 di Laboratorium Bakteriologi Bagian Mikrobiologi Medik Departemen Ilmu Penyakit Hewan dan Kesehatan Masyarakat Veteriner Fakultas Kedokteran Hewan, Laboratorium Penelitian Departemen Biokimia Fakultas Matematika dan Ilmu Pengetahuan Alam Institut Pertanian Bogor (IPB), dan Laboratorium Pengujian Hasil Hutan P3KKPHH Bogor.
Terima kasih, penghargaan, dan apresiasi penulis ucapkan kepada Prof Dr Drh Maria Bintang, MS dan Prof Dr Drh Fachriyan H. Pasaribu sebagai pembimbing atas arahan, bimbingan, perhatian, nasihat, motivasi dan masukkannya selama penelitian serta dalam penyusunan tesis ini. Penulis juga mengucapkan terima kasih kepada Bapak Agus Somantri yang sudah banyak membantu penulis untuk melakukan penelitian di Laboratorium Bakteriologi Bagian Mikrobiologi Medik Departemen Ilmu Penyakit Hewan dan Kesehatan Masyarakat Veteriner Fakultas Kedokteran Hewan, Institut Pertanian Bogor (IPB).
Penelitian ini didanai oleh Direktorat Jenderal Pendidikan Tinggi (DIKTI) melalui Beasiswa Pendidikan Pascasarjana Dalam Negeri (BPPDN). Tidak lupa juga terima kasih penulis ucapkan kepada keluarga, teman-teman Bakteriologi Bagian Mikrobiologi Medik Departemen Ilmu Penyakit Hewan dan Kesehatan Masyarakat Veteriner Fakultas Kedokteran Hewan, Institut Pertanian Bogor (IPB), teman-teman SPs IPB program studi Biokimia 2013 yang selalu mendukung penulis.
Penyusunan tesis ini tentunya tidak terlepas dari kekurangan. Oleh karena itu, penulis mengharapkan adanya saran dan kritik yang bersifat membangun untuk menyempurnakan penyusunan tesis ini. Semoga hasil penelitian ini dapat bermanfaat bagi kemajuan ilmu pengetahuan.
Bogor, Agustus 2015
DAFTAR ISI
Isolat Bakteri Endofit dari Tumbuhan Lantana camara L. 6 Aktivitas Antibakteri Isolat Bakteri Endofit terhadap Bakteri Patogen 8 Sekuen 16S rRNA dan Rekonstruksi Pohon Filogenetik 9 Aktivitas Senyawa Antibakteri Ekstrak Kloroform Isolat BT22 10 Senyawa Antibakteri Ekstrak Kloroform Isolat BT22 dengan GC-MS 11
4 PEMBAHASAN 13
Isolat Bakteri Endofit dari Tumbuhan Lantana camara L. 13 Aktivitas Antibakteri Isolat Bakteri Endofit terhadap Bakteri Patogen 14 Sekuen 16S rRNA dan Rekonstruksi Pohon Filogenetik 14 Aktivitas Senyawa Antibakteri Ekstrak Kloroform Isolat BT22 15 Senyawa Antibakteri Ekstrak Kloroform Isolat BT22 dengan GC-MS 16
DAFTAR GAMBAR
1 Tumbuhan Tembelekan (Lantana camara L.) 7
2 Persentase bakteri endofit yang diisolasi dari tumbuhan
Lantana camara L. 8
3 Zona hambat isolat BT22 yang terbentuk terhadap bakteri
patogen 8
4 Isolat BT22 bakteri endofit dari tumbuhan Lantana camara L 9 5 Sekuen 16S rRNA isolat potensial BT22 hasil pensejajaran 9 6 Zona hambat ekstrak kloroform isolat BT22 terhadap bakteri patogen 11
DAFTAR TABEL
1 Ciri morfologi koloni dan kecepatan pertumbuhan isolat bakteri endofit
dari tumbuhan Lantana camara L. 7
2 Diameter zona hambat yang terbentuk pada seleksi isolat
bakteri endofit 8
3 Diameter zona hambat ekstrak senyawa antibakteri isolat potensial BT22
dengan berbagai pelarut 10
4 Klasifikasi aktivitas antibakteri berdasarkan diameter zona
hambat 10
5 Senyawa yang diduga berperan sebagai senyawa antibakteri
yang terkandung di dalam isolat BT22 12
DAFTAR LAMPIRAN
1 Bagan alir penelitian 25
2 Determinasi tumbuhan Lantana camara L. 26
3 Diameter zona hambat yang terbentuk pada isolat bakteri
endofit terhadap bakteri patogen 27
4 Sekuen 16S rRNA isolat potensial BT22 28 5 Hasil BLAST sekuen 16S rRNA isolat potensial BT22 hasil pensejajaran 29 6 Rekonstruksi pohon filogenetik isolat potensial BT22 30
7 Kondisi operasional alat GC-MS 31
1
PENDAHULUAN
Latar Belakang
Peningkatan kasus penyakit infeksi yang disebabkan oleh bakteri patogen, berdampak pada meningkatnya kejadian resistensi bakteri patogen terhadap antibiotik komersil yang saat ini banyak digunakan. Bakteri patogen yang sudah resisten terhadap antibiotik di antaranya, Staphylococcus aureus yang resisten terhadap antibiotik penisilin, oksasilin, serta metisilin. Escherichia coli yang resisten terhadap antibiotik beta laktam. Pseudomonas aeruginosa yang resisten terhadap antibiotik trimetoprim/sulfametoksasol, tetrasiklin, dan sefalosporin (Mardiastuti et al. 2007).
Menurut WHO pemanfaatan tumbuhan akan menjadi sumber terbaik untuk mendapatkan berbagai obat, sehingga tumbuhan tersebut harus diteliti untuk lebih memahami sifat-sifat tumbuhan terkait keamanan dan khasiatnya (Nascimento et al. 2000). Sekitar 20% dari tumbuhan di dunia telah dilakukan tes farmakologis atau biologis dan sejumlah besar antibiotik yang diperkenalkan di pasar diperoleh dari sumber daya alam atau semi sintesis (Mothana & Lindequist 2005). Lebih dari 50% dari semua obat klinis modern berasal dari alam dan produk alami sangat berperan penting dalam pengembangan obat dalam industri farmasi (Baker
et al. 1995).
Indonesia dikenal sebagai salah satu dari 7 negara yang memiliki keanekaragaman hayati terbesar di dunia setelah Brazil, sehingga sangat potensial dalam pengembangan obat herbal (Radji 2005). Salah satu tumbuhan yang telah diketahui khasiatnya sebagai obat adalah tumbuhan tembelekan (Lantana camara
L.). Menurut Deepak et al. (2009), Lantana camara L. memiliki potensi terapeutik karena adanya senyawa bioaktif flavon, isoflavon, flavonoid, anthosianidin, lignan, katesin, isokatesin, alkaloid, tanin, saponin dan terpenoid.
Barre et al. (1997) melaporkan bahwa triterpenoid pada Lantana camara
L. menunjukkan aktivitas yang dapat menghambat pertumbuhan bakteri
Staphylococcus aureus dan Salmonella typhi. Kajian mengenai Lantana camara
L. dalam bidang kesehatan telah banyak dilakukan. Sebagai contoh, ekstrak daun
Lantana camara L. menunjukkan aktivitas penghambatan terhadap strain
Mycobacterium tuberculosis (Claude et al. 2009).
Lantana camara L. telah diketahui khasiatnya untuk mengobati berbagai jenis penyakit. Bagian akar digunakan untuk mengobati penyakit influenza disertai demam tinggi, TBC kelenjar (skrofulodeurm), rematik, bengkak, memar, keputihan, kencing nanah, gondongan, dan penyakit kulit. Bagian daunnya digunakan sebagai obat sakit batuk, kulit, bisul, bengkak, gatal-gatal, panas tinggi, rematik, dan memar (Dalimartha 2007). Di India, daun Lantana camara L. direbus untuk diminum sebagai obat batuk dan ditumbuk untuk digunakan sebagai obat luka, bisul, dan bengkak (Verma & Verma 2006).
2
dengan pemanfaatan bakteri endofit di dalam jaringan tumbuhan. Bakteri endofit merupakan mikroba yang hidup di dalam jaringan internal tumbuhan hidup tanpa menyebabkan efek negatif langsung yang nyata. Sifat bakteri endofit yang tidak berdampak negatif pada jaringan tumbuhan menunjukkan kemungkinan adanya hubungan simbiosis mutualisme antara bakteri endofit dan inangnya (Strobel & Daisy 2003).
