ECENG GONDOK DAN AGENS HAYATINYA,
NEOCHETINA
SPP. (COLEOPTERA: CURCULIONIDAE), TERHADAP
KOMUNITAS TUMBUHAN AKUATIK DAN SERANGGA
S A P D I
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
Saya menyatakan dengan sebenar-benarnya bahwa disertasi saya dengan judul “Implikasi Keberadaan Spesies Asing Invasif Eceng Gondok dan Agens Hayatinya, Neochetina Spp. (Coleoptera: Curculionidae), terhadap Komunitas Tumbuhan Akuatik dan Serangga” adalah karya saya dengan arahan komisi bimbingan, dan bukan hasil jiplakan atau tiruan dari tulisan siapapun serta belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi manapun.
Bogor, 15 Mei 2007
SAPDI. Implikasi Keberadaan Spesies Asing Invasif Eceng Gondok dan Agens Hayatinya, Neochetina spp. (Coleoptera: Curculionidae), terhadap Komunitas Tumbuhan Akuatik dan Serangga. Dibimbing oleh: DAMAYANTI BUCHORI, UTOMO KARTOSUWONDO, dan SUKISMAN TJITROSEMITO.
Introduksi spesies tumbuhan eksotik invasif dan serangga herbivor eksotik sebagai agens hayati dapat menimbulkan dampak negatif terhadap komunitas tumbuhan dan serangga lokal. Tujuan penelitian adalah untuk mempelajari implikasi keberadaan spesies invasif eceng gondok dan agens hayatinya,
Neochetina spp., terhadap komunitas tumbuhan akuatik dan serangga. Penelitian ini terdiri atas empat bagian yaitu: (1) Evaluasi implikasi keberadaan spesies invasif eceng gondok terhadap komunitas tumbuhan akuatik dan serangga, (2) Evaluasi implikasi keberadaan spesies invasif eceng gondok terhadap komunitas serangga herbivor dan parasitoid, (3) Evaluasi persebaran agens hayati
Neochetina spp. dan dampaknya terhadap eceng gondok, dan (4) Studi potensi dampak non-target introduksi agens hayati N. eichhorniae. Penelitian dilakukan pada beberapa ekosistem perairan di Jawa Barat dan DKI Jakarta yang terinfestasi eceng gondok, berlangsung antara Januari 2004 sampai Maret 2006. Pengamatan komunitas tumbuhan akuatik dilakukan dengan analisis vegetasi, sedangkan pengambilan sampel serangga dilakukan dengan jaring ayun serangga dan perangkap nampan kuning. Distribusi dan kelimpahan Neochetina spp. diamati dengan metode koleksi langsung dengan tangan. Implikasi keberadaan agens hayati tersebut terhadap eceng gondok dievaluasi dengan intensitas kerusakan daun dan parameter pertumbuhan eceng gondok. Potensi dampak non-target introduksi N. eichhorniae dievaluasi dengan uji preferensi dan studi ekspansi kisaran inang di lapangan.
Hasil penelitian menunjukkan bahwa keberadaan eceng gondok secara langsung dapat mengakibatkan terjadinya homogenisasi vegetasi akuatik. Kondisi tersebut secara tidak langsung dapat menurunkan kekayaan dan keanekaragaman spesies serangga pada vegetasi akuatik, baik serangga secara umum, serangga herbivor maupun parasitoid.. Ada indikasi yang kuat bahwa keberadaan eceng gondok dapat memicu terjadinya homogenisasi spesies serangga yang berasosiasi dengan komunitas tumbuhan akuatik.
Agens hayati N. eichhorniae telah menyebar secara luas pada lokasi penelitian, kecuali Karawang, sedangkan N. bruchi tidak ditemukan dalam penelitian ini. Ada indikasi bahwa penyebaran N. eichhorniae secara pasif lebih efektif dibandingkan secara aktif sehingga habitat yang terisolasi tidak dapat dikolonisasi oleh kumbang ini. Meskipun N. eichhorniae secara umum telah mapan di lapangan, namun secara relatif kelimpahannya rendah dan tidak dapat menekan populasi eceng gondok.
Hasil pengujian preferensi dan pengamatan insitu pada tumbuhan akuatik dan terestrial di sekitar ekosistem perairan menunjukkan bahwa N. eichhorniae
memiliki preferensi dan tingkat kekhususan inang yang tinggi terhadap eceng gondok. Hal ini juga mengindikasikan bahwa sejauh ini tidak terjadi ekspansi kisaran inang oleh agens hayati tersebut di lapangan. Selain itu, ada indikasi bahwa keberadaan agens hayati N. eichhorniae sama sekali tidak memiliki implikasi terhadap komunitas serangga yang hidup pada habitat eceng gondok. Kata kunci: Komunitas tumbuhan akuatik, komunitas serangga, Neochetina
ABSTRACT
SAPDI. Implication of The Existence of Invasive Alien Species-Waterhyacinth and Its Biocontrol Agent, Neochetina spp. (Coleoptera: Curculionidae), on Aquatic Plant and Insect Communities. Under supervision of: DAMAYANTI BUCHORI, UTOMO KARTOSUWONDO, and SUKISMAN TJITROSEMITO.
Introduction of invasive exotic plant in a local ecosystem may result in changes of plant communities and insect diversity. The objective of the research is to study the implication of the existence of invasive species-waterhyacinth on plant and insect communities. The research consist of four activities, those are: (1) Evaluation of the existance implication of invasive species-waterhyacinth on plant and insect communities, (2) Evaluation of the existance implication of invasive species-waterhyacinth on herbivorous and parasitoid insect communities, (3) Evaluation of the distribution of biocontrol agents Neochetina
spp. and its implication on waterhyacinth, and (4) Studying of the potential nontarget impacts of biocontrol agents N. eichhorniae introduction. The research was done in several freshwater ecosystems infested by waterhyacinth in West Java and DKI Jakarta, conducted during January 2004 to March 2006. Aquatic plant communities was observed by using vegetation analysis, while insect sample collected by sweep net and yellow pan trap. The distribution and abundance of N. eichhorniae was observed by direct hand-colection technique. Its implication on waterhyacinth was evaluated by leaf damage intensity and growth parameter of the plant. The potential nontarget impacts of N. eichhorniae
introduction was evaluated by preference test and study of host range expansion. The result of the research showed that the existence of invasive species-waterhyacinth may directly resulted in homogenization of aquatic vegetation. This condition may also indirectly resulted in decreasing of species richness and diversity of insects, either in general or herbivorous and parasitoid groups. There is a strong indication that the existence of invasive species-waterhyacinth may lead homogenization of insects species associated with aquatic plants.
Biocontrol agents N. eichhorniae has widely distributed in most sampling sites, except in Karawang, whereas N. bruchi was not found in any sampling sites. Our observation also indicated that N. eichhorniae distributed passively so that the weevil couldn’t colonize the isolated habitat or location. N. eichhorniae
has generally established in West Java and DKI Jakarta, however, its abundance was relatively low and couldn’t result in decreasing of waterhyacith population.
The results of the research also indicated that the weevil showed a high preference and host-specific on waterhyacinth, and so far, the host shifting was not accur. Insitu observation showed that this biocontrol agent was not found in any others aquatic and non aquatic plants living around water areas. This data indicated that host range expansion by the weevil was not consist. Bisides that, there is indication that the existance of N. eichhorniae was have no any implication on insect communities living in waterhyacinth habitat.
©
Hak cipta milik Institut Pertanian Bogor, tahun 2007
Hak cipta dilindungi
ECENG GONDOK DAN AGENS HAYATINYA,
NEOCHETINA
SPP. (COLEOPTERA: CURCULIONIDAE), TERHADAP
KOMUNITAS TUMBUHAN AKUATIK DAN SERANGGA
S A P D I
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Entomologi/Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi Ujian Tertutup: Prof. Dr. Ir. Muhammad Achmad Chozin, M.Sc. Penguji Luar Komisi Ujian Terbuka: 1. Dr. Sih Kahono, M.Sc.
Agens Hayatinya, Neochetina Spp. (Coleoptera: Curculionidae), terhadap Komunitas Tumbuhan Akuatik dan Serangga
Nama : S a p d i
NIM : A461020011
Program Studi : Entomologi/Fitopatologi
Disetujui Komisi Pembimbing
Dr. Ir. Damayanti Buchori, M.Sc. Ketua
Prof. Dr. Ir. Utomo Kartosuwondo, MS. Anggota
Dr. Soekisman Tjitrosemito, M.Sc. Anggota
Diketahui Ketua Program Studi
Entomologi/Fitopatologi
Dekan Sekolah Pascasarjana
Dr.Ir. Sri Hendrastuti Hidayat, M.Sc. Prof. Dr.Ir. Khairil Anwar Notodiputro, MS.
مﻴﺣرﻟاﻦﻣﺣرﻟاﷲامﺴﺒ
Syukur alhamdulillah penulis ucapkan ke hadirat Allah Subhanahu Wata’ala, atas rahmat dan karunia-Nya penulis dapat menyelesaikan penelitian dan penulisan disertasi ini. Disertasi ini berjudul “Implikasi Keberadaan Spesies Asing Invasif Eceng Gondok dan Agens Hayatinya, Neochetina Spp. (Coleoptera: Curculionidae), terhadap Komunitas Tumbuhan Akuatik dan Serangga”. Penelitian in berlangsung antara Januari 2004 sampai Maret 2006.
Pada kesempatan ini penulis menyampaikan penghargaan yang setinggi-tingginya dan ucapan terima kasih yang tidak terhingga kepada Komisi Pembimbing yang terdiri dari Dr. Ir. Damayanti Buchori, M.Sc. sebagai ketua, serta Prof. Dr. Ir. Utomo Kartosuwondo, M.S. dan Dr. Sukisman Tjitrosemito, M.Sc. sebagai anggota. Penyelesaian disertasi ini buah dari jerih payah, perhatian dan ketulusan yang tidak pernah putus dari Komisi Pembimbing dalam memberikan bimbingan dan pengarahan, mulai dari penyusunan rencana penelitian, pelaksanaan penelitian, analisis data, hingga penulisannya.
