IDENTIFIKASI DAN KARAKTERISASI RIZOBAKTERI
PENGHASIL GIBERELIN YANG DIISOLASI DARI TANAH
HUTAN DI BANTEN
HADI SUSILO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Identifikasi dan Karakterisasi
Rizobakteri Penghasil Gibrelin yang Diisolasi dari Tanah Hutan di Banten adalah
benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan
dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang
berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari
penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di
bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut
Pertanian Bogor.
Bogor, November 2015
Hadi Susilo
RINGKASAN
HADI SUSILO. Identifikasi dan Karakterisasi Rizobakteri Penghasil Giberelin
yang Diisolasi dari Tanah Hutan di Banten. Dibimbing oleh NISA RACHMANIA
MUBARIK dan TRIADIATI.
Giberelin adalah zat pengatur tumbuh yang memacu pemanjangan sel,
perkecambahan biji, inisiasi pembungaan, dan pemasakan buah. Rizobakteri
menghasilkan giberelin yang berperan untuk memacu pertumbuhan tanaman.
Tanah hutan merupakan salah satu sumber keragaman rizobakteri. Pandeglang
memiliki sumber daya alam yang belum banyak dieksplorasi terutama rizobakteri
tanah yang menghasilkan
giberelin, salah satunya ialah kawasan Hutan Penelitian
Carita Pandeglang Banten.
Tujuan penelitian ini untuk: (1) mengisolasi rizobakteri penghasil giberelin
asal tanah rizosfer sekitar pohon keruing (
Dipterocarpus
sp.) dari Hutan
Penelitian Carita Pandeglang Banten, (2) menganalisis pengaruh suhu, pH dan
kondisi pencahayaan terhadap pertubuhan sel rizobakteri terpilih dalam
memproduksi giberelin, dan (3) mengidentifikasi isolat terpilih berdasarkan
identifikasi molekuler.
Pengambilan sampel tanah menggunakan metode
Composite sampling
.
Rizobakteri penghasil giberelin diisolasi mengggunakan media seleksi:
Tripticase
Soy Agar
(TSA),
King’s B
,
Nitrogen Free-Base
(NFB), dan
Lactose Glucose
Induce
(LGI). Analisis kandungan giberelin diukur menggunakan
metode
Unyayar
et al.
. Pengaruh faktor lingkungan terhadap pertumbuhan sel dan
produksi giberelin diukur secara bertahap berdasarkan pengaruh: suhu, pH, dan
kondisi cahaya.
Delapan isolat rizobakteri menghasilkan giberelin. Isolat rizobakteri BC2
menghasilkan giberelin tertinggi yaitu: 0.756 mg mL
-1. Isolat rizobakteri BC2
dipilih untuk diidentifikasi berdasarkan karakter fisiologi dan molekuler. Isolat
rizobakteri BC2 dikarakterisasi berdasarkan pengaruh media pertumbuhan sel
terhadap perlakuan suhu, pH, dan kondisi cahaya. Uji fisiologi menunjukkan
bahwa isolat sel rizobakteri BC2 negatif pada produksi indol, urease positif, dan
karbohidrat oksidatif. Analisis pohon filogenetik menunjukkan bahwa isolat sel
rizobakteri BC2 sebagai
Stenotrophomonas maltophilia
dengan tingkat kesamaan
98 %. Pertumbuhan sel dan produksi giberelin isolat rizobakteri BC2 optimum
pada suhu 30 °C, pH 7, dan kondisi cahaya gelap.
Penelitian ini diharapkan dapat menghasilkan isolat rizobakteri penghasil
giberelin unggul yang dapat dimanfaatkan untuk aplikasi praktis sebagai pupuk
hayati.
SUMMARY
HADI SUSILO. Identification and Characterization of Gibberellin Producing
Rhizobacteria Isolated from Forest Soil in Banten.
Supervised byNISA
RACHMANIA MUBARIK
andTRIADIATI.
Gibberellin is plant growth regulator that stimulate cell elongation, seed
germination, flowering, and fruit ripening. Rhizobacteria produce gibberrelin to
improve the plant growth. Forest soil have rhizobacterial biodiversity resources.
Pandeglang district have a lot of natural biodiversity, but not yet to be explored
mostly gibberellins producing rhizobacteria. One of them is Forest Research
Carita, Pandeglang.
This study was conducted: (1) to isolate gibberellin producing rhizobacteria
from forest soil of keruing (
Dipterocarpus
sp.) tree in forest research Carita
Pandeglang Banten, (2) to analyze the effect of temperature, pH, and light
treatments to the growth culture, and (3) to identify selected rizhobacteria with
molecular method.
Soil from rhizosfer was collected by using composite sampling methods.
Tripticase Soy Agar
(TSA),
King’s B
,
Nitrogen Free-Base
(NFB), and
Lactose
Glucose Induce
(LGI) media were used for isolation and selection of
rhizobacteria. Analysis of gibberellin contents was measured by Unyayar
et al.
method. The selected rhizobacteria was identified by using 16S rRNA gene. The
influence of environmental factor to the growth of rhizobacteria and gibberellin
production was measured step by step based on temperature, pH, and light
treatments.
Eight of rhizobacterial isolates produced gibberellins. Among of them BC2
isolate produced the highest of gibberellin (0.756 mg mL
-1). The isolate was
selected to identify based on physiology and molecular, and characterized based
on effect to the growth culture by temperature, pH, and light treatments. The
result of physiological test indicated that BC2 isolate was negative on producing
of indole, positive on urease, and oxidative carbohydrate. The phylogenetic tree
analysis of BC2 isolate was belonged to
Stenotrophomonas maltophilia
with 98 %
similarity level. The environment condition for growth culture media of BC2
isolate was optimum temperature at 30 °C, optimum pH 7, and dark condition.
This research is expected to produce potential gibberellins producing
rhizobacteria which can be exploited for practical application as a biofertilizer.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau
tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan
IPB
IDENTIFIKASI DAN KARAKTERISASI RIZOBAKTERI
PENGHASIL GIBERELIN YANG DIISOLASI DARI TANAH
HUTAN DI BANTEN
HADI SUSILO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2015
Tesis
sebagai salah satu syarat untuk memperoleh gelar
Magister Sains
pada
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya
sehingga penelitian ini berhasil diselesaikan. Penelitian yang berjudul Identifikasi
dan Karakterisasi Rizobakteri Penghasil Giberelin yang Diisolasi dari Tanah
Hutan Di Banten, ini ditujukan sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada Program Studi Bioteknologi Sekolah Pascasarjana IPB
Bogor.
Penulis mengucapkan terima kasih kepada semua pihak yang telah
membantu dalam penyusunan penelitian ini baik secara langsung maupun tidak
langsung. Ucapan terima kasih penulis sampaikan kepada Dr Nisa Rachmania
Mubarik, MSi, sebagai ketua komisi pembimbing, dan Dr Triadiati, MSi, sebagai
anggota komisi pembimbing atas bimbingan dan motivasi yang diberikan kepada
penulis, kepada Prof Dr Ir Suharsono, DEA selaku ketua Program Studi
Bioteknologi IPB yang telah memberi motivasi selama studi, kepada
Dr Rahayu Widyastuti, MSc Agryang telah menjadi dosen penguji dan memberikan saran
penulisan tesis. Kepada DIKTI melalui Beasiswa Program Pendidikan Dalam
Negeri (BPPDN) selama menempuh pendidikan pascasarjana di IPB, dan terima
kasih atas hibah penelitian PDP DIKTI 2015 a.n. Hadi Susilo sehingga penelitian
yang penulis lakukan dapat terlaksana dengan baik.
Ungkapan terima kasih juga disampaikan kepada Istri Hasanah, anak
Hudzaifah Al Faruq dan Hafidz Shalahudin Al Bantani atas doa dan kasih
sayangnya, Ibu Heni dan Bapak Jaka selaku staf Laboratorium Mikrobiologi IPB,
Aldi, Nezharia, Tika, Asa, Rike, Faloe dan teman-teman di Laboratorium
Mikrobiologi IPB serta teman-teman di Sekolah Pascasarjana Bioteknologi
angkatan 2012.
Semoga karya ilmiah ini bermanfaat.
Bogor, November 2015
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
1 PENDAHULUAN
1
Latar Belakang
1
Tujuan Penelitian
2
Manfaat Penelitian
2
2 TINJAUAN PUSTAKA
3
Rizobakteri Pemacu Pertumbuhan Tanaman 3
Giberelin 3
Aplikasi Giberelin 4
Analisis Sekuen Gen 16 S rRNA 4
Pengaruh Pertumbuhan Sel dan Produksi Giberelin 5
3 METODE
7
Tempat dan Waktu Penelitian
7
Bahan
7
Alat
7
Prosedur Kerja
8
4 HASIL DAN PEMBAHASAN
12
Isolasi Rizobakteri
17
Uji Kandungan Giberelin Isolat Rizobakteri
18
Uji Hipersensitivitas Isolat Rizobakteri
18
Identifikasi dan Karakterisasi Isolat Rizobakteri Terpilih
19
Pengaruh Suhu terhadap Pertumbuhan dan Produksi Giberelin
19
Pengaruh pH terhadap Pertumbuhan dan Produksi Giberelin
20
Pengaruh Cahaya terhadap Pertumbuhan dan produksi Giberelin
20
5 SIMPULAN DAN SARAN
22
Simpulan
22
Saran
22
DAFTAR PUSTAKA
23
LAMPIRAN
29
DAFTAR TABEL
1 Mikrob penghasil giberelin pada berbagai kondisi lingkungan
5
2 Karakterisasi isolat rizobakteri asal tanah rizosfer pohon keruing
(
Dipterocarpus
sp.)
9
3 Uji hipersensitivitas isolat rizobakteri BC2 pada daun tembakau
11
4 Karakteristik fisiologi isolat rizobakteri BC2
11
5 Analisis kesamaan sekuen gen 16S rRNA isolat rizobakteri
BC2 menggunakan BLASTN
14
6 Pertumbuhan sel dan produksi giberelin isolat rizobakteri BC2
15
DAFTAR GAMBAR
1 Kerangka penelitian
6
2 Kandungan giberelin yang dihasilkan isolat rizobakteri asal rizosfer
pohon keruing (
Dipterocarpus
sp.)