Tumbuhan tingkat tinggi dapat mengandung beberapa bakteri endofit yang mampu menghasilkan senyawa biologi atau metabolit sekunder yang diduga sebagai akibat koevolusi atau transfer genetik (genetic recombination) dari tumbuhan inangnya ke dalam bakteri endofit (Radji 2005). Endofit di dalam tumbuhan berada di ruang antarsel. Endofit pada awalnya ada di luar tubuh tumbuhan, kemudian masuk jika terjadi luka pada tumbuhan. Jika sudah berada dalam tumbuhan, endofit akan menetap. Tempat hidup bakteri sangat unik sifatnya karena tumbuh dalam jaringan tumbuhan, dimana tumbuhan yang satu tentunya berbeda dengan tumbuhan lainnya. Fisiologi tumbuhan tinggi yang berasal dari spesies yang sama akan berbeda jika tumbuh di lingkungan yang berbeda, sehingga dapat dikatakan bahwa keanekaragaman bakteri endofit sangatlah tinggi. Berdasarkan pertimbangan tersebut endofit dapat menjadi sumber berbagai metabolit sekunder baru yang berpotensi untuk dikembangkan dalam bidang medis, pertanian, dan industri (Prasetyoputri & Ines 2006).
Bakteri endofit dapat menghasilkan senyawa-senyawa bioaktif yang langka dan penting bagi tumbuhan inangnya, maka kebutuhan untuk menumbuhkan tumbuhan yang masa hidupnya panjang dan mungkin termasuk langka akan berkurang dan keanekaragaman hayati dunia juga terlindungi. Bakteri digunakan sebagai sumber suatu produk hayati akan memudahkan proses dan mengurangi biaya produksi, sehingga pada akhirnya menghasilkan produk dengan harga lebih murah. Kemampuan bakteri endofit memproduksi senyawa metabolit sekunder sesuai dengan tumbuhan inangnya merupakan peluang yang sangat besar dan dapat diandalkan untuk memproduksi metabolit sekunder (Radji 2005).
Berbagai penelitian mengenai potensi senyawa bioaktif yang terkandung dalam bakteri endofit telah banyak dilakukan. Beberapa bakteri endofit menghasilkan hormon IAA yang terdapat pada tumbuhan tertentu dan menghasilkan fitohormon yang bermanfaat bagi pertumbuhan tumbuhan tersebut (Aryantha et al. 2005). Beberapa jenis bakteri endofit yang dapat menghasilkan antibiotika berspektrum luas adalah bakteri endofit yang diisolasi dari tumbuhan
Grevillea pteridifolia menghasilkan metabolit kakadumycin yang berkhasiat sebagai anti malaria (Castillo et al. 2003). Bakteri endofit Pestalotiopsis microspora yang diisolasi dari tumbuhan Taxus mampu menghasilkan senyawa metabolit paclitaxel yang berkhasiat sebagai antikanker (Strobel et al. 2002). Endofit Pseudomassaria sp menghasilkan metabolit sekunder yang bekerja seperti insulin dan telah terbukti dapat menurunkan glukosa darah tikus yang terkena diabetes (Zhang et al. 1999).
3 yang diduga berpeluang besar mampu menghasilkan senyawa aktif yang memiliki aktivitas yang sama dengan tumbuhan inangnya.
Perumusan Masalah
Tumbuhan tembelekan (Lantana camara L.) telah terbukti memiliki khasiat sebagai obat, sehingga berpotensi untuk dijadikan sumber antibiotik alami. Penelitian mengenai ekstrak tumbuhan tembelekan telah banyak dilakukan, namun kajian mengenai pemanfaatan senyawa metabolit sekunder bakteri endofit yang diisolasi dari tumbuhan tembelekan sebagai alternatif terbaru yang berpotensi menghasilkan antibiotik alami sejauh ini belum dilaporkan.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengisolasi dan mengidentifikasi bakteri endofit dan aktivitas antibakterinya dari tumbuhan tembelekan (Lantana camara
L.) serta menganalisis senyawa antibakteri yang dihasilkan oleh isolat bakteri endofit potensial dengan menggunakan Gass Chromatography-Mass Spectrometer (GC-MS).
Hipotesis
Bakteri endofit dapat diisolasi dari tumbuhan tembelekan (Lantana camara L.) dan memiliki aktivitas antibakteri terhadap bakteri patogen (S. aureus, E. coli, B. cereus dan S. enteritidis) serta isolat bakteri endofit potensial dapat dikarakterisasi secara molekuler menggunakan analisis sekuen 16S rRNA. Senyawa aktif dari isolat bakteri endofit potensial yang berperan dalam aktivitas antibakteri dapat dianalisis menggunakan GC-MS.
Manfaat Penelitian
4
2 METODE
Waktu dan Tempat Penelitian
Penelitian ini telah dilaksanakan pada bulan September 2014 – Februari 2015 di Laboratorium Bakteriologi Bagian Mikrobiologi Medik Departemen Ilmu Penyakit Hewan dan Kesehatan Masyarakat Veteriner Fakultas Kedokteran Hewan, Laboratorium Penelitian Departemen Biokimia Fakultas Matematika dan Ilmu Pengetahuan Alam Institut Pertanian Bogor (IPB), dan Laboratorium Pengujian Hasil Hutan P3KKPHH Bogor.
Bahan
Bahan yang digunakan antara lain : serabut akar (S), batang (B), dan daun (D) tumbuhan tembelakan (Lantana camara L.) yang diperoleh dari area Kampus IPB Darmaga, Bogor, dan telah dilakukan determinasi di Pusat Konservasi Tumbuhan, Kebun Raya Bogor – LIPI, Bogor (Lampiran 2), media Nutrient Agar
(NA) produk Oxoid, media Nutrient Broth (NB) produk Difco, sodium hipoklorit (NaOCl) 5.25%, nistatin, etanol 96%, pelarut kloroform, n-heksana, etil asetat dan etanol 70%, kloramfenikol, akuades, larutan garam, aseton, primer 27F (5’ – AGA GTT TGA TCM TGG GTC AG – 3’), 1492R (5’ – TAC GGY TAC CTT GTT ACG ACT T – 3’), 518F ( 5’- CCA GCA GCC GCG GTA ATA CG -3’ ) dan 800R ( 5’- TAC CAG GGT ATC TAA TCC -3’), InstaGene Matrix (Bio-Rad, USA), Montage PCR Clean up kit (Milipore), Big Dye terminator cycle sequencing kit (Apllied BioSystem, USA). Kultur bakteri patogen meliputi:
Escherichia coli, Salmonella enteritidis, Bacillus cereus, Staphylococcus aureus
berasal dari koleksi Laboratorium Bakteriologi Bagian Mikrobiologi Medik Fakultas Kedokteran Hewan, Institut Pertanian Bogor, Darmaga, Bogor, Jawa Barat.
Alat
Alat yang digunakan antara lain : pipet mikro, alat gelas, cawan Petri,
shaker incubator, mesin sentrifugasi, mesin Polymerase Chain Reaction (PCR),
Applied Biosystem model 3730XL automated DNA sequencing system (Apllied BioSystem, USA), corong pisah, vakum evaporator, dan mesin GC-MS (Gas Chromatography-Mass Spectrometer) tipe Shimadzu GCMS-QP2010.
Prosedur Analisis Data
Isolasi Bakteri Endofit (Modifikasi Desriani et al. 2013)
5 cairan perendam dibuang dan diganti dengan sodium hipoklorit (NaOCl) lalu didiamkan selama 5 menit. Cairan perendam dibuang kembali dan sampel dibilas dengan etanol 96% sebanyak tiga kali. Sampel yang telah steril dipotong lagi menjadi beberapa bagian lalu ditanam pada media NA yang telah ditambahkan nistatin (200 μL/200mL) dan diinkubasi pada ruang gelap selama 48 jam. Pemurnian dilakukan dengan memindahkan bakteri endofit yang tumbuh pada media NA yang ditambahkan nistatin ke cawan Petri yang berisi NA steril. Setelah diperoleh biakan murni, bakteri endofit disimpan ke agar miring NA. Uji Aktivitas Antibakteri (Simarmata et al. 2007)
Isolat bakteri endofit dari agar miring diregenerasi ke media NA, sedangkan bakteri patogen diregenerasikan ke dalam 5 mL media NB lalu diinkubasi selama 24 jam pada suhu 28-30 ºC. Sebanyak 0.4 mL kultur cair bakteri uji dimasukkan ke dalam 80 mL media NA yang bersuhu ± 40 ºC. Selanjutnya sebanyak 20 mL dituangkan ke dalam cawan Petri steril dan ditunggu hingga memadat. Isolat bakteri endofit yang akan diuji diinokulasikan ke media berisi patogen menggunakan ose, lalu diinkubasi selama 24-48 jam. Zona hambat yang terbentuk diamati kemudian diameter zona hambat diukur. Isolat bakteri endofit yang positif menunjukkan zona hambat terhadap semua patogen dikatakan sebagai isolat potensial.
