ALUMINIUM DAN DEFISIENSI FOSFOR DI TANAH MASAM
KARLIN AGUSTINA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi ”Fisiologi Adaptasi Sorgum (Sorghum bicolor (L.) Moench) terhadap Toksisitas Aluminium dan Defisiensi Fosfor di Tanah Masam” merupakan hasil penelitian saya dengan arahan dari komisi pembimbing. Disertasi ini disusun dari sebagian hasil penelitian payung Hibah Penelitian Tim Pascasarjana (HPTP) yang berjudul ”Pengembangan Sorgum Manis (Sorghum bicolor L. Moench) untuk Bioetanol di Lahan Kering Bertanah Masam: Fisiologi, Genetika dan Pemuliaan” yang diketuai oleh Dr. Ir. Trikoesoemaningtyas, M.Sc. Disertasi ini belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Juli 2011
(L.)Moench) to Aluminum Toxicity and Phosphorous Deficiency in Acid Soils. Under direction of DIDY SOPANDIE, TRIKOESOEMANINGTYAS, and DESTA WIRNAS.
Sorghum (Sorghum bicolor (L) Moench) is an ideal crop to be developed as food diversification and source of bioethanol in Indonesia because this crop has high adaptability to be grown in dry lands. From a total of 148.2 millions ha dry land in Indonesia, an estimated are of 108 millions ha are acid soil (ultisols). Acid soils have low fertility and Al toxicity which are major constraints for crop production in acid soil. For sorghum production in acid dry lands, sorghum varieties having high adaptation to acid soil have to be developed. The objectives of this study was to obtain complete information about the physiological
adaptation to aluminum toxicity and phosphorus deficiency which can be used as the character selection in breeding program to obtain the tolerant sorghum in acid soils. The genotypes used was Numbu (Tolerant), ZH-30-29-07 (Tolerant), B-69 (sensitive) and B-75 (sensitive) from selected on acid soils in Lampung. The study on physiological mechanism of sorghum adaptation to acid soils has
resulted in information that the main limiting factor in soghum growth in acid soil was phosphorous deficiency. Under sufficient P, without liming, the sensitive genotype produced similar yield. The tolerant genotype, Numbu and ZH30-29-07 showed higher ability to maintain root growth under Al stress and P deficiency. The tolerant genotypes also showed higher ability to absorp nutient under low P condition compared to sensitive genotypes. The lowest concentration of 74 μM Al can reduce the growth in all genotypes tested. The longer the sorghum roots exposed to the stress, the higher the value of scoring and the accumulation of aluminum in the roots under Al toxicity. Sorghum genotypes had specific absorption rate was high, but it was not followed by increasing of roots length, roots dry weight and shoot dry weight. Specific absorption rate is more affected by Al stress rather than increasing level of P. Genotype ZH-30-29-07 were tolerant in identified in acid soils showed the moderate response to Al toxicity in nutrient solution. The higher aluminum concentration leading to greater plant growth inhibition. The tolerant genotypes had the internal mechanism (tolerance) in the face under low P condition by increasing the efficiency of internal
phosphor (interrelated), otherwise the sensitive genotype had the external
mechanism (avoidance) through increased P absorption or total P . There was no relationship between the use efficiency of P with total P content of plant both in testing using the acid soils in rhizotron and nutrient solution.
KARLIN AGUSTINA. Fisiologi Adaptasi Sorgum (Sorghum bicolor (L) Moench) Terhadap Toksisitas Aluminium dan Defisiensi Fosfor di Tanah Masam. Dibimbing oleh DIDY SOPANDIE, TRIKOESOEMANINGTYAS dan DESTA WIRNAS.
Sorgum (Sorghum bicolor (L) Moench) ideal dikembangkan sebagai sumber bahan makanan untuk diversifikasi pangan dan bahan baku bioetanol di Indonesia, karena tanaman ini mempunyai kesesuaian yang tinggi untuk dikembangkan di lahan kering. Dari total luas lahan Indonesia yang mencapai 188.2 juta ha,148 juta ha di antaranya merupakan lahan kering yang diperkirakan 102.8 juta ha di antaranya berupa lahan kering bertanah masam (ultisols). Lahan-lahan bertanah masam mempunyai tingkat kesuburan tanah yang rendah dan keracunan Al yang menjadi kendala dalam produksi tanaman. Untuk mencapai tujuan pengembangan sorgum di lahan kering bertanah masam diperlukan upaya mengembangkan varietas-varietas sorgum yang beradaptasi pada kondisi agroekologi lahan kering bertanah masam.
Tujuan penelitian adalah (1) untuk memperoleh informasi tentang tanggap agronomi genotipe sorgum terhadap pertumbuhan dan produksi pada kondisi cekaman Al dan defisiensi fosfor di tanah masam, (2) memperoleh informasi tentang adaptasi sorgum terhadap cekaman Al di larutan hara, (3) memperoleh informasi tentang tanggap morfofisiologi akar sorgum di tanah masam dalam rhizotron, (4) memperoleh informasi tentang mekanisme yang mungkin terjadi yang dapat mendasari perbedaan efisiensi P dalam keadaan tercekam Al baik melalui penilaian serapan P maupun penilaian penggunaan P, (5) memperoleh informasi mengenai distribusi dan akumulasi Al dalam akar sorgum serta kemampuan akar memperbaiki kerusakan akibat cekaman Al, dan (6) mengidentifikasi karakter-karakter tanaman yang berkaitan dengan toleransi terhadap toksisitas Al dan defisiensi P, yang dapat digunakan sebagai karakter untuk seleksi dalam rangka pengembangan tanaman sorgum di tanah masam.
Penelitian dilaksanakan secara terpadu mulai dari lapangan, rumah kaca dan laboratorium. Penelitian lapang dilaksanakan di tanah kering masam di Jasinga Kabupaten Bogor. Rancangan penelitian yang digunakan adalah split plot dengan tiga ulangan. Faktor perlakuan yang digunakan adalah kondisi tanah masam dengan Al tinggi dan Al rendah serta genotipe sorgum. Genotipe yang digunakan adalah dua genotipe toleran yaitu Numbu dan ZH-30-29-07 serta dua genotipe peka yaitu B-69 dan B-75 hasil seleksi Sungkono (2007) di tanah masam Lampung.
Hasil penelitian menunjukkan cekaman Al dan defisiensi P merupakan faktor pembatas pertumbuhan di tanah masam. Pemberian kapur saja tanpa diikuti peningkatan dosis pupuk P hanya mampu meningkatkan bobot kering tajuk dan komponen hasil sorgum di tanah masam untuk genotipe toleran, tetapi untuk genotipe peka harus diikuti dengan peningkatan dosis pupuk P. Bobot kering tanah masam menunjukkan respon moderat terhadap toksisitas Al di larutan hara. Semakin tinggi konsentrasi Al, semakin besar hambatan pertumbuhan tanaman.
Pengujian menggunakan rhizotron menunjukkan diameter penyebaran akar yang tidak berbeda antara sorgum toleran dan peka pada kondisi optimal. Genotipe B-75 memiliki kadar P jaringan tinggi pada media tanah masam, akan tetapi nilai efisiensi penggunaan hara P nya lebih rendah daripada tanaman toleran. Hasil analisis kadar P jaringan dan efisiensi penggunaan ini menunjukkan bahwa genotipe toleran memiliki mekanisme internal (toleransi) dalam menghadapi cekaman P rendah dengan meningkatkan efisiensi penggunaan P internal (interrelated), sebaliknya genotipe peka memiliki mekanisme eksternal (penghindaran) melalui peningkatan serapan P atau kadar P total jaringan. Kadar P jaringan dan efisiensi penggunaan P berkorelasi tinggi dengan diameter akar dan pembentukan biomassa tanaman.
Pada kondisi bercekaman Al, genotipe sorgum memiliki laju serapan spesifik P yang tinggi, tetapi tidak diikuti oleh adanya peningkatan panjang dan bobot kering akar maupun tajuk. Genotipe peka memiliki total serapan P tinggi, tetapi memiliki efisiensi penggunaan P lebih rendah daripada genotipe toleran. Hasil ini sejalan dengan pengujian kemampuan memanfaatkan hara P pada media tanah masam. Laju serapan spesifik P lebih dipengaruhi oleh cekaman Al daripada peningkatan dosis P. Tidak ditemukan adanya hubungan antara efisiensi penggunaan P dengan kadar P total jaringan tanaman baik pada pengujian menggunakan media tanah masam dalam rhizotron maupun di larutan hara.
Metode pewarnaan hematoksilin dengan pengukuran intensitas pewarnaan melalui histokimia (sediaan mikroskopis) untuk melihat penetrasi dan distribusi Al ke dalam akar, dapat digunakan untuk melihat perbedaan toleransi sorgum terhadap Al. Batas konsentrasi tertinggi yang masih bisa ditolerir oleh genotipe sorgum adalah 74 μM Al dengan lama cekaman 24 jam. Semakin lama akar sorgum diberi cekaman, semakin tinggi nilai skoring dan akumulasi Al pada akar.
Cekaman Al tidak mempengaruhi munculnya akar sekunder pada genotipe toleran, tetapi sangat menghambat pertambahan panjang akar sekunder yang muncul pada perkembangan selanjutnya. Kerusakan akar akibat peningkatan konsentrasi Al hingga 148 μM masih mampu di recovery oleh Numbu, tetapi genotipe ZH-30-29-07, B-69 dan B-75 hanya mampu memunculkan kembali akar sekundernya pada konsentrasi cekaman Al 74 μM. Semakin tinggi konsentrasi cekaman Al, semakin berkurang kemampuan genotipe sorgum dalam menumbuhkan kembali akarnya.
