LAMPIRAN
Lampiran 1. Data bobot lahir kelahiran pertama
Lampiran 3. Data bobot lahir kelahiran kedua
Lampiran 4. Data bobot sapih kelahiran kedua
Lampiran 6. Data Tetua Kelinci Lokal
Lampiran 7. Analisis Ragam Heritabilitas Bobot Lahir
Source df ss ms
Between sires 3 820,65 273,55
Progeny Within Sires 109 10015,10 91,88
Hertitabilitas (H2) = 0,26
Lampiran 8. Analisis Ragam Heritabilitas Bobot Sapih
Source df ss ms
Between sires 3 262137,78 87379,26
Progeny Within Sires 109 1260088,35 11560,44
Hertitabilitas (H2) = 0,75
Lampiran 9. Analisis Ragam Heritabilitas Litter Size
Source df ss ms
Between sires 3 1,09 0,36
Progeny Within Sires 28 56,88 2,03
Hertitabilitas (H2) = -0,46
Lampiran 10. Analisis Ragam Ripitabilitas Bobot Lahir
Source df ss ms
Correction term 1 255669,25
Individuals 15 2859,24 190,62
Measurement 97 7976,51 82,23
Lampiran 11. Analisis Ragam Ripitabilitas Bobot Sapih
Source Df ss ms
Correction term 1 22744109,9
Individuals 15 695851,27 46390,085
Measurement 97 826374,85 8519,328
Ripitabilitas (r2) = 0,40
Lampiran 12. Analisis Ragam Ripitabilitas Litter Size
Source df ss ms
Correction term 1 399,03
Individuals 15 37,469 2,4979
Measurement 16 20,5 1,2813
Ripitabilitas (r2) = 0,32
Lampiran 13. Efek Heterosis
Jantan Betina Rata-rata BL Induk
Rata-rata
BL Anak % heterosis
NZ2J L2B 57 58.17 2.05
-4.09
NZ2J L5B 53 44.86 -15.36
NZ2J L6B 55.5 53.89 -2.90
NZ2J L8B 50.5 50.43 -0.14
L2J NZ3B 54 47.83 -11.42
-13.36
L2J NZ4B 49.5 49.00 -1.01
L2J NZ5B 52.5 48.80 -7.05
Lampiran 14. Format Data Pengamatan Jumlah Anak Kelinci Jantan dan Betina
No Perkawinan Jumlah
Lampiran 15. Format Data Pengamatan Bobot Lahir Kelinci
Perkawinan Jumlah Anak
Bobot Lahir (gr) Total Bobot Lahir
Rataan Bobot Lahir 1 2 3 4 5 6 7 8
Note :
1
= Tanda di Kepala
2
= Tanda di Kuping Kanan
3
= Tanda di Kuping Kiri
4
= Tanda di Badan
5
= Tanda di Punggung
6
= Tanda di Kaki Belakang Kanan
7
= Tanda di Kaki Depan Kiri
Lampiran 16. Format Data Bobot Sapih Kelinci
Perkawinan Jumlah Anak
Bobot Sapih (gr) Total Bobot Sapih
Rataan Bobot Sapih 1 2 3 4 5 6 7 8
Note :
1
= Tanda di Kepala
2
= Tanda di Kuping Kanan
3
= Tanda di Kuping Kiri
4
= Tanda di Badan
5
= Tanda di Punggung
6
= Tanda di Kaki Belakang Kanan
7
= Tanda di Kaki Depan Kiri
Lampiran 17. Format Data Pengamatan Perkawinan Kelinci
No Tanggal Kawin I
Perkawinan
Waktu Tanggal Kawin II
Perkawinan
Waktu
Lampiran 18. Foto Penelitian
NZ1J NZ2J
NZ1B NZ2B
NZ5B NZ6B
NZ7B NZ8B
L1J L2J
L3B
L5B
L7B
L6B
DAFTAR PUSTAKA
Adjisoedarmo, S., B. Purnomo, A. Marmono E., S. Haryati, D. Purwantini dan A. Sudewo A.T. 1985. Perormans produksi dan reproduksi kelinci Lokal (Bukan Ras). Pros. Sem, Peternakan dan Forum peternak unggas dan aneka ternak. Pusat Penelitian dan Pengembangan Peternakan, Bogor.
Becker. 1984. Manual of Quantitative Genetics. Washington, Academic Enterprises. Pullman.
Bourdon, R. M.1997. Understanding Animal Breeding. Prentice Hall, Inc, New Jersey.
Brahmantiyo, B. 2008. Kajian Potensi Genetik Ternak Kelinci (Orytolagus Cunniculus) di Bogor, Jawa Barat dan Magelang, Jawa Tengah. IPB Press, Bogor.
Brahmantiyo, B. dan Y. C. Raharjo. 2005. Pengembangan Pembibitan Kelinci di Pedesaan dalam menunjang Potensi dan Prospek Agribisnis Kelinci. Pusat Penelitian dan Pengembangan Peternakan, Bogor.
Brahmantiyo, B. dan Y. C. Raharjo. 2011. Peningkatan Produktivitas Kelinci Rex, satin dan Persilanggannya Melalui Seleksi. Pusat Penelitian dan Pengembangan Peternakan, Bogor.
Brahmantiyo, B., Y. C. Raharjo dan L. H. Prasetyo. 2013. Peningkatan Bobot Badan Umur 10 Minggu Kelinci Pedaging FZ-3 Melalui Seleksi. Pusat Penelitian dan Pengembangan Peternakan, Bogor.
Brahmantiyo, B., Y. C. Raharjo, N. D. Savitri dan M. Duldjaman. 2009. Karakteristik Reproduksi Kelinci Rex, Satin dan Reza. Pusat Penelitian dan Pengembangan Peternakan, Bogor.
Cassady, P. J., L. D. Yung dan K. A. Leymaster. 2002. Heterosis and Rekombinant Effects on Pig Reproductive Traits. J. Anim. Sci. 20(9): 2303-2315.
Cheeke P. R., N. M. Paton., S. D. Lukefahr & J. L. McNitt. 1987. Rabbit Production. 6th Edition. The Interstate Printers and Publisher Inc. Danvile,
Illinois. USA.
Egena, S. S. A., G. N. Akpa, I. C. Alemede and A. Aremu. 2012. Genetic and Non-Genetic Factors Affecting Litter Sizeand Birth Weight of Rabbit in Minna, Niger State, Nigeria. J. Animal Production, 14(3): 160-166.
Falconer, D. S. dan T. F. C. Mackay. 1989. Introduction to Quantitative Genetics. Longman Inc, New York.
Falconer, R. D. and F. C. M. Trudy. 1996. Introduction to Quantitative Genetics Longman, Malaysia.
Farrel, D. J. & Y. C. Raharjo. 1984. Potensi Ternak Kelinci Sebagai Penghasil Daging. Pusat Penelitian dan Pengembangan Peternakan, Bogor.
Hamdan. 2010. Pendugaan Komponen Ragam dan Parameter Genetik Beberapa Sifat Produksi Puyuh pada Seleksi Jangka Panjang. USU Press, Medan. Hardjosubroto, W. 1994. Aplikasi PemuliabiakanTernak di Lapangan. PT.
Gramedia Widiasarana Indonesia, Jakarta.
Harper, M.J.K., 1963. Ovulation in the Rabbit. The Time of Follicular Rupture
and Expulsion of the Egg in Relating to Injection of Luteinizing Hormone.
Journal of Endocrinology 26: 307-316.
Harris I. 1994. The Laboratory Rabbit. Anzccart Facts Sheet. Anzccart News. Vol 7(4);1-8.
http://www.accessadelaide.com/ANZCCART/Publications/FS_Rabbit.Pdf. (6 Juni 2006).
Herman, R. 2000. Produksi Kelinci dan Marmut, Anatomi dan Fisiologi Alat Pencernaan serta Kebutuhan. Edisi Ketiga. Fakultas Peternakan. Institut Pertanian Bogor, Bogor.