Dalam proses penyelesaian disertasi ini juga tidak terlepas dari kontribusi berbagai pihak. Kepada Dr. Ir. Hermanu Triwidodo. M.Sc. yang telah bersedia menjadi Penguji Luar Komisi pada Ujian Prakualifikasi dan telah banyak memberikan masukan dalam pelaksanaan penelitian, khususnya Uji Preferensi, penulis menyampaikan penghargaan dan terima kasih. Penghargaan dan terima kasih juga penulis sampaikan kepada Prof. Dr. Ir. Muhammad Achmad Chozin, M.Sc. yang telah bersedia meluangkan waktu untuk bertindak sebagai Penguji Luar Komisi pada Ujian Tertutup, serta Dr. Sih Kahono, M.Sc. dan Dr. Sri Sudarmiyati Tjitrosoedirdjo, M.Sc. yang telah bersedia meluangkan waktu untuk bertindak sebagai Penguji Luar Komisi pada Ujian Terbuka. Semua masukan yang telah diberikan memiliki kontribusi yang sangat berarti dalam penyempurnaan penulisan disertasi ini.
Kesempatan untuk mengikuti Program Doktor yang telah diberikan oleh Rektor Universitas Syiah Kuala merupakan suatu kepercayaan dan kehormatan yang tidak ternilai. Demikian juga kesempatan yang telah diberikan oleh Rektor Institut Pertanian Bogor dan Sekolah Pascasarjana IPB (SPs IPB) kepada penulis untuk mengikuti Program Doktor pada SPs IPB. Atas kesempatan yang telah diberikan, penulis menyampaikan terima kasih. Ucapan terima kasih juga disampaikan kepada Ketua Program Studi Entomologi/Fitopatologi dan seluruh Staf Pengajar SPs IPB, serta seluruh Karyawan SPs IPB yang telah memberikan segenap perhatian dan bantuan dengan penuh rasa kekeluargaan selama penulis mengikuti Program Doktor pada SPs IPB.
Badan Rehabilitasi dan Rekonstruksi (BRR) Aceh-Nias, serta Perhimpunan Keluarga Musara Gayo di Jakarta. Atas semua bantuan tersebut penulis menyampaikan terima kasih dan penghargaan yang setinggi-tingginya.
Teriring do’a buat Ayahanda Almarhum Muhammad Kasah, Ibunda Almarhumah Siti Ara, Ibunda Ramlah, Bapak Mertua Almarhum Ir. Taharuddin, dan Ibu Mertua Iwanah Djalil, penulis mengucapkan terima kasih yang setulus-tulusnya buat isteri, Yusnidar, S.Pd., M.Si. dan putri tercinta, Zuhria Purnama, yang selalu memberikan dukungan dan pengorbanan dengan penuh cinta kasih dan keikhlasan. Ucapan terima kasih juga penulis sampaikan kepada seluruh anggota keluarga (Kanda Rosmeity dan Kanda Tantawi, Kanda Jurnalis dan Kanda Mastani, Kanda Suastikawati dan Kanda Suryadi, Kanda Firmansyah, SH. dan Kanda Hawariah, S.Pd., Kanda Siti Hajar, S.Pd. dan Kanda Hasyimi, SH., Adinda Sabariah, A.Md. dan Adinda Akhmad, Adinda Sumarni, A.Md dan Adinda Saukani, Adinda Yanti Yusmita dan Adinda Jailani, Adinda Neni Fitri dan Adinda Usman, Kanda Dra. Asnawani dan Kanda M. Syakir, A.Md., Kanda Almarhumah Nadhirah, SE, M.Si. dan Kanda Almarhum Syaifullah, SE, MBA., Adinda Indar Sri Wahyuni, S.Si. dan Erwan Hadi Putra, SE., Adinda Hasril, S.Pd dan Adinda Jurialita, Adinda Fitria dan Adinda Irsad Juarsah, Adinda Isna, AMK. dan Adinda Armiya Putra, dan Adinda Nuriana) yang selalu memberikan dukungan moril dan mendo’akan penulis dalam mengikuti pendidikan.
Terima kasih juga penulis sampaikan kepada Pamanda Drs. H. A. Basic Djalil, SH., MA., Pamanda Drs. Lukman Hakim, Pamanda Drs. Johar Abadi, Bapak Drs. H. A. Wahab Rachmatsyah, dan Kanda Irwansyah SE, MS., dan Kanda Martis, SE., atas dukungan moril dan materil yang telah diberikan. Terima kasih dan penghargaan juga disampaikan kepada Bandung Sahari, SP, M.Si. dan Ahmad Rizali, SP, M.Si. yang telah banyak memberikan masukan dalam penulisan disertasi ini, baik berupa ide, analisis data, maupun referensi. Demikian juga kepada semua anggota Hibah Penelitian Tim Pascasarjana, Hasmiandy Hamid, SP, M.Si., Ahmad Rizali, SP, M.Si., Alal Huda Jaya Sironi, SP, M.Si., Asmaul Husna, SP, M.Si., Murni Indarwa, M.Si, dan Joseph Toisuta, SP, M.Si., serta seluruh anggota Laboratorium Bioekologi Parasitoid dan Predator DPT Faperta IPB. Ucapan terima kasih juga penulis sampaikan kepada Hery Tabadepu, SP. yang telah banyak membantu dalam pengadaan dan operasional fasilitas penunjang pelaksanaan Sidang Komisi, Ujian Tertutup dan Ujian Terbuka. Semoga semua bantuan yang telah diberikan mendapat balasan yang setimpal dari Allah SWT.
Penulis dilahirkan di Tanjung Pura, Aceh Tengah pada tanggal 15 November 1964, merupakan anak keenam dari sepuluh bersaudara dari pasangan Almarhum Muhammad Kasah dan Almarhumah Siti Ara. Penulis menyelesaikan pendidikan sarjana pada Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas Syiah Kuala, Banda Aceh, tahun 1990. Pada tahun 1999 penulis melanjutkan pendidikan Magister Sains pada Program Studi Entomologi/Fitopatologi Program Pascasarjana Institut Pertanian Bogor (IPB) dan lulus tahun 2002. Pada tahun yang sama penulis diterima untuk melanjutkan pendidikan Program Doktor pada Program Studi Entomologi/Fitofatologi Sekolah Pascasarjana IPB, dengan dukungan biaya pendidikan dari BPPS Ditjen Dikti, Depdiknas Republik Indonesia.
Sejak tahun 1993 sampai sekarang, penulis bekerja sebagai staf pengajar pada Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas Syiah Kuala, Banda Aceh. Penulis tercatat sebagai anggota Perhimpunan Entomologi Indonesia (PEI) sejak tahun 2004 sampai sekarang.
xi
Alur Pemikiran dan Landasan Penelitian... 7
II. TINJAUAN PUSTAKA ... 9
Distribusi dan Pemencaran Spesies Tumbuhan Eksotik Invasif ... 9
Eceng Gondok sebagai Gulma Eksotik Invasif... 11
Hubungan antara Keanekaragaman Tumbuhan dan Serangga... 14
Pengendalian Hayati Gulma Eksotik Invasif ... 15
Neochetina spp. Sebagai Agens Hayati Eceng Gondok... 17
Dampak Non-target Pengendalian Hayati Gulma... 24
III. HOMOGENISASI BIOTIK PADA EKOSISTEM PERAIRAN: IMPLIKASI KEBERADAAN SPESIES INVASIF ECENG GONDOK ... 28
Implikasi keberadaan eceng gondok terhadap komunitas tumbuhan akuatik ... 34
Implikasi keberadaan eceng gondok terhadap komunitas serangga ... 40
Implikasi keberadaan eceng gondok terhadap komunitas semut ... 56
Pembahasan... 62
Kesimpulan ... 70
Daftar Pustaka ... 70
IV. IMPLIKASI KEBERADAAN SPESIES INVASIF ECENG GONDOK TERHADAP KOMUNITAS SERANGGA HERBIVOR DAN PARASITOID... 74
Abstraks ... 74
Abstract ... 74
Pendahuluan ... 75
xii
Hasil ... 79
Implikasi keberadaan eceng gondok terhadap komunitas serangga herbivor ... 79
Implikasi keberadaan eceng gondok terhadap komunitas serangga Hymenoptera parasitoid ... 87
Hubungan antara kekayaan spesies dan kelimpahan individu serangga parasitoid dengan keberadaan serangga herbivor ... 96
Pembahasan... 99
Kesimpulan ... 105
Daftar Pustaka ... 105
V. PENYEBARAN AGENS HAYATI NEOCHETINA SPP. (COLEOPTERA: CURCULIONIDAE) DAN DAMPAKNYA TERHADAP ECENG GONDOK... 108
Abstraks ... 108
Abstract ... 108
Pendahuluan ... 109
Bahan dan Metode ... 111
Hasil ... 113
Penyebaran dan kelimpahan individu Neochetina spp... 113
Implikasi Neochetina eichhorniae terhadap enceng gondok ... 119
Pembahasan ... 124
Kesimpulan ... 132
Daftar Pustaka ... 133
VI. INTRODUKSI NEOCHETINA EICHHORNIAE (COLEOPTERA: CURCULIONIDAE) PADA EKOSISTEM: ADAKAH DAMPAK NONTARGET? ... 136
Potensi dampak non target oleh Neochetina eichhorniae... 143
Keberadaan Neochetina eichhorniae pada tumbuhan akuatik dan terestrial di sekitar ekosistem perairan ... 146
Hubungan antara Neochetina eichhorniae dengan komunitas serangga pada habitat eceng gondok... 147
xiii
DAFTAR TABEL
Halaman
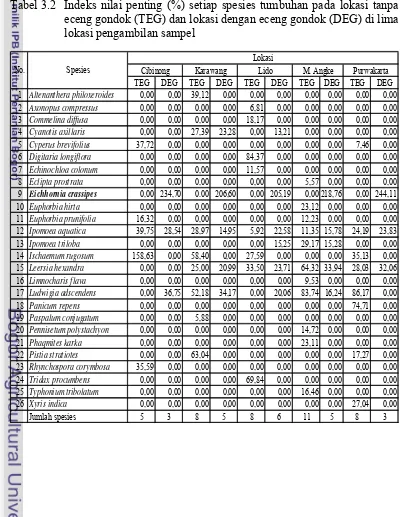
3.1 Deskripsi lokasi penelitian ... 31 3.2 Indeks nilai penting (%) setiap spesies tumbuhan pada lokasi
tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok
(DEG) di lima lokasi pengambilan sampel ... 35 3.3 Kemiripan komunitas tumbuhan (indeks Sorensen) antara lokasi
tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok
(DEG) di lima lokasi pengambilan sampel ... 39 3.4 Komposisi komunitas serangga berdasarkan kekayaan spesies
pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) di lima lokasi pengambilan sampel ... 43 3.5 Kemiripan komunitas serangga (indeks Sorensen) antara lokasi
tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok
(DEG) pada lima lokasi pengambilan sampel... 51 3.6 Komposisi komunitas serangga berdasarkan kekayaan spesies
pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) pada musim hujan dan kemarau ... 54 3.7 Komposisi komunitas semut berdasarkan kelimpahan individu
pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) di lima lokasi pengambilan sampel ... 58 3.8 Matriks kemiripan (indeks Sorensen) komunitas semut antara
lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) di lima lokasi pengambilan sampel ... 59 4.1 Komposisi komunitas serangga herbivor berdasarkan kekayaan
spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
eceng gondok (DEG) di lima lokasi pengambilan sampel... 80 4.2 Kemiripan komunitas serangga herbivor (indeks Sorensen)
antara lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) di lima lokasi pengambilan sampel ... 86 4.3 Komposisi komunitas Hymenoptera parasitoid berdasarkan
kekayaan spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) di lima lokasi pengambilan
sampel ... 89 4.4 Kemiripan komunitas Hymenoptera parasitoid (indeks Sorensen)
antara lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) di lima lokasi pengambilan sampel ... 95 4.5 Daftar famili Hymenoptera parasitoid yang ditemukan pada
xiv
4.6 Korelasi antara kekayaan spesies beberapa famili Hymenoptera parasitoid dengan kekayaan spesies dan kelimpahan individu Homoptera atau Lepidoptera herbivor pada lokasi tanpa eceng
gondok (TEG) dan lokasi dengan eceng gondok (DEG) ... 98 5.1 Populasi kelimpahan individu larva dan imago Neochetina
eichhorniae di empat lokasi pengambilan sampel yang diamati
pada bulan Juli dan Agustus 2004... 116 5.2 Komposisi imago betina Neochetina eichhorniae (% ± SD) pada
empat lokasi pengambilan sampel ... 117 6.1 Spesies tumbuhan yang digunakan pada uji tanpa pilihan... 140 6.2 Jumlah telur Neochetina eichorniae yang diletakkan pada setiap
spesies tumbuhan uji ... 146 6.3 Korelasi Pearson antara kepadatan populasi N. eichhorniae dan
kelimpahan individu setiap ordo serangga yang ditemukan pada
habitat eceng gondok ... 148 6.4 Komposisi komunitas serangga di habitat eceng gondok
berdasarkan kekayaan spesies pada lokasi ditemukan N.