10
3 Gejala hipersensitivitas isolat rizobakteri BC7 pada daun tembakau
seperti pada
Pseudomonas syringae
sebagai kontrol positif
11
4 Isolat rizobakteri BC2 dengan pewarnaan Gram
12
5 Pita gen 16S rRNA isolat rizobakteri BC2 berukuran ±1300 bp
12
6 Konstruksi pohon filogenetik isolat rizobakteri BC2 dengan metode
Neighbour Joining
12
7 Kurva pertumbuhan dan produksi giberelin isolat rizobakteri BC2
14
8 Pertumbuhan sel dan produksi giberelin isolat rizobakteri BC2 pada
berbagai suhu dengan kondisi pH netral dan cahaya terang
18
9 Pertumbuhan sel dan produksi giberelin isolat BC2 pada berbagai
pH berbeda dengan suhu 27 °C
20
DAFTAR LAMPIRAN
1 Diagram alir pengukuran kandungan giberelin
29
2 Komposisi media seleksi isolat rizobakteri
30
3 Karakterisasi dan jumlah sel/g tanah rizobakteri asal tanah rizosfer
pohon keruing (
Dipterocarpus
sp.)
31
4 Kurva standar pertumbuhan isolat rizobakteri BC2
31
5 Uji kualitas DNA isolat rizobakteri BC2
31
6 Data sekuen DNA isolat rizobakteri BC2
32
1 PENDAHULUAN
Latar Belakang
Giberelin adalah produk penting dalam bioteknologi yang mempunyai nilai ekonomi tinggi, sebagai hormon pertumbuhan tanaman alami banyak digunakan dalam bidang pertanian, pembibitan, pemeliharaan anggur, kultur jaringan, dan perkebunan teh (Bandelier dan Renaud 1997; Sukla et al. 2005). Giberelin adalah senyawa organik kelompok diterpenoid, tersusun dari unit isopren yang terdiri atas 5 atom karbon dengan struktur cincin tulang hidrokarbon (giberelan) dan gugus karboksil bebas. Giberelin diisolasi dari cendawan Gibberella fujikuroi yang patogen pada tanaman padi pertama kali di Jepang (Santner et al. 2009). Biosintesis giberelin di dalam sel melalui jalur asam mevalonat (Gomi dan Matsuoka 2003).
Giberelin pada tumbuhan berfungsi sebagai berikut: mematahkan dormansi pembungaan (Bomke dan Tudzynki 2009; Kang et al. 2014), meningkatkan tinggi tanaman kerdil (Swain dan Singh 2005; Pereg dan McMillan 2015), meningkatkan inisiasi pembungaan (Ribeiro dan Cardoso 2012; Goldberg-Moeller et al. 2013; Sumanasiri et al. 2013), memacu proses perkecambahan biji (Komatsu et al. 2001; Taiz dan Zeiger 2010), meningkatkan pembentukan enzim α-amilase dan pemanjangan sel (Fernie dan Willmitzer 2001; Kazmierczak 2003; Miransari dan Smith 2014). Giberelin dihasilkan oleh tanaman, cendawan (MacMillan 2002); lumut (Ergun et al. 2002), mikroalga (Strik et al. 2014), dan bakteri (Bottini et al. 1989; Atzorn et al. 1998). Bakteri yang mampu menghasilkan giberelin, yaitu: Azotobacter, Azospirillum, Pseudomonas, Acetobacter, Burkholderia, dan Bacillus (Gutierrez-Manero et al. 2001; Mansour et al. 2004).
Widiastuti et al. (1993) melaporkan bahwa giberelin yang disemprotkan dengan konsentrasi 50 ppm dapat meningkatkan hasil Phyllanthus niruri L. selain itu giberelin meningkatkan luas daun Plantago major L. (Khristiyana et al. 2005). Aplikasi giberelin dapat meningkatkan perkecambahan biji Parthenium argentatum Gray (Dissanayake et al. 2010), mematahkan dormansi biji Panicum virgatum L. (Duclos et al. 2014), meningkatkan pembentukan enzim α-amilase pada biji gandum (Kondhare et al. 2014).
2
Rumusan Masalah
Penelitian mengenai isolasi, karakterisasi, dan pengaruh media pertumbuhan rizobakteri indigenous penghasil giberelin di Indonesia sangat sedikit. Hutan Penelitian Carita Banten adalah hutan penelitian yang didominasi oleh pohon keruing (Dipterocarpus sp.) dan belum banyak dieksplorasi keanekaragaman hayatinya, khususnya rizobakteri penghasil giberelin. Sehubungan dengan hal tersebut, maka perlu dilakukan penelitian mengenai rizobakteri penghasil giberelin asal tanah hutan di Carita Banten.
Tujuan Penelitian
Tujuan penelitian ini ialah: (1) mengisolasi sel rizobakteri penghasil giberelin, (2) mengidentifikasi isolat rizobakteri berdasarkan karakter morfologi, fisiologi, dan molekuler berdasarkan gen 16S rRNA, dan (3) menganalisis pengaruh suhu, pH, serta kondisi cahaya terhadap pertumbuhan sel rizobakteri dan produksi giberelin.
Manfaat Penelitian
Penelitian ini diharapkan dapat menghasilkan isolat rizobakteri penghasil giberelin unggul yang dapat dimanfaatkan untuk aplikasi praktis sebagai pupuk hayati.
Hipotesis
2 TINJAUAN PUSTAKA
Rizobakteri Pemacu Pertumbuhan Tanaman
Rizobakteri pemacu pertumbuhan tanaman (RPPT) merupakan kelompok bakteri yang hidup bebas dan menguntungkan yang secara aktif mengkoloni rizosfer untuk memacu pertumbuhan tanaman (Shruti et al. 2013). Menurut Agustian et al. (2010), RPPT berperan penting dalam meningkatkan pertumbuhan, hasil panen, dan kesuburan lahan. RPPT dapat bekerja secara langsung atau tidak langsung. Secara langsung, RPPT merangsang pertumbuhan tanaman dengan menghasilkan hormon pertumbuhan, vitamin, dan asam organik, serta meningkatkan serapan unsur hara bagi tanaman. Secara tidak langsung, RPPT akan menghasilkan senyawa antimikrob patogen yang dapat menekan pertumbuhan cendawan penyebab penyakit tumbuhan dan menghasilkan siderofor (MacMillan 2007; Yadzani et al. 2009; Kumar et al. 2012; Shruti et al. 2013; Kumar et al. 2014). Siderofor adalah senyawa pengompleks Fe3+ atau pengkelat ion logam besi (Fe3+) dan mereduksi Fe3+ menjadi Fe2+ (Ahemad dan Kibret 2014). Kelompok utama dari siderofor adalah asam hidroksamat yang mampu mengikat ion logam besi. Hidroksamat akan mengikat ion logam besi yang tidak larut kemudian ditranspor ke dalam sel. Hidroksamat yang telah dipakai akan ditranspor ke luar sel untuk mengikat ion logam besi lainnya (Madigan et al. 2012).
Potensi RPPT antara lain: mampu memproduksi hormon pertumbuhan yaitu: asam indol asetat, asam giberelin, sitokinin dan etilen; menambat N2; menekan pertumbuhan organisme fitopatogen dengan memproduksi siderofor, kitinase, antibiotik, dan sianida; melarutkan fosfat dan menyediakan nutrien lainnya (Glick 2012; Gunes et al. 2014). RPPT berperan penting dalam memacu pertumbuhan tanaman sereal dan legum untuk meningkatkan hasil tanaman (Perez-Montano et al. 2014), dan kandungan metabolit sekunder (Capellari et al. 2013), selain itu rizobakteri tidak berbahaya terhadap lingkungan (Taufik 2010).
Giberelin
Giberelin termasuk dalam metabolit sekunder, berbentuk kristal, sedikit larut dalam air, dan larut dalam etil asetat. Giberelin pertama kali diisolasi dari cendawan Giberella fujikuroi, dengan nama baru Fusarium fujikuroi (O’Donnell et al. 1998). Biosintesis giberelin berasal dari geranil difosfat dengan dikatalislis enzim ent-Kauren siklase, GA3 β-hidroksilase, dan GA 20-oksidase (Morrone et al. 2009; Marti et al. 2010). Giberelin yang dihasilkan rizobakteri dapat berperan sebagai hormon endogen pada tanaman (Karakoc dan Aksoz 2006). Giberelin dapat memacu sintesis protein kinase pada kacang polong (Pisum sativum L.) yang kerdil (Anggarwal dan Sachar 1995), mereduksi gen kerdil Rht-B1b dan meningkatkan tinggi tanaman Triticum aestivum L. (Rebetzke et al. 2012).
4
disintesis di situs yang berbeda pada bagian tumbuhan seperti pada ujung batang, akar, dan biji (Srivastava 2002). Hormon tumbuhan berperan dalam pengikatan membran protein yang berpotensi untuk meningkatkan aktivitas enzim. Hasil pengikatan ini mengaktifkan enzim tersebut dan mengubah substrat menjadi produk. Produk ini selanjutnya menyebabkan serangkaian reaksi-reaksi sekunder, yang salah satunya adalah pembentukan metabolit sekunder (Puga-Freitas dan Blouin 2014).
Hormon tumbuhan memacu pertumbuhan dengan memberi isyarat kepada sel target untuk membelah dan memanjang. Sebagian besar molekul hormon tumbuhan dapat mempengaruhi metabolisme dan perkembangan sel-sel tumbuhan dengan cara mempengaruhi lintasan sinyal transduksi pada sel target. Hormon tumbuhan bekerja dengan cara mengubah ekspresi gen, mempengaruhi aktivitas enzim, atau dengan cara mengubah ciri dan sifat-sifat membran (Campbell et al. 2003). Pengaruh hormon tumbuhan tergantung pada spesies tumbuhan, situs aksi hormon tumbuhan pada tumbuhan, dan konsentrasi hormon tumbuhan (Watimena 1991; Liu et al. 2013; Gunes et al. 2014).
Aplikasi Giberelin
Aplikasi giberelin merupakan salah satu usaha untuk memaksimalkan pertumbuhan dan hasil tanaman. Aplikasi giberelin dapat memacu pembungaan Crysanthemum sp. (Sumitomo et al. 2009), menginisiasi pembungaan Citrus sp. (Goldberg-Moeller et al. 2013), meningkatkan ukuran diameter tangkai bunga Helleborus sp. (Christiaens et al. 2012), meningkatkan ukuran biji dan kandungan pati gandum (Dian-liang et al. 2013).
Aplikasi giberelin secara eksogen dapat meningkatkan pertumbuhan tanaman cabe (Joo et al. 2005), mematahkan dormansi umbi selama penyimpanan (Liu et al. 2013), meningkatkan ukuran buah dan penundaan kerusakan warna buah Prunus avium L. (Zhang dan Whiting 2011), memperlama masa penyimpanan tomat (Solanum lycopersicum L.) (Ding et al. 2015), meningkatkan produktivitas budidaya kapas (Pereg dan McMillan 2015), meningkatkan produktivitas rumput dan mereduksi emisi NO2 (Whitehead dan Edward 2015).