Kadarwati. 2006. Pengaruh Akar Gingseng (Wild gingseng) dalam Ransum
Mencit (Mus Musculus) Terhadap Jumlah Anak dan PertumbuhanAnak dari Lahir Sampai dengan Sapih. Skripsi. IPB Press, Bogor.
Karnaen. 2008. Estimation of Genetic Parameters, Genetic and Phenotypic Correlation on Madura Cattle. Jatinangor.
Kartadisastra. 1994. Kelinci Unggul. Kanisius, Yogyakarta.
Khalil, M. H., E. A. Afifi, Y. M. K. Youssef dan A. F. Khadir. 1995. Heterosis, Maternal and Direct Genetic Effects for Litter Performance and Reproductive Intervals in Rabbit Crosses. Ministry of Agriculture, Egypt. Khalil, M. H., J. B. Owen and E. A. Afifi. 1986. A Revies of Phenotypic and
Kunianto, E. 2009. Pemuliaan Ternak. Graha Ilmu, Yogyakarta.
Larzul, C., G. Florence, C. Sylvie and Rochambeau H. D. 2004. Divergent Selection on 63-day Body Weight in the Rabbit: Response on Growth, Carcass and Muscle Traits.Genet. Sel. Evol. 37(2005)105-122.
Lasley, J. F. 1978. Genetics of Livestock Improvement. Department of Animal Husbandry. University of Missouri. Prentice-Hall, Inc., New Jersey.
Lebas, F., P. Coudert, R. Rouvier, d Rochambeau H. 1986. The Rabbit. Food and Agriculture Organization of the United Nations, Italy.
Lukefahr, S. D., H. B. Odi, J. K. A. Atakora.1996. Mass Selection for 70-Day Body Weight in Rabbit. J. anim Sci. 74: 1481-1489.
Majalah Flona. 2009. Buku Pintar Memelihara Kelinci & Rodensia. Gramedia, Jakarta.
Martojo H. 1992. Peningkatan Mutu Geneti Ternak. Departemen Pendidikan dan Kebudayaan. Direktorat Pendidikan Perguruan Tinggi. Pusat antar Universitas Bioteknologi. Institut Pertanian Bogor, Bogor.
Negara, N. M. 2014. Seleksi Induk Kambing Peranakan Etawah Berdasarkan Nilai Indeks Produktivitas Induk pada Bobot SApih di Desa Dadapan Kecamatan Sumberrejo Kabupaten Tanggamus. Universitas Lampung, Lampung.
Noor, R. R. 2000. Genetika Ternak. Penebar Swadaya, Jakarta.
Ouyed, A. dan J. M. Brun. 2008. Heterosis, Direct and Maternal Additive Effects on Rabbit Growth and Carcass Characteristics. 9th World Rabbit Congress, Italy.
Prihandini, P.W.,L.Hakim dan V.M.A. Nurgiartiningsih. 2011.Seleksi Pejantan Berdasarkan Nilai Pemuliaan Pada Sapi Peranakan Ongole di Loka Penelitian Sapi Potong Grati-Pasuruan. Jurnal Ternak Tropika Vol 12 No.1 hal 97-107.
Raharjo Y. C., Sumanto L., Sastrodiharjo S., Dharsana R., Pasek I. W. dan Purnama D. 1995 Pola Usaha Pemeliharaan Kelinci Rex dalam Sistem
“PIR” (Bapak Angkat) pada Agroklimat Dataran Tinggi Lahan Kering.
Badan Penelitian dan Pengembangan Pertanian, hlm. 115-136.
Rastogi, R. K., S.D. Lukehfahr dan F.B. Lauckner. 2000. Maternal Heritabilitiy and Repeatibility for Lietter Traits in Rabbit in a Humid Tropical Environment. Livestock Production Science 67 (2000): 123-128. www.elsevier.com/locate/livprodsci.
Sartika, T. 2005. Strategi Pemuliaan Sebagai Alternatif Peningkatan Produktivitas Kelinci Pedaging. Pusat Penelitian dan Pengembangan Peternakan, Bogor. Sartika, T., Y.C. Rahardjo, A. Habibie, D. Purnama dan I.W.P. Sumadia, 1995.
Kebutuhan Energi dan Protein pada Induk Kelinci Fase Gestasi dan Laktasi. Balai Penelitian Ternak, Ciawi-Bogor.
Sarwono, B. 2007. Kelinci Potong dan Hias. Agromedia Pustaka, Jakarta.
Setyawan, W. 1982. Beberapa Faktor yang Mempengaruhi Bobot Lahir dan Pertumbuhan Periode Menyusu serta Estimasi Nilai Heritabilitas Bobot Lahir dan Bobot Sapih Kelinci (Oryctolagus cuniculus). IPB Press, Bogor. Sundasesan, D. 1975. Livestock Breeding in India. Vikas Publisher House PVT
Ltd. Delhi, India.
Sutiyono, B., Soedarsono, S. Johari dan Y. S. Ondho. 2011. Efek Heterosis Berbagai Penampilan Tiktok Jantan dan Betina. Universitas Diponegoro, Semarang.
Warwick, E. J. dan J. E. Legates. 1979. Breeding and Improvement of Farm Animals. (7th ed.). Mc graw-Hill Book Co, New York.
Warwick, E. J., J. M. Astuti, W. Hardjosubroto. 1990. Pemuliaan Ternak. Universitas Gadjah Mada Press, Yogyakarta.
Warwick. E. J., J. M. Astuti dan W. Hardjosubroto. 1995. Pemuliaan Ternak. Cetakan ke-5. Universitas Gadjah Mada Press, Yogyakarta.
Wikipedia. 2007. New Zealand White Rabbit https://en.wikipedia.org/wiki/ New_Zealand_white_rabbit (26 Desember 2015).
BAHAN DAN METODE PENELITIAN
Tempat dan Waktu Penelitian
Penelitian ini dilaksanakan di peternakan Dusun Kelinci pada bulan Juli 2014 sampai Maret 2015. Lokasi Dusun Kelinci berada di Jl. Stasiun No. 68 Kelurahan Kedai Durian Kecamatan Medan Johor.
Bahan dan Alat Penelitian
Bahan
Materi penelitian ini adalah anak kelinci sebanyak 113 ekor anak yang berasal dari perkawinan 4 ekor jantan dan 16 ekor betina (2 jantan New Zealand White, 8 betina New Zealand White, 2 jantan Lokal dan 8 betina Lokal) 2 kali kelahiran.
Alat
Metode Penelitian
Mekanisme dan Teknis Pemeliharaan Kelinci
Kandang dan peralatan disiapkan sebelum kelinci masuk kedalam kandang agar mencegah dari hama dan bibit penyakit. Ternak kelinci yang akan digunakan sebagai objek penelitian melalui beberapa syarat sebagai berikut : ternak kelinci dalam keadaan sehat, lincah, tidak cacat dilihat dari bentuk kaki yang lurus dan lincah, ekor melengkung ke atas lurus merapat ke bagian luar mengikuti tulang punggung, telinga lurus ke atas dan telinga tidak terasa dingin, mata jernih dan bulu mengkilat. Kelinci diadaptasikan selama dua minggu dengan tujuan menstabilkan kondisi kelinci dengan lingkungan barunya. Setelah kondisi kelinci stabil dimulai program perkawinan dengan format NZWa x NZWb, NZWa x Lokalb, Lokala x NZWb dan Lokala x lokalb. Perkawinan dilakukan pada pagi dan sore hari pukul 06.00 WIB dan 18.00 WIB dengan cara indukan dibawa kedalam kandang pejantan dan dibiarkan hingga terjadi perkawinan, setelah itu dipisahkan.
(σ2s + σ2d + σ2w)
4σ2s
Parameter Penelitian
1. Sifat Kuantitatif
Sifat Kuantitatif dicirikan dengan sesuatu yang dapat diukur atau ditimbang, fenotipe sifat kuantitatif dipengaruhi banyak pasangan gen, penampilan sifat kuantitatif dipengaruhi lingkungan. Sifat kuantitatif yang akan diamati pada penelitian ini adalah litter size, bobot lahir dan bobot sapih anakan kelinci.
