Studi Peran Interaksi Bakteri Patogen Dan Lingkungan Terhadap Penyakit Ice-Ice Pada Rumput Laut Kappaphycus Alvarezii.
Teks penuh
Gambar
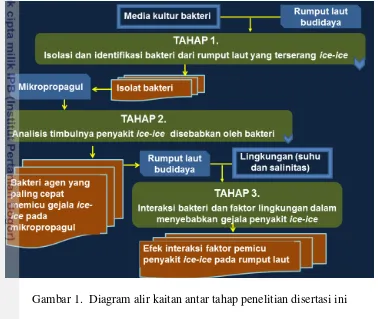
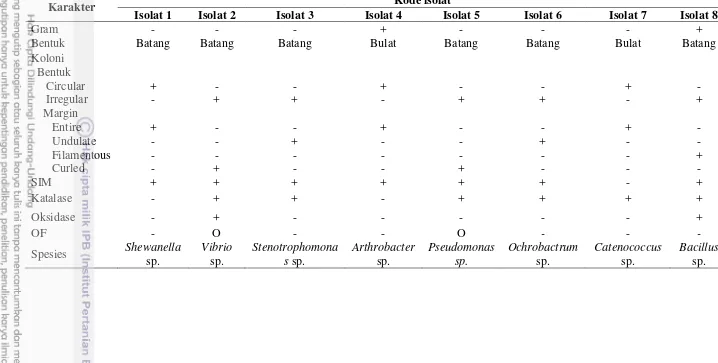
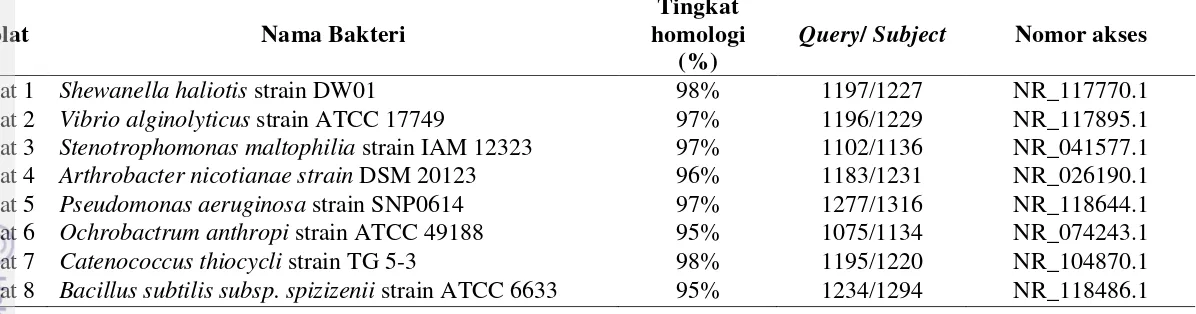
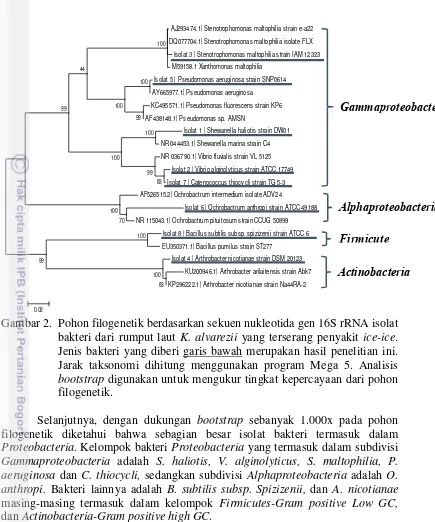
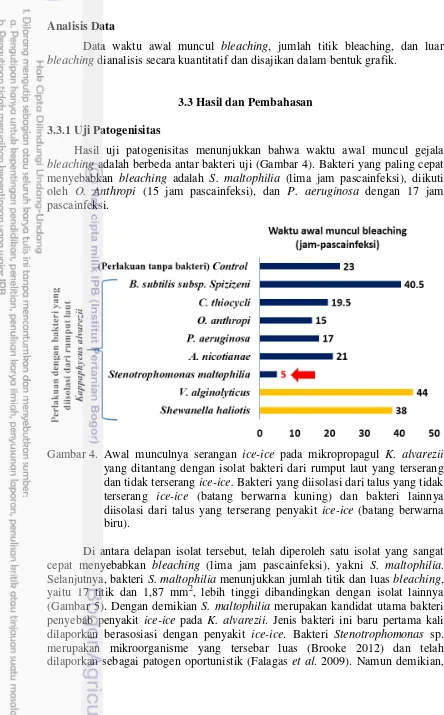
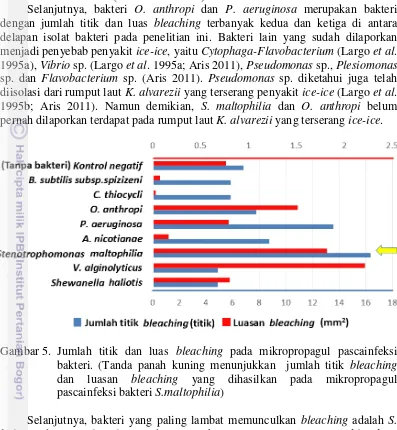
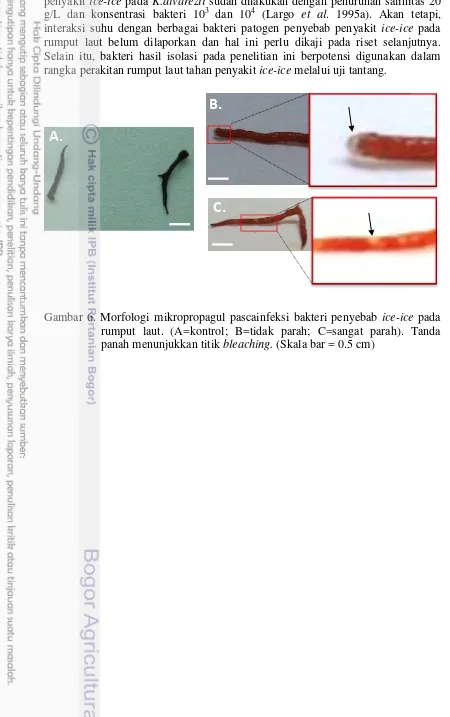
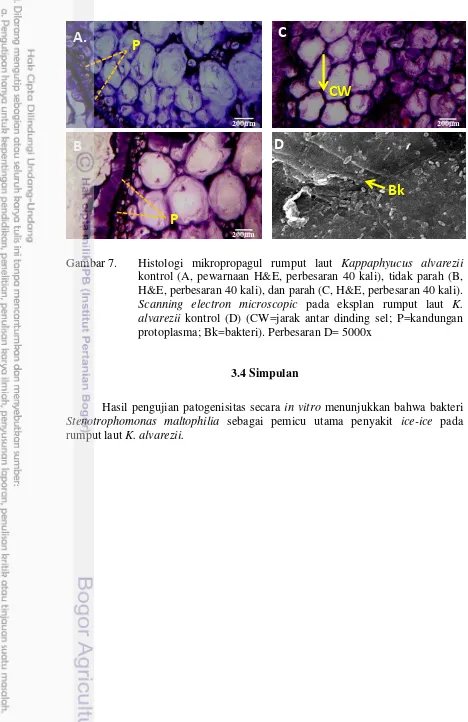
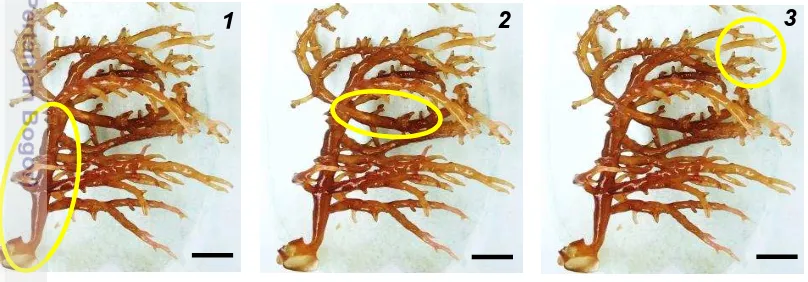

Dokumen terkait
Sumber data dalam penelitian adalah subjek dari mana data dapat diperoleh, apabila peneliti menggunakan kuesioner atau wawancara dalam pengumpulan datanya, maka
Standar tersebut dibagi lagi menjadi lebih spesifik, seperti misalnya standar 802.1 yang memberikan pengenalan bagi sekumpulan standar dan mendefenisikan primitif
Berdasarkan hasil kegiatan pengabdian masyarakat tersebut, dapat disimpulkan bahwa partisipasi masyarakat di daerah Bale-Bale khususnya remaja yang mempunyai jiwa
Seluruh data dan informasi yang tercantum dalam dokumen ini sesuai dengan LHKPN yang diisi dan dikirimkan sendiri oleh Penyelenggara Negara melalui elhkpn.kpk.go.id , serta
Proses Dehumidifikasi, yang merupakan proses pengurangan kadar air dalam gas, sama dengan proses humidifikasi mempunyai dua cara proses, yaitu dengan pemanasan dan tanpa
Berdasarkan hasil dan pembahasan, maka dapat ditarik kesimpulan sebagai berikut : (1) potensi pengembangan sistem minapadi di Provinsi Jambi masih sangat besar,
Karena itulah maka perlunya untuk mempertahankan keberadaan IK.M tersebut, dan salah satu cara dalam mempertahankan keberadaan suatu industri adalah dengan