Analisis Sekuen Gen 16S rRNA
Gen atau genom dua spesies dapat dibandingkan dengan analisis sekuen DNA (DNA sequence analysis), yaitu: membandingan urutan nukleotida bagian dari DNA. Teknik yang dipakai adalah dengan Polymerase chain reaction (PCR), untuk amplifikasi DNA, yang kemudian dilanjutkan dengan pengurutan basa secara otomatis menggunakan mesin pengurut DNA (DNA sequncer). Para ahli sistematika menggunakan data sekuen nukleotida dari DNA nukleus, mtDNA atau keduanya untuk menarik kesimpulan filogeni, membentuk pohon filogenetik, dan mengelompokkan organisme (Madigan et al. 2000).
salah satu molekul RNA yang berperan dalam pembentukan kerangka ribosom dan merupakan organel yang penting dalam proses translasi RNA untuk membentuk asam amino (Glick dan Pasternack 2003).
Molekul rRNA mempunyai sifat homolog, baik secara fungsional maupun evolusinya pada organisme yang berbeda dan merupakan molekul yang strukturnya konservatif. Ukuran molekul rRNA yang cukup besar memudahkan proses isolasi terutama komponen 16S rRNA yang dijadikan sumber analisis filogeni dan klasifikasi makhluk hidup (Broun-Houland et al. 1992). Analisis gen 16S rRNA digunakan untuk menentukan spesies, karena molekul ini terdapat dalam setiap organisme, sehingga dapat dirancang primer universal untuk seluruh kelompok (Pangastuti 2006).
Pengaruh Faktor Lingkungan terhadap Pertumbuhan Sel dan Produksi Giberelin
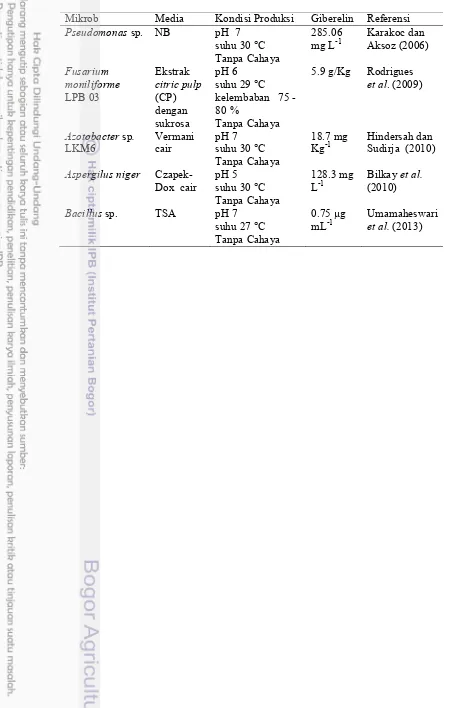
Rizobakteri menghasilkan fitohormon secara ekstraseluler (Bottini et al. 1989; Gray dan Smith 2005; Hindersah dan Sudirja 2010). Rizobakteri dapat menghasilkan giberelin yang optimal bila dipengaruhi oleh beberapa faktor seperti jenis isolat atau galur dan kondisi kultur isolat (Basiacik dan Aksoz 2004). Kondisi kultur isolat dipengaruhi antara lain: pH media pertumbuhan, suhu, waktu inkubasi, dan kondisi inkubasi bergerak atau diam, dan periode gelap atau terang (Bilkay et al. 2010). Mikrob penghasil giberelin (Tabel 1).
6
Tabel 1 Mikrob penghasil giberelin pada berbagai kondisi lingkungan
Mikrob Media Kondisi Produksi Giberelin Referensi Pseudomonas sp. NB pH 7
suhu 30 °C Tanpa Cahaya
285.06
mg L-1 Karakoc dan Aksoz (2006) Fusarium
moniliforme LPB 03
Ekstrak citric pulp (CP) dengan sukrosa
pH 6 suhu 29 °C kelembaban 75 -80 %
Tanpa Cahaya
5.9 g/Kg Rodrigues et al. (2009)
Azotobacter sp.
LKM6 Vermani cair pH 7 suhu 30 °C Tanpa Cahaya
18.7 mg
Kg-1 Hindersah dan Sudirja (2010) Aspergilus niger
Czapek-Dox cair pH 5 suhu 30 °C Tanpa Cahaya
128.3 mg
L-1 Bilkay (2010) et al. Bacillus sp. TSA pH 7
suhu 27 °C Tanpa Cahaya
0.75 µg
3 METODE
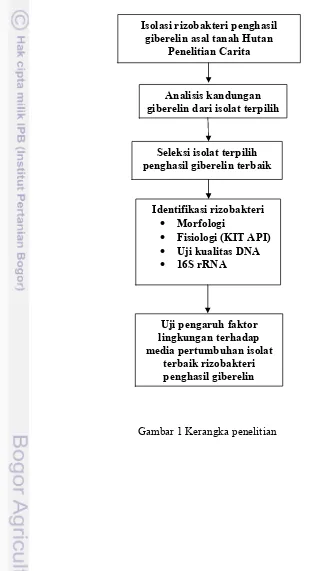
Kerangka Penelitian
Kerangka penelitian meliputi: isolasi rizobakteri dari tanah rizosfer asal Hutan Penelitian Carita, uji kemampuan isolat rizobakteri penghasil giberelin, uji hipersensitivitas, identifikasi isolat terpilih, uji pengaruh faktor lingkungan yaitu: suhu, pH, dan kondisi cahaya gelap terang terhadap pertumbuhan sel rizobakteri dan produksi giberelin (Gambar 1).
Gambar 1 Kerangka penelitian
Isolasi rizobakteri penghasil giberelin asal tanah Hutan
Penelitian Carita
Seleksi isolat terpilih penghasil giberelin terbaik
Identifikasi rizobakteri
Morfologi
Fisiologi (KIT API)
Uji kualitas DNA
16S rRNA
Analisis kandungan giberelin dari isolat terpilih
Uji pengaruh faktor lingkungan terhadap media pertumbuhan isolat
8
Waktu dan Tempat
Penelitian ini dilaksanakan dari bulan Maret sampai Desember 2014. Penelitian dilaksanakan di Laboratorium Mikrobiologi dan Laboratorium Fisiologi Tumbuhan Departeman Biologi, FMIPA, IPB.
Bahan Penelitian
Bahan yang digunakan pada penelitian ini ialah sampel tanah asal rizosfer sekitar perakaran pohon keruing (Dipterocarpus sp.) di Hutan Penelitian Carita, Pandeglang, Banten (0608’ – 06014’LS dan 105050 – 105055’BT).
Pengambilan Sampel Tanah
Sampel tanah diambil dengan menggunakan metode Composite sampling (Hyde et al. 2009) dari 10 titik lokasi pengambilan sampel secara acak dengan jarak ± 1 Km antar titik lokasi pengambilan sampel dan 3 – 5 meter dari batang pohon keruing (Dipterocarpus sp.) di bawah tajuk pohon di kedalaman 15-20 cm dari permukaan tanah yang menempel di perakaran. Pengambilan sampel tanah dilakukan dengan menggunakan bor tanah dengan kondisi bersih dan steril permukaan. Sterilisasi alat dilakukan dengan mencuci peralatan dengan air bersih dan dibilas dengan kapas beralkohol dan dievaporasi dengan nyala api (Saraswati et al. 2007). Tanah kemudian dikompositkan, diambil sebanyak 1 Kg tanah untuk diisolasi rizobakteri penghasil giberelin.
Isolasi Sel Rizobakteri Penghasil Giberelin dari Tanah Hutan
Analisis Kemampuan Produksi Giberelin
Analisis kandungan giberelin diukur secara kuantitatif dengan menggunakan metode Unyayar et al. (1996) (Lampiran 1). Isolat sel rizobakteri ditumbuhkan pada media cair Trypticase Soy Broth (TSB) selama 24 jam inkubasi pada suhu ruang ± 25 °C dan dikocok dengan kecepatan 120 rpm. Sampel kultur isolat bakteri ditimbang 5 g, ditambahkan dengan 100 mL larutan pelarut campuran metanol:kloroform:2N amonium hidroksida (12:5:3 v/v/v). Campuran pelarut ekstrak 100 mL ditambah dengan 22.4 mL air aquades, didiamkan dalam corong pemisah selama 24 jam sampai terjadi pemisahan 2 fase lapisan cairan. Lapisan cair kloroform dibuang, fase cair ekstrak diatur menjadi pH 2.5 dengan menggunakan larutan 5N HCl atau 1N NaOH untuk menaikkan pH. Hasil ekstraksi ditambahkan dengan 15 mL etil asetat sampai 3 kali, didiamkan selama 15 menit sampai terjadi pemisahan 2 lapisan cairan ekstrak, lapisan ekstrak yang mengandung etil asetat kemudian dievaporasi 3 kali masing-masing ditambahkan etil asetat 15mL dengan rotaroevaporator (Buchi Instruments) pada suhu 65 oC. Ekstrak hasil evaporasi kemudian dilarutkan dalam metanol 10 mL. Larutan sampel hasil ekstraksi, dianalisis kandungan giberelinnya dengan menggunakan spektrofotometer (Shimadzu Pharmaspec 1700) pada ג 263 nm.
Uji Hipersensitivitas pada Daun Tembakau
Uji hipersensitif dilakukan pada daun tembakau (Nicotiana tabacum L.) dewasa umur 3 bulan (Vanneste et al. 1990). Isolat sel rizobakteri ditumbuhkan pada media seleksi TSB, LGI dan King’s B cair (107 sel mL-1). Bakteri Pseudomonas syringae digunakan sebagai kontrol positif, karena bersifat patogen pada tanaman, dengan gejala nekrosis pada daun tembakau, sedangkan air dan media kultur sebagai kontrol negatif. Sebanyak 200 µL dari kultur bakteri uji, kontrol positif, dan kontrol negatif diambil menggunakan alat penyuntik steril (tanpa jarum), kemudian disuntikkan ke permukaan bawah daun tembakau. Gejala hipersensitif diamati setelah 48 jam penyuntikan.
Karakterisasi Isolat Rizobakteri
Isolat rizobakteri dikarakterisasi berdasarkan ciri-ciri morfologi dan fisiologi mengikuti Bergey’s Manual of Determinative Bacteriology (Holt et al. 1994). Identifikasi bakteri berdasarkan ciri-ciri biokimia menggunakan kit API® 20NE (bioMeriux, Inc. Durham, USA).
Identifikasi Isolat Sel Rizobakteri Terseleksi Berdasarkan Gen 16S rRNA
10
dilakukan dengan mengikuti prosedur PrestoTMMini gDNA Bakteri Kit (Geneaid). Hasil ekstraksi DNA diukur kosentrasi dan kemurniannya dengan menggunakan NanoDrop 2000 spektrofotometer (Thermo Scientific, Wilmington, DE, USA).