© Hak Cipta milik Institut Pertanian Bogor, tahun 2011 Hak Cipta dilindungi Undang-undang
ALUMINIUM DAN DEFISIENSI FOSFOR DI TANAH MASAM
KARLIN AGUSTINA A.262070021/AGH
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor
pada Mayor Agronomi dan Hortikultura
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi pada Ujian Tertutup: 1. Dr. Ir. Ahmad Junaedi, M.Si
2. Dr. Ir. Komaruddin Idris, MS
Penguji Luar Komisi pada Ujian Terbuka: 1. Prof (R). Dr. Ir. Abdul Karim Makarim, M.Sc
Nama Mahasiswa : Karlin Agustina
NRP : A262070021
Mayor : Agronomi dan Hortikultura
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Didy Sopandie, MAgr. Ketua
Dr. Ir. Trikoesoemaningtyas, M.Sc Dr. Desta Wirnas, SP, M.Si Anggota Anggota
Diketahui
Koordinator Mayor Dekan Sekolah Pascasarjana Agronomi dan Hortikultura
Dr. Ir. Munif Ghulamahdi, MS Dr. Ir. Dahrul Syah, MSc.Agr
Alhamdulillahirabbil ’alamin, segala puji syukur penulis panjatkn kehadirat Allah SWT yang telah melimpahkan rahmat dan karuniaNya sehingga
karya ilmiah ini berhasil diselesaikan.
Disertasi berjudul Fisiologi Adaptasi Sorgum (Sorghum Bicolor L. Moench) terhadap Toksisitas Aluminium dan Defisiensi Fosfor di Tanah Masam disusun berdasarkan 6 (enam) topik hasil penelitian yang dilaksanakan secara terpadu. Keenam topik ini merupakan satu kesatuan penelitian Land to Lab yang bertujuan untuk mendapatkan informasi dari bidang kajian fisiologi tentang
kemampuan sorgum beradaptasi pada kondisi tercekam Al dan defisiensi P.
Disertasi ini disusun dari sebagian hasil penelitian payung Hibah Penelitian Tim Pascasarjana (HPTP) yang berjudul ”Pengembangan Sorgum Manis (Sorghum bicolor L. Moench) untuk Bioetanol di Lahan Kering Bertanah Masam: Fisiologi, Genetika dan Pemuliaan” yang diketuai oleh Dr.Ir. Trikoesoemaningtyas, M.Sc. Hasil penelitian ini telah dipublikasikan pada jurnal
Agronomi Indonesia (terakreditasi) No.38 (2):88-94 (2010). Telah dipresentasikan
pada Seminar Nasional PERAGI di Medan pada bulan Oktober 2009 dan
memenangkan dana Hibah untuk diterbitkan pada jurnal berskala Internasional.
Telah dipresentasikan juga di Maros Sulawesi Selatan pada acara Pekan Serealia
Nasional I bulan Juli 2010. Topik penelitian mengenai análisis root regrowth dan uji pewarnaan hematoksilin juga telah terseleksi oleh DP2M DIKTI sebagai
penelitian berpotensi paten dan telah diikutsertakan dalam program Pelatihan
Pemanfaatan Hasil Penelitian, Pengabdian kepada Masyarakat, dan Kreativitas
Mahasiswa Berpotensi Paten yang diselenggarakan pada tanggal 10 – 12 Maret 2011 di Jakarta.
Dalam pelaksanaan penelitian dan penyelesaian studi, penulis banyak
mendapat bantuan baik dari perorangan maupun lembaga atau instansi tertentu.
Oleh karena itu, penulis mengucapkan terima kasih dan penghargaan secara
khusus kepada komisi pembimbing yang diketuai oleh Bapak Prof. Dr. Ir. Didy
Sopandie, M.Agr dan beranggotakan Ibu Dr. Ir. Trikoesoemaningtyas, M.Sc dan
Terima kasih disampaikan juga kepada Prof. Dr. Ir. Sudirman Yahya dan
Dr. Ir. Anas D.Susila atas kesediaan sebagai penguji luar komisi pada ujian
prakuallifikasi. Kepada Dr. Ir. Ahmad Junaedi, M.Si dan Dr. Ir. Komaruddin Idris,
MS disampaikan terima kasih atas kesediaan sebagai penguji luar komisi pada
ujian tertutup, dan kepada Prof (R). Dr. Ir. Abdul Karim Makarim, M.Sc dan Dr.
Ir. Miftahudin, M.Si sebagai penguji luar komisi pada ujian terbuka.
Terima kasih juga disampaikan kepada DITJENDIKTI KEMDIKNAS atas
bantuan beasiswa BPPS, bantuan biaya penelitian dan penyelesaian studi melalui
program Hibah Penelitian Tim Pascasarjan (HPTP) dan program Hibah Disertasi
Doktor tahun 2010. Ucapan yang sama juga disampaikan kepada Pemda Provinsi
Sumatera Selatan atas dukungan dana penelitian dan penyelesaian studi melalui
program Beasiswa Kemitraan Daerah tahun 2010-2011.
Ucapan terima kasih juga disampaikan kepada PATIR-BATAN, UPTD
Lahan kering Tenjo, Kebun percobaan University Farm IPB, Laboratorium RGCI,
Laboratorium Micro Tehnique IPB, Laboratorium Puslit Tanah dan Laboratorium
SEAMEO BIOTROP sebagai unit kerja dan lembaga yang telah banyak
membantu terlaksananya penelitian ini dengan baik dan lancar.
Kepada Pengurus Yayasan IBA, Rektor Universitas IBA dan Dekan
Fakultas Pertanian disampaikan ucapan terima kasih atas izin dan kesempatan
untuk melanjutkan studi di Sekolah Pascasarjana Institut Pertanian Bogor. Ucapan
serupa disampaikan kepada teman-teman satu tim penelitian sorgum: Sungkono,
Isnaini, Rahmansyah Dermawan, Sumiyati dan Winda yang telah banyak
membantu pelaksanaan penelitian dan bertukar informasi hasil penelitian.
Akhirnya kepada Ibu Misdariyah dan Ayah Bastomi Amin kedua orang
tuaku, terima kasih tak terhingga atas kasih sayang dan doa yang tak pernah putus.
Kepada Ade Munirwan, SE suamiku, dan anak-anakku Agung Anggana Ajie dan
Zhafir Afla Raihan terima kasih atas segala kesabaran, keikhlasan dorongan serta
doa kalian. Terima kasih juga disampaikan kepada semua saudaraku dan semua
pihak yang telah membantu dalam penelitian dan penulisan disertasi.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juli 2011
Penulis dilahirkan di Palembang pada tanggal 13 Agustus 1968 merupakan
anak kedua dari lima bersaudara dari pasangan Bapak Bastomi Amin dan Ibu
Misdaryah. Menikah dengan Ade Munirwan,SE dan dikaruniai dua anak
laki-laki, Agung Anggana Ajie (17 tahun) dan Zhafir Afla Raihan (8 tahun).
Penulis menamatkan pendidikan formal di SDN 134 Palembang tahun
1981, SMPN 4 Palembang tahun 1984, dan SMAN 5 Palembang pada tahun 1987.
Penulis mendapatkan gelar Sarjana Pertanian (Ir.) di Fakultas Pertanian
Universitas IBA Palembang dengan bidang Budidaya Pertanian pada tahun 1992.
Tahun 1995 penulis melanjutkan pendidikan ke Program Pascasarjana Institut
Pertanian Bogor pada Program Studi Agronomi, dan menyelesaikannya pada
tahun 1998. Tahun 2007 penulis melanjutkan pendidikan Program Doktor di
Sekolah Pascasarjana Institut Pertanian Bogor dengan biaya pendidikan dari BPPS
DITJENDIKTI.
Penulis bekerja sebagai staf pengajar Kopertis Wilayah II Palembang
dipekerjakan di Fakultas Pertanian Universitas IBA Palembang pada program
studi Agroteknologi sejak tahun 1994.
Selama mengikuti Program S3, penulis pernah menjadi bendahara Forum
Mahasiswa Pascasarjana (FORSCA) Departeman AGH dan bendahara pada
FORUM WACANA IPB periode 2008-2009. Pada bulan Juli 2009 penulis juga
Halaman Pengaruh Cekaman Aluminium terhadap Tanaman ... Mekanisme Fisiologi Tanaman terhadap Cekaman Aluminium... Interaksi Fosfor dengan Aluminium... Fungsi dan Gejala Defisiensi Fosfor ... Mekanisme Toleransi Tanaman terhadap Defisiensi Fosfor ...
7
TANGGAP AGRONOMI SORGUM (Sorghum bicolor L. Moench) TERHADAP CEKAMAN ALUMINIUM DAN DEFISIENSI FOSFOR DI TANAH MASAM... TOLERANSI SORGUM (Sorghum bicolor L. Moench) TERHADAP
LAJU SERAPAN SPESIFIK PADA SORGUM (Sorghum bicolor L. Moench) DALAM KONDISI BERCEKAMAN ALUMINIUM DAN DEFISIENSI FOSFOR DI LARUTAN HARA ...
SORGUM (Sorghum bicolor L. Moench) MELALUI UJI PEWARNAAN HEMATOKSILIN... ...
