BAB I
PENDAHULUAN 1.1 Latar Belakang
Jumlah kromosom lalat buah Drosophila melanogaster relative sedikit jika dibandingkan kromosommakhluk hidup lainya. Lalat jantan ialah 3AAXY (3 pasang autosom + sebuah kromosom X dan sebuah kromosom Y. Oleh karena kromosom kelamin pada lalat betina itu sejenis (artinya kedua-duanya berupa kromosom X), maka lalat betina dikatakan bersifat homogametic. Lalat jantan bersifat heterogametic, sebab dua buah kromosom kelamin yang dimilikinya satu sama lain berbeda (yang satu kromosom X dan yang satu kromosom Y). Dalam keadaan normal, lalat betina membentuk satu macam sel telur saja yang bersifat haploid (3AX). Tetapi lalat jantan membentuk 2 macam spermatozoa yang haploid. Ada spermatozoa yang membawa kromosom X (3AX) dan ada yang membawa kromosom Y (3AY). Apabila sel telur itu dibuahi spermatozoon yang membawa kromosom X, terjadilah lalat betina yang diploid (3AAXX). Tetapi bila sel telur itu dibuahi spermatozoa yang membawa kromosom Y, terjadilah lalat jantan yang diploid (3AAXY). Kadang-kadang di waktu meiose selama pembentukan sel-sel kelamin, sepasang kromosom kelamin itu tidak memisahkan diri, melainkan tetap berkumpul. Peristiwa ini disebut nondisjun
Peristiwa Nondisjunction (gagal berpisah) adalah kegagalan untuk memisah kromosom pada peristiwa mitosis dan meiosis. Sebagai contoh pada meiosis, tiap-tiap anggota suatu pasangan kromosom menuju hanya pada satu kutub, sehingga kutub yang lain tidak menerima pasangan kromosom. Pai, (1987: 406) menyatakan nondisjunction adalah penyimpangan pembelahan sel, di mana kromosom-kromosom atau kromatid-kromatid yang secara normal berpisah pada waktu anafase tetap tinggal bersama, menghasilkan sel anak dengan kebanyakan atau kekurangan kromosom.
Berdasarkan berbagai hal diatas, peneliti tertarik untuk mengkaji lebih lanjut tentang pengaruh jenis kelamin terhadap frekuensi
nondisjunction pada D. melanogaster. Selain itu penelitian mengenai persilangan D. melanogaster strain Normal jantan dengan strain e betina beserta resiproknya belum banyak dipublikasikan. Oleh karenanya, maka dilakukan penelitian dengan judul “pengaruh jenis kelamin terhadap frekuensi fenomena gagal berpisah (non-disjuction) Drosophila mlanogaster persilangan N♀><e♂ beserta Resiproknya”.
1.2 Rumusan Masalah
Berdasarkan latar belakang di atas, maka dapat dirumuskan masalah sebagai berikut.
Adakah perbedaan frekuensi fenomena gagal berpisah (nondisjunction) persilangan Droshopila melanogaster ♀N × ♂e beserta resiprok pada jenis kelamin yang berbeda?
1.3 Tujuan Penelitian
Berdasarkan rumusan masalah di atas, tujuan dari penelitian ini adalah,
Untuk mengetahui ada atau tidaknya perbedaan frekuensi fenomena gagal berpisah (nondisjunction) persilangan Droshopila melanogaster ♀N × ♂e beserta resiprok pada jenis kelamin yang berbeda.
1.4 Kegunaan Penelitian
Hasil penelitian ini memiliki kegunaan baik dari segi pengembangan ilmu maupun terapan, antara lain:
1. Menambah pengalaman, ketrampilan, dan kecakapan dalam melakukan penelitian.
2. Menambah pengetahuan kepada para pembaca umumnya, dan khususnya kepada mahasiswa Biologi UM angkatan 2014 tentang perbedaan frekuensi fenomena gagal berpisah (nondisjunction) persilangan Droshopila melanogaster ♀N × ♂e beserta resiprok pada jenis kelamin yang berbeda sehingga mahasiswa mampu menerapkannya dalam kehidupannya kelak baik dari segi ilmunya ataupun dari segi keterampilan dalam melakukan penelitian.