eichhorniae (DNE) dan tidak ditemukan N. eichhorniae (TNE)... 148 6.5 Matriks kemiripan (Indeks Sorensen) komunitas serangga secara
umum pada habitat eceng gondok antara lokasi dengan N.
eichhorniae (DNE) dan lokasi tanpa N. eichhorniae (TNE)... 149 6.6 Matriks kemiripan (Indeks Sorensen) komunitas serangga
herbivor pada habitat eceng gondok antara lokasi dengan N.
eichhorniae (DNE) dan lokasi tanpa N. eichhorniae (TNE)... 150 6.7 Matriks kemiripan (Indeks Sorensen) komunitas serangga
Coleoptera pada habitat eceng gondok antara lokasi dengan N.
xv
DAFTAR GAMBAR
Halaman
1.1 Kerangka penelitian implikasi keberadaan spesies invasif eceng gondok dan agens hayatinya, Neochetina spp., terhadap
komunitas tumbuhan akuatik dan serangga ... 8
2.1 Morfologi eceng gondok (Eichhornia crassipes) ... 12
2.2 Morfologi imago N. eichhorniae dan N. bruchi... 19
2.3 Siklus hidup N. eichhorniae... 21
2.4 Gejala serangan imago N. eichhorniae... 23
2.5 Gejala serangan larva N. eichhorniae... 23
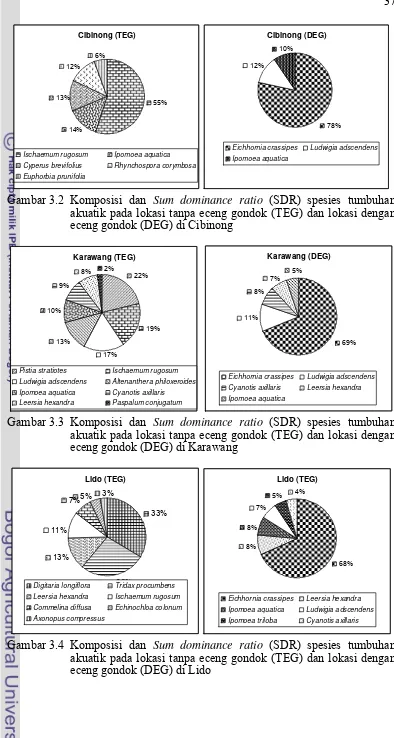
3.1 Total komposisi dan Sum dominance ratio (SDR) spesies tumbuhan akuatik pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) ... 36
3.2 Komposisi dan Sum dominance ratio (SDR) spesies tumbuhan akuatik pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) di Cibinong ... 37
3.3 Komposisi dan Sum dominance ratio (SDR) spesies tumbuhan akuatik pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) di Karawang ... 37
3.4 Komposisi dan Sum dominance ratio (SDR) spesies tumbuhan akuatik pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) di Lido ... 37
3.5 Komposisi dan Sum dominance ratio (SDR) spesies tumbuhan akuatik pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) di Muara Angke... 38
3.6 Komposisi dan Sum dominance ratio (SDR) spesies tumbuhan akuatik pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) di Purwakarta... 38
3.7 Kekayaan spesies tumbuhan akuatik pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) ... 39
3.8 Plot skala dua dimensi yang menunjukkan kemiripan komunitas tumbuhan akuatik pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) di lima lokasi pengambilan sampel ... 40
xvi
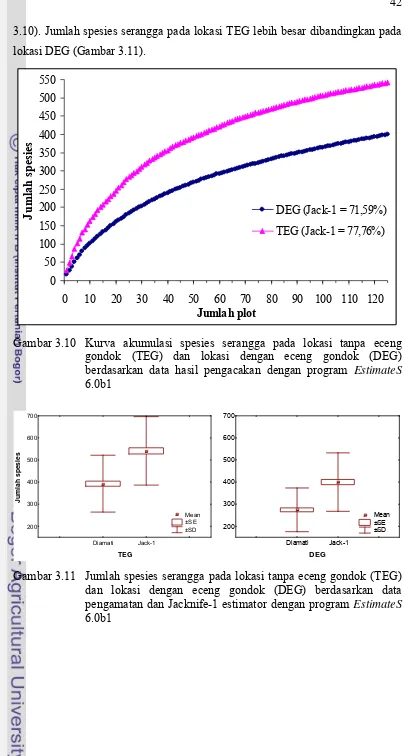
3.10 Kurva akumulasi spesies serangga pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) berdasarkan data hasil pengacakan dengan program EstimateS
6.0b1... 42 3.11 Jumlah spesies serangga pada lokasi tanpa eceng gondok (TEG)
dan lokasi dengan eceng gondok (DEG) berdasarkan data pengamatan dan Jacknife-1 estimator dengan program EstimateS
6.0b1... 42 3.12 Kekayaan spesies serangga pada lokasi tanpa eceng gondok
(TEG) dan lokasi dengan eceng gondok (DEG) ... 44 3.13 Total komposisi komunitas serangga dominan berdasarkan
kekayaan spesies pada lokasi tanpa eceng gondok (TEG) dan
lokasi dengan eceng gondok (DEG) ... 45 3.14 Komposisi komunitas serangga dominan berdasarkan kekayaan
spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
eceng gondok (DEG) di Cibinong... 45 3.15 Komposisi komunitas serangga dominan berdasarkan kekayaan
spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
eceng gondok (DEG) di Karawang ... 46 3.16 Komposisi komunitas serangga dominan berdasarkan kekayaan
spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
eceng gondok (DEG) di Lido... 46 3.17 Komposisi komunitas serangga dominan berdasarkan kekayaan
spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
eceng gondok (DEG) di Muara Angke ... 47 3.18 Komposisi komunitas serangga dominan berdasarkan kekayaan
spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
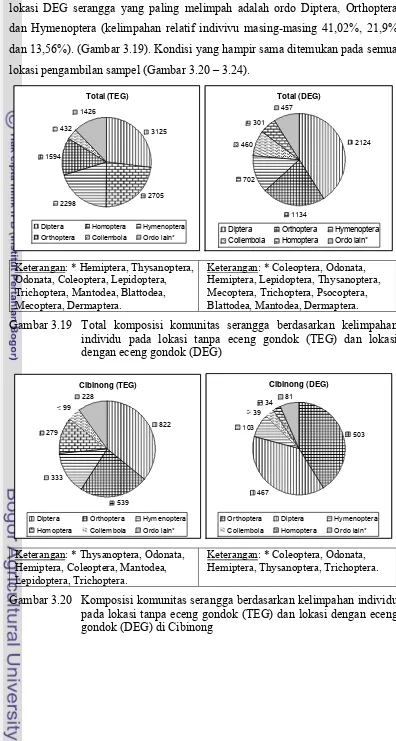
eceng gondok (DEG) di Purwakarta ... 47 3.19 Total komposisi komunitas serangga berdasarkan kelimpahan
individu pada lokasi tanpa eceng gondok (TEG) dan lokasi
dengan eceng gondok (DEG) ... 48 3.20 Komposisi komunitas serangga berdasarkan kelimpahan individu
pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) di Cibinong... 48 3.21 Komposisi komunitas serangga berdasarkan kelimpahan individu
pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) di Karawang ... 49 3.22 Komposisi komunitas serangga berdasarkan kelimpahan individu
pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
xvii
3.23 Komposisi komunitas serangga berdasarkan kelimpahan individu pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) di Muara Angke... 50 3.24 Komposisi komunitas serangga berdasarkan kelimpahan individu
pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) di Purwakarta... 50 3.25 Plot skala dua dimensi yang menunjukkan kemiripan komunitas
serangga pada lokasi tanpa eceng gondok (TEG) dan lokasi
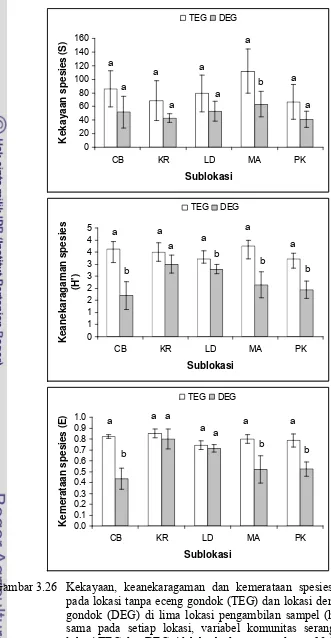
dengan eceng gondok (DEG) di lima lokasi pengambilan sampel ... 52 3.26 Kekayaan, keanekaragaman dan kemerataan spesies serangga
pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) di lima lokasi pengambilan sampel ... 53 3.27 Kekayaan, keanekaragaman dan kemerataan spesies serangga
pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) pada musim kemarau dan hujan ... 55 3.28 Kelimpahan individu setiap spesies semut pada lokasi tanpa
eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) ... 57 3.29 Plot skala dua dimensi yang menunjukkan kemiripan komunitas
semut pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
eceng gondok (DEG) di lima lokasi pengambilan sampel... 60 3.30 Kekayaan, keanekaragaman dan kemerataan spesies semut pada
lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG) di lima lokasi pengambilan sampel ... 61 4.1 Jumlah spesies masing-masing ordo serangga herbivor pada
lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG)... 79 4.2 Kurva akumulasi spesies serangga herbivor pada lokasi tanpa
eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) berdasarkan data hasil pengacakan dengan program EstimateS
6.0b1... 81 4.3 Jumlah spesies serangga herbivor pada lokasi tanpa eceng
gondok (TEG) dan lokasi dengan eceng gondok (DEG) berdasarkan data pengamatan dan Jacknife-1 estimator dengan
program EstimateS 6.0b1 ... 82 4.4 Total komposisi komunitas serangga herbivor berdasarkan
kekayaan spesies pada lokasi tanpa eceng gondok (TEG) dan
lokasi dengan eceng gondok (DEG) ... 82 4.5 Komposisi komunitas serangga herbivor berdasarkan kekayaan
spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
xviii
4.6 Komposisi komunitas serangga herbivor berdasarkan kekayaan spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
eceng gondok (DEG) di Karawang ... 83 4.7 Komposisi komunitas serangga herbivor berdasarkan kekayaan
spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
eceng gondok (DEG) di Lido... 83 4.8 Komposisi komunitas serangga herbivor berdasarkan kekayaan
spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
eceng gondok (DEG) di Muara Angke ... 84 4.9 Komposisi komunitas serangga herbivor berdasarkan kekayaan