Litter size merupakan jumlah anakan kelinci dalam satu kelahiran dari satu
indukan. Bobot lahir kelinci adalah bobot badan seekor kelinci pada umur 1 hari. Bobot ini ditimbang setelah ternak dilahirkan oleh induknya. Bobot sapih kelinci adalah bobot badan seekor kelinci pada umur 6 minggu. Bobot ini ditimbang ketika ternak tersebut dipisahkan dari induknya setelah periode menyusui selesai.
2. Heritabilitas
Pendugaan nilai heritabilitas menggunakan metode rancangan tersarang (nested design) un-balanced dengan metode matematika sebagai berikut:
Yij = μ+αi + βij + εij
Keterangan:
Yij = respon anak ke-k dari induk ke-j dan pejantan ke-i
μ = rataan umum
αi = pengaruh pejantan ke-i
βij = pengaruh betina ke-j yang dikawinkan dengan pejantan ke-i
εij = galat
Nilai pendugaan heritabilitas dihitung dengan rumus sebagai berikut :
σ2 W
σ2
W + σ2E
MSW + MSW
k1 (σ2s + σ2d + σ2w)
4σ2d
h2D =
Keterangan:
h2S = Heritabilitas pejantan h2D = Heritabilitas induk
σ2
s = Ragam antar pejantan
σ2
d = Ragam antar induk
σ2
w = Ragam antar anak dalam pejantan.
3. Ripitabilitas
Sedangkan perhitungan ripitabilitas menggunakan model sebagai berikut (Becker,1984). :
Ykm= μ + αk + ekm
Keterangan:
μ = rataan umum
αk = pengaruh individu ke-k
ekm = pengaruh lingkungan
Tabel 2. Sidik Ragam untuk Menduga Nilai Ripitabilitas Suatu Sifat Berdasarkan Pola Tersarang atau Hieraechial
Sumber Keragaman db JK KT Komponen Ragam
Antara Individu n-1 JKW KTW σ2E+k1σ2W
Galat n(m-1) JKE KTE σ2E
Keterangan: db = derajat bebas; JK = jumlah kuadrat; KT = kuadrat tengah; W = individu; E = catatan per individu; n= jumlah individu; σ2W = komponen ragam antar
individu; σ2E = komponen ragam antar catatan dalam individu
Pendugaan ripitabilitas dihitung dengan rumus (Becker, 1984):
R= dengan σ2E = MSE dan σ2W =
keterangan:
R = ripitabilitas
σ
2
W = ragam sifat yang diamati
σ
2
MPH 100% H (%) =
MSW = kuadrat tengah sifat yang diamati
MSE = kuadrat tengah individu yang diamati
K1 = jumlah pencatatan atau ulangan
4. Efek Heterosis
Rumus pengujian nilai efek heterosis adalah :
x
Keterangan :
F1 = Nilai Hibrida yang dihasilkan MPH = Nilai tengah kedua tetua
Korelasi
Model statistik : Yijk= µ + αi+ βij + eik
Yijk = Nilai pengukuran pada anak ke-k dari betina (induk) ke-j yang dikawini
pejantan ke-i µ = Rataan umum
αi = Pengaruh pejantan ke-i
βij = Pengaruh induk ke-j yang kawin dengan pejantan ke-i
eik = Pengaruh lingkungan dan simpangan genetik yang tidak terkontrol
Tabel 3. Sidik Ragam untuk Menduga Nilai Korelasi Suatu Sifat Berdasarkan Pola Tersarang atau Hieraechial
Sumber Keragaman db JK HKR Komponen Ragam
Antar pejantan S-1 JKs HKRS
CovW + k2
CovD + k3
CovS
Antar induk dalam pejantan D-S JKd HKRD
CovW + k1
CovD
Antar anak dalam induk n.. -D JKw HKRW CovW
Koefisien k1=k2 = jumlah anak per induk, k3 = jumlah anak per pejantan
korelasi genetik ( rg) = 4Covs
4σ2
s(x) . 4σ2s(Y)
Analisis Data
HASIL DAN PEMBAHASAN
Bobot Lahir
Bobot lahir adalah bobot saat dilahirkan atau bobot hasil penimbangan dalam kurun waktu 24 jam setelah dilahirkan (Hardjosubroto, 1994).
Tabel 4. Rataan Bobot Lahir Kelinci
Bobot Lahir N x ± sb
NZa x NZb 26 45,85 ± 8,91
La x Lb 30 47,70 ± 8,01
NZa x Lb 29 51,76 ±10,13
La x NZb 28 44,68 ±11,05
NZ = New Zealand White, L = Lokal, a = Jantan, b = Betina
Rataan bobot lahir kelinci NZa x NZb, La x Lb, NZa x Lb dan La x NZb yang didapat pada penelitian ini berturut-turut sebesar 45,85±8,91; 47,70±8,01; 51,76±10,13 dan 44,68±11,05. Hasil ini lebih rendah dibandingkan dengan hasil penelitian Khalil et al. (1995), rataan bobot lahir kelinci NZ(buck) x NZ(doe), BR(buck) x BR(doe), NZ(buck) x BR(doe) dan BR(buck) x NZ(v) berturut sebesar 52±1,5; 49±1,9; 53±1,9 dan 53±2,1. Lebih tinggi dibandingkan hasil yang dilaporkan oleh Egena et al. (2012), pada kelinci CH(buck) x CH(dam), NZ(doe) x NZ(doe), CH(buck) x NZ(doe) dan NZ(buck) x CH(doe) berturut-turut sebesar 41,18±3,39; 40,65±5,30; 39,07±7,09 dan 39,61±3,45. Brahmantiyo dkk. (2009), mengemukakan bahwa jumlah anak yang tinggi menghasilkan bobot individu lebih rendah. Akan tetapi hasil pada penelitian ini berbanding terbalik dengan pendapat tersebut. Indikasi yang menyebabkan perbedaan tersebut adalah
maternal (induk) disetiap ternak yang berbeda. Hal ini sesuai dengan peryataan
Bobot Sapih
Bobot sapih adalah bobot pada saat dipisahkan dari induknya. Bobot sapih merupakan indikator dari kemampuan induk untuk menghasilkan susu dan kemampuan anak untuk mendapatkan air susu dan tumbuh (Hardjosubroto, 1994).
Tabel 5. Rataan Bobot Sapih Kelinci
Bobot Sapih N x ± sb
NZa x NZb 26 473,15 ± 88,72
391,63 ± 71,54 514,17 ± 102,81 419,07 ± 151,41
La x Lb 30
NZa x Lb 29
La x NZb 28
NZ = New Zealand White, L = Lokal, a = Jantan, b = Betina
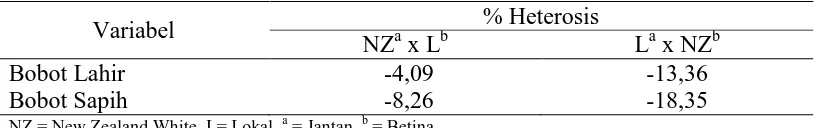
Efek Heterosis
Heterosis adalah rataan keunggulan keturunan dibandingkan dengan rataan kedua kelompok tetuanya sebagai akibat dari perbedaan frekuensi gen diantara tetuanya dan adanya efek dominan dan atau epistasis (Falconer dan Mackay, 1996). Menurut Xu dan Zhu (1999) dan Sutiyono dkk (2011), menyatakan heterosis terjadi akibat adanya interaksi dari pertemuan diantara gen-gen aditif maupun perpaduan aktivitas gen pada suatu lingkungan yang mendukung atau lingkungan yang cocok.
Tabel 6. Efek Heterosis
Variabel % Heterosis
NZa x Lb La x NZb
Bobot Lahir -4,09 -13,36
Bobot Sapih -8,26 -18,35
NZ = New Zealand White, L= Lokal, a = Jantan, b = Betina
Estimasi Nilai Heritabilitas
Heritabilitas adalah istilah yang digunakan untuk menunjukkan bagian dari keragaman total suatu sifat yang diakibatkan oleh pengaruh genetik. Pengetahuan tentang besarnya heritabilitas penting dalam mengembangkan seleksi dan rencana perkawinan untuk memperbaiki ternak (Warwick et al., 1990). Heritabilitas merupakan rasio yang menunjukkan persentase keunggulan tetua yang rata-rata diwariskan kepada anaknya (Warwick et al., 1995).