1.5 Ruang Lingkup dan Batasan Masalah Ruang lingkup dalam penelitian ini meliputi:
1. Strain D. melanogaster yang digunakan dalam penelitian ini adalah strain N, strain e yang diperoleh dari Laboratorium Genetika Jurusan Biologi FMIPA UM.
2. Pengamatan fenotip yang dilakukan meliputi warna mata, faset mata, warna tubuh, dan bentuk sayap.
3. Penelitian ini dibatasi pada Drosophila melanogaster yang memiliki gen terpaut pada kromosom kelamin.
4. Drosophila melanogaster yang disilangkan memiliki usia maksimal 3 hari setelah menetas dari pupa hitam.
5. Pengambilan data diperoleh dari pengamatan pada hasil fenotip persilangan F1.
6. Data yang diperoleh berupa data hasil pengamatan fenotip serta jumlah F1
1.6 Asumsi Penelitian
1. Kondisi medium dari semua persilangan dan ulangan dari awal sampai akhir dianggap sama.
2. Semua jumlah medium dalam tiap-tiap botol ulangan persilangan dari awal sampai akhir penelitian dianggap sama.
3. Semua kondisi lingkungan seperti suhu, cahaya, kelembapan, dan tempat perkembangbiakan selama penelitian berlangsung dianggap sama.
4. Umur dan kondisi D. melanogaster yang digunakan dalam penelitian dianggap sama.
1.7 Definisi Istilah
Adapun beberapa defenisi istilah adalah sebagai berikut.
1. Peremajaan adalah kegiatan mengembangbiakkan lalat sejenis dari stok kedalam medium baru.
2. Pengampulan adalah kegiatan memindahkan pupa hitam kedalam selang yang telah diberi pisang yang kemudian ditutup dengan spon. 3. Perkawinan resiprok adalah perkawinan kebalikan dari perkawinan
4. Strain adalah suatu kelompok intra spesifik yang memiliki hanya satu atau sejumlah kecil ciri yang berbeda, biasanya secara genetik dalam keadaan homozigot untuk ciri-ciri tersebut atau galur murni. 5. Fenotipe adalah ciri tubuh yang dapat diamati pada suatu individu.
6. Genotip adalah sifat yang tidak tampak dari luar; keseluruhan jumlah informasi genetik yang terdapat pada suatu individu.
7. Alela adalah sepasang kromosom homolog sesamanya (memiliki lokus yang sama).
8. Homozigot adalah individu yang genotipnya terdiri dari alel yang sama.
9. Heterozigot adalah individu yang genotipnya terdiri dari sepasang alel yang tidak sama.
10.Nondisjunction adalah kegagalan memisah pada kromosom pada peristiwa meiosis.
BAB II
KAJIAN PUSTAKA 2.1 Sistematika
Drosophila melanogaster meupakan jenis lalat buah, dimasukkan dalam filum Artropoda kelas Insekta bangsa Diptera, anak bangsa Cyclophorpha (pengelompokan lalat yang pupanya terdapat kulit instar 3, mempunyai jaw hooks), seri Acaliptrata (imago menetas dengan keluar dari bagian anterior pupa), suku Drosophilidae. Jenis Drosophila melanogaster di Indonesia terdapat sekitar 600 jenis, pulau Jawa sekitar 120 jenis dari suku Drosophilidae (Wheeler dalam Santoso, 2009).
Menurut Storer (1975) dan Bock (1976) dalam Kusumawati (1995) sistematika Drosophila melanogaster adalah
Filum : Arthropoda
Kelas : Insecta
Anak kelas : Pterygota
Bangsa : Diptera
Anak Bangsa : Cyclorrhapha
Suku : Drosophilidae
Marga : Drosophila
Spesies : Drosophila melanogaster
2.2 Deskripsi D. melanogaster
Lalat buah dan Artrophoda lainnya mempunyai suatu seri segmen yang teratur. Segmen ini menyusun tiga bagian tubuh utama, yaitu; kepala, thoraks, dan abdomen. seperti hewan simetris bilateral lainnya, Drosophila ini mempunyai poros anterior dan posterior (kepala-ekor) dan poros dorsoventral (punggung-perut). Drosophila memiliki ciri morfologi yang berbeda antara jantan dan betinanya. Pada Drosophila jantan memiliki ukuran tubuh yang lebih kecil bila dibandingkan dengan yang betina. Memiliki 3 ruas dibagian abdomennya dan memiliki sisir kelamin. Sedangkan pada yang betina ukuran relatif lebih besar, memiliki 6 ruas pada bagian abdomen dan tidak memiliki sisir kelamin. (Soemartomo.S.S dalam Santoso, 2009).