ANALISIS ROOT REGROWTH AKAR SORGUM (Sorghum bicolor L. Moench) TERHADAP CEKAMAN ALUMINIUM DI LARUTAN
1 Jenis-jenis asam organik yang dieksudasi tanaman dalam
hubungannya dengan toleransi terhadap Al... 11
2 Respon tanaman terhadap Defisiensi P... 20
1.1 Ciri kimia dan fisika contoh tanah pada lokasi penelitian ... 30
1.2 pitu Rekapitulasi analisis ragam pengaruh kondisi cekaman, genotipe dan interaksi antara kondisi cekaman dan genotipe terhadap pertumbuhan dan produksi sorgum di tanah
masam... 32
1.3 Respon genotipe sorgum untuk karakter bobot kering tajuk pada berbagai kondisi cekaman ... 34
1.4 Respon genotipe sorgum untuk karakter bobot kering tajuk pada kondisi cekaman Al tinggi di tanah masam ... 35
1.5 Respon genotipe sorgum untuk karakter bobot kering tajuk pada kondisi cekaman Al rendah di tanah masam... 35
1.6 Respon genotipe sorgum untuk karakter bobot kering tajuk terhadap pemberian pupuk P pada kondisi cekaman Al tinggi... 36
1.7 Respon genotipe sorgum untuk karakter bobot kering tajuk terhadap pemberian pupuk P pada kondisi cekaman Al rendah.. 36
1.8 Respon genotipe sorgum untuk karakter bobot biji per tanaman pada berbagai kondisi cekaman di tanah masam... 37
1.9
Respon genotipe sorgum untuk karakter bobot biji pertanaman pada kondisi cekaman Al tinggi di tanah masam ... 38
1.10 Respon genotipe sorgum untuk karakter bobot biji pertanaman pada kondisi cekaman Al rendah di tanah masam... 38
1.11 Respon genotipe sorgum untuk karakter bobot biji per tanaman terhadap pemberian pupuk P pada kondisi cekaman Al tinggi .. 39
1.13
Respon genotipe pada bobot 1000 biji sorgum terhadap berbagai kondisi cekaman Al dan pupuk P di tanah masam 40
1.14
1.15
Respon genotipe untuk karakter bobot 1000 biji terhadap pemberian pupuk P dengan kondisi cekaman Al tinggi...
Respon genotipe untuk karakter bobot 1000 biji terhadap pemberian pupuk P dengan kondisi cekaman Al rendah...
40
40
1.16 Respon genotipe pada berbagai kondisi cekaman terhadap padatan terlarut (PTT) dan gula total ... 41
Respon genotipe sorgum toleran dan peka terhadap kandungan gula total dan padatan terlarut total (PTT) batang sorgum di
Nilai korelasi antara peubah pertumbuhan, produksi dan padatan terlarut total sorgum di tanah masam...
42
43
2.1 Rekapitulasi nilai analisis ragam pengaruh genotipe, konsentrasi Al dan pengaruh interaksi antara pengaruh genotipe dan konsentrasi Al terhadap pertumbuhan pertumbuhan sorgum
di larutan hara ……….. 50
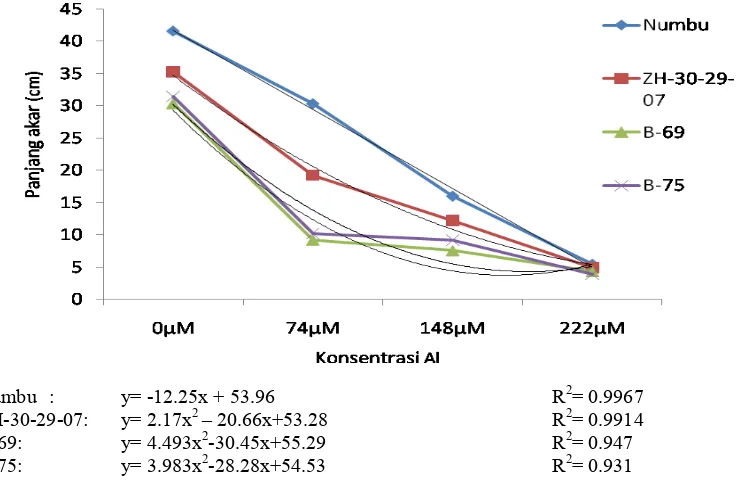
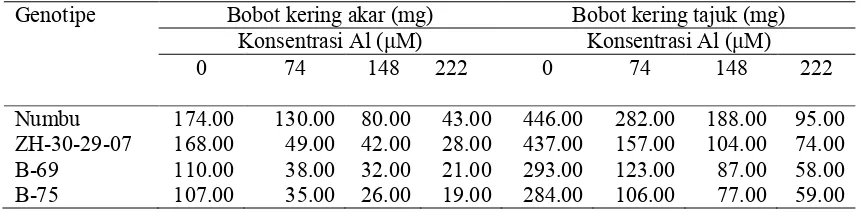
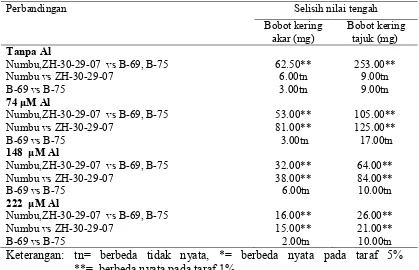
2.2 Respon genotipe sorgum pada berbagai konsentrasi cekaman Al terhadap panjang akar dan panjang tajuk di larutan hara………. 52
2.3 Respon antar genotipe sorgum pada berbagai konsentrasi cekaman Al di larutan hara ………
53
2.4
2.5
Respon genotipe ZH-30-29-07 dan B-75 terhadap panjang akar dan panjang tajuk pada berbagai konsentrasi cekaman Al di larutan hara ……….
Respon genotipe sorgum terhadap panjang akar pada berbagai konsentrasi cekaman Al ...
53
54
2.6 Respon genotipe sorgum terhadap panjang tajuk pada berbagai konsentrasi cekaman Al ... 55
2.8 Respon antar genotipe sorgum pada berbagai konsentrasi
Nilai rataan pengaruh genotipe dan konsentrasi cekaman Al terhadap panjang akar relatif dan Bobot tajuk relatifsorgum
3.1 Rekapitulasi nilai analisis ragam pengaruh genotipe, konsentrasi Al dan interaksi antara pengaruh genotipe dan konsentrasi Al terhadap pertumbuhan sorgum di dalam
rhizotron ………... 68
3.2
3.3
Rata-rata nilai pengaruh interaksi antara kondisi cekaman dan genotipe terhadap bobot kering total dalam rhizotron ...
Respon genotipe sorgum pada berbagai kondisi cekaman P di tanah masam terhadap bobot kering total ..……….
69
70
3.4 Respon genotipe terhadap bobot kering total pada kondisi cekaman Al tinggi dan Al rendah di tanah masam... 70
3.5 Rata-rata nilai terhadap peubah diameter sebaran akar, jumlah akar primer, kadar P jaringan dan efisiensi penggunaan P (EPP) 71
3.6 Respon genotipe sorgum pada dua kondisi cekaman Al di tanah masam terhadap diameter sebaran akar. jumlah akar primer, kadar P jaringan, dan efisiensi penggunaan P... 73
3.7 Respon karakter diameter sebaran akar, jumlah akar primer, kadar P jaringan dan efisiensi penggunaan P genotipe sorgum terhadap pemberian pupuk P pada kondisi cekaman Al tinggi.... 73
3.8 Respon karakter diameter sebaran akar, jumlah akar primer, kadar P jaringan dan efisiensi penggunaan P genotipe sorgum terhadap pemberian pupuk P pada kondisi cekaman Al rendah... 74
4.1
4.2
Rekapitulasi sidik ragam pengaruh genotipe, dan komposisi larutan hara serta interaksi antara genotipe dan komposisi larutan hara terhadap pertumbuhan sorgum fase bibit ………….
Respon genotipe sorgum pada berbagai cekaman Al dan defisiensi P terhadap bobot kering total di larutan hara…………
83
84
4.3 Respon genotipe sorgum pada kondisi cekaman Al berbeda terhadap bobot kering total di larutan hara ……….. 85
4.4 Respon pembentukan bobot kering total genotipe sorgum terhadap pemberian pupuk P pada dua kondisi cekaman Al... 86
4.5
4.6
Nilai tengah pengaruh konsentrasi Al dan genotipe, terhadap nisbah tajuk akar sorgum fase bibit umur 14 HST di larutan
hara ………...
Rataan Laju Serapan Spesifik sorgum yang ditumbuhkan pada larutan hara dengan komposisi yang berbeda selama 14 hari…...
86
88
4.7 Respon genotipe sorgum pada kondisi cekaman Al dan hara P berbeda terhadap laju serapan spesifik P di larutan hara……….. 89
4.8 Rataan Laju Serapan Spesifik pada sorgum toleran dan peka yang ditumbuhkan pada larutan hara selama 14 hari…………... 90
4.9
4.10
Nilai rataan pengaruh komposisi larutan hara terhadap kadar P jaringan sorgum fase bibit umur 14 HST di larutan hara ……...
Nilai rataan pengaruh genotipe terhadap kadar P jaringan sorgum fase bibit umur 14 HST di larutan hara………..
91
Nilai rataan efisiensi penggunaan P (mg.mg BK/mg P) genotipe
sorgum selama 14 hari ……….
Nilai rataan rasio efisiensi P (mg BK/mg P) dalam larutan hara dengan komposisi yang berbeda selama 14 hari …………..
5.1 Rekapitulasi nilai sidik ragam pengaruh konsentrasi Al dan genotipe, terhadap panjang akar dan nilai skoring warna sorgum fase bibit pada lama cekaman Al 6 jam, 24 jam dan 48 jam…… 102
5.2 Nilai rataan pengaruh konsentrasi Al dan genotipe, terhadap
Nilai rataan pengaruh konsentrasi Al dan genotipe, terhadap panjang akar sorgum selama 48 jam tercekam Al ...
Respon genotipe terhadap panjang akar pada konsentrasi dan lama cekaman Al berbeda...
103
104
5.6 Pengaruh konsentrasi Al dan genotipe, terhadap nilai kuantitatif
pewarnaan hematoksilin pada sorgum fase bibit dengan lama cekaman 6 jam ... 105
5.7 Pengaruh konsentrasi Al dan genotipe, terhadap nilai kuantitatif
pewarnaan hematoksilin pada sorgum fase bibit dengan lama cekaman 24 jam ... 105
5.8 Pengaruh konsentrasi Al dan genotipe, terhadap nilai kuantitatif
pewarnaan hematoksilin pada sorgum fase bibit dengan lama cekaman 48 jam ... 105
5.9 pengaruh konsentrasi cekaman Al terhadap skoring pewarnaan hematoksilin pada sorgum dengan berbagai lama waktu cekaman ... 106
6.1 Rekapitulasi nilai sidik ragam pengaruh genotipe, dan komposisi larutan hara terhadap pertumbuhan sorgum pada fase
bibit ……….. 118
6.2 Nilai rataan pertambahan panjang akar, pertambahan volume akar dan pertambahan panjang tajuk sorgum fase bibit pada berbagai genotipe ... 118
6.3 Nilai rataan pertambahan panjang akar, pertambahan volume akar dan pertambahan panjang tajuk sorgum fase bibit pada berbagai konsentrasi Al... 119
Gambar Halaman
1 Bagan alir penelitian mekanisme toleransi sorgum terhadap toksisitas aluminium dan defisiensi fosfor untuk adaptasi di tanah
masam……….