Analisis Sekuen 16S rRNA (Kusumawati 2014) dan Rekonstruksi Pohon Filogenetik (Singh et al. 2012)
Preparasi DNA cetakan dari isolat potensial. Koloni isolat potensial disuspensikan ke dalam 0.5 mL larutan garam steril pada tabung sentrifugasi yang berukuran 1.5 mL, kemudian disentrifugasi dengan kecepatan 10.000 rpm selama 10 menit. Selanjutnya pelet diresuspensi dengan penambahan 0.5 mL InstaGene Matrix (Bio-Rad, USA). Setelah itu, diinkubasi pada suhu 56 ºC selama 30 menit, lalu dipanaskan 100 ºC selama 10 menit. Kemudian setelah proses pemanasan selesai, supernatan yang berisi DNA cetakan siap digunakan untuk proses amplifikasi.
Amplifikasi DNA dan penentuan urutan basa DNA untuk sekuen 16S rRNA. Amplifikasi DNA dilakukan menggunakan mesin PCR (Polymerase Chain Reaction). Primer yang digunakan pada proses PCR adalah 27F dan 1492R. Volume DNA cetakan yang ditambahkan adalah 1 µL dari 20 µL total larutan reaksi. Amplifikasi dilakukan sebanyak 35 siklus dengan kondisi denaturasi awal 94 °C selama 45 detik, anneling 55 ºC selama 60 detik dan elongasi pada suhu 72 ºC selama 60 detik. Kemudian purifikasi produk PCR dilakukan dengan menggunakan Montage PCR Clean up kit (Milipore). Setelah itu, ditentukan urutan basa DNA (sequencing) dari hasil PCR dengan menggunakan dua primer (518F dan 800R) dan Big Dye terminator cycle sequencing kit (Apllied BioSystem, USA), lalu dianalisis menggunakan Applied Biosystem model 3730XL automated DNA sequencing system (Apllied BioSystem, USA).
Analisis hasil sequencing. Pensejajaran urutan basa DNA hasil
6
Rekonstruksi pohon filogenetik (Singh et al. 2012). Pohon filogenetik untuk mengetahui hubungan kekerabatan isolat bakteri endofit potensial dengan spesies lain. Pembuatan pohon filogenetik dengan menggunakan software ClustalX2 dan dilanjutkan dengan NJPlot.
Fermentasi dan Ekstraksi Senyawa Antibakteri Isolat BT22 (Ahamed 2012, Garcia et al. 2012)
Fermentasi dilakukan dengan menumbuhkan isolat bakteri endofit potensial ke dalam 50 mL media cair NB, kemudian diinkubasi dalam “shaker incubator” selama 48 jam pada suhu 28-30 ºC dengan kecepatan 150 rpm. Kultur bakteri hasil fermentasi dipindahkan ke dalam tabung sentrifugasi steril dan disentrifugasi dengan kecepatan 3.600 rpm selama 10 menit untuk memisahkan pelet dan supernatan. Supernatan yang telah terpisah dipindahkan ke corong pemisah dan diekstrak menggunakan berbagai pelarut yaitu kloroform, n-heksana, etil asetat dan etanol 70% dengan perbandingan 1:1 (v/v). Ekstrak dari kloroform,
n-heksana, etil asetat dan etanol 70% dilakukan uji aktivitas antibakteri terhadap bakteri patogen dengan kloramfenikol sebagai kontrol positif. Selanjutnya, larutan metabolit sekunder yang didapat dievaporasi pada vakum evaporator dengan suhu 40 ºC hingga terbentuk ekstrak kering.
Identifikasi Senyawa Antibakteri dengan Gas Chromatography – Mass Spectrometer (GC-MS) (Kusumawati 2014)
Senyawa metabolit sekunder dari hasil ekstraksi dilarutkan dengan aseton, kemudian diinjeksikan ke alat GC-MS. Proses GC-MS menggunakan kolom kapiler tipe Phase Rtx-5MS dengan panjang 60 m dan diameter 0.25 mm. Kondisi alat meliputi : suhu kolom 50 ºC, gas helium, SPL Temperature 280 ºC, MS Interface 280 ºC, pyrolisis temperature 280 ºC, dan ion surface 200 ºC (Lampiran 8).
3
HASIL
Isolat Bakteri Endofit dari Tumbuhan Lantana camara L.
Tumbuhan tembelekan (Lantana camara L.) yang digunakan sebagai sumber bakteri endofit dalam penelitian ini dengan daun berwarna hijau dan tepian bergerigi serta batang berduri (Gambar 1). Sebanyak 21 isolat murni bakteri endofit telah diperoleh dari hasil isolasi tumbuhan tembelekan (Lantana camara L.), yaitu: 13 isolat dari akar (AT), 5 isolat dari batang (BT), dan 3 isolat dari daun (DT). Persentase isolat bakteri endofit disajikan pada Gambar 2.
7
Gambar 1 Tumbuhan Lantana camara L. (sumber : dokumentasi pribadi, 2015)
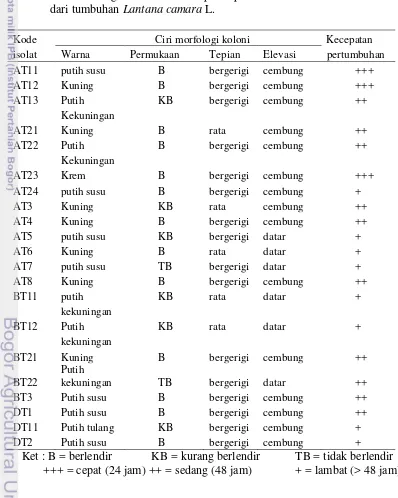
Tabel 1 Ciri morfologi koloni dan kecepatan pertumbuhan isolat bakteri endofit dari tumbuhan Lantana camara L.
Kode Ciri morfologi koloni Kecepatan
isolat Warna Permukaan Tepian Elevasi pertumbuhan
8
Gambar 2 Persentase bakteri endofit yang diisolasi dari tumbuhan Tembelekan Aktivitas Antibakteri Isolat Bakteri Endofit terhadap Bakteri Patogen
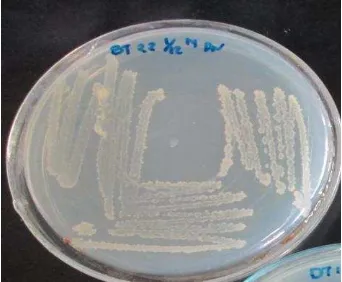
Uji aktivitas antibakteri dari isolat bakteri endofit dilakukan dengan metode inokulasi titik terhadap bakteri patogen Bacillus cereus, Escherichia coli, Staphylococcus aureus, dan Salmonella enteritidis. Berdasarkan hasil uji aktivitas antibakteri, beberapa isolat yang mampu menghasilkan zona hambat di antaranya AT22, AT3, BT21, BT22, dan DT11 (Tabel 2). Hal ini menandakan bahwa isolat tersebut berpotensi sebagai antibakteri. Isolat BT22 merupakan isolat paling potensial karena memiliki aktivitas antibakteri dengan menghambat keempat bakteri patogen (Gambar 3). Isolat BT22 berwarna putih kekuningan dengan tepian bergerigi, permukaan tidak berlendir dan elevasi datar (Gambar 4).