Tabel 7. Nilai Dugaan Heritabilitas Bobot Lahir, Bobot Sapih dan Litter Size Sifat Produksi Jumlah Pejantan
(ekor)
Jumlah Anak (ekor)
Nilai Heritabilitas (h2)
Bobot Lahir 4 113 0,26
Bobot Sapih 4 113 0,75
Litter Size 4 113 -0,46
Hasil analisis statistik menunjukkan nilai heritabilitas kelinci sifat bobot lahir sebesar 0,26 (tabel 7). Hal ini lebih tinggi dibandingkan dengan hasil penelitian Khalil et al. (1986) dan Brahmantiyo dan Raharjo (2011), yang melaporkan dugaan heritabilitas bobot lahir sebesar 0,124 pada kelinci New Zealand White, 0,12 pada kelinci Bouscat dan 0,40 pada kelinci Giza White. Adanya perbedaan ini sesuai dengan pendapat Khalil et al. (1986) dan Brahmantiyo dan Raharjo (2011), bahwa adanya perbedaan nilai dugaan heritabilitas disebabkan oleh (a) metode analisis yang digunakan untuk menduga; (b) ekspresi genetik setiap bangsa di dalam populasi yang berbeda; (c) jumlah data yang digunakan; dan (d) faktor koreksi untuk sifat non-genetik yang dibuat pada setiap data.
dan Raharjo (2011), yang melaporkan dugaan nilai heritabilitas bobot sapih kelinci RR, SS dan RS berturut turut sebesar 0,93 ; 0,82 dan 0,98. Dugaan heritabilitas penelitian ini lebih tinggi dibandingkan dugaan heritabilitas bobot sapih kelinci FZ-3 (Flemish Giant 37,5% dan Reza 62,5%) sebesar 0,36 (Lukefahr, 1996; Brahmantiyo dkk, 2013). Khalil et al. (1986) dan Brahmantiyo dan Raharjo (2011), melaporkan nilai dugaan heritabilitas bobot sapih umur 42 hari kelinci New Zealand White sebesar 0,14; kelinci Bouscat sebesar 0,29 dan kelinci Giza White sebesar 0,62. Nilai heritabilitas suatu sifat akan bervariasi antar populasi dan bangsa. Perbedaan variasi tersebut dapat disebabkan oleh perbedaan faktor genetik (ragam genetik), perbedaan lingkungan (ragam lingkungan), metode dan jumlah cuplikan data yang digunakan (Falconer dan Mackay, 1989; Hamdan, 2010). Warwick et al. (1990), menyatakan bahwa diperlukan jumlah sampel minimal 500 sampel agar nilai heritabilitas yang diperoleh handal.
Nilai heritabilitas litter size pada penelitian ini sebesar -0,46 (Tabel 7). Nilai heritabilitas litter size mempunyai nilai minus meskipun secara teoritis nilai heritabilitas tidak lebih dari satu. Nilai heritabilitas negatif atau lebih dari satu secara biologis tidak mungkin. Hal tersebut dimungkinkan disebabkan oleh keseragaman yang disebabkan oleh lingkungan yang berbeda untuk keluarga kelompok yang berbeda, metode yang digunakan tidak tepat sehingga tidak dapat menunjukkan antara ragam genetik dan ragam lingkungan dengan selektif, kesalahan dalam pengambilan sampel (warwick et al., 1995).
Ripitabilitas merupakan dari ragam total suatu populasi yang disebabkan oleh perbedaan-perbedaan antara individu yang bersifat permanen (warwick et al., 1990). Menurut Sundasesan (1975), jika nilai ripitabilitasnya tinggi, maka ternak dapat dipertahankan atau dikeluarkan dari peternakan berdasarkan catatan pertama pengamatan, namun jika ripitabilitasnya rendah diperlukan pengamatan lebih dari satu kali pada karakter yang sama sebelum menentukan kehidupan produksi ternak yang bersangkutan.
Tabel 8. Nilai Dugaan Ripitabilitas Sifat Bobot Lahir, Bobot Sapih dan Litter Size Sifat Produksi Jumlah Betina
(ekor)
Jumlah Anak (ekor)
Nilai Ripitabilitas (r2)
Bobot Lahir 16 113 0,17
Bobot Sapih 16 113 0,4
Litter Size 16 113 0,3
Korelasi Genetik
[image:31.595.113.530.394.424.2]Terdapat 3 pengertian korelasi, yaitu korelasi fenotipik (rp), korelasi genotipik (rg) dan korelasi lingkungan (re), maka pembahasan yang paling penting adalah korelasi genetik (Kurnianto, 2009). Karnaen (2008) menyatakan bahwa korelasi genetik adalah korelasi dari pengaruh aditif atau nilai pemuliaan sifat kuantitatif, sedangkan korelasi fenotipik merupakan korelasi total dari semua sifat yang dimiliki ternak. Faktor yang diwariskan tetua kepada keturunannya adalah faktor genetik, bukan faktor lingkungan. Korelasi genetik sangat penting jika dua sifat berkorelasi secara genetik, maka seleksi untuk suatu sifat akan menyebabkan perubahan pada sifat lain (Kurnianto, 2009).
Tabel 9. Korelasi Sifat Bobot Lahir Terhadap Bobot Sapih pada Kelinci
Sifat N Nilai Korelasi Genetik
Bobot Lahir – Bobot Sapih 113 0,52
KESIMPULAN DAN SARAN
Kesimpulan
Efek heterosis minus menunjukkan bahwa performans bobot badan anak tidak lebih baik dari rata-rata tetuanya. Nilai heritabilitas digolongkan kedalam kategori rendah untuk sifat bobot lahir, kategori tinggi untuk sifat bobot sapih dan minus pada litter size, atas hasil tersebut maka sifat bobot sapih dapat digunakan sebagai kriteria dalam penyusunan program seleksi. Nilai ripitabilitas sifat bobot lahir termasuk dalam kategori rendah, sifat bobot sapih dikategorikan tinggi dan
litter size termasuk kategori sedang. Korelasi genetik termasuk kedalam kategori
sedang menunjukkan antar sifat cukup efektif dalam hubungan keterkaitannya.
Saran
TINJAUAN PUSTAKA
Klasifikasi Ternak Kelinci
Kingdom: Animalia, Famili: Leporidae, Subfamili: Leporine, Ordo:
Lagomorpha, Genus: 1.Lepus (22 species)=Genuine Hare, 2.Orictolagus (1
species)=O. Cuniculus/European Rabbit, 3.Sylvilagus (22 species)=Cotton Tail
Rabbit, 4.Pronolagus (22 species)= Red Hare, 5.Bunolagus (1 species)=Bushman
Hare, 6.Pentalagus (1 species)=Riu-Kiu Rabbit, 7.Caprolagus (1 species)=Bristle
Rabbit, 8.Poelagus (1 species)=African Rabbit, 9.Nesolagus (1 species)=Sumatera
Rabbit, 10.Romerolagus (1 species)=Volcano Rabbit dan 11.Brachylagus
(1 species)=Dwarf Rabbit (Kartadisastra, 2012).
Biologis Kelinci
rata-rata pada umur 6-8 minggu, serta (6) segera setelah melahirkan, induk kelinci dapat dikawinkan.