5
2 Mekanisme detoksifikasi Al dalam sel tanaman toleran Al ... 12
3 Pola-pola akumulasi Al di dalam akar jagung toleran Al... 13
4 Urutan gen-gen yang berubah ekspresinya saat terinduksi kahat P secara hipotetik...
Kurva logaritmik erapan P contoh tanah dengan Al tinggi...
Kurva logaritmik erapan P contoh tanah dengan Al rendah... 30
31
2.1
2.2
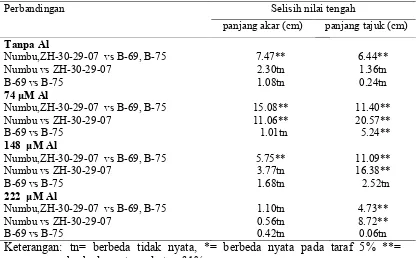
Respon genotipe sorgum terhadap cekaman Al di larutan hara (A) kontrol ( tanpa Al) dan (B) 222 μM Al , dari kiri ke kanan: Numbu, ZH-30-29-07, B-69 dan B-75 ……….
Respon genotipe tanaman terhadap berbagai tingkat konsentrasi Al (A) Numbu/ toleran dan (B) B-75/peka, dari kiri ke kanan: tanpa Al, 74μM Al, 148 μMAl, dan 222μM Al………...
2.6 Bobot kering tajuk sorgum fase bibit pada berbagai konsentrasi Al
3.1 Kondisi tanaman yang ditumbuhkan dalam media Al tinggi-tanpa P (R1), tanaman toleran (T) dan t anaman peka (P) ...
Perbandingan morfologi akar sorgum yang ditumbuhkan dalam media Al tinggi - tanpa P (R1), Al tinggi - P kurang (R2), dan Al tinggi - P cukup (R3) pada tanaman toleran (T) dan peka (P) ...
75
76
3.4 Perbandingan morfologi akar sorgum yang ditumbuhkan dalam media berAl rendah-tanpa P (R4), berAl rendah-P kurang (R5), dan berAl rendah-P cukup (R6) pada tanaman toleran (T) dan peka (P) 76
4.1
5.1 Perbandingan potongan melintang akar sorgum genotipe B-75 pada perlakuan 6 jam tercekam Al. A) 74μM Al, B) 148μM Al,
dan C) 222μM Al ………... 107
5.2
5.3
Genotipe B-69 pada perlakuan 6 jam tercekam Al. A) 74μM Al, B) 148μM Al, dan C) 222μM Al ………
Genotipe ZH-30-29-07 pada perlakuan 6 jam tercekam Al. A) 74μM Al, B) 148μM Al, dan C) 222μM Al ……….
Potongan melintang akar sorgum genotipe B-75 pada perlakuan 24 jam tercekam Al. A) 74μM Al, B) 148μM Al, dan C) 222μM Al...
Genotipe B-69 pada perlakuan 24 jam tercekam Al. A) 74μM Al, B) 148μM Al, dan C) 222μM Al ...
109
110
5.8
5.9
ZH-30-29-07 pada perlakuan 24 jam tercekam Al. A) 74μM Al, B) 148μM Al, dan C) 222μM Al………..
Perbandingan potongan melintang akar sorgum genotipe B-75 pada perlakuan 48 jam tercekam Al. A) 74μM Al, B) 148μM Al, dan C) 222μM Al ...
110
111
5.10 Genotipe B-69 pada perlakuan 48 jam tercekam Al. A) 74μM Al,
B) 148μM Al, dan C) 222μM Al... 111
5.11 Varietas Numbu pada perlakuan 48 jam tercekam Al. A) 74μM Al, B) 148μM Al, dan C) 222μM Al………... 112
5.12 ZH-30-29-07 pada perlakuan 48 jam tercekam Al. A) 74μM Al, B)
148μM Al, dan C) 222μM Al ……… 112
6.1
6.2
Akar sorgum Numbu setelah ditumbuhkan kembali pada larutan tanpa Al. Perlakuan 74μM Al (A), 148μM Al (B), dan 222μM Al
(C) ………..
Akar sorgum ZH-30-29-07 setelah ditumbuhkan kembali pada
larutan tanpa Al. Perlakuan 74μM Al (A), 148μM Al (B), dan 222μM Al (C) ………
120
120
6.3 Perbandingan akar sorgum B-69 setelah ditumbuhkan kembali pada larutan tanpa Al. Setelah perlakuan 74μM Al (A), 148μM Al (B), dan 222μM Al (C) ……….. 120
6.4 Perbandingan akar sorgum B-75 setelah ditumbuhkan kembali pada larutan tanpa Al. Setelah perlakuan 74μM Al (A), 148μM Al (B), dan 222μM Al (C) ……….. 121
7 Pengelompokan empat genotipe sorgum berdasarkan efisiensi penggunaan P pada kondisi bercekaman Al dan P rendah di larutan
1
2
Denah penelitian toleransi terhadap Al dan defisiensi di tanah masam Jasinga ...
Kriteria Penilaian Sifat Kimia Tanah (Pusat Penelitian Tanah, 1983) ...
141
Latar Belakang
Bahan pangan karbohidrat yang berasal dari komoditi tanaman pangan di
negara kita masih berasal dari tanaman padi. Secara teoritis pemenuhan
ketersediaan pangan dapat ditempuh melalui produksi domestik dan impor, namun
bagi bangsa Indonesia dengan jumlah penduduk yang besar dan tersebar di banyak
kepulauan, pemenuhan kebutuhan yang mengandalkan impor akan menciptakan
kerentanan ketahanan pangan nasional. Ketahanan pangan dapat dioptimalkan
dengan mempercepat peningkatan diversifikasi konsumsi pangan (Direktorat
Jenderal Tanaman Pangan 2007). Upaya diversifikasi pangan memberi dampak
terhadap pengurangan konsumsi beras dan berkembangnya komoditi selain padi.
Salah satu tanaman pangan non beras yang telah diidentifikasi sangat
cocok untuk agroklimat Indonesia dan mempunyai prospek sangat bagus untuk
usaha agribisnis lainnya adalah sorgum (Sorghum bicolor L. Moench) Sorgum merupakan tanaman serealia yang potensial untuk dibudidayakan dan
dikembangkan khususnya pada daerah-daerah marjinal dan lahan kering di
Indonesia. Keunggulan sorgum terletak pada daya adaptasi agroekologi yang luas,
tahan terhadap kekeringan, produksi yang tinggi, dan kebutuhan input rendah.
Direktorat Gizi Departemen Kesehatan RI (1992) menyatakan, sorgum
memiliki kandungan nutrisi yang tinggi sehingga sangat baik digunakan sebagai
sumber bahan pangan maupun pakan ternak alternatif. Beberapa kandungan nilai
gizi yang dimiliki sorgum melebihi kandungan yang dimiliki penghasil
karbohidrat utama (beras) seperti protein, lemak, kalsium, besi, fosfor, dan
vitamin B-1.
Menurut Rajvanshi dan Nimbkar (2001), Yudiarto (2006), Reddy dan Dar
(2007), selain untuk pangan sorgum juga dapat dijadikan bahan baku bioenergi.
Batang dan biji sorgum dapat dikonversi menjadi bioetanol melalui proses
fermentasi. Di Amerika Serikat, India, dan Cina, sorgum digunakan sebagai bahan
baku pembuatan bahan bakar bioetanol. Sorgum dapat pula dijadikan bahan baku
Bioetanol merupakan salah satu bahan bakar nabati dengan sumber bahan
baku tanaman yang mengandung karbohidrat seperti tebu, jagung, ubi kayu dan
sorgum. Produksi bahan bakar nabati semakin mendesak untuk terus ditingkatkan.
Perpres Nomor 5 Tahun 2006 dan Inpres Nomor 1 Tahun 2006 menjadi dasar
pengembangan sumber bahan baku untuk bahan bakar nabati dengan target
minimal 5% terhadap total penggunaan energy mix atau setara dengan produksi 400.000 barel minyak per hari pada tahun 2025 (Direktorat Jenderal Tanaman
Pangan 2007). Jika sepertiganya dipenuhi dengan bioetanol, maka harus
diproduksi sekitar 9 juta kilo liter bioetanol kualitas bahan bakar pada tahun 2025.
Peningkatan produksi bioetanol memerlukan pengembangan ketersediaan sorgum
sebagai salah satu bahan bakunya.
Agar sorgum dapat sejajar dengan bahan pangan dan industri lainnya,
perlu diupayakan peningkatan produksi melalui intensifikasi dan ekstensifikasi.
Intensifikasi dapat dilaksanakan dengan memanfaatkan varietas unggul dan
ekstensifikasi melalui perluasan areal penanaman dengan pemanfaatan
lahan-lahan marjinal. Varietas unggul bisa didapatkan dengan program pemuliaan
tanaman melalui perubahan susunan genetik yang diarahkan pada kondisi
lingkungan. Varietas unggul yang dikembangkan harus disesuaikan dengan target
area produksi.
Di Indonesia, kondisi lahan untuk pengembangan tanaman sebagian besar
merupakan lahan marjinal yang kering dan bersifat masam. Kendala utama
pengembangan tanaman pada tanah masam adalah cekaman aluminium dan
kurangnya unsur hara makro terutama fosfor (Taylor 1991). Menurut Sopandie
(2006), perbaikan tanaman pada lahan marjinal sangat memerlukan pemahaman
tentang mekanisme yang berperan dalam peningkatan potensi hasil (yield potential) dan adaptasi tanaman terhadap berbagai cekaman lingkungan abiotik. Pendekatan fisiologi untuk perbaikan potensi hasil dapat dilakukan melalui
perbaikan indeks panen (harvest indeks), efisiensi penggunaan hara, efisiensi fotosintesis, serta keseimbangan source-sink.