Amplifikasi gen 16S rRNA menggunakan mesin Polymerase Chain Reaction (PCR) (EBSCO) dengan primer 67f (5’-CAG GCC TAA CAC ATG CAA GTC-3’) dan 1387r (5’-GGG CGG WGT GTA CAA GGC-3’) (Marchesi et al. 1998). Total volume untuk PCR yaitu 25 µL terdiri atas: 12.5 µL GoTag Green Master Mix 2X (Promega, Madison, W1, USA), 2.5 µL primer 63f dan 1387r (10 pmol), 6.5 µL Nuclease Free Water dan 1 µL DNA template. Tahap PCR yang dilakukan yaitu: pre-denaturation (95 °C, 5 menit), denaturation (95 °C, 1 menit), annealing (55 °C, 1 menit), elongation (72 °C, 1.5 menit), dan extension (72 °C, 5 menit) dengan total sebanyak 30 siklus. Produk hasil PCR dielektroforesis dengan 1 % (w/v) gel agarosa dengan voltase 80 V selama 45 menit, hasil elektroforesis diamati dengan menggunakan UV transiluminator GelDoc (Labquip) dengan pewarna Ethidium Bromida (EtBr). Penentuan urutan nukleotida dilakukan di Laboratorium 1st Base PT Genetika Science, Singapura dengan mengirimkan sampel DNA isolat BC2. Data sekuen DNA dari jasa sekuensing selanjutnya dilakukan BLASTN (Basic Local Alignment Search Tool Nucleotide) dengan data genom di GenBank kemudian disejajarkan menggunakan program MEGA 5.05. (MegaSoftware, Inc, Arizona, USA). Konstruksi pohon filogenetik dibuat dengan metode Neighbour Joining (NJ) (Altschul et al. 1997).
Kurva Pertumbuhan Sel dan Produksi Giberelin Isolat Terpilih
Kultur starter disiapkan dengan menginokulasikan 2 ose isolat rizobakteri terpilih ke dalam 50 mL kultur media TSB cair, diinkubasi pada suhu ruang ± 27 °C dan kecepatan agitasi 120 rpm hingga kepadatan sel rizobakterinya mencapai 108 CFU mL-1. Selanjutnya, sebanyak 1% inokulum dikultivasi ke dalam media TSB cair produksi 200 mL pada pH 7 pada suhu ruang ± 27 °C dan kecepatan agitasi 120 rpm. Kultur sel isolat terpilih diambil sebanyak 5 mL setiap 3 jam sekali selama 24 jam. Jumlah sel isolat rizobakteri diukur kerapatannya pada ג 590 nm. Hasil pengukuran nilai absorbansi dimasukkan ke dalam persamaan kurva standar jumlah sel rizobakteri (Lampiran 4). Sebanyak 5 mL kultur sel dipanen kemudian diukur produksi giberelin dengan menggunakan metode Unyayar et al. (1996).
Pengaruh Faktor Lingkungan terhadap Pertumbuhan Sel Rizobakteri dan Produksi Giberelin
40 oC, pada agitator, (2) pH media pertumbuhan sel untuk produksi giberelin diatur dengan menggunakan bufer sitrat fosfat 0.2 M untuk pH 5 dan bufer fosfat 0.2 M untuk pH 6, 7, 8; dan (3) kondisi inkubasi cahaya gelap dan terang, kondisi gelap dilakukan dengan penutup aluminium foil dan kondisi terang pada cahaya ruang 400 cd M-2.
Analisis Data
12
4 HASIL DAN PEMBAHASAN
Hasil
Isolasi Rizobakteri Penghasil Giberelin
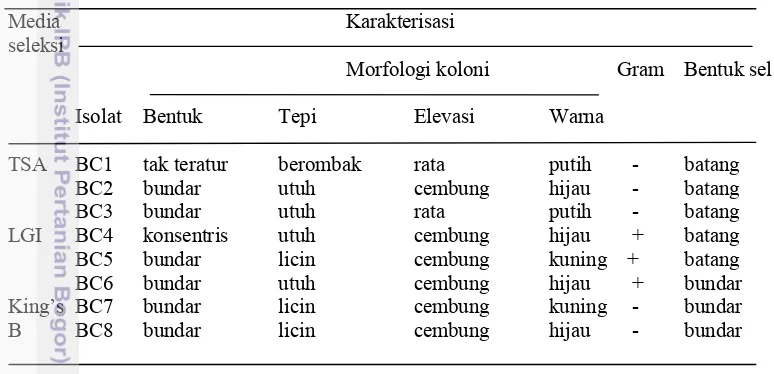
Media pertumbuhan isolat rizobakteri menggunakan media seleksi yaitu: Media TSA, King’s B, LGI, dan media NFB. Media TSA digunakan untuk isolasi Bacillus, King’s B untuk isolasi Pseudomonas, media LGI untuk isolasi Azotobacter, dan media NFB untuk isolasi Azospirillum. Sebanyak 8 isolat rizobakteri dapat tumbuh di media seleksi, yaitu: BC1, BC2, BC3 tumbuh di media TSA, BC4, BC5, BC6 tumbuh di media LGI, BC7, dan BC8 tumbuh di media King’s B. Isolat rizobakteri tidak ada yang tumbuh pada media NFB (Tabel 2).
Tabel 2 Karakterisasi isolat rizobakteri asal tanah rizosfer pohon keruing (Dipterocarpus sp.)
Media Karakterisasi
seleksi
Morfologi koloni Gram Bentuk sel
Isolat Bentuk Tepi Elevasi Warna
TSA BC1 tak teratur berombak rata putih - batang
BC2 bundar utuh cembung hijau - batang
BC3 bundar utuh rata putih - batang
LGI BC4 konsentris utuh cembung hijau + batang
BC5 bundar licin cembung kuning + batang
BC6 bundar utuh cembung hijau + bundar
King’s BC7 bundar licin cembung kuning - bundar
B BC8 bundar licin cembung hijau - bundar
Analisis Kemampuan Produksi Giberelin
Gambar 2 Kandungan giberelin yang dihasilkan isolat rizobakteri asal tanah rizosfer pohon keruing (Dipterocarpus sp.)
Uji Hipersensitivitas pada Daun Tembakau
Uji hipersensitivitas dilakukan untuk mengetahui sifat patogen isolat rizobakteri asal tanah rizosfer akar pohon keruing (Dipterocarpus sp.) terhadap tanaman inang. Hasil uji hipersensitivitas menunjukkan bahwa daun tembakau yang diinjeksi dengan Pseudomonas syringae (kontrol positif) mengalami gejala hipersensitif, yang ditunjukkan dengan perubahan warna daun tembakau dari warna hijau menjadi kuning atau nekrosis (Gambar 3), sedangkan kontrol negatif menunjukkan hasil negatif ditandai dengan tidak terbentuknya perubahan warna pada daun tembakau.
Hasil pengamatan 48 jam setelah diinjeksi, daun tembakau yang diinjeksi perlakuan kultur isolat rizobakteri BC7 dan BC8 menunjukkan gejala hipersensitif di permukaan atas daun tembakau pada daerah penyuntikan, sedangkan 6 isolat rizobakteri lainnya tidak menunjukkan gejala hipersensitif di permukaan atas daun tembakau (Tabel 3).
Tabel 3 Uji hipersensitivitas isolat rizobakteri pada daun tembakau
Gambar 3 Gejala hipersensitivitas isolat
rizobakteri BC7 pada daun tembakau (A) seperti pada Pseudomonas
syringae sebagai kontrol positif (B) Isolat
rizobakteri hipersensitivitas hasil uji
BC1 -
BC2 -
BC3 -
BC4 -
BC5 -
BC6 -
BC7 +
BC8 +
Kontrol
positif +
Kontrol
negatif -
14
Identifikasi dan Karakterisasi Isolat Sel Rizobakteri BC2
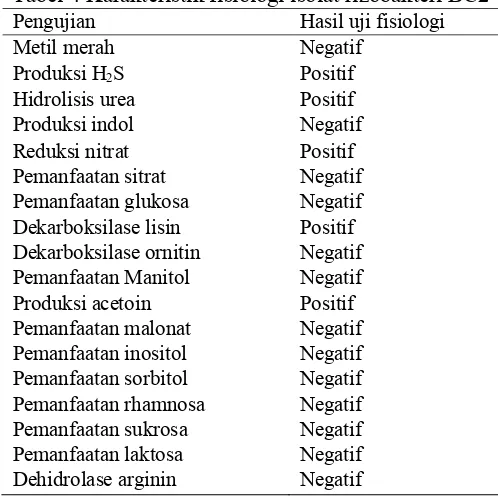
Isolat rizobakteri BC2 dipilih untuk uji lebih lanjut karena tidak menunjukkan gejala hipersensitif pada daun tembakau dan giberelin yang dihasilkan lebih tinggi di antara isolat lainnya yang tidak menunjukkan gejala hipersensitif. Morfologi isolat rizobakteri BC2 dengan pewarnaan Gram menunjukkan bentuk koloni bundar, tepian utuh, bentuk sel batang, ukuran panjang 3 μm, elevasi cembung, dan Gram negatif (Gambar 4). Identifikasi isolat rizobakteri BC2 didasarkan pada karakter fisiologi dan reaksi biokimia yang terjadi dalam metabolisme bakteri (Tabel 4).
Gambar 4 Sel isolat rizobakteri BC2 dengan pewarnaan Gram
Tabel 4 Karakteristik fisiologi isolat rizobakteri BC2
Pengujian Hasil uji fisiologi
Metil merah Negatif
Produksi H2S Positif
Hidrolisis urea Positif
Produksi indol Negatif
Reduksi nitrat Positif
Pemanfaatan sitrat Negatif
Pemanfaatan glukosa Negatif
Dekarboksilase lisin Positif
Dekarboksilase ornitin Negatif
Pemanfaatan Manitol Negatif
Produksi acetoin Positif
Pemanfaatan malonat Negatif
Pemanfaatan inositol Negatif
Pemanfaatan sorbitol Negatif
Pemanfaatan rhamnosa Negatif
Pemanfaatan sukrosa Negatif
Pemanfaatan laktosa Negatif
Dehidrolase arginin Negatif
Hasil uji biokimia menggunakan kit API® 20NE menunjukkan isolat rizobakteri BC2 merupakan Pseudomonas maltophilia dengan tingkat kesamaan 99 %. Hasil uji kualitas DNA isolat rizobakteri BC2 menunjukkan perbandingan ג
260/280 senilai 2.02 (Lampiran 5). Hasil visualisasi amplifikasi gen 16S rRNA pada gel agarosa 0.8 % dihasilkan produk pita DNA dengan ukuran ± 1300 pasang basa (Gambar 5).