Drosophila melanogaster normal memiliki mata yang berwarna merah berbentuk elips. Terdapat pula mata oceli yang ukurannya jauh lebih kecil dari mata majemuk, berada pada bagian atas kepala, di atas di antara mata dua mata majemuk, berbentuk bulat. Selain itu, Drosophila melanogaster normal memiliki sungut yang berbentuk tidak runcing dan bercabang-cabang. Kepala berbentuk elips. Thorax terlihat berwarna krem, ditumbuhi banyak bulu, dengan warna dasar putih. Abdomen bersegmen lima, segmen terlihat dari garis-garis hitam yang terletak pada abdomen. Sayap Drosophila normal memiliki ukuran yang panjang hingga melebihi abdomen lalat, lurus, dan
bermula dari thorax dengan warna transparan (Ghostrecon dalam Santoso, 2009).
2.3 Penetuan Kelamin pada Drosophila melanogaster
Tidak semua organisme yang bereproduksi secara seksual mempunyai sistem penetuan kelamin seperti pada manusia, misalnya D. melanogaster, lalat buah, mempunyai suatu mekanisme yang “seimbang”. Suatu keseimbangan antara jumlah perangkat otosom dan jumlah kromosom X, menentukan fenotip seksual lalat buah (Pai,1985).
Menurut Corebima (2004) pada Drosophila melanogaster terdapat kromosom kelamin X dan Y. Dalam keadaan diploid normal ditemukan pasangan kromosom kelamin XX dan XY, atau pasangan kromosom secara lengkap sebagai AAXX dan AAXY (jumlah autosom sebanyak tiga pasang). Mekanisme ekspresi kelamin pada Drosophila melanogaster dikenal sebagai suatu mekanisme perimbangan antara X dan A atau X/A. Lanjut Pai (1985) dalam Corebima (2004) menyebutkan mekanisme itu sebagai suatu mekanisme keseimbangan determinasi kelamin.
Dari eksperimen Bridges dalam Novitasari (1992) dihasilkan variasi kombinasi kromosom X dengan autosom pada Drosophila. Dari perbandingan-perbandingan tersebut disimpulkan bahwa perbandingan satu kromosom X dan dua set autosom menghasilkan lalat jantan Normal (XAA) atau jantan diploid, dengan rasio kromososm X dengan autosom 0,5. Kombinasi dua kromosom X dengan dua perangkat autosom (2X + 2A, rasio 2 : 2 = 1) menghasilkan betina diploid. Pada rasio perbandingan kromosom X dengan autosom 3:2 dihasilkan lalat betina yang metafemales. Lalat bergenotip XXY (2X/2A) adalah betina normal sedangkan genotip XO (1X/2A) adalah lalat jantan tetapi steril. Pada lalat XO secara fisiologi sperma terbentuk tetapi sperma tersebut non motil. Hal tersebut menunjukkan bahwa kromosom Y pada D. melanogaster tidak
berperanan pada penetuan jenis kelamin. Tetapi kromosom Y berperanan dalam menentukan fertilitas jantan.
2.4 Peristiwa Meiosis
Meiosis merupakan cara pembelahan sel yang khusus, terjadi pada waktu pematangan sel-sel benih, yang membagi angka kromosom menjadi setengahnya. Pemisahan (penyebaran) kromosom homolog sewaktu meiosis melalui pembelahan reduksi (Pai, 1985). Lanjut Pai bahwa “beberapa dari tahap meiosis sangat menyerupai tahap-tahap terkait yang terdapat pada mitosis. Meiosis seperti halnya mitosis, didahului oleh replikasi kromosom”.