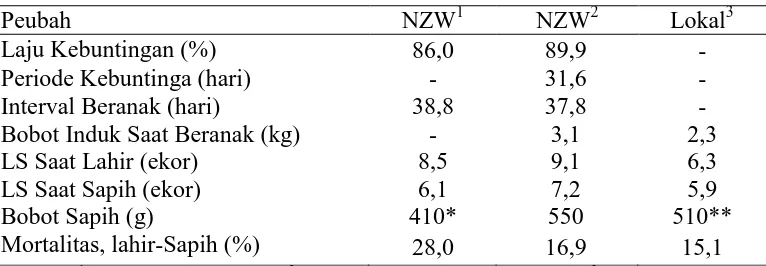
Tabel 1. Performans Produksi Kelinci New Zealand White dan Lokal (Raharjo,1994)
Peubah NZW1 NZW2 Lokal3
Laju Kebuntingan (%) 86,0 89,9 -
Periode Kebuntinga (hari) - 31,6 -
Interval Beranak (hari) 38,8 37,8 -
Bobot Induk Saat Beranak (kg) - 3,1 2,3
LS Saat Lahir (ekor) 8,5 9,1 6,3
LS Saat Sapih (ekor) 6,1 7,2 5,9
Bobot Sapih (g) 410* 550 510**
Mortalitas, lahir-Sapih (%) 28,0 16,9 15,1
Sumber : 1Partridge (1988) Inggris; 2Raharjo et al. (1986), Oregon, USA; 3Sartika dan Diwyanto (1986), Bogor, Indonesia
Keterangan : LS = litter size; *sapih umur 25 hari; **sapih umur 35 hari;
Bangsa Kelinci
Dalam The New Rabbit Handbook ditulis, kelinci (Orytolagus cuniculus
forma domestica) dan terwelu (Lepus europaeua) berasal dari Eropa. Kemudian
menyebar ke seluruh dunia, termasuk Asia. Kelinci adalah anggota ordo
logomorpha. Sampai awal abad ke-20, ordo ini dimasukkan superfamili dalam
ordo Rodentia, keluarga pengerat. Karena ada perbedaan nyata antara Rodentia
dan Lagomorpha, mereka dipisah dalam ordo sendiri (Flona, 2009).
New Zealand White
laboratorium (Sarwono, 2007). Lebas et al. (1986), menerangkan bahwa bobot dewasa 4,1-5,0 kg. umur kawin pertama 144 hari, rataan litter size lahir 8.5 ekor,
litter size hidup 8,0 ekor dan litter size sapih 6,5 ekor. Kelinci New Zealand White
terkenal dengan mothering abilitynya yang baik, produksi susunya juga baik (Lukefahr et al., 1983; Sartika, 2005). Serta merupakan ternak prolifik.
Keunggulan lain dari kelinci New Zealand White adalah kelinci yang umum dipergunakan dalam penelitian sebagai hewan percobaan untuk penelitian biomedis (Cheeke et al., 1987; Brahmantiyo, 2008). Menurut Wikipedia (2015), kelinci NZW menampilkan respon yang sama sebagaimana manusia pada penyakit dan pengobatannya. Ditambahkan, reaksi ini menjadikan kelinci NZW selalu dipergunakan di laboratorium pharmasi pada rumah sakit umum di Amerika Serikat, pusat penelitian kanker dan rumah sakit universitas.
Lokal
dan mempunyai toleransi terhadap panas serta berbadan besar (Farrel dan Raharjo, 1984).
Herman (2000), menyatakan bahwa kelinci lokal lebih toleran terhadap panas (suhu tinggi) dibandingkan kelinci impor. Hal ini disebabkan kelinci lokal telah beradaptasi di daerah tropis sehingga lebih tahan terhadap lingkungan panas dibandingkan kelinci impor yang berasal dari daerah iklim sedang. Kelinci lokal diternakkan dengan tujuan sebagai penghasil daging. Daging yang dihasilkan pun mempunyai kualitas yang cukup baik.
Cross Breeding (kawin silang)
Perkawinan silang adalah perkawinan ternak-ternak dari bangsa yang berbeda (Warwick et al., 1990). Teknisnya Crossbreeding ini hanya berlaku untuk persilangan pertama pada breed asli, tetapi secara umum berlaku juga untuk sistem crisscrossing dari dua jenis atau rotasi persilangan dari tiga atau lebih bibit dan untuk menyilangkan pejantan murni dari satu ras untuk menaikkan tingkatan betina dari ras yang yang lain (Warwick dan Legates, 1979).
Sifat Kuantitatif
Adanya variasi bobot badan dari berbagai penelitian bahkan pada lokasi yang sama dapat disebabkan oleh faktor waktu, lokasi dan jumlah ternak yang berbeda (Prihandini et al., 2011). Diterangkan Lebas et al. (1986), bahwa pengaruh lingkungan yang mempengaruhi sifat kuantitatif antara lain iklim, habitat, kelembaban, aliran udara, peralatan pemeliharaan, teknik pemuliabiakan, pemberian pakan dan faktor manusia (peternak).
Bobot Lahir
Bobot lahir adalah bobot saat dilahirkan atau bobot hasil penimbangan dalam kurun waktu 24 jam setelah dilahirkan (Hardjosubroto, 1994). Khalil et al. (1995), menyatakan bahwa rataan bobot lahir kelinci NZ(buck) x NZ(doe), BR(buck) x BR(doe), NZ(buck) x BR(doe) dan BR(buck) x NZ(v) berturut sebesar 52±1,5; 49±1,9; 53±1,9 dan 53±2,1. Lebih tinggi dibandingkan hasil yang dilaporkan oleh Egena et al. (2012), pada kelinci CH(buck) x CH(dam), NZ(doe) x NZ(doe), CH(buck) x NZ(doe) dan NZ(buck) x CH(doe) berturut-turut sebesar 41,18±3,39; 40,65±5,30; 39,07±7,09 dan 39,61±3,45.
Bobot Sapih
Bobot sapih adalah bobot pada saat dipisahkan dari induknya. Bobot sapih merupakan indikator dari kemampuan induk untuk menghasilkan susu dan kemampuan anak untuk mendapatkan air susu dan tumbuh (Hardjosubroto, 1994). Khalil et al. (1995), rataan bobot sapih kelinci NZ(buck) x NZ(doe), BR(buck) x BR(doe), NZ(buck) x BR(doe) dan BR(buck) x NZ(doe) berturut sebesar 404±10; 401±12; 418±13 dan 426±15. Perbedaan ini mungkin disebabkan karena terbatasnya kemampuan induk untuk mencukupi kebutuhan susu yang diperlukan anaknya selama periode menyusu. Besar litter size yang tinggi, sehingga anak tidak dapat menerima susu secara optimal sesuai kebutuhan untuk pertumbuhan normal, sehingga pertumbuhan terganggu (Setyawan, 1982).
Litter Size
Jumlah litter size tergantung pada jumlah sel telur yang diovulasikan dan dibuahi oleh sperma serta tumbuh dan berkembang normal sampai dilahirkan (Sartika dkk., 1995). Harper (1963), menyatakan bahwa sel telur dilepaskan secara bertahap selama ovulasi berlangsung. Semakin banyak jumlah sel telur diovulasikan, jumlah sel telur yang dibuahi akan semakin banyak sehingga jumlah anak sekelahiran akan meningkat pula.
Kadarwati (2006), menyebutkan bahwa besarnya anak per kelahiran dipengaruhi oleh bangsa ternak, umur induk, musim kelahiran, dan kondisi lingkungan. Faktor-faktor lingkungan sangat mempengaruhi jumlah kelahiran antara lain musim kawin, jumlah sel telur yang dihasilkan serta tingkat kematian embrio yang sangat berpengaruh terhadap jumlah anak per kelahiran.
Heterosis
Hetero-genetik adalah pertemuan antara berbagai gen yang mengontrol bermacam-macam sifat dalam menumbuhkan karakter, baik karakter kualitatif maupun kuantitatif. Menurut Xu dan Zhu (1999) dan Sutiyono dkk (2011), menyatakan heterosis terjadi akibat adanya interaksi dari pertemuan diantara gen-gen aditif maupun perpaduan aktifitas gen-gen pada suatu lingkungan yang mendukung atau lingkungan yang cocok.
heterosigot lebih unggul daripada individu homosigot. Kemudian teori epistasis menyebutkan bahwa heterosis merupakan perwujudan dari segala bentuk interaksi antar lokus. Peningkatan performa pertumbuhan pada hasil persilangan berkisar 0- 10% dan untuk sifat-sifat fertilitas berkisar antara 5-25% (Noor, 2000).