spesies pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
eceng gondok (DEG) di Purwakarta ... 84 4.10 Kekayaan, keanekaragaman dan kemerataan spesies serangga
herbivor pada lokasi tanpa eceng gondok (TEG) dan lokasi
dengan eceng gondok (DEG) di lima lokasi pengambilan sampel ... 85 4.11 Plot skala dua dimensi yang menunjukkan kemiripan komunitas
serangga herbivor pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) di lima lokasi pengambilan
sampel ... 87 4.12 Jumlah spesies masing-masing famili Hymenoptera parasitoid
pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng
gondok (DEG)... 88 4.13 Kurva akumulasi spesies Hymenoptera parasitoid pada lokasi
tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) berdasarkan data hasil pengacakan dengan program
EstimateS 6.0b1... 90 4.14 Jumlah spesies Hymenoptera parasitoid pada lokasi tanpa eceng
gondok (TEG) dan lokasi dengan eceng gondok (DEG) berdasarkan data pengamatan dan Jacknife-1 estimator dengan
program EstimateS 6.0b1 ... 90 4.15 Total komposisi komunitas Hymenoptera parasitoid berdasarkan
kekayaan spesies pada lokasi tanpa eceng gondok (TEG) dan
lokasi dengan eceng gondok (DEG) ... 91 4.16 Komposisi komunitas Hymenoptera parasitoid berdasarkan
kekayaan spesies pada lokasi tanpa eceng gondok (TEG) dan
lokasi dengan eceng gondok (DEG) di Cibinong ... 91 4.17 Komposisi komunitas Hymenoptera parasitoid berdasarkan
kekayaan spesies pada lokasi tanpa eceng gondok (TEG) dan
lokasi dengan eceng gondok (DEG) di Karawang ... 92 4.18 Komposisi komunitas Hymenoptera parasitoid berdasarkan
kekayaan spesies pada lokasi tanpa eceng gondok (TEG) dan
xix
4.19 Komposisi komunitas Hymenoptera parasitoid berdasarkan kekayaan spesies pada lokasi tanpa eceng gondok (TEG) dan
lokasi dengan eceng gondok (DEG) di Muara Angke ... 93 4.20 Komposisi komunitas Hymenoptera parasitoid berdasarkan
kekayaan spesies pada lokasi tanpa eceng gondok (TEG) dan
lokasi dengan eceng gondok (DEG) di Purwakarta ... 93 4.21 Kekayaan, keanekaragaman dan kemerataan spesies
Hymenoptera parasitoid pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) di lima lokasi
pengambilan sampel... 94 4.22 Plot skala dua dimensi yang menunjukkan kemiripan komunitas
Hymenoptera parasitoid pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) di lima lokasi
pengambilan sampel... 96 5.1 Peta penyebaran N. eichhorniae di Jawa Barat dan DKI Jakarta... 114 5.2 Kelimpahan individu imago Neochetina eichhorniae per m2 (A)
dan per rumpun eceng gondok (B) di empat lokasi ... 115 5.3 Korelasi antara kelimpahan individu imago dan larva Neochetina
eichhorniae yang ditemukan di empat lokasi pengambilan
sampel ... 116 5.4 Rata-rata jumlah imago betina dan jantan Neochetina
eichhorniae di empat lokasi pada musim hujan dan kemarau ... 117 5.5 Fluktuasi kelimpahan individu imago Neochetina eichhorniae
per m2 (A) dan per rumpun eceng gondok (B) di empat lokasi
dari April sampai Agustus 2004... 118 5.6 Rata-rata kelimpahan individu imago Neochetina eichhorniae
(individu/m2) di empat lokasi pada musim hujan dan kemarau ... 119 5.7 Intensitas kerusakan daun eceng gondok (A) dan jumlah bekas
ketaman (B) yang disebabkan oleh Neochetina eichhorniae... 120 5.8 Korelasi antara bekas ketaman dan kelimpahan individu imago
Neochetina eichhorniae per m2 (A) dan per rumpun eceng
gondok (B) di empat lokasi pengambilan sampel ... 121 5.9 Korelasi antara kelimpahan imago Neochetina eichhorniae dan
populasi eceng gondok... 122 5.10 Korelasi antara kelimpahan imago Neochetina eichhorniae dan
biomassa eceng gondok... 122 5.11 Korelasi antara kelimpahan imago Neochetina eichhorniae dan
jumlah daun eceng gondok... 123 5.12 Korelasi antara kelimpahan imago Neochetina eichhorniae dan
xx
6.1 Lama hidup imago Neochetina eichhorniae pada beberapa
spesies tumbuhan uji ... 143 6.2 Perkembangan aktivitas makan imago Neochetina eichhorniae
pada beberapa spesies tumbuhan uji ... 144 6.3 Perkembangan preferensi Neochetina eichhorniae pada beberapa
DAFTAR LAMPIRAN
Halaman
1 Keberadaan setiap spesies tumbuhan akuatik pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan eceng gondok (DEG) di
lima lokasi pengambilan sampel ... 182 2 Keberadaan spesies dari masing-masing ordo dan famili serangga
dominan pada lokasi tanpa eceng gondok (TEG) dan lokasi dengan
eceng gondok (DEG) di lima lokasi pengambilan sampel... 184 3 Keberadaan setiap spesies semut pada lokasi tanpa eceng gondok
(TEG) dan lokasi dengan eceng gondok (DEG) di lima lokasi
pengambilan sampel... 210 4 Keberadaan spesies serangga herbivor dari masing-masing ordo
dan famili pada lokasi tanpa eceng gondok (TEG) dan lokasi
dengan eceng gondok (DEG) di lima lokasi pengambilan sampel ... 212 5 Keberadaan spesies dari masing-masing famili Hymenoptera
parasitoid pada lokasi tanpa eceng gondok (TEG) dan lokasi
dengan eceng gondok (DEG) di lima lokasi pengambilan sampel ... 217 6 Spesies tumbuhan yang digunakan dalam uji preferensi Neochetina
eichhorniae menggunakan metode dengan dan tanpa pilihan ... 226 7 Gejala kerusakan pada beberapa spesies tumbuhan yang digunakan
dalam uji preferensi Neochetina eichhorniae menggunakan metode
tanpa pilihan ... 233 8 Hasil pengamatan keberadaan Neochetina eichhorniae pada
tumbuhan akuatik di sekitar eceng gondok... 233 9 Hasil pengamatan keberadaan Neochetina eichhorniae pada
Latar Belakang Penelitian
Spesies asing invasif telah menjadi issu hangat yang banyak dibicarakan dalam beberapa tahun belakangan ini, terutama berkaitan dengan spesies tumbuhan invasif. Banyak spesies tumbuhan terbawa oleh manusia ke bagian dunia yang lain, baik secara sengaja maupun tidak. Aktivitas dan mobilitas manusia telah menyebabkan spesies tumbuhan tersebut terbawa dan menyebar ke berbagai belahan bumi. Hal ini dimungkinkan dengan dimulainya era eksplorasi yang dapat menghilangkan penghalang biogeografi, yang sebelumnya mengisolasi biota benua selama jutaan tahun (Mooney & Cleland, 2001). Penyebaran spesies tumbuhan akibat aktivitas manusia telah melebihi yang pernah terjadi sebelumnya dalam sejarah geologi. Ribuan spesies tumbuhan pada saat ini telah mapan secara permanen pada zona fitogeografi yang tidak akan pernah terjadi dalam proses evolusi yang normal (Wilson. 1988)
Introduksi suatu spesies tumbuhan asing dapat menimbulkan masalah yang serius pada habitat yang baru. Setelah keluar dari habitat alaminya, tumbuhan asing tersebut seringkali berkembang menjadi penginvasi agresif yang lebih kompetitif daripada spesies tumbuhan lokal. Beberapa spesies invasif dapat mengubah jalur evolusi dari spesies lokal melalui kompetisi, pemindahan relung, hibridisasi, dan akhirnya kepunahan (Mooney & Cleland, 2001). Schoonhoven et al. (1996) melaporkan bahwa di beberapa bagian dunia, 60-97% gulma merupakan spesies asing, yang telah merusak ekosistem alam atau menimbulkan kehilangan yang nyata bagi produksi pertanian.