Tabel 2 Diameter zona hambat yang terbentuk pada seleksi isolat bakteri endofit
Keterangan : V = terdapat zona hambat namun sangat kecil dan sulit terukur
Gambar 3 Zona hambat isolat BT22 yang terbentuk terhadap bakteri patogen
No Kode Diameter zona hambat (mm)
Isolat B. cereus E. coli S. aureus S.enteritidis
1 AT22 - V - -
2 AT3 - V - V
3 BT21 2 3 V -
4 BT22 3 V 8 5
9
Gambar 4 Isolat BT22 bakteri endofit dari tumbuhan Lantana camara L. Sekuen 16S rRNA dan Rekonstruksi Pohon Filogenetik
Identifikasi isolat potensial BT22 dengan menggunakan analisis 16S rRNA. Urutan sekuen basa 16S rRNA dibandingkan dengan urutan basa yang telah tersimpan dalam NCBI-GenBank database. Hasil sekuensing isolat BT22 yang telah disejajarkan menunjukkan bahwa gen 16S rRNA pada isolat tersebut berukuran 1477 pb (pasang basa) (Gambar 5). Berdasarkan hasil analisis sekuen 16S rRNA menggunakan program BLAST, isolat BT22 memiliki persentase kemiripan dengan Bacillus amyloliquefaciens YB-1402 sebesar 99% (Lampiran 5). Hal ini didukung dengan rekonstruksi pohon filogenetik untuk mengetahui hubungan kekerabatan antar spesies bakteri. Pensejajaran sekuen hasil penelitian dan beberapa sekuen bakteri lain yang diambil dari NCBI-GenBank database
dengan menggunakan clustalX2. Kemudian hasil pensejajaran dimasukkan ke dalam NJPlot untuk merekonstruksi pohon filogenetik. Hasil rekonstruksi pohon filogenetik menunjukkan bahwa isolat BT22 memiliki hubungan kekerabatan yang paling dekat dengan Bacillus amyloliquefaciens (Lampiran 6).
>BT22_Consensus (1477 pb)
10
Aktivitas Senyawa Antibakteri Ekstrak Kloroform Isolat Potensial BT22
Ekstraksi senyawa antibakteri isolat potensial BT22 dilakukan dengan menggunakan pelarut kloroform, n-heksana, etil asetat, dan etanol 70%. Selanjutnya ekstrak dari kloroform, n-heksana, etil asetat, dan etanol 70% isolat potensial BT22 diujikan kembali dengan bakteri patogen Bacillus cereus, Escherichia coli, Staphylococcus aureus, dan Salmonella enteritidis.
Kloramfenikol 60µg/mL digunakan sebagai kontrol positif. Berdasarkan hasil uji, ekstrak kloroform isolat BT22 memiliki aktivitas antibakteri yang paling baik karena mampu menghambat pertumbuhan Bacillus cereus, Escherichia coli, Staphylococcus aureus, dan Salmonella enteritidis. Hal ini terlihat dengan terbentuknya zona bening di sekitar lubang sumur masing-masing sebesar 3 mm, 7 mm, 2 mm, dan 1 mm (Tabel 3). Diameter zona hambat ekstrak kloroform isolat BT22 terhadap Bacillus cereus, Escherichia coli, Staphylococcus aureus, dan
Salmonella enteritidis ditunjukkan pada Gambar 6. Menurut Zahro & Agustini (2013), aktivitas antibakteri dapat digolongkan berdasarkan besarnya zona hambat yang terbentuk dapat diklasifikasikan dalam Tabel 4.
Tabel 3 Diameter zona hambat ekstrak senyawa antibakteri isolat potensial BT22 dengan berbagai pelarut
Jenis Diameter zona hambat (mm)
Pelarut B. cereus E. coli S. aureus S. enteritidis
Kloroform 3 7 2 1
n-heksana 3
Etil asetat 1 Etanol 70%
Tabel 4 Klasifikasi aktivitas antibakteri berdasarkan diameter zona hambat Aktivitas antibakteri Diameter zona hambat (mm)
Lemah < 5
Sedang 5 – 10
Kuat 10 – 20
11
Gambar 6 Zona hambat ekstrak kloroform terhadap bakteri patogen
Senyawa Antibakteri Ekstrak Kloroform Isolat BT22 dengan GC-MS
12
Tabel 5 Senyawa yang diduga berperan sebagai senyawa antibakteri yang terkandung di dalam ekstrak kloroform isolat BT22
No Nama Senyawa Rumus
Molekul
Berat Molekul
Aktivitas Biologi
1 Sikloheksanon C6H10O 98.15 Antibakteri dan
antifungi (Jha et al.
7 Nonakosan C29H60 408 Antibakteri (Saracoglu et al. 2012)
8 Metil eugenol (ME) C11H14O2 178 Antibakteri terhadap
Campylobacter jejuni
C16H22O4 278 Antifungi, antibakteri, dan anti malaria (Elija et al. 2012)
11 Asam heksadekanoat C17H34O2 270 Antibakteri terhadap
Salmonella typhi,
Shigella sp., dan
Enterococcus faecalis (Manilal et al. 2009) 12 Asam heksanedioat C22H42O4 370 Antibakteri terhadap S.
aureus, K. pneumoniae,
dan Shigella dysentriae
13
4
PEMBAHASAN
Isolat Bakteri Endofit dari Tumbuhan Lantana camara L.
Isolasi tumbuhan tembelekan (Lantana camara L.) untuk menghasilkan bakteri endofit yang berpotensi sebagai penghasil senyawa antibakteri merupakan alternatif terbaru tanpa memerlukan ekstraksi tumbuhan obat dalam jumlah yang banyak dan waktu yang cukup lama. Bakteri endofit tidak berdampak negatif bagi tumbuhan inang, namun terbentuk simbiosis mutualisme antara bakteri endofit dan tumbuhan inangnya (Strobel & Daisy2003). Bakteri endofit bermanfaat bagi tumbuhan inangnya karena mampu menghasilkan produk alami yang dapat digunakan sebagai obat, bidang pertanian, dan industri (Ryan et al. 2008).
Pemilihan tumbuhan tembelekan dikarenakan tumbuhan ini telah terbukti memiliki khasiat sebagai obat karena mengandung senyawa bioaktif flavon, isoflavon, flavonoid, anthosianidin, lignan, katesin, isokatesin, alkaloid, tanin, saponin dan terpenoid (Deepak et al. 2009) dan ekstraknya banyak digunakan untuk mengobati penyakit influenza disertai demam tinggi, rematik, bengkak, memar, serta penyakit kulit (Dalimartha 2007).
Hasil isolasi bakteri endofit dari tanaman Lantana camara L. didapatkan sebanyak 21 isolat di antaranya 13 isolat dari akar (AT), 5 isolat dari batang (BT), dan 3 isolat dari daun (DT). Hal ini sesuai dengan pernyataan Lodewyckx et al. (2002) yang menyatakan bahwa bakteri endofit dapat diisolasi dari bagian akar, daun, batang, bunga, buah, dan biji. Bakteri endofit banyak terdapat di akar dan semakin menurun jumlahnya pada batang dan daun (Lamb et al. 1996). Jumlah bakteri endofit yang dihasilkan banyak terdapat di bagian akar. Hal ini disebabkan karena bakteri endofit masuk ke jaringan tumbuhan pertama kali melalui akar (Zinniel et al. 2002). Perakaran tanaman (rhizosfer) banyak mengandung asam amino dan gula yang digunakan sebagai sumber nutrisi bagi bakteri endofit (Sylvia 2005), sehingga jaringan internal bagian perakaran memiliki kerapatan populasi bakteri paling tinggi dibandingkan bagian tanaman lain (Hallman 1997).
Isolasi bakteri endofit dari sampel tumbuhan dilakukan dengan metode sterilisasi permukaan (Hallman et al. 1997). Sodium hipoklorit (NaOCl) digunakan untuk sterilisasi permukaan yang bersifat bakterisidal dan virusidal (Bolfoni et al. 2014). Metode surface sterilization digunakan untuk menghilangkan mikroorganisme epifit pada sampel tumbuhan (Larran et al.
2001). Penambahan nistatin ke dalam media isolasi NA berfungsi sebagai antifungi (Silva et al. 2008).
14
Aktivitas Antibakteri Isolat Bakteri Endofit terhadap Bakteri Patogen
Hal ini sesuai dengan pernyataan Simarmata et al. (2007) yang menyatakan bahwa isolat mikroba endofit dikatakan mempunyai aktivitas antimikroba jika terbentuk zona jernih di sekeliling isolat mikroba endofit yang ditumbuhkan pada media yang telah diinokulasi oleh mikroba patogen.
Perbedaan diameter zona hambat kemungkinan disebabkan oleh kandungan senyawa antibakteri yang berbeda (Kusumawati 2014) dan diameter dinding sel bakteri patogen (Junanto et al. 2008). Beberapa isolat bakteri endofit tidak menunjukkan aktivitas antibakteri. Hal ini terjadi karena adanya kemungkinan bakteri endofit memiliki kandungan senyawa aktif namun jumlahnya sangat kecil atau mungkin juga mengandung senyawa aktif potensial lain (Son & Cheah 2002) seperti asam indol asetat (IAA) (Dudeja & Giri 2014) dan enzim (Kannan et al. 2015).
