Original paper
ANTIBACTERIAL ACTIVITY OF A SECONDARY
METABOLITE-PRODUCING CORAL BACTERIUM
Pseudoalteromonas SPECIES
Ocky Karna Radjasa1,2*), Torben Marten3, Thorsten Brinkoff 3, Hans-Peter Grossart3,4, Agus Sabdono1,2 and Meinhard Simon3
1
Department of Marine Science, Diponegoro University, Semarang – 50275, Central Java, Indonesia 2
Center for Tropical Coastal and Marine Studies, Diponegoro University, Widya Puraya, Semarang – 50275, Central Java, Indonesia
3
Institute for Chemistry and Biology for Marine environment, University of Oldenburg, 26111 Oldenburg, Germany
4
Institute of Freshwater Ecology and Inland Fisheries (IGB), Dept. Limnology of Stratified Lakes, 16775 Neuglobsow, Germany
Received: October 13, 2003 ; Accepted: December 20, 2004
ABSTRACT
A bacterium, collected at the surface of coral Acropora sp., TAB4.2 was successfully screened for secondary metabolites production based on PCR amplification of the non-ribosomal peptide synthetase gene. It was identified as closely related to Pseudoalteromonas luteoviolacea based on its 16S rDNA. TAB4.2 was found to inhibit the growth of all 5 coral-associated and all 5 pathogenic bacteria tested. To characterize the inhibiting metabolite, a 279 bp long DNA fragment was obtained and the deduced amino acid sequence showed conserved signature regions for peptide synthetases and revealed a high similarity to NosD (40 % identity), a multifunctional peptide synthetase from Nostoc sp. GSV224, and NdaB (44 % identity), a peptide synthetase module of Nodularia spumigena.
Key words: Coral-associated bacterium, secondary metabolites, antibacterial activity,
Pseudoalteromonas
*) Correspondence
: Phone: +62-24-7460038; Fax: +62-24-7460039; E-mail: [email protected]
I
NTRODUCTION
Coral reefs are the most diverse marine ecosystems, however, little is known about their microbial diversity in these ecosystems. It is well understood that
become sources of great interest to natural product chemistry, since they provide a large proportion of bioactive metabolites with different biological activities (Faulkner, 2000). In particular, marine invertebrates with high species diversity in the Indo-Pacific regions (Coll and Sammarco, 1986) are often rich in secondary metabolites and are preferential targets in the search for bioactive natural products (Sammarco and Coll, 1992).
Perhaps the most significant problem that has hampered the investigation of secondary metabolites is their low concentration (Munro et al, 1999). In marine invertebrates many highly active compounds contribute to < 10-6% of the body-wet weight (Procksch et al 2002). Providing sufficient amounts of these biologically active substances, hence, may be a difficult task. Limited amounts found in the producing organism, limited quantities of the organism itself, and geographic or seasonal variations in the produced secondary metabolites (Kelecom, 2002), further complicate the study of secondary metabolites of aquatic organisms.
It is a widely observed phenomenon that microbial cells attach firmly to almost any surface submerged in marine environments, grow, reproduce, and produce extracellular polymers that provide structure to the assemblage termed as biofilm (Kioerboe et al. 2003).
Recently many coral-associated bacteria have been characterized as sources of marine natural products (Moore, 1999), especially since the coral surface is more nutrient rich than seawater or even sediments (Unson et al, 1994; Bultel-Ponce et al, 1999). However, colonization of coral surfaces by bacteria and other microorganisms is mostly nondestructive to corals (Paul et al, 1986; Coffroth, 1990 and Kim, 1994).
Due to the close spatial vicinity of these biofilm-forming bacteria, it can be expected that the indigenous microbial population is adapted to competitive conditions, e.g. for available nutrients and space (Slattery et al, 2001). The production of secondary metabolites is a common adaptation of these bacteria to compete in such microenvironments.