Hasil dari penelitian Khalil et al. (1995), heterosis bobot lahir dan sapih berturut-turut sebesar 2,5 dan 19,1 pada kelinci persilangan New Zealand White x Baladi Red. Persilangan kelinci New Zealand White x Californian menghasilkan efek heterosis berat hidup 35 hari dan 63 hari sebesar 36 dan 60 (Ouyed dan Brun, 2008). Efek heterosis suatu karakter dapat juga negatif, yang disebabkan oleh perpaduan gen yang menimbulkan suatu sifat menjadi lebih rendah dari rata-rata penampilan kedua tetuanya. Efek heterosis yang negatif pada persilangan antar spesies lebih tampak pada karakter reproduksi (Cassady et al., 2002; Sutiyono dkk, 2011).
Parameter Genetik
Keragaman dan mutu genetik sifat-sifat yang merupakan potensi genetik individu-individu dalam suatu populasi akan tercermin pada nilai parameter genetiknya meliputi nilai heritabilitas, ripitabilitas dan korelasi genetik (Hardjosubroto, 1994).
Heritabilitas
merupakan rasio yang menunjukkan persentase keunggulan tetua yang rata-rata diwariskan kepada anaknya (Warwick et al., 1995).
Noor (2000) menyatakan bahwa, ada dua macam heritabilitas, yaitu heritabilitas dalam arti luas yang merupakan rasio antar keragaman genetik dengan keragaman fenotipik yang melibatkan pengaruh gen aditif dan non aditif. Heritabilitas dalam arti sempit merupakan rasio antara keragaman aditif dan keragaman fenotip yang dalam hal ini aksi gen non aditif (dominan dan epistasis). Secara teori, nilai heritabilitas dapat berkisar antara 0 hingga 1, tetapi angka ekstrim ini jarang diperoleh untuk sifat-sifat kuantitatif ternak. Heritabilitas yang bernilai nol maka hal tersebut berarti semua keragaman disebabkan oleh pengaruh lingkungan, dan sebaliknya, jika heritabilitas bernilai satu, maka semua keragaman disebabkan oleh keturunan (Warwick et al., 1990).
Pada penelitian Khalil et al. (1986) dan Brahmantiyo dan Raharjo (2011), dugaan heritabilitas bobot lahir sebesar 0,124 pada kelinci New Zealand White, 0,12 pada kelinci Bouscat dan 0,40 pada kelinci Giza White. Adanya perbedaan ini sesuai dengan pendapat Khalil et al. (1986) dan Brahmantiyo dan Raharjo (2011), bahwa adanya perbedaan nilai dugaan heritabilitas disebabkan oleh (a) metode analisis yang digunakan untuk menduga; (b) ekspresi genetik setiap bangsa di dalam populasi yang berbeda; (c) jumlah data yang digunakan; dan (d) faktor koreksi untuk sifat non-genetik yang dibuat pada setiap data.
(Lukefahr, 1996; Brahmantiyo dkk, 2013). Khalil et al. (1986) dan Brahmantiyo dan Raharjo (2011), melaporkan nilai dugaan heritabilitas bobot sapih umur 42 hari kelinci New Zealand White sebesar 0,14; kelinci Bouscat sebesar 0,29 dan kelinci Giza White sebesar 0,62. Nilai heritabilitas suatu sifat akan bervariasi antar populasi dan bangsa. Perbedaan variasi tersebut dapat disebabkan oleh perbedaan faktor genetik (ragam genetik), perbedaan lingkungan (ragam lingkungan), metode dan jumlah cuplikan data yang digunakan (Falconer dan Mackay, 1989; Hamdan, 2010). Warwick et al. (1990), menyatakan bahwa diperlukan jumlah sampel minimal 500 sampel agar nilai heritabilitas yang diperoleh handal.
Martojo (1992), menyatakan bahwa heritabilitas bukan suatu konstanta, karena itu nilainya dipengaruhi oleh besar komponen ragam aditif, ragam genetik, dan lingkungan. Sifat reproduksi dan daya hidup pada umumnya mempunyai nilai heritabilitas yang rendah, sedangkan sifat-sifat pertumbuhan dan perdagingan mempunyai nilai heritabilitas tinggi. Nilai heritabilitas negatif atau lebih dari satu secara biologis tidak mungkin. Hal tersebut dimungkinkan disebabkan oleh keseragaman yang disebabkan oleh lingkungan yang berbeda untuk keluarga kelompok yang berbeda, metode yang digunakan tidak tepat sehingga tidak dapat menunjukkan antara ragam genetik dan ragam lingkungan dengan selektif, kesalahan dalam pengambilan sampel (warwick et al., 1995).
Ripitabilitas
(Warwick et al., 1990). Ragam ripitabilitas disebabkan oleh perubahan fisiologi dan fluktuasi lingkungan yang bersifat sementara. Menurut Sundasesan (1975), jika nilai ripitabilitasnya tinggi, maka ternak dapat dipertahankan atau dikeluarkan dari peternakan berdasarkan catatan pertama pengamatan, namun jika ripitabilitasnya rendah diperlukan pengamatan lebih dari satu kali pada karakter yang sama sebelum menentukan kehidupan produksi ternak yang bersangkutan. (Martojo, 1992).
Korelasi
Terdapat 3 pengertian korelasi, yaitu korelasi fenotipik (rp), korelasi genotipik (rg) dan korelasi lingkungan (re), maka pembahasan yang paling penting adalah korelasi genetik (Kurnianto, 2009). Karnaen (2008) menyatakan bahwa korelasi genetik adalah korelasi dari pengaruh aditif atau nilai pemuliaan sifat kuantitatif, sedangkan korelasi fenotipik merupakan korelasi total dari semua sifat yang dimiliki ternak. Faktor yang diwariskan tetua kepada keturunannya adalah faktor genetik, bukan faktor lingkungan. Korelasi genetik sangat penting jika dua sifat berkorelasi secara genetik, maka seleksi untuk suatu sifat akan menyebabkan perubahan pada sifat lain (Kurnianto, 2009).
PENDAHULUAN
Latar Belakang
Permasalahan kelinci saat ini ditinjau dari segi produksi adalah mutu bibit yang kurang memadai dan berakibat pada menurunnya produktivitas dan mutu produk, mortalitas anak lepas sapih yang tinggi dan harga pakan yang tinggi untuk suatu pemeliharaan intensif (Raharjo et al., 1995). Untuk keberhasilan pengembangan yang lebih baik, perlu pula dilakukan peningkatan mutu genetik ternaknya. Untuk meningkatkan mutu genetik ternak kelinci salah satu cara yang dapat dilakukan yaitu dengan mengadakan seleksi. Akan tetapi pelaksanaan seleksi tidak dapat terlepas dari perhitungan nilai ekonomisnya. Nilai ekonomis dari kegiatan seleksi dipengaruhi oleh berbagai faktor, salah satu faktor yang mempengaruhi yaitu daya pewarisan sifat (heritabilitas) dari sifat yang hendak diseleksi. Pewarisan sifat pada semua ternak dipengaruhi keadaan lingkungan tempat ternak hidup serta faktor genetik ternak yang bersangkutan.
Nilai heritabilitas dapat digunakan sebagai indikator untuk memperkirakan keekonomisan suatu kegiatan seleksi, dengan memperkirakan waktu yang diperlukan untuk mencapai tujuan seleksi yang dikehendaki. Pada suatu sifat dengan nilai heritabilitas tinggi, seleksi akan lebih efektif dibandingkan seleksi untuk sifat dengan nilai heritabilitas yang rendah, dan pada sifat dengan nilai heritabilitas tinggi, seleksi dapat didasarkan atas fenotip individu.
terutama faktor lingkungan yang mungkin akan menghambat pertumbuhan kelinci, seperti angin, suhu, makanan dan kelembaban udara.