Hasil uji aktivitas antibakteri menunjukkan bahwa bakteri endofit dari tumbuhan tembelekan (Lantana camara L.) memiliki kemampuan untuk menghasilkan senyawa antibakteri (Tabel 2). Penelitian sebelumnya melaporkan bahwa ekstrak dari bunga, daun, batang, dan akar Lantana camara L. menunjukkan aktivitas antibakteri terhadap Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus, dan Staphylococcus saprohiticus (Mary 2011). Hal ini menandakan bahwa bakteri endofit yang diisolasi dari tumbuhan Tembelekan (Lantana camara L.) mampu mensintesis senyawa antibakteri seperti tumbuhan inangnya.
Berdasarkan hasil penelitian, isolat BT22 merupakan isolat paling potensial karena memiliki kemampuan menghambat keempat bakteri patogen yang diujikan. Selanjutnya, isolat potensial BT22 dilakukan uji lanjut untuk karakterisasi molekuler analisis sekuen 16S rRNA.
Analisis Sekuen 16S rRNA dan Rekonstruksi Pohon Filogenetik
Identifikasi bakteri endofit isolat BT22 dilakukan dengan analisis sekuen 16S rRNA. Analisis sekuen 16S rRNA merupakan tahap penting identifikasi tingkat molekuler (Singh et al. 2012). Analisis ini dapat digunakan untuk klasifikasi dan identifikasi berbasis filogenetik dengan menggunakan parameter yang tidak bergantung pada kondisi pertumbuhan dan media yang digunakan (Case et al. 2007). Menurut Singh et al. (2012), operon ribosom terutama 16S rRNA telah terbukti menjadi penanda molekuler yang stabil dan spesifik untuk identifikasi bakteri. Alasan menggunakan rRNA adalah dapat mengakses
database dengan mudah. Sifat rRNA yang sangat konservatif dapat mensintesis primer universal untuk proses PCR yang mampu melekat pada sekuen terkonservasi dari gen rRNA ketiga domain filogenetik : Archaea, Bacteria, dan
Eukarya. Daerah terkonservasi tersebut menjadi tempat perlekatan primer untuk mengamplifikasi gen 16S rRNA secara in vitro yang diisolasi dari lingkungan (Drancourt et al. 2000).
15 morfologi berbentuk batang dan banyak terdapat di dalam tanah (Niazi 2014).
Bacillus amyloliquefaciens termasuk bakteri Gram positif yang dapat menghasilkan senyawa metabolit sekunder dan berperan sebagai antimikroba. Jenis senyawa antimikroba yang mampu disintesis di antaranya senyawa antibakteri berupa poliketida (basilaen, diffisidin, dan makrolaktin), senyawa antifungi berupa lipopeptida (surfaktin, fengysin, dan basilomisin D) (He et al.
2013), siderophora (basilibaktin) (Scholz et al. 2011), dan antibiotik iturin A (Lin
et al. 2007).
Bacillus amyloliquefaciens memiliki kemampuan untuk mempercepat pertumbuhan tumbuhan dan menekan pertumbuhan fungi serta bakteri fitopatogenik (Qiao et al. 2014). Nasrin et al. (2015) dalam penelitiannya mengungkapkan bahwa senyawa metabolit sekunder yang dihasilkan oleh
Bacillus amyloliquefaciens mampu menghambat pertumbuhan Methicilin-resistant Staphylococcus aureus (MRSA). Filho et al. (2013) melaporkan bahwa
Bacillus amyloliquefaciens mampu mensintesis makromolekul protein dari alam yang dapat menginduksi daya resistensi tumbuhan tomat (Solanum lycopersicum)
terhadap bercak bakteri yang disebabkan oleh Xanthomonas vesicatoria.
Aktivitas Senyawa Antibakteri Ekstrak Kloroform Isolat Potensial BT22
Fermentasi dilakukan untuk mengisolasi senyawa metabolit sekunder pada isolat potensial BT22. Pemilihan supernatan pada hasil sentrifugasi dikarenakan senyawa antibakteri yang dihasilkan oleh isolat potensial BT22 bersifat ekstraseluler. Hal ini diperkuat dengan pernyataan Seo et al. (2010) yang menyatakan bahwa metabolit sekunder dihasilkan secara ekstraseluler sehingga dilakukan pemisahan sel bakteri dalam supernatan untuk mengekstrak metabolit sekunder. Supernatan diperoleh melalui proses sentrifugasi untuk mengendapkan sel bakteri, sehingga supernatan hanya mengandung hasil metabolisme bakteri (Savadogo et al. 2004).
16
Senyawa Antibakteri Ekstrak Kloroform Isolat BT22 dengan GC-MS
Hasil analisis GC-MS menunjukkan bahwa di dalam ektrak kloroform isolat BT22 terdapat beberapa senyawa yang berpotensi sebagai antibakteri (Tabel 5). Senyawa yang terdeteksi dalam analisis GC-MS dengan konsentrasi tertinggi adalah sikloheksanon. Jha et al. (2013) melaporkan bahwa reaksi asam aminobenzoat / aminopiridin dengan keton (asetil aseton / sikloheksanon) dan benzaldehid menghasilkan senyawa asam benzoat / piridin yang menunjukkan aktivitas antibakteri dan antifungi.
Senyawa dominan yang paling sering terdeteksi dalam analisis GC-MS adalah tetrakosan. Tetrakosan merupakan senyawa hidrokarbon alifatik (Kalegari
et al. 2011) golongan alkana (Sonibare et al. 2007). Senyawa ini belum ada laporan ilmiah terkait perannya sebagai antibakteri, namun penelitian sebelumnya telah melaporkan bahwa senyawa tetrakosan memiliki aktivitas sitotoksik terhadap sel kanker usus besar HT-29 dengan menginduksi apoptosis (Uddin et al.
2012). Hal ini menandakan bahwa meskipun belum diketahui perannya sebagai antibakteri, namun tetrakosantelah diketahui berperan dalam bidang medis. Selain itu, oktadekan juga merupakan senyawa dominan yang sering terdeteksi dalam analisis GC-MS. Oktadekan merupakan senyawa golongan alkana yang memiliki efek antimikroba, terutama terhadap S. aureus dan E. coli (Shkiri et al. 2014).
Dokosanterdeteksi sebanyak 1.76%, 3.50%, dan 1.51% pada hasil analisis GC-MS. Ekstrak T. alexandri dengan salah satu komponen senyawa utamanya dokosan bersifat antibakteri terhadap bakteri Gram-positif maupun Gram-negatif (Uma & Parvathavarthini 2010). Eikosanmerupakan senyawa yang muncul pada waktu retensi 20.240 dan 20.910 (0.51% dan 3.07%). Senyawa eikosan dari ekstrak tumbuhan Ceratonia siliqua menunjukkan aktivitas antibakteri, antifungi, serta efek sitotoksik terhadap sel kanker HeLa dan MCF-7 (Hsouna et al. 2011).
Tetratetrakontan merupakan senyawa yang terdeteksi pada waktu retensi 21.111 (1.60%) merupakan senyawa golongan alkana (Kanimozhi & Bai 2012).
Senyawa ini diketahui sebagai komponen dari daun dan biji Syzygium cumini, tumbuhan obat yang digunakan sebagai antibakteri dan antiinflamasi (Kumar et al
2011). Nonakosan terdeteksi pada waktu retensi 26.127 (1.69%) adalah senyawa golongan alkana (Kanimozhi & Bai 2012). Senyawa ini terdapat pada akar B. lancifolium dan memiliki aktivitas antibakteri terhadap Staphylococcus aureus
ATCC 6538, Staphylococcus aureus ATCC 29213, E. coli ATCC 25923, E. coli
ATCC 29988, Proteus mirabilis ATCC 43071 (Saracoglu et al. 2012).
Metil eugenolterdeteksi pada waktu retensi 16.748 (2.63%), biasa dikenal dengan nama Methyl Eugenol (ME). ME merupakan senyawa phenylpropanoid,
turunan dari phenylalanin yang banyak terkandung dalam tumbuhan obat (Tan & Nishida 2012). Pertumbuhan Campylobacter jejuni, bakteri penyebab penyakit gastroenteritis pada manusia dapat dihambat oleh tumbuhan wortel dengan kandungan minyak esensial berupa MEdan elemisin (Rossi et al. 2007).