Gambar 5 Pita gen 16S rRNA isolat BC2 berukuran ±1300 bp, M=Marker 1kb
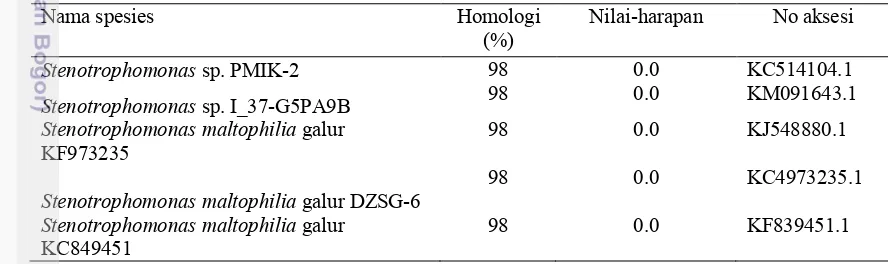
Analisis sekuen gen 16S rRNA (Lampiran 6 ) dengan data pada GenBank pada program BLAST-N menunjukkan bahwa isolat rizobakteri BC2 termasuk ke dalam genus Stenotrophomonas dengan nilai kesamaan 98 % (Tabel 5).
Tabel 5 Analisis kesamaan sekuen gen 16S rRNA isolat BC2 menggunakan BLAST-N
Nama spesies Homologi
(%) Nilai-harapan No aksesi
Stenotrophomonas sp. PMIK-2 98 0.0 KC514104.1
Stenotrophomonas sp. I_37-G5PA9B 98 0.0 KM091643.1
Stenotrophomonas maltophilia galur
KF973235 98 0.0 KJ548880.1
Stenotrophomonas maltophilia galur DZSG-6 98 0.0 KC4973235.1
Stenotrophomonas maltophilia galur
KC849451 98 0.0 KF839451.1
16
Stenotrophomonas_maltophilia_KC136833 Xanthomonadaceae_bacteriumJ_N846918
Stenotrophomonas_maltophilia_KF973235 Bacillus_anthracis_KF973291
Xanthomonas_sp_GQ381284 hadi
Stenotrophomonas_maltophilia_KC849451 45
48 95
31
0.002
Gambar 6 Konstruksi pohon filogenetik isolat rizobakteri BC 2 dengan metode Neighbour-Joining
Kurva Pertumbuhan Sel dan Produksi Giberelin Isolat Rizobakteri BC2
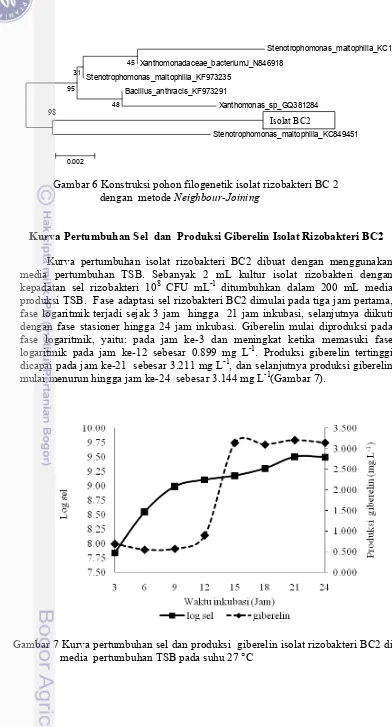
Kurva pertumbuhan isolat rizobakteri BC2 dibuat dengan menggunakan media pertumbuhan TSB. Sebanyak 2 mL kultur isolat rizobakteri dengan kepadatan sel rizobakteri 108 CFU mL-1 ditumbuhkan dalam 200 mL media produksi TSB. Fase adaptasi sel rizobakteri BC2 dimulai pada tiga jam pertama, fase logaritmik terjadi sejak 3 jam hingga 21 jam inkubasi, selanjutnya diikuti dengan fase stasioner hingga 24 jam inkubasi. Giberelin mulai diproduksi pada fase logaritmik, yaitu: pada jam ke-3 dan meningkat ketika memasuki fase logaritmik pada jam ke-12 sebesar 0.899 mg L-1. Produksi giberelin tertinggi dicapai pada jam ke-21 sebesar 3.211 mg L-1, dan selanjutnya produksi giberelin mulai menurun hingga jam ke-24 sebesar 3.144 mg L-1(Gambar 7).
Gambar 7 Kurva pertumbuhan sel dan produksi giberelin isolat rizobakteri BC2 di media pertumbuhan TSB pada suhu 27 °C
Pengaruh Suhu Media terhadap Pertumbuhan Sel dan Produksi Giberelin
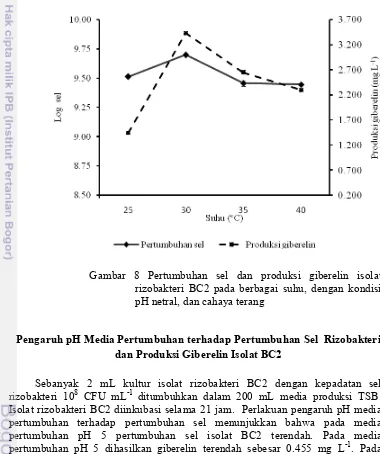
Sebanyak 2 mL kultur isolat rizobakteri BC2 dengan kepadatan sel rizobakteri 108 CFU mL-1 ditumbuhkan dalam 200 mL media produksi TSB. Isolat rizobakteri BC2 diinkubasi selama 21 jam. Hasil perlakuan suhu terhadap media pertumbuhan isolat sel rizobakteri BC2 menunjukkan bahwa pada suhu 25 °C, isolat rizobakteri BC2 mengalami pertumbuhan sel terendah dan dihasilkan giberelin terendah sebesar 1.546 mg L-1, sedangkan pada suhu 30 °C pertumbuhan sel isolat rizobakteri BC2 tertinggi dan dihasilkan giberelin tertinggi sebesar 3.428 mg L-1 (Gambar 8).
Gambar 8 Pertumbuhan sel dan produksi giberelin isolat rizobakteri BC2 pada berbagai suhu, dengan kondisi pH netral, dan cahaya terang
Pengaruh pH Media Pertumbuhan terhadap Pertumbuhan Sel Rizobakteri dan Produksi Giberelin Isolat BC2
18
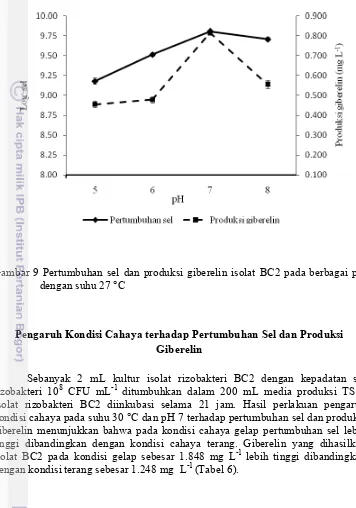
[image:30.595.113.469.148.656.2]media pertumbuhan pH 7 dihasilkan giberelin tertinggi sebesar 0.815 mg L-1 (Gambar 9). Pertumbuhan sel isolat rizobakteri BC2 terjadi peningkatan jumlah sel dan produksi giberelin sampai pada pH 7, kemudian mengalami penurunan jumlah sel dan produksi giberelin sampai pada pH 8.
Gambar 9 Pertumbuhan sel dan produksi giberelin isolat BC2 pada berbagai pH dengan suhu 27 °C
Pengaruh Kondisi Cahaya terhadap Pertumbuhan Sel dan Produksi Giberelin
Tabel 6 Pertumbuhan sel dan produksi giberelin isolat rizobakteri BC2 pada suhu 30 °C, pH 7, dan kondisi cahaya terang dan tanpa cahaya
Kondisi inkubasi Log sel Giberelin (mg L-1)
Terang 9.550 1.248
Gelap 9.809 * 1.848*
Keterangan: tanda * menunjukkan angka beda nyata pada α =5%
Pembahasan
Isolat rizobakteri dapat tumbuh di sekitar rizosfer perakaran tumbuhan. Namun di laboratorium, hanya isolat rizobakteri yang dapat dikulturkan yang dapat tumbuh dan berhasil diisolasi. Pada penelitian ini tidak didapatkan isolat rizobakteri yang tumbuh pada media Nitrogen Free-Base (NFB) yang diduga dipengaruhi oleh ketersediaan sumber nutrisi yang berbeda dengan media Tripticase Soy Agar (TSA), Lactose Glucose Induce (LGI) dan King’s B.
Jumlah populasi sel rizobakteri menunjukkan jumlah populasi yang berbeda pada setiap sampel tanah rizosfer (Lampiran 2). Isolat BC1 jumlah sel/g tanah paling tinggi (4.9 x 106 sel/g tanah) sedangkan isolat BC7 jumlah sel rizobakteri paling rendah (2.5 x 105 sel/g tanah). Keragaman jumlah spesies ditunjukkan dengan adanya isolat-isolat yang dapat tumbuh pada media seleksi. Di alam, eksudat perakaran yang merupakan sumber nutrisi berperan sebagai penghambat dan stimulator terhadap keragaman populasi rizobakteri (Lebuhn et al. 1997) menjadi pembeda, penentu keragaman, dan jumlah populasi di rizosfer tanaman (Broekling et al. 2008; Piromyou et al. 2011; Gunes et al. 2014).
Populasi isolat rizobakteri juga dipengaruhi oleh faktor abiotik seperti pH tanah, suhu, dan kelembapan (Bardgett 2005; Arruda et al. 2013). Semakin banyak keragaman dan jumlah populasi sel rizobakteri semakin menguntungkan tanaman karena dapat menjadi sumber rizobakteri tanaman lainnya (Agustian et al. 2010; Bratkova et al. 2012; Ahemad dan Kibret 2014).
Sebanyak delapan isolat rizobakteri yang diisolasi dari tanah rizosfer pohon keruing (Dipterocarpus sp.) menghasilkan giberelin (Gambar 2) dengan kemampuan yang berbeda dalam menghasilkan giberelin. Rizobakteri diketahui menghasilkan giberelin (Bomke dan Tudzynski 2009). Giberelin merupakan metabolit sekunder yang dihasilkan dalam metabolisme sel sebagai molekul sinyal untuk mengenal inang tumbuhan (Bottini et al. 2004; Ahemad dan Kibret 2014; Puga-Freitas dan Blouin 2014). Kemampuan isolat rizobakteri dalam menghasilkan giberelin tidak sama (Capellari et al. 2013). Hal ini dipengaruhi oleh karakteristik biokimia dan faktor lingkungan (Ahmad et al. 2008; Kumar et al. 2014).
20
warna kekuningan atau nekrosis pada daun tembakau. Hipersensitif merupakan reaksi inang terhadap adanya serangan patogen. Reaksi ini biasanya menyebabkan kematian sebagian sel inang yang bertujuan untuk menghambat pertumbuhan patogen (Lindsay et al. 1993; Arwiyanto et al. 2007).