Due to cultivation biases only a minor fraction of heterotrophic microorganisms in the coral reefs has yet been isolated. More information on coral-associated bacteria might be desirable, as many of these bacteria serve as sources of secondary metabolites including novel antibiotics. Here, we report on isolation, screening and characterization of a novel secondary metabolite-producing coral bacterium closely related to
Pseudoalteromonasluteoviolacea.
M
ATERIALS AND
M
ETHODS
Sampling and isolation of
Jepa ra North Java Sea
Scale : 1 : 5.000.000 -6
-7
-8
-9
111 110
109 108
107
106 112 113 114
Fig 1. Sampling site for the collection of coral from Teluk Awur water, Jepara .
PCR-based
screening
of
NRPS
producing bacterial strain
To obtain genomic DNA of strain TAB 4.2 for PCR analysis, cell material was taken from an agar plate, suspended in sterile water (Sigma, Germany) and subjected to five cycles of freeze (-70 °C) and thaw (95 °C). Amplification of peptide synthetase gene fragments was carried out with the degenerated primers A2gamF (5´-AAG GCN GGC GSB GCS TAY STG CC-3´) and A3gamR (5´-TTG GGB IKB CCG GTS GIN CCS GAG GTG-3´) (MWG-Biotech, Ebersberg, Germany) designed from conserved regions of adenylation domains of various bacterial peptide synthetase sequences (GenBank accession numbers: AAK81824, AAK81827, AAK81826, AAC82549, CAA40561, CAC48362, CAA11796, CAC48369, CAC48369, AAF42473, BAB69322, CAB38518, AAG02364, AAG02355, AAG02356, CAA67248, CAB93684, CAB93684, CAB93683, AAC68816, AAC44129, CAA65394, AAG05812, AAG05789, AAG05789, AAF40220, AAD51026, CAC11137, AAB96629). The sequence of the reverse primer was based on the signature sequence of the
synthetases, but also in acetyl-CoA synthetases. The sequence of the forward primer, based on the motif KAGGAY(LV)P (motif A2), is highly conserved for peptide synthetases which are involved in non ribosomal peptide synthesis (Marahiel et al., 1997).
PCR was performed with an Eppendorf Mastercycler (Eppendorf Inc., Germany) as follows: 2 µl template DNA, 40 pmol of each of the appropriate primers, 125 mol of each deoxyribonucleoside triphosphate, 5 µl of 10 x RedTaqTM PCR buffer (Sigma, Germany), 1.2 mg ml-1 (final concentration) bovine serum albumin (Sigma) and 0.75 unit RedTaqTM DNA polymerase (Sigma) were adjusted to a final volume of 50 µl with sterile water (Sigma). A PCR run comprised 40 cycles with denaturing conditions for one minute at 95°C, annealing for one minute at 70 °C and extension for two minutes at 72 °C, respectively.
Cloning and sequencing of a (putative) peptide synthetase domain
protocol. Recombinant clones containing an insert were prepared using the DYEnamic Direct cycle sequencing kit (Amersham Life Science, Inc, UK) for subsequent sequencing on an automated DNA sequencer Model 4200 (LI-COR, Inc, UK). Both strands were sequenced twice using M13F and M13R labeled with IRDyeTM800 as sequencing primers (Messing, 1983). Prior to further analysis of the gene fragment the primer sequences on both sides of the fragment were
PCR amplification and sequencing of
16S rRNA gene fragments
PCR amplification of the almost complete 16S rRNA gene of strain TAB4.2, purification of PCR products and subsequent sequencing analysis were performed according to the method of Brinkhoff and Muyzer (1997). The determined 1204 bp DNA sequence of strain TAB 4.2 was then compared for homology to the BLAST database.