Hal ini didasarkan pada kemungkinan terjadinya kompetisi interspesifik. Jika spesies asing introduksi tersebut lebih dominan daripada spesies lokal, besar kemungkinan akan terjadi dampak negatif terhadap keanekaragaman hayati lokal, bahkan tidak mustahil akan terjadi kepunahan spesies Selain itu, beberapa spesies asing mempunyai kemampuan untuk menginvasi dan mendominasi habitat baru sekaligus menggantikan kedudukan spesies lokal. Hal ini disebabkan antara lain karena ketidakhadiran musuh alami dan kompetitornya di habitat yang baru.
Spesies asing invasif pada habitat yang baru dapat menyebabkan terjadinya homogenisasi biotik dan pergantian spesies lokal dengan spesies introduksi (Olden et al. (2004). Hal ini dapat terjadi karena spesies asing invasif mampu beradaptasi dan mendominasi suatu habitat baru yang awalnya didominasi oleh spesies lokal. Salah satu contoh adalah introduksi gulma
Hypericum perforatum L. dari Eropa ke Amerika Utara. Spesies gulma ini secara cepat menyebar pada padang rumput California hingga menutupi 59% komunitas tersebut dan mengakibatkan spesies rumput lokal yang sebelumnya dominan menurun menjadi 22% dari penutupan lahan (Bellows 2001).
Pergeseran komposisi vegetasi akibat invasi spesies asing diprediksi akan mempengaruhi komunitas serangga yang berasosiasi dengan tumbuhan tersebut. Menurut Kruess (2003), keanekaragaman komunitas serangga pada suatu ekosistem sangat dipengaruhi oleh struktur spasial, keanekaragaman habitat dan komposisi habitat. Selanjutnya Altieri dan Nicholls (2004) menyatakan bahwa populasi serangga secara langsung dapat dipengaruhi oleh konsentrasi atau dispersi spasial tumbuhan inang mereka, sebab kemampuan serangga untuk menemukan dan menggunakan tumbuhan inangnya dipengaruhi oleh asosiasi spesies tumbuhan.
spesiesnya. Secara ekologi, serangga memegang peranan penting dalam kelestarian keanekaragaman hayati karena serangga tersebut dapat bertindak sebagai herbivor, karnivor (parasitoid dan predator), polinator, dan dekomposer (Borror et al. 1996; Gullan & Cranston 1994).
Salah satu jenis tumbuhan yang dikenal sangat invasif dan telah menyebar ke seluruh dunia adalah eceng gondok, Eichhornia crassipes (Mart.) Solms. (Pontederiaceae) (Gopal & Sharma 1981; Center et al. 2002). Keberadaan eceng gondok pada suatu ekosistem perairan dikhawatirkan akan menggeser komposisi spesies tumbuhan akuatik lain. Hal ini dapat terjadi karena tingginya daya adaptasi dan laju reproduksi eceng gondok sehingga hampir selalu memenangkan kompetisi dengan spesies tumbuhan lain (Gopal & Sharma 1981; Center et al. 2002). Eceng gondok dapat hidup di daerah tropis dan subtropis (Gopal & Sharma 1981), terdistribusi mulai dari latitude 40o Lintang Utara hingga 40o Lintang Selatan (Center et al. 2002). Populasi eceng gondok berkembang dengan sangat cepat karena tumbuhan ini dapat berreproduksi secara seksual melalui biji dan aseksual dengan stolon. Reproduksi secara aseksual atau vegetatif memegang peranan penting karena dengan cara ini laju pertumbuhan jumlah rumpun eceng gondok dapat berkisar antara 1,2%-13,8% per hari (Gopal & Sharma 1981).
Keberadaan spesies invasif eceng gondok di Indonesia pada saat ini cukup memprihatinkan. Keprihatinan ini cukup beralasan karena sebagian besar ekosistem air tawar di Indonesia telah terinvestasi oleh eceng gondok, meliputi hampir seluruh wilayah mulai dari Sumatera, Jawa, Kalimantan dan Irian Jaya. Tjitrosoedirdjo dan Wijaya (1991) melaporkan bahwa perairan terbuka yang telah terinvestasi oleh eceng gondok meliputi Danau Situ Bagendit, Danau ‘Danau Rawa’, Danau Curug, dan Waduk Saguling di Jawa Barat; Danau Rawa Pening di Jawa Tengah; Waduk Wlingi Raya, Waduk Bureng, dan Sungai Surabaya di Jawa Timur; Danau Kerinci di Jambi; Danau Maninjau di Sumatera Barat; Danau Sentani di Irian Jaya.
pengendalian yang dikembangkan sejak tiga dekade yang lalu, yaitu dengan memanfaatkan musuh alami gulma tersebut yang berupa serangga herbivor. Kegiatan ini dimulai dengan diintroduksinya dua spesies Neochetina spp. (Coleoptera: Curculionidae), yaitu N. eichhorniae (Subagyo et al. 1977) dan N. bruchi (Widayanti et al. 1998). Introduksi dan pelepasan agens hayati tersebut diharapkan dapat menekan perkembangan populasi eceng gondok. Menurut Bellows (2001), introduksi agens hayati berupa serangga herbivor dapat mengurangi dominasi spesies tumbuhan asing invasif dan merestorsi spesies-spesies tumbuhan lokal.
Introduksi serangga herbivor sebagai agens hayati suatu spesies gulma ke suatu ekosistem yang baru perlu mendapat perhatian yang serius sebab dikhawatirkan dapat menimbulkan efek negatif terhadap komunitas spesies bukan sasaran (nontarget effect), baik tumbuhan maupun serangga. Efek nontarget introduksi agens hayati bisa bersifat langsung (direct effect) maupun tidak langsung (indirect effect). Efek langsung terjadi bila agens hayati introduksi tersebut menyerang spesies tumbuhan lain yang bukan target pengendalian. Sementara itu, efek tidak langsung dapat terjadi melalui penggantian ekologi ketika agens hayati secara fisik dan fungsional menggantikan spesies lokal (Pearson & Callaway, 2003).
akibat dari pergantian ekologi, respon kompensasi, dan subsidi jaring-jaring makanan (Pearson & Callaway, 2003).
Masalah lain yang perlu mendapat perhatian adalah fakta bahwa kebanyakan introduksi serangga herbivor untuk pengendalian gulma tidak diikuti dengan upaya monitoring yang serius terhadap distribusinya dan dampaknya terhadap keanekaragaman hayati di lapangan. Tanpa adanya upaya monitoring, degradasi keanekaragaman hayati dalam suatu ekosistem tidak dapat dipantau, terutama tempat-tempat atau habitat yang jauh dari titik pelepasan (Simberloff dan Stiling, 1996).
Introduksi N. eichhorniae ke Indonesia sebagai agens hayati eceng gondok juga perlu mendapat perhatian yang serius. Selain aspek efektivitasnya dalam mengendalikan populasi eceng gondok, evaluasi tentang implikasinya terhadap keanekaragaman hayati lokal juga perlu dilakukan. Apa yang akan terjadi apabila serangga herbivor tersebut menyerang tumbuhan lokal dan mempengaruhi eksistensi keanekaragaman serangga lokal akibat terjadinya kompetisi interspesifik? Pertanyaan ini sangat penting karena dari hasil penelitian diketahui bahwa N. eichhorniae dapat hidup dalam periode tertentu pada beberapa tanaman inang alternatif seperti ganyong (Canna edulis), jahe (Zingiber officinale), galangale (Kaempa galangale) dan temu lawak (Curcuma domestica) (Widayanti et al. 1998).
Bertitik tolak pada uraian di atas, pertanyaan yang saat ini muncul adalah: (1) Seberapa jauh implikasi keberadaan eceng gondok terhadap komunitas tumbuhan akuatik? (2) Seberapa jauh implikasi keberadaan eceng gondok terhadap komunitas serangga? (3) Bagaimana penyebaran dan kelimpahan
Neochetina spp. di lapangan dan bagaimana pengaruhnya terhadap eceng gondok? (4) Apakah ada dampak nontarget introduksi agens hayati Neochetina eichhorniae?
Tujuan Penelitian
Sejalan dengan permasalahan yang telah disebutkan di atas, secara umum penelitian ini bertujuan untuk mengevaluasi implikasi keberadaan spesies invasif eceng gondok dan agens hayatinya, Neochetina spp., terhadap komunitas tumbuhan akuatik dan serangga. Sesuai dengan topik-topik penelitian, secara khusus rangkaian penelitian ini bertujuan untuk mengevaluasi:
1. Implikasi keberadaan spesies invasif eceng gondok terhadap komunitas tumbuhan akuatik.
2. Implikasi keberadaan spesies invasif eceng gondok terhadap komunitas serangga.
3. Penyebaran agens hayati Neochetina spp. dan dampaknya terhadap eceng gondok.
4. Potensi dampak non-target introduksi agens hayati Neochetina eichhorniae.
Manfaat Penelitian
penelitian ini diharapkan dapat menjadi bahan pertimbangan bagi praktisi pengendalian hayati dalam introduksi spesies serangga asing sebagai agens pengendalian hayati gulma.
Alur Pemikiran dan Landasan Penelitian
Ruang lingkup penelitian implikasi spesies invasif eceng gondok dan agens hayatinya, Neochetina spp., terhadap komunitas tumbuhan akuatik dan serangga meliputi empat topik penelitian sebagai berikut:
Penelitian 1: Evaluasi implikasi keberadaan spesies invasif eceng gondok terhadap komunitas tumbuhan akuatik dan serangga.
Penelitian 2: Evaluasi implikasi keberadaan spesies invasif eceng gondok terhadap komunitas serangga herbivor dan parasitoid.
Penelitian 3: Evaluasi penyebaran agens hayati Neochetina spp. dan dampaknya terhadap eceng gondok.
Penelitian 4: Studi potensi dampak non-target introduksi agens hayati
Neochetina eichhorniae.