17 Mekanisme kerja senyawa fenol sebagai zat antibakteri dengan cara meracuni protoplasma, merusak dan menembus dinding sel, serta mengendapkan protein sel bakteri. Komponen fenol juga dapat mendenaturasi enzim yang bertanggung jawab dalam germinasi spora atau berpengaruh terhadap asam amino yang terlibat dalam proses germinasi. Senyawa fenolik bermolekul besar mampu menginaktifkan enzim esensial di dalam sel bakteri meskipun dalam konsentrasi yang sangat rendah. Senyawa fenol mampu memutus senyawa peptidoglikan saat menerobos dinding sel. Ikatan peptidoglikan ini secara mekanis memberi kekuatan pada sel bakteri. Bakteri Gram negatif dengan dinding sel terdapat peptidoglikan yang sedikit sekali berada di antara selaput luar dan selaput dalam dinding sel. Dinding sel Gram negatif mengandung fosfolipid, lipopolisakarida dan lipoprotein. Setelah menerobos dinding sel, senyawa fenol akan menyebabkan kebocoran isi sel dengan cara merusak ikatan hidrofobik komponen membran sel (seperti protein dan fosfolipid) serta larutnya komponen-komponen yang berikatan secara hidrofobik yang berakibat meningkatnya permeabilitas membran. Terjadinya kerusakan pada membran sel mengakibatkan terhambatnya aktivitas dan biosintesis enzim-enzim spesifik yang diperlukan dalam reaksi metabolisme pada bakteri patogen segingga mampu menghambat pertumbuhan sel atau bahkan mematikan sel bakteri patogen tersebut (Darsana et al. 2012).
1,2- Asam benzendikarboksilat, Butil pthlat terdeteksi sebanyak 0.84% dan 2.08% merupakan senyawa golongan triterpenoid berstruktur siklik, kebanyakan berupa alkohol, aldehid, dan asam karboksilat (Suhada 2013). Senyawa ini biasa dikenal dengan nama Dibuthyl phthalate (DBP). Senyawa ini memiliki aktivitas antifungi, antibakteri, dan antimalaria (Elija et al. 2012). Triterpenoid mempunyai potensi sebagai antibakteri terhadap Escherichia coli dan
Staphylococcus aureus pada konsentrasi 1000 ppm (Sukadana et al. 2008). Kandungan triterpenoid membuktikan perannya sebagai senyawa antibakteri.
Mekanisme kerja senyawa triterpenoid dengan cara merusak membran sel bakteri karena senyawa triterpenoid bersifat lipofilik. Hal ini mengakibatkan lisisnya membran sel dan koagulasi sitoplasma dari sel bakteri (Darsana et al.
2012).
Asam heksadekanoattermasuk senyawa golongan asam palmitat terdeteksi pada waktu retensi 20.490 dengan konsentrasi 1.10%. Asam heksadekanoat merupakan komponen utama dari alga merah F. hillebrandii yang memiliki potensi sebagai antibiotik karena bersifat antibakteri terhadap Salmonella typhi, Shigella sp., dan Enterococcus faecalis (Manilal et al. 2009). Asam heksanedioat, senyawa organik yang biasa dikenal dengan asam adipat. Asam heksanedioatyang diisolasi dari larva Hermetia illucens mampu menghambat pertumbuhan bakteri patogen S. aureus, K. pneumoniae, dan Shigella dysentriae (Chu et al. 2014).
18
5
KESIMPULAN DAN SARAN
Kesimpulan
Isolat bakteri endofit BT22 yang diisolasi dari tumbuhan tembelekan (Lantana camara) mampu menghambat pertumbuhan bakteri Bacillus cereus, Escherichia coli, Staphylococcus aureus, dan Salmonella enteritidis. Berdasarkan analisis 16S rRNA, isolat BT22 memiliki persentase kemiripan dengan Bacillus amyloliquefaciens strain YB-1402 sebesar 99%. Hasil GC-MS menunjukkan bahwa senyawa aktif yang diduga berperan sebagai antibakteri dari ekstrak kloroform BT22 di antaranya : senyawa dengan konsentrasi tertinggi yaitu sikloheksanon, senyawa dominan yang terdeteksi yaitu tetrakosan dan oktadekan.
Saran
Perlu dilakukan pengukuran zona hambat ekstrak kloroform terhadap bakteri patogen berdasarkan indeks penghambatan pada uji aktivitas antibakteri untuk mengetahui kisaran indeks penghambatan ekstrak kloroform tergolong medium, resisten atau sensitif terhadap bakteri Escherichia coli, Salmonella enteritidis, Bacillus cereus, dan Staphylococcus aureus.
DAFTAR PUSTAKA
Ahamed N. 2012. Isolation and identification of secondary metabolites producing organisms from marine sponge. Discovery. 1 (1) : 14-17.
Aryantha I, Dian PL, Nurmi PDP. 2005. Mikroba Penghasil Fitohormon. Departemen Biologi: FMIPA ITB.
Asriani, Laksmi BS, Yasni S, Sudirman I. 2007. Mekanisme antibakteri metabolit
Lb. plantarum dan monoasilgliserol minyak kelapa terhadap bakteri patogen pangan. Jurnal. Teknol. dan Industri Pangan. 18 (2) : 126-133.
Baker JT, Borris RP, Carte B. 1995. Natural product drug discovery and development. New Perspective on International Collaboration. J Nat Prod. 58: 1325-1357.
Barre JT, Coll BF, Jesus JCD, Fuente JDL, Janairo VE, Ragasa GC. 1997. A bioactive triterpene from Lantana camara. Phytochemistry. 45(2): 321-4. Bolfoni MR, Ferla MDS, Sposito ODS, Giardino L, Jacinto RDC, Pappen FG.
2014. Effect of a surfactant on the antimicrobial activity of sodium hypochlorite solutions. Braz Dent J 24 (5): 416-419.
Case RJ, Boucher Y, Dahllof I, Holmstrom C, Doolittle WF, Kjelleberg S. 2007. Use of 16S rRNA and rpoB Gen as Moleculer Markers for Microbial Ecology Studies. Appl. Environ. Microbial. 73(1): 278-288.
19 from Streptomyces sp. NRRL 30566, an endophytes of Grevillea pteridifolia. FEMS Lett. 24: 183-190.
Chu KB, Jeon GC, Quan FS. 2014. Hexanedioic acid from Hermetia illucens
larvae (Diptera: Stratiomyidae) protects mice against Klebsiella pneumoniae infection. Entomological Research. 44:1-8. DOI: 10.1111/1748-5967.12043.
Claude K, Paul W, Moses J, Olwa O. 2009. The anti-mycobacterial activity of
Lantana camara a plant traditionally used to treat symptoms of
tuberculosis in Southwestern Ugand. African Health Sci. 9(1): 40-45.
Dalimartha S. 2007. Atlas tumbuhan obat Indonesia. Jakarta : Trubus Agriwidya. Darsana IGO, Besung INK, Mahatmi H. 2012. Potensi daun binahong (Anredera
cordifolia) dalam menghambat pertumbuhan bakteri Escherichia coli
secara in vitro. Jurnal Indonesia Medicus Veterinus. 1 (3) : 337-351. Deepak G, Silviya S, Kishwar HK. 2009. Biochemical Compotitions and
Antibacterial Activities of Lantana camara Plants with Yellow Lavender, Red and White Flowers. Eurasia. Jour. Bio. Sci. 3:69-77.
Desriani, Kusumawati DE, Rivai A, Hasanah N, Amrinola W, Triatna L, Sukma A. 2013. Potential endophytic bacteria for increasing paddy var rojolele productivity. Int. J. on Adv. Sci., Eng. and Information Tech. 3 (1) : 76-78.
Drancourt M, C. Bollet, A. Carlioz, R. Martelin, J.P Gayral, D. Raoult. 2000. 16S Ribosomal DNA Sequence Analysis of a Large Collection of Environtmental and Clinical Unidentifitable Bacterial Isolates. J. Clin. Microbiol. 38: 3623-3630.
Dudeja SS, Giri R. 2014. Beneficial properties, colonization, establishment and molecular diversity of endophytic bacteria in legumes and non legumes.
Afr. J. Microbiol. Res. 8(15): 1562-1572. DOI: 10.5897/AJMR2013.6541. Elija K, Vaishali B, Adsul MK, Deshpande NR, Kashalkar RV. 2012.
Antibacterial activity of Dibutyl Phthalate : A secondary metabolite isolated from Ipomoea carnea stem. Journal of Pharmacy Research 5(1). Filho RL, Souza RM, Magalhaes MM, Villela L, Zanotto E, Junior PMR, Resende
MLV. 2013. Induced defense responses in tomato against bacterial spot by protein synthesized by endophytic bacteria. Tropical Plant Pathology.
38(4): 295-302.
Garcia A, Rhoden SA, Bernardi WJ, Orlandelli RC, Azevedo JL, Pamphile JA. 2012. Antimicrobial activity of crude extracts of endophytic fungi isolated from medicinal plant Sapindus saponaria L. journal of Applied Pharmaceutical Science. 2 (10) : 35-40.