Secara umum, tujuan penelitian ini adalah mendapatkan informasi tentang
adaptasi sorgum terhadap cekaman aluminium dan defisiensi fosfor untuk
Penelitian ini menjadi penting karena masih terbatasnya informasi mengenai
mekanisme toleransi sorgum terhadap cekaman aluminium dan defisiensi fosfor
pada lahan masam.
Tujuan Penelitian
Penelitian ini merupakan bagian dari penelitian yang bertujuan untuk
mengembangkan sorgum sebagai bahan pangan dan penghasil bioetanol di
lingkungan area produksi lahan masam, yaitu pembentukan varietas sorgum yang
toleran terhadap cekaman Aluminium dan efisien dalam penggunaan hara mineral
terutama fosfor. Secara khusus penelitian ini bertujuan untuk :
1. Memperoleh informasi tentang tanggap agronomis genotipe sorgum
terhadap produksi biomassa dan kandungan padatan terlarut total pada
kondisi cekaman Al dan defisiensi fosfor di lahan masam
2. Memperoleh informasi tentang dasar fisiologi dari sifat efisiensi hara
fosfor dalam keadaan tercekam Al pada sorgum
3. Memperoleh informasi tentang mekanisme yang mungkin terjadi yang
dapat mendasari perbedaan efisiensi P dalam keadaan tercekam Al baik
melalui penilaian serapan P maupun penilaian penggunaan P dari sorgum
yang toleran dan peka Al
4. Mengidentifikasi karakter-karakter tanaman yang berkaitan dengan
toleransi terhadap toksisitas Al dan defisiensi P, yang dapat digunakan
sebagai karakter untuk seleksi dalam rangka pengembangan tanaman
sorgum sebagai penghasil bioetanol di tanah masam.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat memberi manfaat 1) sebagai
masukan dari bidang kajian fisiologi untuk dijadikan karakter seleksi dalam
program pemuliaan tanaman untuk perbaikan genotipe sorgum sebagai bahan
baku penghasil bioetanol untuk toleransi terhadap toksisitas Al dan efisiensi P di
tanah masam, dan 2) memperkaya informasi tentang mekanisme toleransi
Hipotesis
Hipotesis yang dikemukakan pada penelitian ini adalah :
1. Terdapat perbedaan tanggap agronomi genotipe sorgum terhadap toksisitas
Al dan defisiensi P di tanah masam
2. Terdapat perbedaan toleransi terhadap cekaman Al pada genotipe sorgum
di larutan hara.
3. Genotipe adaptif tanah masam memiliki akar lebih panjang, diameter
sebaran akar lebih luas serta mampu mempertahankan bobot kering tajuk
yang lebih tinggi dibandingkan genotipe peka
4. Genotipe toleran memiliki laju serapan spesifik P dan efisiensi
penggunaan P lebih tinggi dibandingkan genotipe peka..
5. Genotipe toleran lebih mampu menahan distribusi dan akumulasi Al ke
dalam jaringan akar dibandingkan genotipe peka.
6. Genotipe toleran mampu menumbuhkan akarnya kembali setelah terkena
cekaman Al.
Kerangka dan Ruang Lingkup Penelitian
Untuk mencapai tujuan penelitian dan menjawab hipotesis, dilakukan
serangkaian percobaan yang merupakan satu kesatuan, dan tahapan yang
ditempuh merupakan proses yang sistematis dalam mencapai tujuan. Penelitian
ini lebih menekankan pada studi fisiologi mekanisme toleransi sorgum terhadap
toksisitas Al dan defisiensi P. Percobaan 1 merupakan percobaan lapangan yang
dilaksanakan di tanah masam Kebun Percobaan UPTD Lahan Kering Dinas
Pertanian Kabupaten Bogor Kecamatan Tenjo, Jasinga. Berdasarkan peta tanah
Lembaga Pusat Penelitian Tanah Bogor, tanah di daerah ini tergolong Podzolik
Merah Kekuningan dengan bahan induk batuan liat. Percobaan 2, 3, 4, 5 dan 6
dilaksanakan di rumah kaca Kebun Percobaan University Farm IPB di Cikabayan.
Analisis kimia jaringan tanaman dan media tumbuh dilaksanakan di Laboratorium
RGCI Fakultas Pertanian Institut Pertanian Bogor, Laboratorium Micro Technique
IPB dan Laboratorium SEAMEO BIOTROP. Ruang lingkup penelitian secara
Gambar 1. Bagan alir penelitian fisiologi adaptasi sorgum terhadap toksisitas aluminium dan defisiensi fosfor di tanah masam
Pertumbuhan Akar pada
Genotipe sorgum toleran dan peka hasil uji daya adaptasi di Taman Bogo Lampung Timur dan
Sulusuban Lampung Tengah
Studi Lapang Studi di Larutan Hara, Rhizotron dan
Morfologi dan Fisiologi Sorgum
Sorgum memiliki sistem perakaran serabut. Keunggulan sistem perakaran
pada tanaman sorgum yaitu sanggup menopang pertumbuhan dan perkembangan
tanaman ratun (ratoon) hingga dua atau tiga kali ratoon dengan akar yang sama.
Saat proses perkecambahan akar primer mulai tumbuh dan seiring dengan proses
pertumbuhan tanaman akan diikuti pula dengan pertumbuhan akar sekunder pada
ruas pertama. Tahap berikutnya akar sekunder lebih dominan berfungsi menyerap
air dan hara dari media tumbuh serta memperkokoh tegaknya tanaman (House
1985).
Sorgum memiliki batang dengan tinggi bervariasi antara 0.5 – 4.0 m. Tinggi batang sorgum yang dikembangkan di China dapat mencapai 5.0 m (FAO,
2002),. Beberapa varietas sorgum memiliki batang yang menghasilkan
percabangan dan anakan baru (Steenis 1975). Batang tanaman sorgum
merupakan rangkaian berseri dari ruas (internodes) dan buku (nodes). Bentuk batang silinder dengan ukuran diameter batang bagian pangkal antara 0.5 – 5.0 cm (House 1985).
Daun sorgum mirip tanaman jagung, berbentuk pita dengan struktur daun
terdiri atas helai dan tangkai daun. Panjang rata-rata daun sorgum adalah 1 m
(House 1985). Posisi daun terdistribusi secara berlawanan sepanjang batang
dengan pangkal daun menempel pada buku. Menurut Martin (1970) jumlah total daun berkisar antara 13-40 helai per batang. Jumlah daun sorgum berkorelasi
tinggi dengan panjang periode vegetatif yang dibuktikan oleh setiap penambahan
satu helai daun memerlukan waktu 3-4 hari ( Bullard dan York 1985). Tanaman
sorgum juga memiliki daun bendera (leaf blades) yang muncul paling akhir bersamaan dengan inisiasi malai. Daun bendera muda bentuknya kaku, tegak dan
berfungsi penting dalam transportasi fotosintat (Freeman 1970).
Sorgum tergolong tanaman C4, yaitu tanaman yang dalam proses
metabolisme karbon (C) menghasilkan asam berkarbon empat (malat dan aspartat)
sebagai produk awal penambatan CO2. Produk asam malat dan aspartat yang
mengalami dekarboksilasi melepaskan CO2 yang selanjutnya ditambat Rubisco
dan diubah menjadi 3-PGA (asam fosfo gliserat). Sel seludang pembuluh
tanaman C4 lebih tebal dibandingkan tanaman C3, sehingga lebih banyak
mengandung kloroplas, mitokondria dan organel lain yang berperan sangat
penting dalam proses fotosintesis (Taiz dan Zeiger 2002). Daun-daun spesies C4
mempunyai laju pertukaran CO2 yang lebih tinggi, rasio antara luas potongan
melintang floem dengan luas daun yang lebih besar dan memiliki laju translokasi
lebih besar dibandingkan tanaman C3 (Salisburry dan Ross 1995). Ekspor hasil
asimilasi yang lebih baik oleh tanaman C4 disebabkan oleh anatomi khususnya,
yaitu sel-sel seludang ikatan pembuluhnya yang mempunyai kloroplas (anatomi
kranzs) atau hasil dari luas potongan melintang floem yang lebih besar.
Karakteristik Tanah Masam
Tanah di lingkungan tropika basah pada umumnya bersifat masam dan
merupakan ciri khas sebagian besar wilayah di Indonesia. Tanah jenis ini tersebar
di bebarapa daerah di luar Jawa, seperti Sumatera dan Kalimantan. Di Sumatera
terdapat sekitar 21 juta hektar, Kalimantan 15.5 juta hektar, dan Jawa 2 juta hektar
(Van der Heide et al., 1992). Menurut Hidayat dan Mulyani (2002), luas tanah masam yang berupa lahan kering mencapai 99.5 juta hektar dan berpotensi untuk
dikembangkan menjadi lahan pertanian. Kondisi tropika basah di Indonesia
dengan curah hujan tinggi dapat mengakibatkan pencucian, sehingga cadangan
unsur hara dan kesuburan tanah rendah, kandungan Al dapat ditukar (Al-dd) dan
kapasitas retensi P tinggi, kandungan nitrogen yang rendah, kapasitas tukar kation
(KTK) rendah, serta keracunan alumunium di lapisan bawah (Hairiah et al. 2000). Di Indonesia potensi tanah masam ini cukup tinggi. Menurut data Pusat
Penelitian Tanah dan Agroklimat tahun 2000, luas areal tanah bereaksi masam
seperti podsolik, ultisol, oxisol dan spodosol masing-masing sekitar 47.5; 18.4;
5.0; dan 56.4 juta ha atau seluruhnya sekitar 67% dari luas total tanah di
Indonesia. Pengelolaan kesuburan di tanah masam diarahkan untuk menurunkan
kemasaman tanah, menambah hara dan menekan tingkat kejenuhan Al. Teknologi
untuk meningkatkan kesuburan tanah masam yang dapat diterapkan antara lain
pemupukan berimbang, pengelolaan hara P, pengapuran serta pemberian bahan
Pengaruh Cekaman Aluminium terhadap Tanaman
Aluminium dapat mempengaruhi tanaman secara morfologis, fisiologis
dan ekspresi gen tanaman. Gejala yang umum dijumpai akibat cekaman Al adalah
terjadinya klorosis, defisiensi nutrisi, dan tanaman menjadi kerdil (Taiz dan
Zeiger 2002). Respon morfologi nyata akibat cekaman Al adalah terjadinya
penebalan pada ujung akar dan akar cabang. Respon fisiologi berupa
pembentukan kompleks Al-asam organik dan peningkatan kandungan asam
organik pada akar tanaman dengan cara: 1) Al mengaktivasi kerja enzim yang
berperan dalam biosintesis asam organik, serta adanya asam organik yang
ditransportasikan dari batang menuju akar (Matsumoto et al. 2003). Respon pada tingkat gen untuk tanaman sorgum belum diteliti, tetapi pada beberapa tanaman
lain antara lain pada arabidopsis menunjukkan ekspresi spesifik gen WAK1 (cell wall-associated reseptor kinase 1) dan tipe sel yang merupakan lokasi spesifik dari protein WAK (gen WAK ini terekspresi di akar) (Kochian et al. 2005).