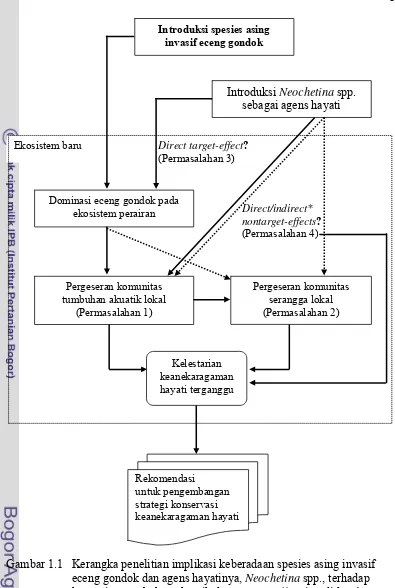
Gambar 1.1 Kerangka penelitian implikasi keberadaan spesies asing invasif eceng gondok dan agens hayatinya, Neochetina spp., terhadap komunitas tumbuhan akuatik dan serangga (*garis solid = direct nontarget-effect; garis putus-putus = indirect nontarget-effect).
Ekosistem baru
Pergeseran komunitas serangga lokal (Permasalahan 2)
Introduksi spesies asing invasif eceng gondok
Dominasi eceng gondok pada ekosistem perairan
Introduksi Neochetina spp. sebagai agens hayati
Pergeseran komunitas tumbuhan akuatik lokal
(Permasalahan 1)
Direct/indirect* nontarget-effects?
(Permasalahan 4)
Rekomendasi
untuk pengembangan strategi konservasi keanekaragaman hayati
Kelestarian keanekaragaman hayati terganggu Direct target-effect?
Distribusi dan Pemencaran Spesies Tumbuhan Asing Invasif
Banyak spesies tumbuhan yang secara sengaja maupun tidak terbawa oleh manusia ke bagian dunia yang lain. Aktivitas dan mobilitas manusia telah menyebabkan spesies tumbuhan ini terbawa dan menyebar ke berbagai belahan bumi. Hal ini dimungkinkan dengan dimulainya era eksplorasi yang dapat menghilangkan penghalang biogeografi yang sebelumnya mengisolasi biota benua selama jutaan tahun (Mooney & Cleland, 2001). Wilson (1988) melaporkan bahwa penyebaran spesies tumbuhan akibat aktivitas manusia telah melebihi yang pernah terjadi sebelumnya dalam sejarah geologi. Ribuan spesies tumbuhan pada saat ini telah mapan secara permanen pada zona fitogeografi yang tidak akan pernah terjadi dalam proses evolusi yang normal.
Setelah keluar dari habitat alaminya, tumbuhan asing tersebut seringkali berkembang menjadi penginvasi agresif yang lebih kompetitif daripada spesies tumbuhan lokal. Beberapa spesies invasif dapat mengubah jalur evolusi dari spesies lokal melalui kompetisi, pemindahan relung, hibridisasi, dan akhirnya kepunahan (Mooney & Cleland, 2001). Schoonhoven et al. (1996) melaporkan bahwa di beberapa bagian dunia, 60-97% gulma merupakan spesies asing, yang mengakibatkan pengaruh merusak dalam ekosistem alam atau menimbulkan kehilangan yang nyata bagi produksi pertanian. Kehilangan hasil pertanian oleh gulma melampaui kehilangan hasil oleh serangga dan pengeluaran untuk herbisida di seluruh dunia 30% lebih tinggi daripada insektisida.
ini secara cepat menyebar pada padang rumput California hingga menutupi 59% komunitas tersebut dan mengakibatkan spesies rumput lokal yang sebelumnya dominan menurun menjadi 22% dari penutupan lahan (Bellows 2001).
Sifat invasif spesies tumbuhan asing dapat terjadi pada tumbuhan akuatik maupun tumbuhan terestrial. Spesies tumbuhan akuatik yang hidup dan berkembang pada habitat perairan dapat dianggap sebagai gulma apabila keberadaannya telah mengganggu kepentingan manusia untuk memanfaatkan perairan, serta gangguan terhadap pemenfaatan habitat perairan oleh flora dan fauna akuatik lokal (Forno & Julien 2000). Banyak contoh tumbuhan akuatik yang menjadi invasif ketika diintroduksi ke luar dari daerah aslinya. Tumbuhan akuatik asing dapat menjadi gulma diduga antara lain terjadi karena tumbuhan tersebut tidak lagi dikendalikan oleh musuh alami, ataupun adanya tekanan kompetisi dari spesies tumbuhan lain yang mempunyai karakter yang mirip. Eceng gondok, Eichhornia crassipes (Mart.) Solms. (Pontederiaceae), merupakan salah satu spesies tumbuhan akuatik paling merusak di dunia, menginvasi danau, kolam, saluran irigasi dan sungai. Menurut beberapa literatur menunjukkan bahwa eceng gondok berasal dari Brasil, sedangkan menurut literatur yang lain menyatakan bahwa eceng gondok adalah tumbuhan asli perairan Amerika Selatan. (Gopal & Sharma 1981).
Spesies invasif eceng gondok menyebar ke berbagai negara dalam kurun waktu akhir abad ke-19 dan awal abad ke-20 dan mengakibatkan degradasi ekosistem perairan (Center et al. 2002). Gopal dan Sharma (1981) menyatakan bahwa pada saat ini tersedia cukup data yang membuktikan bahwa manusia telah membantu distribusi dan pemencaran eceng gondok ke seluruh penjuru dunia, baik secara sengaja maupun tidak.
Jawa Barat (Gopal & Sharma 1981). Tumbuhan akuatik ini diintroduksi sebagai tanaman hias dan penutup kolam ikan (Soerjani et al. 1987).
Pada saat ini, penyebaran eceng gondok di Indonesia sangat luas dan hampir meliputi seluruh wilayah Indonesia, mulai dari Sumatera, Jawa, Kalimantan dan Irian Jaya. Dari laporan inventori pada beberapa perairan terbuka di Indonesia menunjukkan bahwa beberapa danau secara serius terinfestasi oleh eceng gondok. Tjitrosoedirdjo & Wijaya (1991) melaporkan bahwa perairan terbuka yang telah diokupasi oleh eceng gondok meliputi Danau Situ Bagendit, Danau ‘Danau Rawa’, Danau Curug, dan Waduk Saguling di Jawa Barat; Danau Rawa Pening di Jawa Tengah; Waduk Wlingi Raya, Waduk Bureng, dan Sungai Surabaya di Jawa Timur; Danau Kerinci di Jambi; Danau Maninjau di Sumatera Barat; Danau Sentani di Irian Jaya.
Eceng Gondok sebagai Gulma Asing Invasif
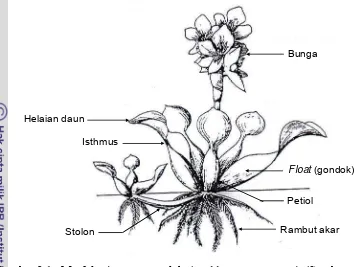
Eceng gondok merupakan tumbuhan mengapung yang tumbuh pada habitat air tawar. Eceng gondok tergolong dalam ordo Liliales, famili Pontederiaceae (Gopal & Sharma 1981). Tumbuhan ini memiliki akar serabut. Daun merumpun mengelilingi pangkal rumpun, hijau mengkilat, dan membulat berbentuk seperti jantung dengan ujung meruncing, lebar daun rata-rata 7,25 cm. Bunga biseksual dan berwarna ungu dengan enam tangkai sari yang melekat pada pembuluh kelopak bunga dengan kepala putik yang panjangnya 1,5–2,0 mm (Soerjani et al. 1987). Morfologi eceng gondok sebagaimana terlihat pada Gambar 2.1.
Isthmus Helaian daun
Rambut akar Stolon
Float(gondok) Bunga
Petiol
Gambar 2.1 Morfologi eceng gondok (Eichhornia crassipes) (Sumber: Gopal & Sharma 1981; Gurure 1999).
Gulma eceng gondok dikenal sangat invasif karena spesies tumbuhan ini memiliki daya adaptasi dan laju reproduksinya yang tinggi. Eceng gondok dapat hidup di daerah tropis dan subtropis (Gopal & Sharma 1981) dan sangat toleran terhadap iklim sedang dan tropis (Julien et al. 1999). Tumbuhan ini terdistribusi mulai dari latitude 40o Lintang Utara hingga 40o Lintang Selatan (Center et al.
2002). Populasi eceng gondok berkembang dengan sangat cepat karena tumbuhan ini dapat berreproduksi secara seksual melalui biji dan aseksual dengan stolon. Reproduksi secara aseksual atau vegetatif memegang peranan penting karena dengan cara ini laju pertumbuhan jumlah rumpun eceng gondok dapat berkisar antara 1,2%-13,8% per hari (Gopal & Sarma 1981).
tumbuhan lokal. Pada daerah perairan, tumbuhan asli dan satwa air tidak dapat bertahan dan mati (Tjitrosemito 1999).
Center et al. (2002) mengindentifikasi dua masalah besar yang dapat timbul akibat keberadaan gulma ini yakni kerugian ekonomi dan kerusakan ekologi. Kerugian ekonomi terjadi karena pertumbuhan gulma yang cepat dapat menutupi wilayah perairan yang luas dan mengakibatkan terhambatnya aktivitas pelayaran pada daerah-daerah yang mengandalkan sarana transportasi air. Biomassa yang besar juga seringkali menutupi saluran irigasi menghambat aliran air. Selain itu, infestasi gulma ini juga dapat menurunkan nilai jual daerah tujuan wisata dan seringkali mengganggu perekonomian masyarakat nelayan perikanan darat. Eceng gondok dapat menyebabkan penurunan pertumbuhan ikan dan tumbuhan air karena rendahnya kandungan oksigen di dalam air. Sebagai contoh, di perairan Sungai Mahakam, Kalimantan Timur, invasi eceng gondok telah menyebabkan daerah perairan yang subur dan kaya ikan lokal menjadi daratan serta mempersempit daerah perairan sehingga menimbulkan bahaya banjir (Tjitrosemito 1999).
Kerusakan ekologi terjadi akibat hamparan eceng gondok yang padat mengurangi cahaya yang mencapai tumbuhan di bawah permukaan air sehingga mengurangi oksigen dalam air. Kondisi ini akan mengurangi fitoplankton yang mengakibatkan perubahan komposisi komunitas invertebrata dan pada akhirnya mempengaruhi perikanan. Daya kompetisi eceng gondok yang kuat dapat mengakibatkan perubahan komposisi vegetasi, merusak tumbuhan lokal, bahkan seringkali mengambil alih habitat tumbuhan liar.