Analisis kualitas DNA isolat rizobakteri BC2 menunjukkan nilai 2.02 (Lampiran 5). Kualitas DNA yang baik dihitung dengan membandingkan nilai serapan cahaya oleh molekul DNA dengan konsentrasi yang sama (50 mg/ml) ג
260/280 dengan kisaran nilai 1.8-2.0 (Yuwono 2005). Hal ini menunjukkan bahwa kualitas DNA isolat rizobakteri BC2 adalah baik.
Analisis filogenetik menunjukkan bahwa isolat rizobakteri BC2 kekerabatannya dekat dengan Stenotrophomonas maltophilia dengan tingkat kesamaaan 98 % (Gambar 6). Stenotrophomonas maltophilia adalah nama baru dari Pseudomonas maltophilia, bakteri bentuk batang, aerob, Gram negatif, nonpatogen pada tanaman (Palleroni dan Bradbury 1993; Urszula et al. 2009). Stenotrophomonas maltophilia bersifat katalase dan oksidase positif, mengakumulasi β-polihidroksi butirat sebagai sumber karbon, kemoorganotrof (Deswhal et al. 2013) dan memiliki kandungan GC tinggi berkisar 58-68 % (Broun-Howland et al. 1992) ditemukan juga pada rizosfer tanaman tebu (Mehnaz et al. 2010), jagung (Arruda et al. 2013), dan kacang tanah (Sholichatun et al. 2013).
Stenotrophomonas maltophilia termasuk dalam rizobakteri ekstraseluler yang dapat memacu pertumbuhan tanaman dengan menghasilkan fitohormon (Liba et al. 2006), yaitu hormon giberelin (Owen et al. 2015), mampu menginduksi ketahanan bawang merah terhadap hawar daun bakteri (Dunne et al. 1997; Ernita et al. 2010), secara in vitro efektif menekan pertumbuhan Fusarium culmorum (Kamil et al. 2007).
Jumlah sel isolat rizobakteri dapat dihitung dengan menggunakan rapat optis (Optical density atau OD) kemudian dibuat kurva standar jumlah sel (Lampiran 4). Pertumbuhan sel dan produksi giberelin isolat rizobakteri BC2 dipengaruhi oleh suhu inkubasi media pertumbuhan sel. Pada suhu 25 °C pertumbuhan sel rizobakteri BC2 terendah dan produksi giberelin terendah, sedangkan pada suhu 30 °C pertumbuhan sel rizobakteri BC2 tertinggi dan produksi giberelin tertinggi. Hal ini ditunjukkan dengan peningkatan jumlah sel isolat rizobakteri BC2 dan produksi giberelin pada media pertumbuhan sel. Jumlah sel isolat rizobakteri BC2 dan produksi giberelin pada suhu 35 °C lebih kecil dibandingkan pada suhu 25 °C. Jumlah sel rizobakteri BC2 dan produksi giberelin menunjukkan penurunan pada suhu di atas 30 °C. Bakteri dapat tumbuh baik di kisaran suhu 25 °C – 30 °C (Gambar 8).
Jumlah sel isolat rizobakteri BC2 mencapai 9.85 pada suhu 30 °C, pada suhu ini bakteri dapat melangsungkan proses metabolisme dengan baik. Glick (2012) menyatakan bahwa, bakteri peka terhadap suhu lingkungan. Kecepatan reaksi hampir semua metabolisme dapat meningkat dua kali lebih cepat pada setiap kenaikan suhu 10 °C (Murray et al. 2009). Pertumbuhan sel dan produksi giberelin isolat Pseudomonas sp. optimum pada suhu 30 0C (Karakoc dan Aksoz 2006; Shruti et al. 2013).
sedangkan pada pH media 7, pertumbuhan sel dan produksi giberelin tertinggi (Gambar 9). Pseudomonas sp. menghasilkan giberelin sebesar 285,06 mg L-1 pada suhu 30 °C, pH 7, dan 72 jam inkubasi (Karakoc dan Aksoz 2006). Azotobacter sp. LKM6 menghasilkan giberelin sebesar 18,7 mg Kg-1 pada suhu 30 °C, pH 7, dan 48 jam inkubasi (Hindersah dan Sudirja 2010).
Pada saat pH <6 pertumbuhan sel dan produksi giberelin lebih kecil dibanding saat pH 7. Hal tersebut ditunjukkan dengan jumlah sel isolat rizobakteri BC2 dan kadar giberelin yang dihasilkan lebih kecil dibandingkan dengan jumlah sel dan kadar giberelin pada saat pH 7. Pada saat pH >7 mulai terjadi penurunan pertumbuhan sel dan produksi giberelin. Hal ini ditunjukkan dengan turunnya jumlah sel isolat BC2 dan produksir giberelin yang dihasilkan lebih kecil dibandingkan saat pH 7 (Gambar 9). Karakoc dan Aksoz (2006) menyatakan bahwa pertumbuhan sel dan produksi giberelin Pseudomonas sp. optimum pada media pertumbuhan dengan pH, suhu 30 °C, dan kondisi gelap.
Kondisi pH yang asam atau basa dapat menyebabkan perubahan aktivitas metabolisme sel. Perubahan pada pH dapat mengubah penyebaran muatan ion sehingga aktivitas enzim akan mengubah metabolisme dalam sel. Bufer fosfat berfungsi untuk menjaga agar konsentrasi ion H+ pada media kultur cair tidak berubah. Selain itu, K2HPO4 dan MgSO4.7H20 juga berperan sebagai sumber fosfor dan magnesium (Sukmadi 2012).
Pertumbuhan sel dan produksi giberelin isolat rizobakteri BC2 pada suhu inkubasi 30 oC, pH 7, dan kondisi cahaya gelap menghasilkan pertumbuhan sel dan produksi giberelin yang lebih tinggi dibandingkan dengan kondisi cahaya terang (Tabel 5). Hal ini ditunjukkan dengan jumlah sel dan produksi giberelin pada kondisi cahaya gelap lebih tinggi dibandingkan dengan kondisi cahaya terang. Lugterberg dan Kamilova (2009) dan Glick (2012) menyatakan bahwa rizobakteri peka terhadap intensitas cahaya. Cahaya dapat menghambat biosintesis giberelin (Karakoc dan Aksoz 2006; Bomke dan Tudzynki 2009; Kang et al. 2014), termasuk lama penyinaran menghambat biosintesis giberelin (Strik et al. 2014). Pengaturan cahaya dalam biosintesis giberelin melalui gen-gen dioksigenase (Hedden dan Phillip 2000).
22
5 SIMPULAN DAN SARAN
Simpulan
Dari sampel tanah rizosfer akar pohon keruing (Dipterocarpus sp.) di Hutan Penelitian Carita, Kabupaten Pandeglang, Banten diperoleh 8 isolat rizobakteri penghasil giberelin. Isolat rizobakteri BC2 menghasilkan giberelin tertinggi (0.756 mg L-1) dan tidak menyebabkan nekrosis atau bersifat hipersensitif negatif pada daun tembakau. Isolat rizobakteri BC2 bersifat Gram negatif, bentuk sel batang, dengan karakter fisiologi: uji produksi indol negatif, urease positif, dan nitrat positif. Isolat rizobakteri BC2 mempunyai kesamaan 98 % dengan Stenotrophomonas maltophilia KC 849451. Pertumbuhan sel dan produksi giberelin isolat rizobakteri BC2 optimum pada suhu 30 °C, pH 7, dan kondisi cahaya gelap.
Saran
DAFTAR PUSTAKA
Agustian N, Maira L, Emalinda O. 2010. Rizobakteri penghasil fitohormon IAA pada rizosfir tumbuhan semak Karamunting, Titonia, dan tanaman pangan. J Solum. 7(1):49-60.
Ahemad M, Kibret M. 2014. Mechanisms and applications of plant growth promoting rhizobacteria: current perspective. J King Saud Univ. 26:1-20. Ahmad F, Ahmad L, Khan MS. 2008. Screening of free-living rhizospheric
bacteria for their multiple plant growth promoting activities. Microbiol Res. 163:173-181.
Altschul SF, Madden TL, Schaffer AA, Zhang J, Miller W. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25:3389-3402.
Arruda L, Beneduzi A, Martins A, Lisboa B, Lopes C, Bertolo F, Passaglia LMP, Vargas LK. 2013. Screening of rhizobacteria isolated from maize (Zea mays L.) in Rio Grande do Sul State (South Brazil) and analysis of their potensial to improve plant growth. Appl Soil Eco J. 63: 15-22.
Arwiyanto T, Maryudani YMS, Prasetyo AE. 2007. Karakterisasi dan uji aktivitas Bacillus spp. sebagai agensia pengendalian hayati penyakit lincat pada tembakau temanggung. Berk Penel Hayati. 12:93-98.
Atzorn R, Crozier A, Wheeler C, Sandberg G. 1998. Production of gibberellin and indole 3-acetic acid by Rhizobium phasseoli L. in relation to nodulation of Phaseolus vulgaris roots. Planta. 75(9):532-538.
Anggarwal KK, Sachar RC. 1995. Giberrellin stimulates synthesis of a protein kinase in dwarf pea (Pisum sativum L.) epicotyls. Phytochemistry. 40(2):383-387.
Bandelier S, Renaud R. 1997. Production of gibberellic acid by fed-batch solid state fermentation in aseptic pilot scale reactor. Proc Biochem. 32(4):141-145.
Bardgett RD. 2005. The Biology of Soil: A Community and Ecosystem Approach. Oxford (US): Oxford Univ Pr.
Basiacik KS, Aksoz N. 2004. Optimation of carbon-nitrogen ratio for production of gibberellic acid by Pseudomonas sp. J Microbiol. 53(5):117-120.
Bilkay S, Karakoc S, Aksoz N. 2010. Indole -3-acetic aid and gibberellic acid production in Aspergillus niger. Turk J Biol. 34(7):313-318.
Bomke C, Tudzynski B. 2009. Diversity, regulation, and evolution of the gibberellins biosynthetic pathway in fungi compared to plants and bacteria. Phytochemistry. 70:1876-1893.
Bottini R, Fulchieri M, Pearce D, Pharis RP. 1989. Identification of gibberellin A1, A3, and isoA3, in culture of Azospirillum lipoferum. Plant Physiol. 90(7):45-47.
24
Bratkova S, Nikolova D, Evstatieva Y, Dimitrov I, Nikolova K. 2012. Analysis of rhizospheric bacterial community in soil affected by the formation of calcarate. J Geochem Explor. 119:44-50.