Pada tanaman gandum, resistensi terhadap Al bersifat multigenik.
Gen-gen tersebut menGen-gendalikan pengeluaran beberapa senyawa pengkelat ion Al+3.
Hasil penelitian Pelle et al. (1996) menunjukkan, gandum kultivar resisten akan mengeksudasikan P tinggi yang diduga dikendalikan oleh gen yang berbeda lokus.
Kelarutan Al yang tinggi berpengaruh langsung terhadap metabolisme
tanaman dan tidak langsung terhadap ketersediaan unsur hara sehingga
pertumbuhan tanaman tertekan. Menurut Alam et al. (1999), secara umum pengaruh Al pada tanaman yang ditumbuhkan pada tanah masam adalah: 1)
mengurangi kation bervalensi dua yang diserap oleh akar tanaman (khususnya
Ca). Menurut Matsumoto (2003), hal ini terjadi karena penghambatan Al dengan
cara menggantikan posisi Ca yang melekat pada Calmodulin (dinding sel), ikatan
Al dengan karboksil (RCOO-) membentuk ikatan kuat sehingga sel tidak mampu
membesar. 2) menghambat fungsi sel-sel pada jaringan meristem akar melalui
penetrasi Al ke dalam protoplasma akar dan menghasilkan morfologi akar yang
tidak normal dan dapat mengganggu proses penyerapan hara tanaman, dan 3)
menurunkan adsorpsi anion (SO4-2, PO4-3, dan Cl-) karena meningkatnya daerah
Kesulitan dalam mempelajari Al berhubungan dengan proses-proses yang
terdapat dalam tanaman disebabkan karena kompleksnya Al ( Kinroide, 1991).
Al dihidrolisis dalam larutan sebagai ion trivalent Al3+ dan dominan pada kondisi
pH <5, sedangkan Al(OH)2+ merupakan bentuk yang dominan dengan makin
tingginya pH. Pada keadaan tanah yang bereaksi netral, Al berbentuk Al(OH)3
atau gibsit, sedangkan pada tanah alkalin dijumpai bentuk Al(OH)4-. Kation Al
monomer membentuk ikatan dengan berbagai ligand asam organik dan anorganik
seperti PO43-, SO42-, asam organik, protein dan lemak.
Tanah dengan pH rendah memiliki kapasitas ion H+ tinggi sehingga
penyerapan unsur-unsur lainnya menjadi berkurang dan unsur Al meningkat.
Meningkatnya konsentrasi Al terlarut mengakibatkan terjadinya defisiensi P, K
dan hara mikro seperti Mo. Hasil penelitian Yamamoto et al. (1992) mendapatkan bahwa toksisitas Al selain mengakibatkan tanaman kekurangan
hara juga mengubah struktur dan fungsi dari membran plasma dan menghalangi
pembelahan sel pada ujung-ujung akar. Akhirnya, Al akan menghambat
pertumbuhan akar dan menunjukkan berbagai gejala kekurangan hara akibat
keracunan Al (MacDiarmid dan Gardner, 1996).
Kasus pada tanaman jagung menunjukkan bahwa cekaman Al terhadap
tanaman mula-mula akan menekan pertumbuhan akar yaitu akar menjadi pendek,
tebal dan rapuh. Terhambatnya pertumbuhan akar disebabkan karena Al
berasosiasi dengan DNA pada inti sel dan menghentikan proses pembelahan sel
meristem apikal (Pellet et al. 1995). Daerah yang paling peka terhadap keracunan Al terutama pada bagian ujung akar (tudung akar, meristem, dan zona
pemanjangan) sekitar 2 mm. Ujung akar mengakumulasi Al lebih banyak
(Delhaize dan Ryan 1995).
Sasaran utama cekaman Al pada akar adalah tudung akar. Rusaknya
tudung akar akan mengakibatkan berkurangnya sekresi mucilage. Keracunan Al
dapat menghambat pertumbuhan tajuk dengan cara menghambat pasokan hara, air
dan sitokinin dari akar karena buruknya penetrasi akar ke sub-soil atau kondisi
hidrolik akar rendah (Marschner 1995). Rusaknya akar oleh Al menyebabkan
terganggunya penyerapan dan transpor hara Ca, K, P, Mg dan N, serta peka
produktivitas tanaman (Polle dan Konzak 1990). Akumulasi Al akan
menyebabkan kebocoran membran, disintegrasi struktur dan berkurangnya
kandungan K dalam jaringan ujung akar, serta menurunkan viabilitas protoplasma.
Terbentuknya ikatan polimer Al dengan membran plasma akar akibat cekaman Al
akan menyebabkan kerusakan pada membran dan kebocoran K dari sel akar
(Matsumoto et al. 2003).
Toksisitas Al mempengaruhi efluks unsur K dan Ca. Gangguan Al
terhadap Ca pada ujung akar menyebabkan defisiensi Ca pada sel apikal akar atau
mengubah homeostatis Ca. Perubahan ini akan memicu penyimpangan fungsi
metabolisme dalam sel ujung akar yang selanjutnya dapat menghambat
pemanjangan akar (Huang et al. 1992).
Mekanisme Toleransi Tanaman terhadap Cekaman Aluminium
Suatu tanaman yang toleran terhadap keracunan Al mempunyai kriteria
antara lain: 1) pertumbuhan akar normal 2) mampu meningkatkan pH tanah di
sekitar perakaran, 3) sebagian besar Al tertahan di akar dan sedikit
ditranslokasikan ke bagian atas tanaman, dan 4) ion Al tidak dapat menghambat
serapan dan translokasi Ca, Mg, K dan P (Kochian 1995).
Pengaruh cekaman Al+3 tidak sama pada setiap spesies, bahkan pada
tanaman dalam satu spesies. Adanya perbedaan tersebut menunjukkan adanya
mekanisme toleransi yang berbeda pada setiap tanaman dalam mengatasi
cekaman. Mekanisme toleransi terhadap Al menurut Marschner (1995) adalah: 1)
ekslusi Al di membran akar, 2) KTK dinding sel rendah, 3) alkalisasi di daerah
perakaran, 4) proteksi ujung akar oleh mucilage, dan 5) efluks Al.
Mekanisme toleransi tanaman secara umum terbagi dalam dua kelompok
yaitu: 1) mekanisme penolakan secara eksternal (external tolerance exclusion mechanism), dengan cara mencegah Al masuk ke dalam simplas dan bagian metabolik yang sensitif melalui immobilisasi dinding sel, permeabilitas selektif
membran plasma, barier pH di rizosfir dan apoplas akar, eksudasi ligan pengkelat
(eksudasi asam organik), eksudasi fosfor dan efluks Al, dan 2) mekanisme secara
pengikat Al, enzim yang tahan Al serta peningkatan aktivitas enzim (Taylor,
1991).
1. Kelatasi Al oleh Eksudasi Asam Organik Tanaman
Salah satu penyebab terjadinya detoktisifikasi Al oleh tanaman yang
toleran adalah karena adanya asam organik yang dieksudasikan sehingga Al
terkelatasi oleh asam organik baik di dalam jaringan maupun di media/larutan
(Delhaize dan Ryan 1995). Asam organik yang dieksudasikan oleh akar tanaman
umumnya adalah asam malat, asam sitrat dan asam oksalat (Tabel 1). Jumlah dan
jenis yang dieksudasikan tergantung spesies dan kultivar tanaman.
Hasil penelitian Magalhaes et al. (2004) menunjukkan bahwa asam sitrat merupakan asam organik yang dieksudasikan tanaman sorgum dalam menghadapi
cekaman Al, asam malat merupakan asam organik yang paling banyak
dieksresikan oleh ujung akar pada tanaman gandum yang toleran Al (Delhaize et al. 1993; Ryan et al. 1995 dan Pellet et al. 1996).
Tabel 1 Jenis-jenis asam organik yang dieksudasi tanaman dalam hubungannya dengan toleransi terhadap Al Sitrat Nicotiana tabacum Delhaize et al. (2001)
Sitrat Oryza sativa Ma et al. (2002)
Sitrat Sorghum bicolor Magalhaes (2002)
Sitrat Zea mays Ishikawa et al (2000)
Sitrat dan Malat Avena sativa Zheng et al. (1998) Sitrat dan Malat Raphanus sativus Zheng et al. (1998) Sitrat dan Malat Triticale sp. Ma et al. (2000) Sitrat dan oxalat Zea mays Kid et al. (2001) Malat Triticum aestivum Huang et al. (1996), Ishikawa et al. (2000)
Oksalat Colocasia esculenta Ma (1998)
Oksalat Fogopyrum esculentum Zheng et al. (1998) __________________________________________________________________ Sumber: Kochian et al. (2005).