Hallmann J, Quadt-Hallmann A, Mahaffee WF, Kloepper JW. 1997. Bacterial endophytes in agricultural crops. Can J Microbiol. 43 (10): 895-914. DOI: 10.1139/m97-131.
He P, Hao K, Blom J, Ruckert C, Vater J, Mao Z, Wu Y, Hou M, He P, He Y, Borriss R. 2013. Genome sequence of the plant growth promoting strain
Bacillus amyloliquefaciens subsp. Plantarum B9601-Y2 and expression of mersacidin and other secondary metabolites. J. biotechnol. Page: 1-11. DOI: 10.1016/j.jbiotec.2012.12.014.
20
Hsouna AB, Trigui M, Mansour RB, Jarraya RM, Damak M, Jaoua S. 2011. Chemical composition, cytotoxicity effect and antimicrobial activity of
Ceratonia siliqua essential oil with preservative effects against Listeria
inoculated in minced beef meat. International Journal of Food Microbiology. 148: 66-72. DOI: 10.1016/j.ijfoodmicro.2011.04.028. Jha A, Yashmeen S, Kumar DN. 2013. An innovative green synthesis of some
Schiff bases and their antimicrobial activity. Int J Pharm Bio Sci. 4(4) : 197-204.
Junanto T, Sutarno, Supriyadi. 2008. Aktivitas antimikroba estrak Angsana (Pterocarpus indicus) terhadap Bacillus subtilis dan Klebsiella pneumoniae. Bioteknologi 5(2): 63-69.
Kalegari M, Miguel MD, Dias JDFG, Lordello ALL, Lima CPD, Miyazaki CMS, Zanin SMW, Verdam MCDS, Miguel OG. 2011. Phytochemical constituents and preliminary toxicity evaluation of leaves from Rourea induta Planch. (Connaraceae). Brazilian Journal of Pharmaceutical Sciences. 47(3): 635-642.
Kanimozhi D, Bai VR. 2012. Analysis of bioactive components of ethanolic extract of Coriandrum sativum L. International Journal of Research in Pharmacy and Science. 2(3): 97-110.
Kannan R, Damodaran T, Umamaheswari S. 2015. Sodicity tolerant polyembrionic mango root stock plants: A putative role of endophytic bacteria. Afr. J. Biotechnol. 14(4): 350-359. DOI: 10.5897/AJB2014.14259.
Katzung BG, Lofholn PW. 2004. Penentuan Penggunaan Obat dan Penggunaan Resep Secara Rasional dalam Farmakologi Dasar dan Klinik. Buku 3. Edisi Kedelapan. Jakarta: Penerbit Salemba Medika. p 609.
Kumar V, Bhatnagar AK, Srivastava JN. 2011. Antibacterial activity of crude extracts of Spirulina platensis and its structural elucidation of bioactive compound. J. Med. Plants Res. 5(32): 7043-7048. DOI: 10.5897/JMPR11.1175.
Kusumawati DE. 2014. Isolasi dan Karakterisasi Senyawa Antibakteri dari Bakteri Endofit Tumbuhan Miana (Coleus scutellariodes [L.] Benth.) [Tesis]. Bogor : Institut Pertanian Bogor.
Lamb TG, Tonkyn DW, Kluepfel DA. 1996. Movement of Pseudomonas aureofaciens from the rhizosphere to aerial plant tissue. Can. J. Microbiol.
42:1112-1120.
Larran S, Monaco C, Alippi HE. 2001. Endophytic fungi in leaves of
Lycopersicon esculentum. World J. Microbiol. Biotechnol. 17 (2): 181-184.
Lin HY, Rao YK, Tzeng YM. 2007. Ferrous ion enhanced lipopeptide antibiotic Iturin A production from Bacillus amyloliquefaciens B128. Int. J. Appl. Sci. Eng. 5(2): 123-132.
21 Manilal A, Sujith S, Selvin J, Shakir C, Kiran GS. 2009. Antibacterial activity of
Falkenbergia hillebrandii (Born) from the Indian coast against human pathogens. International Journal of Experimental Botany. 78:161-166. Mano H, Tanaka F, Nakamura C, Kaba H, Morisaki H. 2007. Culturable
endophytic bacterial flora of the meaturing leaves and roots of Rice plants (Oryza sativa) cultivated in a paddy field. Microbes Environmental. 22(2): 175-185.
Mardiastuti HW, Anis K, Ariyani K, Ikaningsih, Retno K. 2007. Emerging resistance pathogen: situasi terkini di Asia, Eropa, Amerika Serikat, Timur Tengah, dan Indonesia. Maj. Kedokt. Indon. 57 (3) : 75-79.
Mary KV. 2011. Studies on phytochemical screening and antibacterial activities of Lantanacamara. Plant. Sci. Feed. 1: 74-79.
Mothana RA, Lindequist U. 2005. Antimicrobial Activities of some of the island Soqotra. J Ethnopharmacol. 96:177-181.
Nascimento GGF, Locatelli J, Paulo CF, Giuiliana LS. 2000. Antibacterial activity of plant extracts and phytochemicals on antibiotic resistant bacteria. Brazilian Journal of Microbiology. 31: 247-256.
Nasrin S, Hossain MJ, Liles MR. 2015. Draft genome sequence of Bacillus amyloliquefaciens AP183 with anti-bacterial activity against Methicillin-Resistant Staphylococcus aureus. JournalsASM.org 3(2): 1-2.
Niazi A. 2014. Genome-wide analyses of Bacillus amyloliquefaciens strain provide insights into their beneficial role on plants [Disertasi]. Uppsala: Swedish University of Agricultural Sciences.
Prasetyoputri A, Ines A. 2006. Mikroba endofit sumber acuan baru yang berpotensi. Biotrend. 1(2): 13-15.
Proestos C, Chorianopoulos N, Nychas GJ, Komaitis M. 2005. RPHPLC analysis of the phenolic compounds of plant extracts. Investigation of their antioxidant capacity and antimicrobial activity. J Agric Food Chem.
53:1190–1195.
Qiao JQ, Wu HJ, Huo R, Gao XW, Borriss R. 2014. Stimulation of plant growth and biocontrol by Bacillus amyloliquefaciens subsp. Plantarum FZB42 engineered for improved action. Chemical and Biological Technologies in Agriculture. 1(12): 1-14.
Radji M. 2005. Peranan Bioteknologi dan Mikroba Endofit dalam Pengembangan Obat Herbal. Majalah Ilmu Kefarmasian. 3: 113-126.
Rossi PG, Bao L, Luciani A, Panighi J, Desjobert JM, Costa J, Casanova J, Bolla JM, Berti L. 2007. (E)-Methylisoeugenol and Elemicin: Antibacterial Components of Daucus carota L. Essential Oil against Campylobacter jejuni. J. Agri. Food Chem. 55(18): 7332-7336. DOI: 10.1021/jf070674u. Roy S, Rao K, Bhuvaneswari CH, Giri A, Mangamoori LN. 2010. Phytochemical
analysis of Andrographis paniculata extract and its antimicrobial activity.
World J Microbiol Biotechnol. 26: 85-91. DOI: 10.1007/s11274-009-0146-8.
Ryan RP, Germaine K, Franks A, Ryan DJ, Dowling DN. 2008. Minireview: Bacterial endophytes: recent development and application. FEMS Microbiol Lett. 278: 1-9.
22
Bupleurum L. species. Afr. J. Microbiol. Res. 6(12): 2899-2908. DOI: 10.5897/AJMR11.1097.
Savadogo A, Cheik ATO, Imael HNB, Alfred ST. 2004. Antimicrobial Activities of Lactic Acid Bacteria Strains Isolated from Burkina Faso Fermented Milk. Pakistan Journal of Nutrition. 3 (3): 174-179.
Scholz R, Molohon KJ, Nachtigall J, Vater J, Markley AL, Susmuth RD, Mitchell DA, Borriss R. 2011. Plantazolicin, a novel Microcin B17/Streptolysin S-like natural product from Bacillus amyloliquefaciens FZB42. J. Bacteriol.
193(1): 215-224. DOI: 10.1128/JB.00784-10.
Seo WT, Lim WJ, Kim EJ, Yun HD, Lee YH, Cho KM. 2010. Endophytic bacterial diversity in the Young Radish and their antimicrobial activity against pathogens. J. Korean Soc. Appl. Biol. Chem. 53(4): 493-503. Shittu LAJ, Bankole MA, Ahmed T, Bankole MN, Shittu RK, Saalu CL, Ashiru
OA. 2007. Antibacterial and antifungal activities of essential oils of crude extracts of Sesame radiatum against some common pathogenic microorganisms. Iran J Pharmacol Ther 6:165–170.