Meiosis terjadi selama gametogenesis. Meiosis merupakan dua pembelahan inti yang berurutan, bertujuan untuk pembentukan gamet. Selama meiosis setiap sel membelah dua kali, sedangkan kromosom hanya sekali diduplikasi, sehingga gamet-gamet yang dihasilkan mempunyai jumlah kromosom setengah dari sel induknya. Pembelahan dua kali tersebut disebut sebagai meiosis I dan meiosis II. Masing-masing dari pembelahan meiosis tersebut mempunyai fase-fase yang sama seperti pada mitosis, yaitu profase-fase, metafase-fase, anafase-fase, dan telofase. Akan tetapi terdapat perbedaan-perbedaan penting pada perilaku kromosomnya, terutama dalam pembelahan yang pertama (meiosis I). Meiosis I merupakan pembelahan reduksi, sebab inti-inti yang dihasilkan hanya mempunyai setengah dari jumlah kromosom sel induk, sedangkan meiosis II merupakan pembelahan eukasional, sebab inti-inti yang dihasilkan dari meiosis II identik satu dengan yang lain. Berkenaan dengan hal ini, pada bagian lain menyatakan bahwa meiosis I dimulai dengan sel yang mengandung 2n kromosom dan berakhir dengan dua sel, setiap selnya mengandung n kromosom, sedangkan meiosis II diakhiri dengan empat sel, setiap selnya mengandung n kromosom (Ayala 1984 dalam Balqis 1995).
Fenomena gagal berpisah (non-disjunction) pada persilangan Drosophila melanogaster pertama kali dilaporkan oleh T.H. Morgan dan Bridges. Mereka menemukan penyimpangan pada persilangan antara individu betina bermata putih dengan jantan bermata merah, dimana hasil turunan (F1) yaitu jantan bermata putih dan betina bermata merah. Akan tetapi, dilaporkan pula bahwa satu diantara 2000 turunan F1 tersebut mempunyai warna mata menyimpang, entah betina bermata putih atau jantan bermata merah. Bridges menduga bahwa penyimpangan itu terjadi karena gagal berpisah pada kromosom kelamin X. Dalam hal ini kedua kromosom kelamin X gagal memisah selama meiosis sehingga keduanya menuju kutub yang sama, dan terbentuklah telur yang memilki dua kromosom kelamin X maupun yang tidak memiliki kromosom kelamin X (Corebima, 2003).
Nondisjunction (gagal berpisah) adalah kegagalan untuk memisah pada kromosom pada peristiwa mitosis dan meiosis. Sebagai contoh pada meiosis, tiap-tiap anggota suatu pasangan kromosom menuju hanya satu kutub, sehingga kutub yang lain tidak menerima pasangan kromosom (Gardner, dkk., 1991). Pai, (1987: 406) menyatakan nondisjunction adalah penyimpangan pembelahan sel, di mana kromosom-kromosom atau kromatid-kromatid yang secara normal berpisah pada waktu anaphase tetap tinggal bersama, menghasilkan sel anak dengan kebanyakan atau kekurangan kromosom. Peristiwa nondisjunction dibedakan menjadi nondisjunction primer dan sekunder. Nondisjunction primer dapat terjadi pada induk lalat yang belum mengalami nondisjunction atau lalat “Normal”, sedangkan nondisjunction sekunder terjadi pada keturunan yang merupakan hasil nondisjunction primer.
Berkenaan dengan kejadian nondisjunction pada Drosophila seperti yang dikemukakan pertama kali oleh Bridges tahun 1916, Tamarin, dkk., (1991) menjelaskan, bahwa kejadian nondisjunction tersebut dapat dijelaskan melalui kejadian nondisjunction pada betina Normal, dalam hal ini betina Normal yang mengalami nondisjunction
saat meiosis, akan menghasilkan telur XwXw dan 0 telur (tanpa kromosom sek). Jika telur XwXw dibuahi oleh Y yang dibawa sperma, akan dihasilkan turunan betina bermata putih (XwXwY). Jika telur yang tanpa kromosom sek dibuahi oleh X yang dibawa sperma, akan menghasilkan keturunan jantan Normal (X+O). Tipe lain dari kejadian nondisjunction adalah telur XX yang dibuahi oleh Y yang dibawa sperma dan telur O yang akan dibuahi oleh Y yang dibawa sperma. Zigot XXX bergenotip XwXwX+ (betina Normal) biasanya mati dan lalat YO (selalu mati). Contoh persilangan antara D. melanogaster strain Normal jantan dan strain white betina yang menghasilkan keturunan nondisjunction.