Phylogenetic analysis
All sequences used were at least 1200 bp long. A phylogenetic tree was constructed using maximum-likelihood analysis. Only sequences of type strains were included in tree calculation. Alignment positions at which less than 50 % of sequences of the entire set of data had the same residues were excluded from the calculations to prevent uncertain alignments within highly variable positions of the 16S rDNA. Phylogenetic analysis was performed with
the ARB software package
(http://www.mikro.biologie.tu-muenchen.de (Strunk et al., 1998).
DNA sequence accession numbers.
The 16S rRNA gene sequence of strain TAB4.2 has been entered into the GenBank database under the sequence accession number AY338404, the putative peptide synthetase sequence obtained from strain TAB4.2 under AY338405.
Inhibitory interaction tests
Inhibitory interaction tests of isolate TAB4.2 against other bacteria were performed by using the agar disk-diffusion method (Conception et al, 1994). The following coral-associated bacteria were used: Salinicoccus roseus, Oceanobacillus iheyensis, Halomonas salina, Bacillus iodinum, and Silicibacter lacuscaeruensis
obtained from the collection of the Microbial Ecology group, ICBM, University of Oldenburg, Germany. Pathogenic bacteria used were
Staphylococcus aureus, Escherichia coli, Vibrio harveyi, V. parahaemolyticus, and
V. anguillarum obtained from the culture collection of the Laboratory of pests and diseases, Center for research on brackish water aquaculture, Ministry of Marine Affairs and Fisheries, Jepara, Indonesia.
R
ESULTS AND
D
ISCUSSION
Results
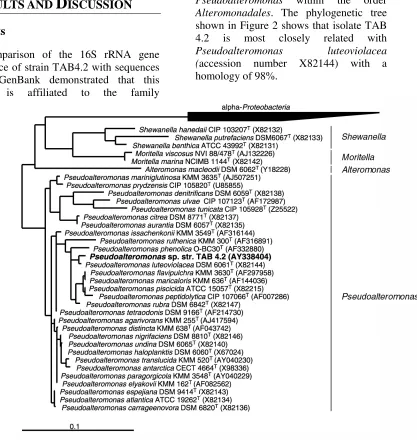
A comparison of the 16S rRNA gene sequence of strain TAB4.2 with sequences from GenBank demonstrated that this strain is affiliated to the family
Pseudoalteromonas within the order
Alteromonadales. The phylogenetic tree shown in Figure 2 shows that isolate TAB 4.2 is most closely related with
Pseudoalteromonas luteoviolacea
(accession number X82144) with a homology of 98%.
Fig 2. Phylogenetic tree based on comparative 16S rRNA gene sequence analysis of
Pseudoalteromonas species showing the phylogenetic affiliation of strain TAB4.2. Selected sequences from the alpha subclass of Proteobacteria were used to root the tree. Accession numbers of the 16S rRNA gene sequences are given in parenthesis. The bar indicates 10% sequence divergence.
To estimate antimicrobial activity of strain TAB4.2 and its biotechnological potential, inhibitory interaction tests with other coral-associated and pathogenic
Table 1. Inhibitory interaction of bacterial strain TAB4.2 against coral bacteria and pathogenic bacteria
No Coral bacteria Antibacterial
activity
Pathogenic bacteria
Antibacterial activity
1 Bacillus iodinum + Escherichia coli +
2 Salinicoccus roseus + Staphylococcus aureus +
3 Silicibacter lacuscaeruensis + Vibrio parahaemolyticus +
4 Oceanobacillus iheyensis + Vibrio harveyi +
5 Halomonas salina + Vibrio anguillarum +
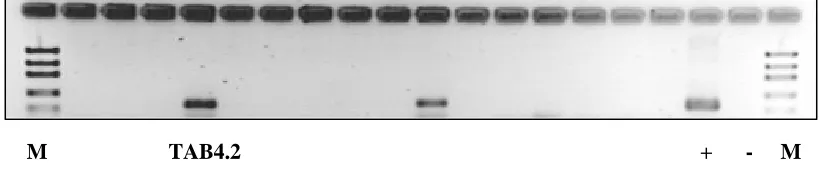
PCR-based screening revealed that the coral-associated bacterial strain TAB4.2 was capable of producing secondary metabolites, in particular a non-ribosomal polypeptides. As indicated in
Figure 3, bacterial strain TAB4.2 possesses the NRPS gene as represented by the occurrence of a single DNA band similar to the positive control on the agarose gel.