Tanaman jagung yang toleran Al mengeksudasikan asam sitrat lebih
(Yelow Biloxi) mengeksudasikan asam oksalat dan asam sitrat lebih tinggi
dibanding kedelai peka (Lumut) (Sopandie, 2006).
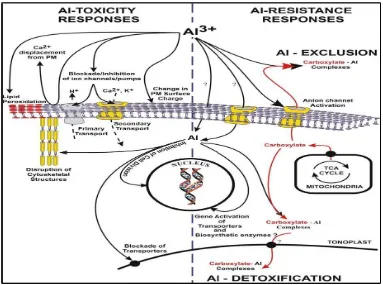
Gambar 2 menunjukkan mekanisme pengikatan Al oleh asam organik
dengan membentuk kompleks sehingga Al tidak toksik dalam sel tanaman yang
toleran terhadap Al, dan aktivasi ion channel pengikat Al serta perbandingannya
dengan mekanisme pada tanaman respon toksisitas Al (peka). Tanaman toleran
Al akan membentuk ikatan kompleks Al-COOH (Carboxylate-Al) sehingga Al
menjadi tidak aktif. Ikatan kompleks ini akan ditranslokasikan ke vakuola
menembus membran tonoplas dan akan terjadi detoksifikasi Al. Tanaman peka
tidak membentuk kompleks Al-COOH sehingga Al tetap aktif dan dapat
menghambat pembelahan sel serta penghambatan aktivitas ion channel yang dapat
mempengaruhi transportasi dalam sel tanaman.
Sumber : Kochian et al. (2004).
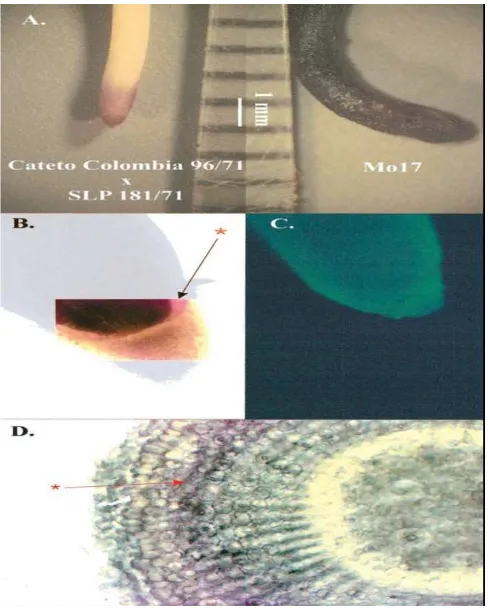
2. Akumulasi Al pada Ujung Akar/ Hambatan Transpor Al ke Tajuk
Tanaman toleran Al akan melakukan akumulasi Al pada ujung-ujung akar
sebagai upaya hambatan terhadap transpor dan akumulasi Al pada bagian tajuk.
Gambar 6 dibawah ini menunjukkan hasil uji pewarnaan hematoxylin terhadap
jagung cultivar cateto columbia dan Mo17 (Pineros et al. 2002).
Sumber: Pineros et al. (2002).
Gambar 3. Pola-pola akumulasi Al di dalam akar jagung toleran Al Cateto-Colombia 96/71(kiri) dan cv.Mo17 (kanan) hasil uji pewanaan Hematoxylin (A) B. Penampang memanjang akar daerah apikal (tanda panah menunjukkan lapisan sel bagian luar tudung akar yang mengakumulasi Al. C. Distribusi akumulasi Al pada ujung akar cv Cateto-Colombia. D. Penampang melintang akar hasil uji pewarnaan (tanda panah menunjukkan penetrasi pewarnaan ke dalam lapisan sel akar bagian dalam tanaman yang mengakumulasi Al
3. Kemampuan Menaikkan pH Rizosfir
Peningkatan pH rizosfir pada larutan/media merupakan salah satu
meningkatkan ketersediaan unsur hara seperti P dan menurunkan ketersediaan Zn,
Cu, Fe, Mn dan Al (Haynes 1990). Genotipe toleran tanaman jagung, gandum,
barley dan padi yang ditumbuhkan pada larutan hara meningkatkan pH larutan
dan menurunkan kelarutan dan toksisitas Al, sedangkan genotipe yang peka tidak
mempunyai pengaruh terhadap pH dan konsentrasi kelarutan Al nya tetap tinggi
(Gupta 1997).
Tanaman Arabidopsis thaliana yang diinkubasi dengan 300 μM AlCl3 selama 12 jam menunjukkan bahwa, genotipe toleran mampu menaikkan pH
larutan dari 4.3 – 4.4 menjadi 4.53, sedangkan genotipe peka hanya mampu menaikkan pH sampai 4,39. Kenaikan pH rizosfir sebesar 0.1 unit pada kondisi
cekaman Al menyebabkan kenaikan laju pertumbuhan akar dua kali lipat pada
genotipe toleran dibandingkan genotipe peka (Degenhardt et al. 1998). Hasil penelitian Delhaize et al. (1995), pada tanaman kedelai menunjukkan bahwa genotipe toleran memiliki kemampuan dua kali lipat dalam meningkatkan pH
rizosfir dibandingkan genotipe peka.
4. Eksudasi Fosfor Organik
Ujung akar tanaman jagung disamping dapat mengeksudasi asam organik,
juga mampu mengeksudasikan fosfor organik. Eksudasi P dari akar tanaman
merupakan proses alami hara P dan bagian dari keseimbangan hara P dalam
tanaman (Pelle et al. 1996). Eksudasi fosfor organik oleh akar tanaman merupakan mekanisme sekunder yang membuat kemampuan tanaman toleran
terhadap Al. Detoksifikasi Al oleh fosfor karena terbentuknya kompleks Al-P
(Delhaize et al. 1993; Pelle et al. 1995 dan 1996). Ketika suplai P terbatas, secara absolut eksudasi P berkurang, tetapi secara relatif meningkat dan menjadi
komponen utama dalam penyerapan hara P. Jika tanaman disuplai P dengan baik,
maka akan meningkatkan eksudasi P dibandingkan dengan tanaman dalam kondisi
cekaman P. Jumlah fosfor organik yang dieksudasi pada tanaman jagung maupun
pada gandum yang toleran nyata lebih banyak dibanding tanaman yang peka
Interaksi Fosfor dengan Aluminium
Tanaman menyerap P dalam bentuk ion H2PO4- atau HPO4-2. Bentuk
dominan yang diserap pada pH rendah adalah H2PO4-, pH sekitar netral (pH 6-7)
kedua bentuk tersebut dapat diserap, sedangkan pada tanah alkalis, bentuk hara P
dominan yang diserap adalah HPO4-2 (Marschner 1995).
Kelarutan Al+3 sangat tinggi di lahan masam menyebabkan P tidak larut
dan kurang tersedia bagi tanaman (hanya sebagian kecil saja seperti bentuk
H2PO4- yang tersedia bagi tanaman). Al tidak hanya menghambat ketersediaan P,
tetapi juga menghambat transpor dan penggunaan P (Rao et al. 1999). Ion Al bermuatan positif dapat berasosiasi dengan gugus fosfor dari ATP atau fosfolipid
pada membran sehingga mempengaruhi efektivitas transportasi proton. Hal ini
akan mengakibatkan penyerapan hara yang dikatalis pompa proton menurun
(Matsumoto et al. 1992). Al secara langsung berinteraksi dan memiliki kapasitas fiksasi yang tinggi terhadap unsur P, baik dalam larutan tanah maupun jaringan
tanaman membentuk kompleks Al-P yang tidak larut sehingga tidak tersedia bagi
tanaman (Marscner 1995). Reaksi sederhana pengendapan P oleh Al adalah
sebagai berikut:
Al+3 + H2PO4- (larut) + 2H2O.2H2 → Al(OH)2H2PO4 (tidak larut)
Keracunan Al dapat diasosiasikan dengan gejala defisiensi P, sebaliknya P efektif
sebagai agen detoksifikasi Al. Interaksi Al-P dalam tanaman adalah: 1)
pengendapan Al-P dan 2) mengganggu metabolisme P (Baligar et al. 1997). Kemiripan gejala toksisitas Al dengan defisiensi P antara lain nekrosis pada ujung
daun, penghambatan pertumbuhan, daun berwarna hijau tua, dan kadang-kadang
daun atau batang berwarna ungu. Defisiensi P disebabkan karena menurunnya
penyerapan dan transpor P oleh pengendapan Al-P pada akar tanaman, dan
gangguan Al dalam metabolisme P yang sudah terdapat pada pucuk tanaman
(Matsumoto et al. 1992).
Hasil penelitian Syafruddin (2002) menunjukkan, genotipe jagung yang
toleran Al jika ditumbuhkan pada larutan hara dengan konsentrasi Al rendah (2.5
ppm) dikombinasikan dengan konsentrasi P rendah (6 ppm) menunjukkan
ditunjukkan oleh genotipe AMATL-HS-C2-S0 (toleran) yang mempunyai Panjang
Akar Relatif (PAR) lebih tinggi pada konsentrasi 2.5 ppm Al disertai 6 ppm P
dibandingkan tanpa Al pada konsentrasi P sama atau pada konsentrasi 2.5 pm Al
dengan 11.5 ppm P. Pemanjangan akar kemungkinan dimaksudkan untuk
mencegah tanaman kekurangan P di akar. Bertambahnya panjang akar,
menyebabkan bertambahnya luas permukaan akar, sehingga kontak akar dengan
larutan untuk mengambil hara P akan lebih tinggi. Sebaliknya, tanaman peka Al
akan memperlihatkan penurunan pertumbuhan akar, bobot kering tanaman serta
kadar dan serapan hara P. Pengaruh buruk ini dapat ditekan dengan meningkatkan
konsentrasi P di larutan/media.