Berkenaan dengan waktu terjadinya nondisjunction, Strickberger, (1976) menyatakan bahwa ada dua alternative kegagalan memisah atau “disjoin” kromosom kelamin; pertama adalah yang terjadi pada saat metaphase meiosis pertama, ketika dua kromosom homolog yang tidak saling berhadapan dan kemudian memisah menuju ke salah satu kutub yang sama, sedangkan kedua adalah yang terjadi pada saat pembelahan meiosis kedua. Dijelaskan lebih lanjut, bahwa jika nondisjunction pada meiosis pertama terjadi pada individu jantan maka akan dihasilkan gamet XY dan O (tanpa X) dan jika terjadi pada meiosis kedua kemungkinan akan menghasilkan sperma XX, YY, dan O. Jika individu betina yang mengalami nondisjunction pada meiosis pertama atau meiosis kedua akan menghasilkan sel telur XX atau O.
Herskowitz, (1977) menyatakan bahwa nondisjunction dapat terjadi (1) baik pada aotosom ataupun kromosom kelamin, (2) selama mitosis ataupun meiosis, dan (3) pada individu jantan maupun betina. 2.6 Faktor-Faktor Penyebab Gagal Berpisah (Non Disjunction)
Banyak faktor yang menyebabkan peristiwa gagal berpisah terjadi, misalnya radiasi tinggi dan zat kimia (Herskowits 1965 dalam Balqis 1995). Selain itu peristiwa gagal berpisah dapat disebabkan faktor dari dalam, yaitu faktor-faktor yang berkaitan dengan kelainan-kelainan pada tingkah laku genetik. Salah satu fenomena genentik
yang dapat menyebabkan peristiwa gagal berpisah adalah hybrid disgenesis (Sved 1979 dalam Balqis 1995). Hibrid disgenesis dapat diartikan sebagai sebuah sindrom yang berhubungan dengan sifat-sifat genetis yang terjadi secara spontan, disebabkan saling berinteraksinya beberapa strain yang disilangkan (Kidwell dan Kidwell 1977 dalam Balqis 1995). Beberapa sifat genetis lain yang juga berhubungan dengan fenomena hybrid disgenesis dikemukakan oleh Sved (1979), yaitu rekombinasi pada jantan, kerusakan kromososm pada saat meiosis, perubahan pada rekombinasi betina, sterilitas, dan lain sebagainya. Lebih lanjut Sved (1979) menyatakan bahwa mekanisme interaksi antara strain-strain yang disilangkan tersebut belum jelas (Balqis,1995)
Salah satu penyebab terjadinya hybrid disgenesis adalah adanya urutan DNA yang dapat bergerak dan menyelip diantara urutan DNA yang ada. Menurut Gardner(1991), urutan DNA yang dapat bergerak itu disebut sebagai trasnposibel elemen atau secara singkat disebut sebagai trasnposon. Watson(1987) menyatakan bahwa trasnpososn yang bertanggung jawab terhadap fenomena genetic hybrid disgenesis adalah elemn P. Hal ini juga ditegaskan olah Gardner (1991) dengan pernyataan senada, yaitu elemen P yang hadir dalam suatu hybrid dapat menyebabkan sindrom abnormalitas genetik yang dikenal dengan hybrid disgenesis (Balqis,1995)
Gardner(1991) menyatakan bahwa diperkirakan elemen P masuk dengan cara’membonceng’ virus yang secara alami menginfeksi Drosophila. Penelitian lebih lanjut berusaha mencari mekanisme pergerakan elemen P pada populasi Drosophila yang mengandung elemen P. Penelitian yang dilakukan pada beberapa strain Drosophila menunjukkan bahwa mekanisme pergerakan elemen P tergantung pada sifat maternal yang diturunkan. Dalam hal ini terdapat (dibedakan) dua tipe Drosophila yaitu P-cytotype yang mengandung elemen P dan mewakili pihak ayah (paternal) dan M-cytotype yang tidak mengandung elemen dan mewakili pihak ibu (maternal) (Balqis,1995).
Sehubungan dengan hal tersebut , (Watson1987 dalam Balqis 1995) menyatakan bahwa bila Drosophila tipe P-cytotype jantan disilangkan dengan Drosophila tipe M-cytotype betina, elemen P yang terkandung pada individu jantan akan diturunkan ke dalam hybrid dan dapat dengan bebas mentrasnposase.Tetapi bila yang disilangkan adalah Drosophila tipe P-cytotype betina dengan Drosophila tipe M-cytotype jantan, maka elemen P tidak dapat ditrasnposasekan sebab sitoplasma telur (individu P-cytotype betina) mengandung molekul tipe repressor yang menghalangi trasnposase elemen P.