M TAB4.2 + - M
Fig 3. PCR-based screening of NRPS producing-TAB4.2 strain.
+ control (Pseudomonas fluorescens DSM No. 50117); M: DNA markers.
To investigate the genetic potential of strain TAB 4.2 to produce secondary metabolites, a 279 bp long DNA fragment was obtained. The deduced amino acid sequence indeed showed conserved signature regions for peptide synthetases. A comparison with proteins in the GenBank database revealed a high similarity to NosD (accession number AAF17281; 40 % identity), a multifunctional peptide synthetase from
Nostoc sp. GSV224, and also to NdaB (accession number AAO64402; 44 % identity), a peptide synthetase module of
Nodularia spumigena.
Discussion
Inhibitory interactions among coral-associated bacteria that occur on the coral surface are of great interest to search for
secondary metabolite-producing bacteria. Isolation and screening for secondary metabolite-producing bacteria in coral reef ecosystems have been strongly neglected until now. Our results highlight one coral-associated bacterium (TAB4.2) carrying the NRPS gene. This bacterium is 98% identical to Pseudoalteromonas
luteoviolacea based on its 16S rRNA gene
sequence. Alteromonadales and Vibrionales of the δ Proteobacteria were among the dominant producers of antibiotics on marine snow from the Southern California Bight (Long and Azam 2001).
competition for space and nutrients with other coral-associated bacteria. This assumption is supported by the fact that our NRPS positive strain, TAB4.2 exhibited antibacterial activity against all tested bacteria. The efficient inhibition of pathogenic bacteria by strain TAB4.2 may further protect the coral from infection (Rohwer et al, 2002).
Not all proteins are synthesized on ribosomes, and small polypeptides can be assembled by peptide synthetases just as other compounds. Most non-ribosomal peptides from microorganisms are classified as secondary metabolites. They rarely play a role in primary metabolism, such as growth or reproduction but have evolved to somehow benefit the producing organisms (Neilan et al, 1999). Products of the microbial non-ribosomal peptide synthesis include the immunosuppressant cyclosporine and other antibiotics such as gramicin S, tyrocin A and surfactins (Kleinkauf and von Dohren, 1996).
The comparison of the derived amino acid sequence of the putative non ribosomal peptide synthetase of strain TAB4.2 revealed a high homology to sequence fragments of known peptide synthetases. Highest similarity was found with sequences of organisms belonging to the phylum Cyanobacteria, from which most genera possess non-ribosomal peptide synthetase genes (Christiansen et al., 2001). Neilan et al (1999) mentioned that
Cyanobacteria produced a myriad array of
secondary metabolites, including alkaloids, polyketides, and non ribosomal peptides, some of which are potent toxins.
The occurrence of structurally related peptides in diverse microorganisms might be due to horizontal gene transfer events of biosynthetic clusters (Kleinkauf and von Doehren, 1996). Interestingly, the organism closest related to TAB4.2,
Pseudoalteromonas luteoviolacea, owns a non-ribosomal peptide synthetase, which produces the siderophore alterobactin
gene product remains unknown, the feasibility that the respective gene detected in strain TAB 4.2 codes for a non ribosomal peptide synthetase is high.
C
ONCLUSION
The present work highlights the production of secondary metabolites by a symbiotic coral bacterium (TAB4.2) carrying the NRPS gene. The expression of the NRPS gene accounts for the biosynthesis of various natural products with different biological activity (Silakowski et al, 2000). Hence, the application of molecular approach through PCR using specific NRPS primers provides rapid detection and is suitable to greatly improve the screening efficiency for secondary metabolite-producer among coral-associated bacteria. Understanding the genetic basis as well as the biochemistry of specific bacteria such as TAB4.2 will facilitate genetic engineering aimed at improving design of antimicrobial substances.