Rasio P akar/tajuk meningkat secara nyata dengan penambahan Al larutan
pada tanaman yang toleran dibandingkan genotipe peka. Peningkatan rasio kadar
P akar/tajuk ini diduga untuk mencegah berkurangnya kadar P di akar oleh adanya
Al, karena P di akar sangat penting dalam eksudasi P ke larutan (Swasti 2004).
Fungsi dan Gejala Defisiensi Fosfor
Fosfor termasuk hara makro dibutuhkan tanaman dalam jumlah banyak
seperti halnya N, K, Ca, Mg dan S. Dalam tanaman P dijumpai dengan kadar
0.1-0.4% (Tisdale et al., 1985). Menurut Marschner (1995), fungsi P pada tanaman dapat digolongkan dalam tiga bagian. Fungsi pertama adalah sebagai penyusun
makro molekul. Dua makro molekul yang utama dan terpenting yang melibatkan
P adalah asam nukleat (DNA dan RNA) dan fosfolipid biomembran. Asam
nukleat adalah senyawa yang berperan dalam pewarisan sifat dan perkembangan
tanaman. Pada biomembran P membentuk ikatan atau jembatan antara digliserida
dan molekul lainnya seperti asam amino, amina atau alkohol, membentuk
fosforidilkolin (lesitin) yang menjaga integritas membran. Fungsi kedua P adalah
sebagai unsur pembentuk senyawa penyimpan dan perpindahan energi. Dua
senyawa kaya energi yang paling umum adalah ATP dan ADP. Energi dalam
ATP/ADP terletak pada ikatan pirofosfor yang pemecahannya akan melepaskan
energi, yang dikenal dengan proses fosforilasi. ATP merupakan sumber energi
untuk hampir semua proses biologi yang membutuhkan energi. Unsur P juga
ribulosa 1.5-bifosfor. Fungsi ketiga P adalah regulator reaksi biokimia melalui
fosforilasi yang dapat mengaktivasi atau inaktivasi protein yang dianggap sebagai
faktor kunci dalam transduksi sinyal.
Secara agronomis unsur P diketahui berperan dalam percepatan
pematangan biji, kekuatan batang sereal, serta mutu buah, hijauan dan biji-bijian.
Benih yang dihasilkan dari tanaman yang cukup P akan memiliki daya kecambah
dan vigour yang tinggi karena kandungan senyawa phytin meningkat (Mengel dan
Kirby 1982).
Tanaman menyerap P dari larutan tanah terutama dalam bentuk
orthofosfor primer dan sekunder (H2PO4- dan HPO42-) dan sedikit dalam bentuk
senyawa organik (Tisdale et al., 1985). Orthofosfor sekunder lebih dominan pada pH di atas 7.22, namun tanaman menyerap P lebih lambat dibanding orthofosfor
primer. Bagian tanaman yang aktif menyerap P adalah jaringan muda dekat ujung
akar. Konsentrasi P yang relatif tinggi menumpuk di ujung akar diikuti oleh
akumulasi rendah pada bagian pemanjangan.
Penyerapan P oleh tanaman dari tanah adalah penyerapan aktif karena
melawan gradien konsentrasi (Clarkson dan Grignon 1991). Kadar P larutan
tanah di luar sel akar umumnya 1µM atau kurang, sedangkan kadar dalam
sitoplasma adalah 103 sampai 104 lebih tinggi. Setelah diserap, fosfor dapat tetap
sebagai P inorganik atau teresterifikasi (melalui gugus hidroksil) dengan rantai
karbon sebagai ester P sederhana (gula P) atau terikat dengan P lainnya dengan
ikatan pirofosfor kaya energi (ATP, ADP) atau diester.
Berbeda dengan unsur hara N dan K, P bergerak menuju akar melalui
difusi yang berjalan lambat. Pergerakan secara difusi merupakan mekanisme
pergerakan P menuju akar yang paling penting. Difusi P sangat dipengaruhi oleh
kadar air tanah, kapasitas penyangga P tanah, temperatur dan bentuk lintasan
difusi (Tisdale et al., 1985). Mobilitas P yang rendah pada tanah menyebabkan karakter morfologi akar seperti panjang akar dan luas permukaan ditemukan
sangat mempengaruhi serapan P pada beberapa spesies tanaman. Dalam
penelitian dengan beberapa kultivar padi ditemukan bahwa toleransi terhadap P
rendah seluruhnya tergantung variasi genetik dalam serapan P, yang sangat
Gejala khas kekahatan sering sukar terlihat, tidak seperti gejala kekahatan
unsur lainnya seperti K dan Mg. Kekerdilan dan pengurangan jumlah anakan
pada tanaman monokotil atau cabang pada dikotil, daun pendek dan tegak, serta
penundaan pembungaan adalah gejala yang umum pada kebanyakan tanaman
(Rao dan Terry 1999). Penurunan luas dan jumlah daun juga merupakan gejala
kekahatan P akibat tertekannya perkembangan sel epidermis daun. Tanaman yang
kekahatan P juga sering memperlihatkan daun sempit berwarna hijau gelap, hal ini
terjadi karena pertambahan luas daun lebih tertekan dibandingkan pembentukan
kloroplas dan klorofil.
Pemberian pupuk fosfor adalah salah satu usaha untuk mengatasi
kekahatan P. Namun, beberapa hasil penelitian menunjukkan bahwa efisiensi
pemupukan fosfor pada tanaman sangat rendah, hanya 15-20% dari P yang
diberikan dapat diserap tanaman (Baharsyah 1990). Rendahnya efisiensi
pemupukan P ini menurut Tisdale et al., (1985) disebabkan terjadinya transformasi (fiksasi dan presipitasi) P yang diberikan dengan ion-ion besi,
alumunium, kalsium, magnesium dan mangan menjadi bentuk yang sukar larut
dalam air.
Mekanisme Toleransi Tanaman terhadap Defisiensi Fosfor
Toleransi terhadap cekaman hara rendah adalah kemampuan tanaman
untuk mempertahankan hasil pada kondisi hara terbatas. Sifat toleran ini tidak
dapat dipisahkan dengan efisiensi seperti terlihat pada mekanisme yang
mendasarinya (Rao et al. 1999). Toleransi lebih menggambarkan dinamika atau respon tanaman terhadap perubahan lingkungan dan menggambarkan kemampuan
adaptasi tanaman (Wissuwa dan Ae 2001).
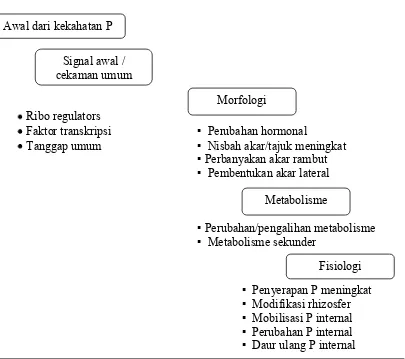
Secara umum adaptasi tanaman terhadap defisiensi P dicapai melalui
mekanisme peningkatan penyerapan dan peningkatan efisiensi penggunaan (Rao
et al., 1999). Peningkatan penyerapan dicapai melalui perbedaan morfologi (pertumbuhan dan distribusi, diameter, rambut akar), dan fisiologi akar (sistem
penyerapan dan mobilisasi P pada rizosfir), sedangkan efisiensi penggunaan
dicapai melalui partisi (mobilisasi) P dalam tanaman dan efisiensi penggunaan
Menurut Peng dan Ismail (2004), mekanisme adaptasi tanaman terhadap P
rendah dapat dikelompokkan ke dalam dua kelompok, yaitu: (1) mekanisme
internal yang berkaitan dengan efisiensi penggunaan P oleh jaringan, dan (2)
mekanisme eksternal yang memungkinkan efisiensi serapan P yang lebih tinggi
oleh akar. Mekanisme Internal dicapai melalui kemampuan tanaman untuk: (a)
memanfaatkan P dengan efisien, dan (b) memobilisasi P dari jaringan yang tidak
aktif lagi bermetabolisme. Menurut Kochian et al. (2004), ketersediaan P tanah yang rendah akibat mudahnya P terfiksasi oleh bahan organik menyebabkan
mekanisme eksternal menjadi lebih penting karena tanaman mengembangkan
berbagai mekanisme untuk membuat P menjadi tersedia dan untuk meningkatkan
kemampuan menyerap P.
Mekanisme eksternal meliputi: (a) kemampuan tanaman untuk membentuk
perakaran yang lebih panjang, (b) kemampuan meningkatkan luas serapan dengan
pertumbuhan rambut-rambut akar, (c) kemampuan melarutkan P tidak tersedia
melalui perubahan pH atau sekresi senyawa pengkelat, (d) kemampuan
menggunakan P organik melalui sekresi fosfatase, dan (e) kemampuan dalam
bersimbiosis dengan mikorhiza. Menurut Kochian et al. (2004), salah satu mekanisme eksternal penting dalam meningkatkan kemampuan menyerap P
adalah dengan peningkatan kinetika serapan P. Perubahan fisiologi akar juga
merupakan mekanisme toleransi tanaman terhadap terbatasnya suplai P yang
terutama disebabkan oleh perubahan kinetika serapan. Kinetika serapan P
menunjukkan adanya high affinity transporter yang lebih aktif pada konsentrasi P rendah dibanding low affinity transporter. Hasil penelitian Syarief (2005) juga menunjukkan afinitas karier akar yang tinggi terhadap P yang ditunjukkan oleh
konstanta Michaelis-Menten (Km) yang rendah yang merupakan salah satu
mekanisme toleransi terhadap P rendah pada tanaman padi. Gordon-Weeks et al. (2003) melaporkan peningkatan ekspresi gen-gen dari kelompok Pht 1 yang menyandikan proton-Pi cotransporter pada akar tanaman kentang yang ditumbuhkan pada keadaan kahat P. Penyerapan P melalui transporter dimediasi
oleh H+-Pi cotransport yang mendapat energi dari H+-ATPase. Mekanisme
adaptasi tanaman terhadap kekahatan P yang meliputi respon morfologi, fisiologi,