Hubungan antara Keanekaragaman Tumbuhan dan Serangga
Keanekaragaman spesies adalah keanekaan jenis organisme yang menempati suatu ekosistem, baik di darat maupun di lautan (Primack et al.
1998). Lebih jauh mereka menyatakan bahwa keanekaragaman hayati di bumi mencakup keseluruhan makhluk hidup, variasi genetik yang ada di antara individu dalam spesies, komunitas biologi di mana spesies itu hidup, dan interaksi tingkat ekosistem dari komunitas dengan lingkungan fisik dan kimianya.
Dewasa ini, para ahli ekologi telah menjadikan keanekaragaman spesies sebagai tema utama penelitian mereka. Penelitian-penelitian untuk mengkaji pengaruh perubahan kondisi lingkungan terhadap keanekaragaman spesies, serta sebaliknya bagaimana keanekaragaman spesies mempengaruhi stabilitas komunitas alami, telah banyak dilakukan (Schowalter 2000).
Secara umum dapat dikatakan bahwa ada kecenderungan peningkatan keanekaragaman serangga mengikuti peningkatan keanekaragaman tumbuhan. Schoonhoven et al. (1996) melaporkan bahwa spesies serangga herbivor berkorelasi positif dengan jumlah spesies tumbuhan dan ukuran area distribusi tumbuhan. Hal ini dapat dimengerti karena sebagian besar spesies serangga sangat tergantung pada tumbuhan tertentu karena mereka bersifat monofag. Di dalam komunitas biologi setiap spesies menggunakan sumberdaya yang membentuk relung (niche)-nya (Primack et al. 1998). Pemilihan tanaman inang oleh serangga herbivor tergantung pada ciri-ciri tumbuhan tersebut, yang meliputi ciri fisik dan kimianya (Bernays & Chapman 1994).
pergerakan, kolonisasi dan kelimpahan beberapa spesies serangga tertentu (Nicholls et al. 2001).
Tanaman inang suatu serangga juga menjadi faktor penting bagi parasitoid untuk menemukan inangnya. Quicke (1997) melaporkan bahwa pengaruh tanaman inang terhadap tingkat parasitisasi oleh parasitoid Hymenoptera dapat terjadi pada fase penemuan tanaman inang, pencarian inang setelah tanaman inang ditemukan, serta pada perkembangan parasitoid setelah pemarasitan. Selain itu, kebanyakan imago parasitoid Hymenoptera sangat membutuhkan sumber makanan tambahan berupa tepung sari dan nektar yang dapat diperoleh dari tumbuhan berbunga tertentu, serta embun madu yang dihasilkan oleh serangga dari ordo Homoptera (Altieri & Nicolls 2004).
Struktur komunitas tumbuhan dapat mempengaruhi musuh alami serangga yang hidup pada tumbuhan tersebut. Menurut Hawkins (1994), perilaku makan suatu spesies serangga herbivor sangat mempengaruhi jumlah spesies parasitoid yang didukung oleh spesies herbivora tersebut. Selanjutnya Jervis dan Kidd (1996) menyatakan bahwa secara umum musuh alami menunjukkan perilaku yang spesifik dalam pencarian inang, yang sangat dipengaruhi oleh komunitas tumbuhan dimana mangsa atau inangnya hidup.
Pengendalian Hayati Gulma Asing Invasif
Untuk mengendalikan gulma asing dapat digunakan beberapa teknik pengendalian. Dalam kondisi tertentu pengendalian hayati merupakan salah satu teknik pengendalian yang memiliki banyak keuntungan, diantaranya aman bagi lingkungan, agens pengendalian memiliki kemampuan untuk bertahan dan menyebar sendiri, serta biaya pengendalian yang dikeluarkan tidak terlalu besar (Schoonhoven et al. 1996). Dengan keunggulan-keunggulan tersebut, pengedalian hayati dianggap sebagai salah satu teknik pengendalian yang penting dalam mengendalikan gulma, terutama spesies asing.
teknik pengendalian hayati klasik (Forno & Julien 2000). Salah satu contoh pengendalian hayati klasik pada spesies gulma asing terestrial adalah pengendalian kirinyuh (Cromolaena odorata (L.) King & Robinson) menggunakan musuh alminya berupa serangga herbivor, yaitu Pareuchaetes pseudoinsulata Rego Barros (Lepidoptera: Arctiidae) dan Cecidochares connexa
Macquart (Diptera: Tephritidae) (Tjitrosemito 1999).
Pengendalian hayati klasik pada tumbuhan asing invasif dimulai pada tahun 1964 melalui program pengendalian gulma akuatik Alternanthera philoxeroides (Martius) Grisebach (aligator weed). Sejak saat itu, program pengendalian hayati terus dilakukan terhadap gulma tersebut dan beberapa spesies gulma permukaan air lainnya seperti eceng gondok (Eichhornia crassipes), salvinia (Salvinia molesta D.S. Mitchell), kiambang (Pistia stratiotes
L.), dan mata lele (Azolla filiculoides Lamarck), serta tumbuhan bawah permukaan air seperti ganggang hijau (Hydrilla verticillata (L.F.) Royle) dan paris (Myriophyllum spp.) (Forno & Julien 2000).
Menurut Schoonhoven et al. (1996), ada dua kasus yang terkenal yang berhasil dari pengendalian biologi gulma invasif dengan menggunakan serangga herbivor dari habitat asalnya, yaitu pengendalian biologi terhadap gulma
Opuntia dan Salvinia. Opuntia merupakan tanaman yang diintroduksi ke Australia sebagai tanaman pot dan kebun, namun kemudian berkembang menjadi gulma yang sangat invasif. Perkembangan populasi gulma tersebut menurun secara drastis setelah adanya pelepasan ngengat Argentina
Cactoblastis cactorum, yang merupakan pengorok batang kaktus sekitar tahun 1927-1930. Salvinia (Salvinia molesta), merupakan tumbuhan air asli Brazil bagian tenggara yang sejak 1939 disebarkan oleh manusia ke beberapa bagian dunia, baik daerah tropis maupun subtropis (Forno & Julien 2000). Di luar wilayah alaminya, perkembangan tumbuhan ini sangat luar biasa sehingga menimbulkan masalah yang serius. Pelepasan spesies kumbang Cyrtobagous salviniae ke seluruh Australia sangat efektif menurunkan populasi salvinia. Di Afrika dan India penurunan populasi salvinia hampir 100% (Schoonhoven et al.
Tidak semua usaha pengendalian gulma mencapai keberhasilan yang memuaskan seperti yang diuraikan di atas. Bahkan kemapanan populasi agens pengendali biologi tersebut bukan merupakan jaminan terhadap penurunan kelimpahan gulma. Jadi, walaupun sedikitnya 69% dari spesies arthropoda yang dilepaskan mapan pada gulma asing, hanya beberapa saja yang berhasil menekan populasi gulma dan tingkat keberhasilannya beragam pada keadaan yang berbeda (Schoonhoven et al. 1996).
Neochetina spp. Sebagai Agens Hayati Eceng Gondok
Berbagai upaya telah dilakukan di berbagai negara untuk mengendalikan perkembangan populasi eceng gondok. Teknik pengendalian baik secara mekanik maupun kimia pada umumnya tidak berhasil. Pengendalian secara mekanik dengan cara mengangkat dan memindahkan eceng gondok ke lahan kering di sekitar perairan hanya efektif untuk jangka pendek (Kasno et al. 2001). Sementara itu, pengendalian gulma air secara kimia tidak banyak dilakukan karena perairan digunakan untuk berbagai macam keperluan sehingga penggunaan herbisida untuk pengendalian gulma air dikhawatirkan dapat mencemari lingkungan, disamping memerlukan biaya yang cukup besar (Tjitrosoedirdjo 1994). Pengendalian gulma secara kimia juga dapat mengancam kelestarian keanekaragaman hayati. Hill dan Olckers (2001) melaporkan bahwa di Afrika Selatan, kandungan formulasi herbisida yang digunakan untuk pengendalian gulma, khususnya dengan kandungan surfactant yang tinggi, menyebabkan tingginya kematian musuh alami gulma.
Berdasarkan beberapa literatur, Forno & Julien (2000) menyatakan bahwa survey untuk musuh alami eceng gondok dimulai pada tahun 1962 di Amerika Selatan. Sejumlah agens hayati dikoleksi, meliputi kumbang Neochetina bruchi
Hustache (yang belakangan dibedakan menjadi dua spesies, N. eichhorniae dan
N. bruchi), ngengat Xubida (Acigona) infusella (Walker), tungau Orthogalumna terebrantis Walker, ngengat Niphograpta (Sameodes) albiguttalis (Warren), dan kepik Eccritotarsus catarinensis (Carvalho). Keberhasilan pengendalian hayati eceng gondok pertama kali terjadi di White Nile, Sudan setelah dua spesies kumbang Neochetina dan Niphograpta albiguttalis dilepaskan dalam kurun waktu tahun 1978 hingga 1980.
Kumbang N. eichhorniae pertama kali diintroduksi sebagai agens hayati eceng gondok adalah di Amerika Serikat sekitar tahun 1970-an (Perkins 1973
dalam Mangoendihardjo 1978). Pengendalian eceng gondok menggunakan agens hayati N. eichhorniae juga sudah dilakukan di berbagai negara lain, diantaranya Afrika Selatan pada tahun 1974 (Cillers 1991), dan China pada tahun 1995 (Jianqing et al. 2001). Kumbang N. eichhorniae setelah diintroduksi dan dilakukan pelepasan, dapat berkembang dan mapan di daerah baru seperti Afrika Selatan, Uganda, China, dan Kenya (Julien et al. 1999, Julien 2001).
Di Indonesia, pengendalian hayati terhadap eceng gondok juga telah dikembangkan sejak tiga dekade yang lalu, yaitu dengan memanfaatkan musuh alami gulma tersebut yang berupa serangga herbivor. Kegiatan ini dimulai dengan diintroduksinya dua spesies Neochetina spp. (Coleoptera: Curculionidae), yaitu N. eichhorniae (Subagyo et al. 1977) dan N. bruchi
Biologi Neochetina spp.