A
CKNOWLEDGEMENTS
R
EFERENCES
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman D J. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic. Acids. Res. 25: 3389-3402.
Brinkhoff T, Muyzer G. 1997. Increased species diversity and extended habitat range of sulfur-oxidizing
Thiomicrospira spp. Appl.
Environ. Microbiol. 63:
3789-3796.
Bultel-Ponce V, Berge J-P, Debitus C, Nicolas J-L, Guyot M. 1999. Metabolites from the
sponge-associated bacterium
Pseudomonas species. Mar.
Biotechnol. 1: 384-390.
Burgess JG, Boyd KG, Amstrong E, Jiang Z, Yan L, Berggren M, May U, Pisacane T, Granmo A, Adams DR. 2003. Development of a marine natural product-based antifouling paint. Biofouling. 19: 197- 205. 2001. Nonribosomal peptide synthetase genes occur in most cyanobacterial genera as evidenced by their distribution in axenic strains of PCC. Arch. Microbiol.
176(6): 452-458.
Coffroth MA. 1990. The function and fate mucous sheets produced by reef coelenterates. Procc The 6th Int
Coral Reef Symp. 2: 15-20.
Australia.
Coll JC, Sammarco PW. 1986. Soft corals: Chemistry and ecology. Oceanus.
29 (2): 33-37.
Deng J G, Hamada Y, Shioiri T. 1995. Total synthesis of alterobactin-A, a super siderophore from an open-ocean bacterium. J. Am. Chem. Soc. 117 (29): 7824-7825.
Faulkner DJ. 2000. Marine pharmacology.
Antonie van Leeuwenhoek. 77:
135-145.
Fenical W. 1993. Chemical studies of marine bacteria: developing a new source. Chem. Rev. 93: 1673-1683.
Jensen PR, Fenical W. 2000. Marine
microorganisms and drug
discovery: current status and future potential. In: Fusetani N (ed). Drugs from the sea, Basel: Karger, pp 6-29.
Kelecom A. 2002. Secondary metabolites from marine microorganisms. An. Acad. Bras. Cienc. 74:151-170.
Kim K. 1994. Antimicrobial activity in gorgonian corals (Coelenterata, Octocorallia). Coral. Reefs. 13: 75-80.
Kiorboe T, Grossart HP, Ploug H, Kam T. 2003. Microbial dynamics on particles: colonization, growth, detachment, and grazing mortality of attached bacteria. Appl. Environ. Microbiol. 69:3036-3047.
Kleinkauf H, von Doehren H. 1996. A nonribosomal system of peptide biosynthesis. Eur. J. Biochem.
Kushmaro A, Loya Y, Fine M, Rossenberg E. 1996. Bacterial infection and coral bleaching. Nature. 380: 396.
Kushmaro A, Rossenberg E, Fine M, Loya Y. 1997. Bleaching of the coral
Oculina patagonica by Vibrio
AK-1. Mar. Ecol. Prog. Ser. 147: 159-165.
Long R , Azam F. 2001. Antagonistic interactions among marine pelagic bacteria. Appl. Environ. Microbiol.
67:4975-4983.
Ludwig W, Strunk O, Klugbauer S, Klugbauer N, Weizenegger M, Neumaier J, Bachleitner M, Schleifer KH. 1998. Bacterial phylogeny based on comparative
sequence analysis.
Electrophoresis. 19(4): 554-68.
Madigan MT, Martinko JM, Parker J, Brock T.D. 2000. Biology of
microorganisms. Prentice-Hall,
Inc., Upper Saddle River, New Jersey 07458.
Marahiel MA, Stachelhaus T, Mootz HD.
1997. Modular peptide
synthetases involved in nonribosomal peptide synthesis.