Shkiri FH, Soussi LSR, Zakhama AEA, Mahjoub A, Flamini G, Jannet HB. 2014. Chemical composition and antibacterial activity of essential oils from the tunisian Allium nigrum L. EXCLI Journal. 13: 526-535.
Silva WJD, Rached RN, Rosalen PL, Del Bel Cury AA. 2008. Effects of Nystatin, Fluconazole and Propolis on Poly(Methyl Methacrylate) Resin Surface.
Braz Dent J 19(3): 190-196.
Simarmata R, Lekatompessy S, Sukiman H. 2007. Isolasi mikroba endofit dari tumbuhan obat sambung nyawa (Gymura procumbens) dan analisis potensinya sebagai antimikroba. Berk Penel Hayati 13 : 85-90.
Singh V, Chaudhary DK, Mani I. 2012. Molecular characterization and modeling of secondary structure of 16s rrna from Aeromonas veronii. International Journal of Applied Biology and Pharmaceutical Technology. 3 (1): 253-260.
Son R, Cheah YK. 2002. Preliminary screening of endophytic fungi from medical plants in Malaysia for antimicrobial and antitumor activity. Malaysian Journal of Medical Sciences 9(2): 23-33.
Sonibare MA, Soladoye MO, Ogunlana YE. 2007. A chemotaxonomic approach to the alkane content of three species of Anthocleista Afzel. (Loganiaceae).
Afr. J. Biotechnol. 6(13): 1516-1520.
Strobel GA, Woapong EFJ, Harper JK, Arif AM, Grant DM, Fung PCW, Chan K. 2002. Isopestacin, an isobenzopuranone from Pestalotiopsis microspora,
possesing antifungal and antioxidant activities. Phytochemistry. 60: 179-183.
Strobel GA, Daisy. 2003. Bioprospecting for Microbial Endophytess and Their Natural Product. Microbiology and Moleculer Biology Review. 67: 491-502.
23 Sukadana IM, Santi SR, Juliarti NK. 2008. Aktivitas antibakteri senyawa golongan triterpenoid dari biji Pepaya (Carica papaya L.). Jurnal Kimia.
2(1): 15-18.
Sylvia D, Fuhrmann J, Hartel P, Zuberer D. 2005. Principles and Application of Soil Microbiology. Pearson Education Inc. New Jersey.
Tan KH, Nishida R. 2012. Methyl eugenol: Its occurrence, distribution, and role in nature, especially in relation to insect behavior and pollination. Journal of Insect Science. 12(56): 1-74.
Uddin SJ, Grice D, Tiralongo E. 2012. Evaluation of cytotoxic actvity of patriscabratine, tetracosane, and various flavonoids isolated from the Bangladeshi medicinal plant Acrotichum auruem. Pharmaceutical Biology. 50(10): 1276-1280. DOI: 10.3109/13880209.2012.673628. Uma B, Parvathavarthini R. 2010. Antibacterial effect of hexane extract of sea
Urchin, Temnopleurus alexandri (Bell,1884). Int. J. PharmTech Res. 2(3): 1677-1680.
Verma RK, Verma SK. 2006. Phytochemical and termiticidal studies of Lantana camara var aculeata leaves. Fitoterapia. 77: 466-468.
Zahro L, Agustini R. 2013. Uji efektivitas antibakteri ekstrak kasar saponin jamur tiram putih (Pleurotus ostreatus) terhadap S. aureus dan E. coli. Journal of chemistry. 2 (2) : 120 – 129.
Zhang B, Salituro G, Szalkowski D, Li Z, Zhang Y, Royo I, Vilella D, Dez M, Pelaes F, Ruby C, Kendall RL, Mao X, Griffin P, Calaycay J, Zierath JR, Heck JV, Smith RG, Moller DE. 1999. Discovery of small molecule insulin mimetic with antidiabetic activity in mice. Science. 284: 974-981. Zinniel DK, Lambrecht P, Harris BN, Feng Z, Kuczmarski D, Higley P, Ishimaru
24
25 Lampiran 1 Bagan Alir Penelitian
Tumbuhan Lantana camara L.
Isolasi bakteri endofit
Isolat bakteri endofit Isolat bakteri uji
Regenerasi bakteri endofit dan bakteri uji
Uji aktivitas antibakteri endofit terhadap bakteri uji
Isolat potensial
Analisis sekuen 16S rRNA Fermentasi
Sentrifugasi
Pelet Supernatan
Ekstraksi dengan kloroform
Ekstrak kloroform
Uji antibakteri terhadap bakteri uji Analisis GC - MS
Senyawa antibakteri Spesies bakteri endofit
26
27 Lampiran 3 Diameter zona hambat yang terbentuk pada isolat bakteri endofit
terhadap bakteri patogen
Keterangan : V = terdapat zona hambat namun sangat kecil dan sulit terukur
No Kode Diameter zona hambat (mm)
isolat B. cereus E. coli S. aureus S.enteritidis
1 AT11 - - - -
2 AT12 - - - -
3 AT13 - - - -
4 AT21 - - - -
5 AT22 - V - -
6 AT23 - - - -
7 AT24 - - - -
8 AT3 - V - V
9 AT4 - - - -
10 AT5 - - - -
11 AT6 - - - -
12 AT7 - - - -
13 AT8 - - - -
14 BT11 - - - -
15 BT12 - - - -
16 BT21 2 3 V -
17 BT22 3 V 8 5
18 BT3 - - - -
19 DT1 - - - -
20 DT11 - 1 3 1
28
Lampiran 4 Sekuen 16S rRNA isolat potensial BT22 >BT22_518F
29 Lampiran 5 Hasil BLAST Sekuen 16S rRNA isolat potensial BT22 hasil
30
31 Lampiran 7 Kondisi operasional alat GC-MS
Merk : Shimadzu Type GCMS-QP2010
Gas : Helium
Detektor : MS (Mass Spectrometer)
Kolom : Capiler Type Phase Rtx-5MS ; 60 M; 0.25 mmID
Suhu kolom : 50 ºC
Column Flow : 0.85 mL/menit
Split Ratio : 112.3 SPL Temperature : 280 ºC MS Interface : 280 ºC
Ion Source : 200 ºC
32
33
RIWAYAT HIDUP
Penulis dilahirkan di Sragen, 3 Juli 1991 dari Ayah Drs. H. Jumari, M.H dan Ibu Hj. Herning Sutati. Penulis merupakan putri pertama dari tiga bersaudara. Penulis menyelesaikan pendidikan menengah atas di SMA Negeri 1 Gemolong, Sragen, Jawa Tengah tahun 2009. Penulis meneruskan pendidikan di Program Studi Pendidikan Biologi Fakultas Keguruan dan Ilmu Pendidikan, Universitas Sebelas Maret (UNS), Solo tahun 2009 dan penulis mendapatkan gelar sarjana (S1) pada tahun 2013. Tahun 2013 penulis melanjutkan studinya dan diterima di Departemen Biokimia, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor (IPB), Bogor dengan Beasiswa Pendidikan Pascasarjana Dalam Negeri (BPPDN) Direktorat Jenderal Pendidikan Tinggi (DIKTI) 2013.
Selama mengikuti perkuliahan penulis sering mengikuti pelatihan dan seminar yang diadakan baik di dalam kampus maupun di luar kampus. Penulis pernah menjadi panitia dalam Seminar Nasional X Pendidikan Biologi Volume II dengan tema “Biologi, Sains, Lingkungan, dan Pembelajarannya” yang diadakan di UNS tahun 2013. Selama perkuliahan, penulis juga aktif dalam kegiatan organisasi kemahasiswaan UNS sebagai Pengurus Himpunan Mahasiswa Biologi (HIMABIO) tahun (2009/2010, 2010/2011). Penulis juga pernah menjadi asisten mata kuliah praktikum Genetika (2011/2012) dan Biokimia (2010/2011, 2011/2012) di Laboratorium Pendidikan Biologi UNS.
Selama mengikuti perkuliahan di Pascasarjana IPB, penulis pernah mengikuti kegiatan Seminar Internasional SEAMEO BIOTROP dengan tema “Mega Diversity in the Tropics of Southest Asia : Why and How to Regard Them?”. Penulis telah menulis jurnal yang berjudul Isolasi dan Karakterisasi dengan Analisis 16S rRNA Bakteri Endofit dari Tumbuhan Tembelekan (Lantana camara L.) sebagai Penghasil Senyawa Antibakteri dan dipublikasikan di jurnal