Broun-Howland EB, Danielsen SA, Niezwicki-Bouer SA. 1992. Development of rapid method for detecting bacterial cell in situ using 16S rRNA targeted probes. Biotechnique. 13:928-933.
[BPS] Badan Pusat Statistik (ID). 2013. Statistik pertanian Indonesia. [Internet]. [diunduh 2013 Des 17]. Tersedia pada: http://www.bps.go.id/exim-frame.php?kat=2. Pertanian Menurut Komoditi.
Broeckling CD, Broz AK, Bergelson J, Manter DK, Vivanco JM. 2008. Root exudates regulate soil fungal community composition and diversity. Appl Environ Microbiol. 18(8):738-744.
Campbell NA, Reece JB, Mitchell LG. 2003. Biologi. Ed ke-5. Jakarta (ID): Erlangga.
Capellari LR, Santoro MV, Nievaz F, Giordano W, Banchio E. 2013. Increase of secondary metabolite content in marigold by inoculation with plant growth-promoting rhizobacteria. Appl Soil Ecol. 70:16-22.
Christiaens A, Dhooghe E, Pinxteren D, Van Labake MC. 2012. Flower development and effects of a cold treatment and supplemental gibberelilic acid application on flowering on Helleborus niger and Helleborus ericsmithii. Sci Hortic. 136:145-151.
Deshwal VK, Vig K, Amisha, Dwivedi M, Yadav P, Verma M. 2013, Synergistic effects of the inoculation with plant growth-promoting Rhizobium and Pseudomonas on the performance of Mucuna. Annals Forest. 19(1):13-20. Dian-liang P, Tie C, Yan-ping Y, Wei-bing Y, Ying-li N, Dong-qing Y, Zhen-lin
W. 2013. Exogenous application of abscisic acid and gibberellins acid has different effects on starch granule distribution in grain of wheat. J Integrat Agric. 12(9):1551-1559.
Ding Y, Sheng J, Li S, Nie Y, Zhao J, Zhu Z, Wang Z, Tang X. 2015. The role of gibberellins in the mitigation of chilling injury cherry tomato (Solanum lycopersicum L.) fruit. Postharv Biol Technol. 101:88-95.
Dissanayake P, George DL, Gupta ML. 2010. Effect of light, gibberellic acid and absisic acid on germination of guayule (Parthenium argentums Gray) seed. Indust Crops Prod. 32:111-117.
Duclos DV, Altobello CA, Taylor AG. 2014. Investigating seed dormancy in swichgrass (Panicum virgatum L.) elucidating the effect of temperature regimes and plant hormones on embrio dormancy. Indust Crops Prod. 58: 148-159.
Dunne CJ, Crowly J, Moenne-loccoz DN, Dowling FJ, O’Gara F. 1997. Biological control of Phytium ultimum by Stenotrophomonas maltophilia is mediated by an extracellular proteolytic activity. Microbiology. 143:3291-3303.
Ergun N, Topcuolu SF, Yildiz A. 2002. Auxin (Indole-3-acetic acid), gibberellic acid (GA3), absisic acid (ABA) and cytokinin (Zeatin) production by some species of mosses and lichens. Turk J Bot. 26:13-18.
Fernie AR, Willmitzer L. 2001. Molecular and biochemical triggers of potato tuber development. Plant Physiol. 127(12):1459-1465.
Glick BR, Pasternack JJ. 2003. Moleculer Biotechnology, Principles and Applications of Recombinant DNA. Washington (US): ASM Pr.
Glick BR. 2012. Plant Growth Promoting Bacteria Mechanisms and Application. Cairo (EG): Hindawi Publishing.
Goldberg-Moeller R, Shalom L, Shlizerman L, Samuls S, Zur N, Sadka A. 2013. Effects of gibberllin treatment during flowering induction period on global gene expression and the transcription of flowering-control genes in Citrus buds. Plant Sci. 198:46-57.
Gomi K, Matsuoka M. 2003. Gibberellin signaling pathway. Curr Opin Plant Biol. 6(4):489-493.
Gray EJ, Smith DL. 2005. Intercelluler and extracellular PGPR: commonalities and distinctions in plant-bacterium signaling processes. Soil Biol Biochem. 37:395-407.
Gunes A, Turan M, Gulluce M, Sahin F. 2014. Nutritional content analysis of plant growth-promoting rhizobacteria species. Eur J Soil Biol. 60:88-97. Gutierrez-Manero L, Alias-Villegas C, Carerra E. 2001. The
plant-growth-promoting rhizobacteria Bacillus pumilus and Bacillus lichenisformis produce high amounts of physiologically active gibberellins. Physiol Plant 111(9):206-211.
Hedden P, Phillip AL. 2000. Gibberelin metabolism: a new insight revealed by the genes. Trend Plant Sci. 5:12.
Hindersah R, Sudirja R. 2010. Suhu dan waktu inkubasi untuk optimasi kandungan dan fitohormon inokulan cair Azotobacter sp. LKM6. J Natur Indones. 13(1):61-67.
Holt JG, Krieg NR, Sneath PHA, Staley JT, William ST. 1994. Bergey’s Manual of Determinative Bacteriology. Ed ke-9. Baltimore (US): William & Wilkins.
Hyde DK, Aung S, Jeewon R, Pointing BS. 2009. Diversity and abundance of nematode-trapping fungi from decaying litter in terrestrial freshwater and mangrove habitats. J Biodiv Conser. 18:1695-1704.
Joo GJ, Gunes A, Turan M, Gulluce M. 2005. Gibberellins producing rhizobacteria increase endogenous gibberellins content and promote growth of red pepper. J Microbiol. 43:510-515.
Kamil Z, Saleh M, Moustofa S. 2007. Isolation and identification of rhizosphere soil chitinolytic bacteria and their potential in antifungal biocontrol. Global J Mole Sci. 2:57-66.
Kang SM, Radhakrishnan R, Khan AL, Kim M, Park J. 2014. Gibberellin secreting rhizobacterium, Pseudomonas putida H-2-3 moduleates the hormonal and stress physiology of soybean to improve the plant growth under saline and drought conditions. Plant Physiol Biochem. 84:115-124. Karakoc S, Aksoz N. 2006. Some optimal cultural parameter for gibberellic acid
biosynthesis by Pseudomonas sp. Turk J Biol. 30(7):81-85.
26
Khristiyana L, Anggarwulan E, Marsusi. 2005. Pertumbuhan, kadar saponin, dan nitrogen jaringan tanaman daun sendok (Plantago major L.) pada pemberian asam giberelat (GA3). Biofarmasi. 3(1):11-15.
Komatsu S, Yang G, Unno K, Park P. 2001. Characterization of a membrane-associated phosphoprotein (pp47) in rice (Oryza sativa L.) seedling treated by gibberellin. J Plant Physiol. 159:121-128.
Kondhare KR, Hedden P, Kettwell P, Farrell AD, Monaghan JM. 2014. Use of the hormone-biosynthesis inhibitor fluridone and paclobutrazol to determine the
effect of altered absisic acid and gibberellins levels on pre-maturity α-amilase formation in wheat grains. J Cereal Sci. 60:210-216.
Kumar A, Kumar A, Devi S, Pati S, Payal C, Negi S. 2012. Isolation, screening and characterization of bacteria from rhizospheric soils for different plant growth promotion (PGP) activities: an in vitro study. Rec Res Sci Technol. 4(1):01-15.
Kumar A, Maurya BR, Raghuwanshi R. 2014. Isolation and characterization of PGPR and their effects on growth, yield, and nutrient content in wheat (Triticum aestivum L.). Biocatal Agric Biotechnol. 7:234-242.
Lay BW. 1994. Analisa Mikroba di Laboratorium. Ed ke-2. Jakarta (ID): Raja Grafindo Persada.
Lebuhn M, Heulin T, Hartmann A. 1997. Production of auxin and other indolic and phenolic compounds by Phaenibacillus polymixa strains isolated from different proximity to plant root. FEMS Microbiol Ecol. 22:325-334.
Liba CM, Ferrara FIS, Manfio GP, Fantinatti G, Albuquerque RC, Pavan C, Ramos PL. 2006. Nitrogen-fixing chemo-organotrophic bacteria isolated from cyanobacteria-derived lichens and their ability to solubilize phosphate and release amino acids and phytohormones. J Appl Microbiol. 101:1076-1086.
Lindsay WP, Lamb CJ, Dixon RA. 1993. Microbial recognition and activation of plant defence system. Trends Microbiol. 5(1):181-187.
Liu Y, Sun Y, He S, Zhu Y, Ao M, Li J, Cao Y. 2013. Synthesis and characterization of gibberellins-chitosan conjugate for controlled-release applications. Int J Biol Macromol. 57:213-217.
Lugtenberg B, Kamilova F. 2009. Plant growth promoting rhizobacteria. Annu Rev Microbiol. 63:541-556.
MacMillan J. 2002. Occuerence of gibberellin in vascular plants, fungi, and bacteria. J Plant Growth Regul. 20(6):387-443.
MacMillan S. 2007. Promoting Growth with PGPR Soil Foodweb. Vulcan (CA): Soil Biology Laboratory and Learning Centre.
Madigan MT, Martinko JM, Parker J. 2000. Brock Biology of Microorganisms. Upper Sadle River (US): J Wiley.
Madigan M, Martinko J, Stahl D, Clark D. 2012. Brock Biology of Microorganisms. Ed ke-13. New York (US): Pearson.
Marchesi JR, Sato T, Weightman AJ, Martin TA, Fry JC, Hiom SJ, Wade WG. 1998. Design and evaluation of useful bacterium-specific PCR primer that amplify genes coding for bacterial 16S rRNA. Appl Environ Microbiol. 64(2):796-799.
Marti E, Carerra E, Ruiz-Rivero, Garcia-Martines JL. 2010. Hormonal regulation of tomato gibberellins 20-oxidase 1 expressed in Arabidopsis. J Plant Physiol. 167:1188-1196.
Mehnaz S, Baig DN, Bazaroots G. 2010. Genetic and phenotypic diversity of plant growth promoting rhizobacteria isolated from sugarcane plants growing in Pakistan. J Microbiol Biotechnol. 20:1614-1623.
Miransari M, Smith DL. 2014. Plant hormones and seed germination. Environ Exp Bot. 99:110-121.
Morrone D, Chambers J, Lowry L, Kim G, Anterola A, Bender K, Peters RJ. 2009. Gibberellin biosynthesis in bacteria: separate ent-copalyl diphosphate and ent-kaurent synthases in Bradyrhizobium japonicum. FEBS lett. 583:475-480.
Murray RK, Granner DK, Rodwell VW. 2009. Biokimia. Jakarta (ID): EGC. O’Donnell K, Cigelnik E, Nirenberg HI. 1998. Moleculer systematic and
phylogeography of Giberella fujikuroi species complex. Mycologia. 90:465-470.