Tujuan Penelitian
Tujuan penelitian ini adalah untuk mengetahui litter size, bobot lahir dan bobot sapih dalam 2 (dua) kelahiran, menduga parameter genetik terhadap litter
size, bobot lahir dan bobot sapih serta untuk menduga korelasi antara bobot lahir
dengan bobot sapih dari kelinci New Zealand White, Lokal dan persilangan.
Kegunaan Penelitian
ABSTRAK
AHMAD AWALUDDIN, 2016: Pendugaan Parameter Genetik dan Korelasi Sifat Bobot Lahir, Bobot Sapih dan Litter Size pada Kelinci New Zealand White, Lokal dan Persilangan. Dibimbing oleh HAMDAN dan HASNUDI.
Kelinci New Zealand White merupakan alternatif penghasil daging yang potensial. Informasi genetik persilangan kelinci New Zealand White dengan Lokal menentukan strategi pemuliaan di masa mendatang. Penelitian ini bertujuan mengetahui litter size, bobot lahir dan bobot sapih dalam 2 kelahiran, menduga parameter genetik terhadap litter size, bobot lahir dan bobot sapih serta untuk menduga korelasi antara bobot lahir dengan bobot sapih dari kelinci New Zealand White, Lokal dan persilangan. Penelitian dilakukan di peternakan Dusun Kelinci dari bulan Juli 2014 sampai Maret 2015. Rancangan yang digunakan adalah pola tersarang tidak seimbang. Materi penelitian berupa data produksi sebanyak 113 ekor anak kelinci dari 4 ekor jantan dan 16 betina dua kali kelahiran.
Hasil penelitian menunjukkan nilai heritabilitas bobot lahir, bobot sapih dan litter size masing-masing yaitu 0,26; 0,75 dan -0,46. Nilai ripitabilitas yang didapat masing-masing yaitu 0,17; 0,4 dan 0,3 dengan nilai korelasi genetik bobot lahir-bobot sapih sebesar 0,52. Kesimpulan penelitian adalah efek heterosis minus menunjukkan bahwa performans bobot badan anak tidak lebih baik dari rata-rata tetuanya. Nilai heritabilitas sifat bobot sapih tinggi sehingga dapat digunakan sebagai kriteria dalam penyusunan program seleksi. Nilai ripitabilitas sifat bobot lahir termasuk dalam kategori rendah, sifat bobot sapih dikategorikan tinggi dan litter size termasuk kategori sedang. Namun, korelasi genetik termasuk kedalam kategori sedang menunjukkan antar sifat cukup efektif dalam hubungan keterkaitannya.
ABSTRACT
AHMAD AWALUDDIN, 2016. Estimation of Genetic Parameter and Correlation of Birth Weight, Weaning Weight and Litter Size in New Zealand White, Local and Crossbreed Rabbit. Supervised by HAMDAN and HASNUDI.
New Zealand White rabbit has potential as a producer of meet. Genetic information of New Zealand White crossbreed with local breed is important to determine the effect of litter size, birth weight, weaning weight of 2 natal periode, estimation of genetic prematur litter size, birth weight, weaning weight and correlation of New Zealand White, Local and Crossbred. Has study was conducted at Dusun kelinci farm on July 2014 to March 2015. This research used unbalanced nested design. Data consisted of production record from 113 little rabbit of cross 4 buck and 16 doe.
The result showed heritability of birth weight, weaning weight and litter size were : 0.26; 0.75 and -0.46. Ripitability value were : 0.17; 0.4 dan 0.3 with correlation value of birth weight-weaning weight was other. The conclusion heterosis value minus and this showed birth weight performance louwer than their parents. Heritability value of birth weight higher than all parameter, caused this used as breeding program criteria. Ripitability value of birth weight were low category. Ripitability value of weaning weight were high category. Ripitability value of litter size were middle category. However, genetic correlation was middle category and this showed efectivity of the other.
Keywords : New Zealand White breed, Local breed, heritability, ripitability and
PENDUGAAN PARAMETER GENETIK DAN KORELASI
SIFAT BOBOT LAHIR, BOBOT SAPIH DAN LITTER SIZE
PADA KELINCI NEW ZEALAND WHITE, LOKAL
DAN PERSILANGAN
SKRIPSI
Oleh :
AHMAD AWALUDDIN 100306056
PROGRAM STUDI PETERNAKAN FAKULTAS PERTANIAN UNIVERSITAS SUMATERA UTARA
PENDUGAAN PARAMETER GENETIK DAN KORELASI
SIFAT BOBOT LAHIR, BOBOT SAPIH DAN LITTER SIZE
PADA KELINCI NEW ZEALAND WHITE, LOKAL
DAN PERSILANGAN
SKRIPSI
Oleh :
AHMAD AWALUDDIN 100306056
Skripsi sebagai salah satu syarat untuk memperoleh gelar sarjana di Program Studi Peternakan Fakultas Pertanian Universitas Sumatera Utara
PROGRAM STUDI PETERNAKAN FAKULTAS PERTANIAN UNIVERSITAS SUMATERA UTARA
Judul Penelitian : Pendugaan Parameter Genetik dan Korelasi Sifat Bobot Lahir, Bobot Sapih dan Litter Size pada Kelinci New Zealand White, Lokal dan Persilangan
Nama : Ahmad Awaluddin
NIM : 100306056
Program Studi : Peternakan
Disetujui oleh: Komisi Pembimbing
Hamdan, S.Pt., M.Si. Prof. Dr. Ir. Hasnudi, MS. Ketua Anggota
Mengetahui,
Dr. Ir. Ma’ruf Tafsin, M.Si Ketua Program Studi Peternakan
ABSTRAK
AHMAD AWALUDDIN, 2016: Pendugaan Parameter Genetik dan Korelasi Sifat Bobot Lahir, Bobot Sapih dan Litter Size pada Kelinci New Zealand White, Lokal dan Persilangan. Dibimbing oleh HAMDAN dan HASNUDI.
Kelinci New Zealand White merupakan alternatif penghasil daging yang potensial. Informasi genetik persilangan kelinci New Zealand White dengan Lokal menentukan strategi pemuliaan di masa mendatang. Penelitian ini bertujuan mengetahui litter size, bobot lahir dan bobot sapih dalam 2 kelahiran, menduga parameter genetik terhadap litter size, bobot lahir dan bobot sapih serta untuk menduga korelasi antara bobot lahir dengan bobot sapih dari kelinci New Zealand White, Lokal dan persilangan. Penelitian dilakukan di peternakan Dusun Kelinci dari bulan Juli 2014 sampai Maret 2015. Rancangan yang digunakan adalah pola tersarang tidak seimbang. Materi penelitian berupa data produksi sebanyak 113 ekor anak kelinci dari 4 ekor jantan dan 16 betina dua kali kelahiran.
Hasil penelitian menunjukkan nilai heritabilitas bobot lahir, bobot sapih dan litter size masing-masing yaitu 0,26; 0,75 dan -0,46. Nilai ripitabilitas yang didapat masing-masing yaitu 0,17; 0,4 dan 0,3 dengan nilai korelasi genetik bobot lahir-bobot sapih sebesar 0,52. Kesimpulan penelitian adalah efek heterosis minus menunjukkan bahwa performans bobot badan anak tidak lebih baik dari rata-rata tetuanya. Nilai heritabilitas sifat bobot sapih tinggi sehingga dapat digunakan sebagai kriteria dalam penyusunan program seleksi. Nilai ripitabilitas sifat bobot lahir termasuk dalam kategori rendah, sifat bobot sapih dikategorikan tinggi dan litter size termasuk kategori sedang. Namun, korelasi genetik termasuk kedalam kategori sedang menunjukkan antar sifat cukup efektif dalam hubungan keterkaitannya.
ABSTRACT
AHMAD AWALUDDIN, 2016. Estimation of Genetic Parameter and Correlation of Birth Weight, Weaning Weight and Litter Size in New Zealand White, Local and Crossbreed Rabbit. Supervised by HAMDAN and HASNUDI.