Neochetina spp. dikenal sebagai kumbang moncong, tergolong ke dalam ordo Coleoptera, famili Curculionidae (Bennett 1970). Secara sepintas imago N. eichhorniae dan N. bruchi sangat mirip, kecuali pada bagian dorsal elitra imago
N. bruchi terdapat suatu motif berbentuk huruf ‘V’ (Gambar 2.2).
Neochetina eichhorniae Neochetina bruchi
Gambar 2.2 Morfologi imago N. eichhorniae dan N. bruchi (Sumber: Julien et al. 1999).
Tubuh imago N. eichhorniae sebagian besar tertutup sisik berwarna abu-abu dengan dua bintik berwarna coklat pada sayap depan. Panjang tubuh imago
1994). Antena berbentuk gada dan berwarna merah kecoklatan (Julien et al. 1999). Menurut Kasno dan Mangoendihardjo (1978), imago yang baru saja muncul dari pupa berwarna coklat dan kemudian berubah menjadi hitam.
Imago N. bruchi memiliki tubuh yang lebih bundar, kokoh, tertutup dengan sisik berwarna coklat yang padat dan menyatu. Pada bagian dorsal elitra terdapat sisik keras berbentuk huruf ‘V’ yang membedakan spesies ini dari N. eichhorniae. Panjang tubuh imago N. bruchi jantan 3,5 mm dan betina 4,5 mm (tidak termasuk kepala). Moncong pada imago jantan agak pipih dan sedikit melengkung sampai lurus, sedangkan imago betinanya memiliki moncong yang lebih bulat, melengkung dan lebih panjang. Bagian ujung moncong pada imago betina kedua spesies lebih mengkilat tanpa ditutupi sisik (Center 1994).
Neochetina spp. termasuk jenis serangga yang aktif pada malam hari, sedangkan pada siang hari bersembunyi di tempat gelap (Center 1994). Imago mulai makan 24 jam setelah muncul dari pupa. Bekas ketaman imago pada tanaman inang berukuran 0,5 mm2 – 2,5 mm2. Imago betina N. eichhorniae
mulai bertelur rata-rata 6 hari setelah menjadi imago (Center 1994; Julien et al. 1999). Sementara itu, dari pengamatan di Bogor ditemukan imago betina kumbang ini baru meletakkan telur setelah berumur 1 bulan (Kasno & Mangoendihardjo 1978). Satu individu betina dapat meletakkan 200-400 telur selama hidupnya (Center et al. 2002). Di Florida, lama waktu generasi kumbang adalah 70 sampai 140 hari (Center 1994), sedangkan di Bogor waktu generasinya adalah 2,5 bulan (Kasno & Mangoendihardjo 1978).
13, 12 minggu; laju pertumbuhan relatif (r) 0,137; laju pertumbuhan (λ) = 1,373 per minggu.
N. eichhorniae mengalami metamorfosis sempurna (holometabola). Perkembangannya dimulai dari telur, larva, pupa dan imago. Siklus hidup dan morfologi N. eichhorniae sebagaimana terlihat pada Gambar 2.3. Telur N. eichhorniae berukuran 0,8 mm x 0,6 mm, berwarna putih, berbentuk oval dan lunak. Telur diletakkan secara tunggal di bawah lapisan epidermis bagian tanaman. Umumnya telur diletakkan pada daun atau tangkai daun, kadang-kadang pada daun muda, namun juga sering ditemukan pada daun yang sudah tua (Subagyo et al. 1977; Center 1994; Julien et al. 1999). Suhu optimal untuk aktivitas makan dan peletakan telur N. eichhorniae sekitar 30 oC (Center 1994). Lama stadium telur kumbang ini bervariasi tergantung pada kondisi pemeliharaan, terutama dipengaruhi oleh suhu lingkungan. Di Florida, lama stadium telur 7-17 hari (Center 1994), sedangkan di Bogor 13-15 hari (Subagyo at al. 1977; Kasno & Mangoendihardjo 1978).
Telur Pupa
Larva ♀ ♂
I mago
Kokon
Gambar 2.3 Siklus hidup N. eichhorniae
stadium larva adalah 36-90 hari (Center 1994; Center et al. 2002), sedangkan di Bogor lama stadium larva sekitar 40 hari (Kasno & Mangoendihardjo 1978). Larva makan dan berkembang di dalam jaringan tanaman, terutama pada daun, tangkai daun, dan batang (Julien et al. 1999). Center (1994) melaporkan bahwa larva kumbang tersebut makan dan berkembang pada tangkai daun. Larva menggerek menuju bagian bawah dan biasanya mencapai dasar tangkai daun sebelum instar tiga.
Pembentukan pupa terjadi pada perakaran eceng gondok. Larva yang telah berkembang sempurna bergerak ke dalam air menuju bagian atas zona perakaran, membuat kokon dari rajutan rambut-rambut akar. Stadium pupa berkisar antara 7 sampai 10 hari pada suhu 27 oC (Center 1994). Sementara itu, di Indonesia stadium pupa sekitar 20 hari (Kasno & Mangoendihardjo 1978), sedangkan di Afrika Selatan dilaporkan bisa mencapai beberapa bulan (Center et al. 2002).
Gambar 2.4 Gejala serangan imago N. eichhorniae
Gambar 2.5 Gejala serangan larva N. eichhorniae
Kisaran Inang Neochetina spp.
Hasil uji kekhususan inang pada 274 jenis tumbuhan dari 77 famili yang mewakili tumbuhan air, tumbuhan bernilai ekonomi, tumbuhan asing dan lokal di Florida, N. eichhorniae hanya menimbulkan satu atau beberapa gejala serangan pada 25 jenis tumbuhan uji. Gejala makan hanya ditemukan pada tumbuhan yang lebih berhubungan secara taksonomi dengan tumbuhan eceng gondok. Gejala serangan yang disebabkan oleh kumbang sangat sedikit dan tidak menyebabkan kerusakan serius pada tumbuhan uji (Julien et al. 1999).
Dari hasil penelitian juga dilaporkan bahwa betina N. eichhorniae hanya dapat meletakkan telur pada 7 spesies tumbuhan uji yang termasuk ke dalam famili Pontederiaceae atau Commelinaceae, tetapi beberapa telur yang diletakkan tidak dapat menetas (infertile), dan bila telur dapat menetas, larvanya segera mati. Larva yang dapat masuk ke dalam batang tumbuhan uji tidak dapat makan dan kemudian mati. Larva hanya dapat berkembang pada tumbuhan
Pontederia cordata L. (Pontederiaceae), namun tidak dapat menyelesaikan siklus hidupnya (Julien et al. 1999). Penelitian lain menunjukkan bahwa N. eichhorniae juga dapat hidup selama periode tertentu pada beberapa tumbuhan lain selain eceng gondok seperti Canna edulis, namun demikian agens hayati ini tidak dapat menyelesaikan siklus hidupnya (Widayanti et al. 1998).
Dampak Nontarget Pengendalian Hayati Gulma
lokal, bahkan tidak mustahil akan terjadi kepunahan spesies lokal. Simberloff dan Stiling (1996) melaporkan bahwa herbivor generalis yang diintroduksi untuk pengendalian hayati gulma dapat menyerang spesies tumbuhan lokal non-target sehingga mengurangi vegetasi spesies tersebut secara nyata dan mengubah komposisi komunitas tumbuhan dan herbivor lokal. Sebagai salah satu contoh, introduksi ulat ngengat Cactoblastis cactorum ke Florida Keys untuk agens pengendalian hayati pada Opuntia spp. telah mendorong O. Spinosissima dan O. triacantha menjadi langka dan terancam punah.
Hasil penelitian dewasa ini juga menunjukkan bahwa agens pengendali hayati yang inangnya spesifik menimbulkan efek nontarget yang besar melalui interaksi tidak langsung dan subsidi jaring-jaring makanan. Efek tidak langsung dapat terjadi melalui penggantian ekologi ketika agens pengendali hayati secara fisik dan fungsional mengganti spesies lokal (Pearson & Callaway, 2003). Lebih jauh Lynch et al. (2002) menyatakan bahwa meskipun agens pengendalian hayati memiliki tingkat penerimaan atau prefensi yang sangat rendah terhadap spesies nontarget, namun kemungkinan memiliki dampak yang besar terhadap populasi nontarget.
Sebagaimana dikemukakan sebelumnya bahwa seyogyanya dalam mengintroduksi agens pengendali hayati perlu dipertimbangkan pengaruhnya terhadap keanekaragaman hayati lokal. Apa yang akan terjadi apabila serangga herbivor tersebut menyerang tumbuhan lokal dan mempengaruhi eksistensi keanekaragaman serangga lokal akibat terjadinya kompetisi interspesifik? Pertanyaan ini sangat penting karena jika spesies-spesies herbivor yang diintroduksi ini lebih dominan akan dapat mengalahkan serangga herbivora lokal sehingga menimbulkan dampak negatif terhadap keanekaragaman spesies lokal, dan bahkan sangat mungkin terjadi kepunahan.
hayati meningkat secara cepat sebagai respon terhadap tingginya kepadatan spesies target berakibat tingginya kepadatan populasi agens tersebut. Kondisi ini dapat mempengaruhi preferensinya terhadap spesies non-target, yang sebelumnya rendah, dan mengakibatkan penurunan populasi spesies non-target secara tajam (Lynch et al. 2002).
Masalah lain yang perlu mendapat perhatian adalah fakta bahwa kebanyakan introduksi serangga herbivor untuk pengendalian gulma tidak diikuti dengan upaya monitoring yang serius terhadap distribusinya, serta eksistensi musuh alami dan serangga lain yang ada di lapangan. Tanpa adanya upaya monitoring, degradasi keanekaragaman hayati dalam suatu ekosistem tidak dapat dipantau, terutama tempat-tempat atau habitat yang jauh dari titik pelepasan (Simberloff dan Stiling, 1996). Lebih jauh mereka menyatakan, pernyataan bahwa introduksi agens pengendalian hayati pada berbagai proyek sebagai bukti bahwa secara umum pengendalian hayati adalah aman tidak cukup kuat. Hal ini didasarkan pada dua alasan: 1) dari beberapa kerusakan yang terjadi, yang terdeteksi hanya merupakan bagian kecil dari yang sebenarnya terjadi, sebab sedikit saja taksa yang cukup menarik perhatian manusia untuk diamati lebih jauh, bahkan oleh para ilmuan, dan 2) tidak seperti kepunahan yang dapat diamati, kerusakan berbagai proses komunitas atau ekosistem nampaknya kurang cukup diamati, bahkan pada proyek penelitian intensif hanya memonitor sebagian kecil karakter dan proses ekosistem.