Chem. Rev. 97: 2651-2673.
Messing J. 1983. New M13 vectors for cloning. Methods. Enzymol. 10: 20-78.
Moore BS. 1999. Biosynthesis of marine natural products : microorganisms and macroalage. Nat. Prod. Rep. 16: 653-674.
Munro MHG, Blunt JW, Dumdei EJ, Hickford SJH, Lill RE, Li S, Battershill CN, Duckworth AR. 1999. The discovery and
compounds with pharmaceutical potential. J. Biotechnol. 70: 15-25.
Neilan BA, Dittmann, Rouhiainen L, Bass RA, Schaub V, Sivonen K, Borner T. 1999. Nonribosomal peptide synthesis and toxigenicity of cyanobacteria. J. Bacteriol. 181:4089-4097.
Paul JH, DeFlaun ME, Jefffrey WH. 1986. Elevated levels of microbial activity in the coral surface
Pietra F. 1997. Secondary metabolites from marine microorganisms- bacteria, protozoa, algae and
fungi-achievements and
prospectives. J. Nat. Prod. 14: 453-464.
Proksch P, Edrada RA, Ebel R. 2002. Drugs from the seas-current status and microbiological implications.
Appl. Microbiol. Biotechnol. 59: 125-134.
Reid R T, Live DH, Faulkner DJ, Butler A. 1993. A siderophore from a marine bacterium with an exceptional ferric ion affinity constant. Nature. 366(6454):455-8.
Rohwer F, Breitbart M, Jara J, Azam F, Knowlton N. 2001. Diversity of bacteria associated with the Caribean coral Montastraea franksi. Coral. Reefs 20: 85-95.
bacteria. Mar. Ecol. Prog. Ser. 243: 1-10.
Rublee, P.A., Lasker, H.R., Gottfried, M, and Roman, M.R. 1980. Production and bacterial colonization of mucus from the soft coral Briarium asbestinum. Bull. Mar. Sci. 30:888-893.
Sammarco PW, Coll JC. 1992. Chemical adaptation in the Octocorallia: Evolutionary considerations. Mar. Ecol. Prog. Ser. 88: 93-104.
Santavy, DL., Peters, E.C., Kozlowski, J., and Wilkinson, S. 1995. Characterization of the bacterium suspected in the incidence of white band disease. Abstr. Gen. Mee.t Am. Soc. Microbiol. p. 332.
Shashar, N., Cohen, Y., Loya, Y., and Sar, N. 1994. Nitrogen fixation (acetylene reduction) in stony corals: evidence for coral-bacteria interactions. Mar. Ecol. Prog. Ser. 111: 259-264.
Silakowski, B., G. Nordsiek., B. Kunze., H. Blöker., and R. Müller. 2000. Novel features in a combined polyketide synthase/non-ribosomal peptide synthetase: the myxalamid biosynthetic gene cluster of the myxobacterium Stigmatella
aurantiaca Sg a151. Chem. Biol.
53:1-11.
Slattery M, Rajbhandari I, Wesson K. 2001. Competition-mediated antibiotic induction in the marine
bacterium Streptomyces
tenjimariensis. Microb. Ecol.
41:90-96.
Strunk, O., Gross, O., Reichel, B., May, M., Hermann, S., Stuckmann, N., Nonhoff, B., Lenke, M., Ginhart, A., Vilbig, A., Ludwig, T., Bode, A., Schleifer, K.-H., and Ludwig, W. 1998. ARB: a software environment for sequence data. http://www.mikro.biologie.tu-muenchen.de/pub/ARB.
Department of Microbiology, Technische Universität München, Munich, Germany.
Unson, M.D., Holland, N.D., Faulkner, D.J. 1994. A brominated secondary metabolite synthesized by the cyanobacterial symbiont of marine sponge and accumulation of the crystalline metabolites in the sponge tissue. Mar. Biol. 119: 1-11.