New Zealand White rabbit has potential as a producer of meet. Genetic information of New Zealand White crossbreed with local breed is important to determine the effect of litter size, birth weight, weaning weight of 2 natal periode, estimation of genetic prematur litter size, birth weight, weaning weight and correlation of New Zealand White, Local and Crossbred. Has study was conducted at Dusun kelinci farm on July 2014 to March 2015. This research used unbalanced nested design. Data consisted of production record from 113 little rabbit of cross 4 buck and 16 doe.
The result showed heritability of birth weight, weaning weight and litter size were : 0.26; 0.75 and -0.46. Ripitability value were : 0.17; 0.4 dan 0.3 with correlation value of birth weight-weaning weight was other. The conclusion heterosis value minus and this showed birth weight performance louwer than their parents. Heritability value of birth weight higher than all parameter, caused this used as breeding program criteria. Ripitability value of birth weight were low category. Ripitability value of weaning weight were high category. Ripitability value of litter size were middle category. However, genetic correlation was middle category and this showed efectivity of the other.
Keywords : New Zealand White breed, Local breed, heritability, ripitability and
RIWAYAT HIDUP
Penulis dilahirkan di Medan, Sumatera Utara pada tanggal 23 Januari 1993 dari Ayah Haryono dan Ibu Ridayati. Penulis merupakan anak ketiga dari enam bersaudara.
Penulis menyelesaikan pendidikan taman kanak-kanak di TK Dewantara pada tahun 1998 dan pendidikan dasar di SDN 060928 pada tahun 2004. Pendidikan menengah pertama diselesaikan pada tahun 2007 di SMP Swasta Istiqlal dan dilanjutkan ke pendidikan sekolah menengah atas diselesaikan pada tahun 2010 di SMA Swasta Harapan Mandiri. Penulis diterima sebagai mahasiswa Universitas Sumatera Utara melalui jalur Seleksi Nasional Masuk Perguruan Tinggi Negeri (SNMPTN) dengan pilihan Program Studi Peternakan.
Selama mengikuti perkuliahan, penulis aktif sebagai Asisten Praktikum di Laboratorium Penyuluhan dan Komunikasi Peternakan. Penulis juga aktif dalam organisasi internal dan eksternal universitas, diantaranya pernah menjabat sebagai Koordinator Bidang Humas Asosiasi Mahasiswa Wirausaha USU (2011-2012), Koordinator Departemen Ukhuwah Islamiyah HIMMIP (2012-2013) dan Koordinator Bidang Humas Ikatan Mahasiswa Peternakan (2012-2013). Penulis juga mengikuti Kompetisi Program Mahasiswa Wirausaha (2010), Program Kreativitas Mahasiswa (2010), Big Idea Competition (2011). Penulis juga mendapatkan pelatihan wirausaha dari Ipteks Bagi Kewirausahaan (2012) dan Dinas Perindustrian dan Perdagangan (2012).
KATA PENGANTAR
Puji dan syukur penulis panjatkan kehadirat Allah subhanahu wa ta’ala atas segala karunia dan rahmat-Nya shingga skripsi ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Juli 2014 adalah “Pendugaan Parameter Genetik dan Korelasi Sifat Bobot Lahir, Bobot Sapih dan Litter Size pada Kelinci New Zealand White, Lokal dan Persilangan ”.
Terimakasih penulis ucapkan kepada Hamdan, S.Pt.,M.Si selaku ketua komisi pembimbing dan Prof.Dr.Ir. Hasnudi, MS selaku anggota komisi pembimbing dan Ir. Armyn Hakim Daulay, MBA selaku Penanggungjawab Akademik penulis selama perkuliahan. Penghargaan penulis sampaikan kepada Beasiswa Bidikmisi Kementerian Riset Teknologi dan Perguruan Tinggi.
Ungkapan terimakasih kepada Ayahanda Haryono, ibunda Ridayati dan andong Hj. Deliana Lubis atas segala doa dan perhatian yang diberikan kepada penulis. Terimakasih penulis ucapkan kepada abangda Galih Ari Wirawan Siregar, Ferdy Saputra, Suprayoga dan Adinda Arista Pamel yang senantiasa memberikan dukungan sampai dengan saat ini dalam pembuatan skripsi ini. Semoga Allah SWT membalas dengan kebaikan.
Ucapan terimakasih juga kepada staff pengajar, teman-teman mahasiswa Program Studi Peternakan khususnya angkatan 2010 yang sampai saat ini tidak lelah untuk memberikan semangat kepada penulis dalam penyelesaian skripsi ini.
DAFTAR ISI
Hal.
ABSTRAK ... i
ABSTRACT ... ii
RIWAYAT HIDUP ... iii
KATA PENGANTAR ... iv
DAFTAR ISI ... v
DAFTAR TABEL ... vii
DAFTAR LAMPIRAN ... vii
i PENDAHULUAN Latar Belakang ... 1
Tujuan ... 2
Kegunaan ... 2
TINJAUAN PUSTAKA Klasifikasi Kelinci ... 3
Biologis Kelinci ... 3
Bangsa Kelinci ... 4
New Zealand White ... 4
Lokal ... 5
Cross Breeding ... 6
Sifat Kuantitatif ... 6
Bobot Lahir ... 7
Bobot Sapih ... 8
Litter Size ... 9
Heterosis ... 9
Parameter Genetik ... 0
Heritabilitas ... 0
Ripitabilitas ... 2
Korelasi ... 4
BAHAN DAN METODE PENELITIAN Tempat dan Waktu Penelitian ... 7
Bahan dan Alat Penelitian ... 7
Bahan ... 7
Alat ... 7
Metode Penelitian ... 8
Parameter Penelitian ... 9
Analisis Data ... 2
HASIL DAN PEMBAHASAN Bobot Lahir ... 3
Bobot Sapih ... 4
Efek Heterosis ... 5
Estimasi Nilai Heritabilitas ... 6
Estimasi Nilai Ripitabilitas ... 8
Korelasi Genetik ... 9
KESIMPULAN DAN SARAN Kesimpulan ... 0
Saran ... 0 DAFTAR PUSTAKA
DAFTAR TABEL
Hal.
1. Performans Produksi Kelinci New Zealand White dan Lokal ... 4
2. Sidik Ragam untuk Menduga Nilai Ripitabilitas Suatu Sifat Berdasarkan Pola Tersarang atau Hieraechial ... 20
3. Sidik Ragam untuk Menduga Nilai Korelasi Suatu Sifat Berdasarkan Pola Tersarang atau Hieraechial ... 21
4. Rataan Bobot Lahir Kelinci ... 23
5. Rataan Bobot Sapih Kelinci ... 24
6. Efek Heterosis ... 25
7. Nilai Dugaan Heritabilitas Bobot Lahir, Bobot Sapih dan Litter Size ... 26
8. Nilai Dugaan Ripitabilitas Bobot Lahir, Bobot Sapih dan Litter Size ... 28
DAFTAR LAMPIRAN
Hal.
1. Data Bobot Lahir Kelahiran Pertama ... 35
2. Data Bobot Sapih Kelahiran Pertama ... 35
3. Data Bobot Lahir Kelahiran Kedua ... 36
4. Data Bobot Sapih Kelahiran Kedua ... 36
5. Data Tetua Kelinci New Zealand White ... 36
6. Data Tetua Kelinci Lokal ... 37
7. Analisis Ragam Heritabilitas Bobot Lahir ... 37
8. Analisis Ragam Heritabilitas Bobot Sapih ... 37
9. Analisis Ragam Heritabilitas Litter Size ... 37
10.Analisis Ragam Ripitabilitas Bobot Lahir ... 37
11.Analisis Ragam Ripitabilitas Bobot Sapih ... 38
12.Analisis Ragam Ripitabilitas Litter Size ... 38
13.Efek Heterosis ... 38
14.Format Data Pengamatan Jumlah Anak Kelinci Jantan dan Betina ... 39
15.Format Data Pengamatan Bobot Lahir Kelinci ... 40
16.Format Data Bobot Sapih Kelinci ... 41
17.Format Data Pengamatan Perkawinan Kelinci ... 42