Owen D, William AP, Griffith GW, Withers PJA. 2015. Use of commercial bio-inoculant to increase agricultural production through improved phosphorus acquisition. Appl Soil Ecol. 86:41-54.
Palleroni NJ, Bradbury JF. 1993. Stenotrophomonas, a new bacterial genus for Pseudomonas maltophilia (Hugh 1980). Int J Syst Bacteriol. 43(3):606-609. Pangastuti A. 2006. Definisi spesies prokaryota berdasarkan urutan basa gen
penyandi 16s rRNA dan gen penyandi protein. Biodiversitas. 7(3):292-296. Pereg L, McMillan M. 2015. Scoping the potential uses of beneficial
microorganism for increasing productivity in cotton cropping systems. Soil Biol Biochem. 80:349-358.
Perez-Montano F, Villegas CA, Bellogin RA, Espuny MR, Cubo OT. 2014. Plant growth promotion in cereal and leguminous agricultural important plant: from microorganism capacities to crop production. Microb Res. 169:325-336.
Piromyu P, Buranabanyat B, Tantasawat P, Tittabutr P, Boonkerd N, Teaumroong N. 2011. Effect of plant growth promoting rhizobacteria (PGPR) inoculation on microbial community structure in rhizosphere of forage corn cultivated in Thailand. Eur J Soil Biol. 47:44-54.
Puga-Freitas R, Blouin M. 2014. A review of effect of soil organisms on plant growth signaling pathways. Environ Exp Bot. 13:2836.
Rao S. 1996. Root Exudation and Rhizosphere Biology. Columbus (US): Ohio State Univ.
28
Ribeiro CM, Cardoso EJBN. 2012. Isolation, selection, and characterization of root-associated growth promoting bacteria in Brazil pine (Araucaria angustifolia L.). Microbiol Res. 167(10):60-78.
Rodrigues C, Vandenberghe LP, Teodoro J, Os JF, Pandey A, Soccol CR. 2009. A new alternative to produce gibberellic acids by solid state fermentation. Braz Arch Biol Technol. 52:181-188.
Santner A, Calderon-Villalobos L, Estelle M. 2009. Plant hormones are versatile chemical regulator of plant growth. Nat Chem Biol. 5:201-2017.
Saraswati R, Husen E, Simanungkalit RDM. 2007. Metode Analisis Biologi Tanah. Bogor (ID): Balai Besar Penelitian dan Pengembangan Sumberdaya Lahan Pertanian.
Shruti K, Arun K, Yuvnet R. 2013. Potential plant growth-promoting activity of rhizobacteria Pseudomonas sp. in Oryza sativa. J Nat Prod Plant Res. 3(4):38-50.
Solichatun, Khalimi K, Sudarma IM. 2013. Isolasi dan identifikasi rizobakteri dari rizosfer kacang tanah dan uji efektifitasnya dalam mengendalikan penyakit layu Fusarium sp. pada tanaman tomat. E-J Agroekoteknol Trop. 2(4): 260-270.
Sukmadi B. 2012. Aktvitas fitohormon indole-3-acetic acid (IAA) dari beberapa isolat bakteri rizosfer dan endofit. J Sains Teknol Indones. 14(3):221-227. Sukla R, Chand S, Srivastava AK. 2005. Batch kinetics and modeling of
gibberellic acid production by Giberella fujikuroi. Enzyme Microbiol Technol. 36(7):492-499.
Srivastava LM. 2002. Plant Growth and Development, Hormones and Environment. California (US): Academic Pr.
Strik WA, Balint P, Tarkowska D, Novak O, Strnad M, Ordog V, Staden JV. 2014. Effect of light on growth and endogenous hormones in Chlorella minutissima (Trebouxiophyceae). Plant Physiol Biochem. 79:66-76.
Sumanasiri H, Krisnarajah SA, Eeswara JP. 2013. Effect of gibberellic acid on growth and flowering of Henckelia humboldtianus Gardner (Ceylon Rock Primrose). Sci Hortic. 159:29-32.
Sumitomo K, Li T, Hisamatsu T. 2009. Gibberellin promotes flowering of Chrysanthemum sp. by upregulating CmFL, a chrysanthemum FLORICAULA/LEAFY homologous gene. Plant Sci. 176:643-649.
Swain SM, Singh DP. 2005. Tall tales from sly dwarves: novel function of gibberellins in plant development. Trends Plant Sci. 1:3.
Taiz L, Zeiger E. 2010. Plant Physiology. Ed ke-5. Sunderland (US): Sinauer. Taufik M. 2010. Pertumbuhan dan produksi tanaman cabai yang diaplikasi
rizobakteri pemacu pertumbuhan tanaman. J Agrivigor. 10(1):99-107. Umamaheswari T, Anbukkarasi K, Hemalatha T, Chendrayan K. 2013. Studies on
phytohormone producing ability of indigenous endophytic bacteria isolated from tropical legume crops. Int J Curr Microbiol Appl Sci. 2(6):127-136. Unyayar S, Topcuoglu SF, Unyayar A. 1996. A modified methods for extraction
Urzula G, Izabela G, Danuta W, Sylwia L. 2009. Isolation and characterization of a novel strain of Stenotrophomonas maltophilia possessing various dioxygenases for monocylic hydrocarbon degradation. Braz J Microbiol. 40:285-291.
Vanneste JL, Paulin JP, Expert D. 1990. Bacteriophage as a genetic tool to study Erwinia amylovora pathogenicity and hypersensitive reaction on tobacco. J Bacteriol. 172(2):932-941.
Wattimena GA. 1991. Zat Pengatur Tumbuh Tanaman. Bogor (ID): PAU IPB. Whitehead D, Edward GR. 2015. Assesment of the application of gibberellin to
increase productivity and reduce nitrous oxide emission in grazed grassland. Agric Eco Environ. 207:40-50.
Widiastuti Y, Hutapea JR, Suhadi. 1993. Usaha peningkatan hasil biomassa Phyllanthus niruri L. melalui pemberian asam giberelat. Warta Tumb Obat Indones. 2(4):1-37.
Yadzani MA, Bahmanyar H, Pirdashti MA, Esmaili. 2009. Effect of phosphate solubilization microorganisms (PSM) and plant growth promoting rhizobacteria (PGPR) on yield and yield component of corn (Zea mays L). World Acad Sci Eng Technol. 3(7):90-92.
Yuwono T. 2005. Biologi Molekuler. Jakarta (ID): Erlangga.
30
LAMPIRAN
Lampiran 1 Diagram alir pengukuran kadar giberelin
Etilasetat (IAA, GA, ABA)
bebas
Etilasetat (zeatin bebas)
Etilasetat (IAA, GA, ABA)
terikat
Dievaporasi
Etilasetat (zeatin terikat)
pH disesuaikan dengan HCl atau NaOH sampai pH 7. Diesktraksi 3 kali dengan etil asetat (15 mL)
pH gabungan (100 mL) disesuaikan dengan HCl atau NaOH sampai pH 2,5. Diekstraksi 3 kali dengan etil asetat (15 mL)
Medium kultur
pH disesuaikan dengan HCl atau NaOH sampai pH 7. Diekstraksi 3 kali dengan etil asetat (15 mL)
pH disesuaikan dengan HCl atau NaOH sampai pH 11. Dihidrolisis selama 1 jam pada 70 oC
pH disesuaikan dengan HCl atau NaOH sampai pH 2,5.
Diekstraksi 3 kali dengan etil asetat (15 mL)
Spektro UV-VIS ג 263 nm
Fase air
Fase air
Fase air
Fase air
Dievaporasi
Spektro UV-VIS
ג 263 nm
+
Lampiran 2 Komposisi media seleksi rizobakteri
1. King’s B Medium
Bahan Jumlah
Protease pepton K2HPO4
MgSO4.7H2O Gliserol Agar bacto K2HPO Akuades 20 g 1.5 g 1.5 g 15 mL 20 g 1.5 g 1000 mL
2. NFB Medium
Bahan Jumlah
Asam malat KOH 4 g, K2HPO4 FeSO4
MnSO4.7H2O MgSO4.7H2O NaCl
CaCl Na2Mo.O4
Bromolthymol Blue (BTB) 0.5% (alk) Agar bacto Akuades pH 6.8 5 g 0.5 g 0.05 g 0.01 g 0.1 g 0.02 g 0.01 g 0.002 g 2 mL 1.75 g 1000 mL
3. LGI Medium
Bahan Jumlah
Sukrosa K2HPO4 KH2PO4 CaCl2 Mg3O4 Na2MoO4 FeCl2 CaCO3 Agar bacto
32
Lampiran 3 Karakterisasi dan jumlah sel/g tanah rizobakteri asal rizosfer akar
pohon keruing (
Dipterocarpus
sp.)
Morfologi koloni Gram Jumlah sel/g
Isolat Bentuk Tepi Elevasi Warna
BC1 Tak teratur berombak rata putih - 4.9 x 10 6 BC2 bundar utuh cembung hijau - 4.1 x 10 6 BC3 bundar utuh rata putih - 3.0 x 10 6 BC4 konsentris utuh cembung hijau + 2.5 x 105 BC5 bundar licin cembung kuning + 3.1 x 105 BC6 bundar utuh cembung hijau + 4.9 x 105 BC7 bundar licin cembung kuning - 2.5 x105 BC8 bundar licin cembung hijau - 3.3 x 105
Lampiran 4 Kurva standar log sel isolat rizobakteri BC2
Lampiran 5 Uji kualitas DNA isolat rizobakteri BC2
Sampel Konsentrasi
nukleotida
Unit
A260
A280
A260/280 Jenis sampel
Blanko
0.0
ng/µl -0.001
0.001
-1.15
DNA
BC2
890.3
ng/µl 17.807
8.862
2.01
DNA
Lampiran 6 Data sekuen DNA isolat rizobakteri BC2
34
36
RIWAYAT HIDUP
Penulis dilahirkan di Ngawi pada tanggal 27 Juni 1973 sebagai anak sulung dari enam bersaudara dari pasangan ayah Kusman dan ibu Siti Mardhiyah. Penulis meneruskan ke jenjang sarjana di Jurusan Biologi Lingkungan, Fakultas Biologi Universitas Gadjah Mada Yogyakarta, lulus tahun 1996. Pada tahun 2012, penulis diterima di Program Studi Bioteknologi, Sekolah Pascasarjana IPB dengan Beasiswa Program Pendidikan Dalam Negeri (BPPDN) DIKTI 2012.
Selama kuliah penulis aktif di HIMMPAS IPB tahun 2013 bidang keilmuan. Penulis pernah mengikuti Lomba Karya Tulis Ilmiah Tingkat Dosen se-Banten tahun 2013, mendap