Lebih lanjut, Gardner (1991) menyatakan bahwa penyelipan elemen P pada rantai DNA yang sudah ada dapat meningkatkan frekuensi mutasi dan kerusakan kromosom, dalam kasus yang ekstrem dapat menyebabkan cacat pada perkembangan gonad dan pada akhirnya menyebabkan kondisi steril. Bila hal ini dikaitkan dengan peristiwa gagal berpisah, peristiwa ini merupakan salah satu peristiwa yang mengakibatka mutasi kromososm. Sehingga dimungkinkan kejadiannya disebabkan oleh peritiwa trasnposabel elemen dalam fenomena hybrid disgenesis (Balqis ,1995).
Faktor dari dalam lainnya yang berpengaruh terhadap peristiwa gagal berpisah adalah umur. Menurut Pai (1985:39),”experiments on lower forms of life have indicated that age tends to increase the incidence of an aberration of meiosis called’nondisjunction’. Dari pernyataan tersebut dapat dikatakan bahwa peristiwa gagal berpisah cenderung meningkat dengan semakin bertambahnya umur, khususnya pada bentuk kehidupan yang rendah. Tidak dijelaskan lebih lanjut tentang artikata ”lower forms of life” (Balqis,1995).
BAB III
METODE PENELITIAN A. Rancangan Penelitian
Penelitian ini merupakan penelitian eksperimen perlakuan dalam penelitian ini adalah jenis kelamin. Pada penelitian ini dilakukan persilangan D. melanogasterstrain N♀ ><e♂ beserta resiproknya. Selanjutnya data akan dianalisis dengan deskriptif kuantitatif.
B. Variable Penelitian
1. Variabel bebas: jenis kelamin
2. Variabel terikat: frekuensi nondisjunction 3. Variabel kontrol: usia induk, suhu, cahaya
C. Waktu Dan Tempat Pelaksanaan 1. Waktu Penelitian
Penelitian ini dilakukan dari bulan Oktober 2016 hingga bulan November 2016.
2. Tempat penelitian
Laboratorium Genetika, gedung O5 lantai 3 ruang 310, FMIPA UM.
D. Populasi dan Sampel 1. Populasi
D. melanogaster yang dibiakkan di laboratorium genetika jurusan Biologi FMIPA UM
2. Sampel
Sampel dalam penelitian ini adalah D. melanogaster rstrain N, dan e yang didapatkan di Laboratorium Genetika Jurusan Biologi FMIPA UM.
E. Alat dan Bahan 1. Alat
Alat yang digunakan dalam penelitian ini adalah sebagai berikut; blender, spons, selang ampul, pisau, botol selai, panci, sendok, kertas pupasi, kain kasa, kompor gas, Pengaduk kayu, kuas, plastik, gunting, karet, spidol, Mikroskop stereo, Lemari es, kardus, tisu.
2. Bahan
Bahan yang digunakan antara lain: pisang raja mala, air, gula merah, yeast, tape singkong.
F. Prosedur Kerja 1. Pembuatan medium
Menimbang bahan pisang Rajamala, tape singkong dan gula merah dengan perbandingan 7:2:1
Menghaluskan ketiga bahan dengan blender, kemudian menuangkannya ke dalam panic
Menambahkannya dengan air secukupnya
Memasaknya selama 45 menit sambil diaduk (usahakan tidak terlalu encer dan tidak terlalu kental), kemudian didinginkan Memasukkan medium yang telah masak ke dalam botol
persilangan sebanyak seperlima bagian dari tinggi botol persilangan
Memberikan yeast secukupnya dan meletakkan kertas pupasi ke dalam botol tersebut
Menutup botol tersebut dengan spon yang telah dipotong sesuai ukuran
2. Persiapan stok induk
Menyiapkan botol selai yang telah diisi medium untuk membiakkan stok induk strain N, dan e.
Memberi label pada botol sesuai strain dan tanggal memasukannya.
Mengamati perkembangan D. melanogastersampai muncul pupa.
Mengisolasi pupa yang telah menghitam dengan menggunakan kuas yang sudah di basahi dengan air.
Meletakkan pupa pada selang ampul yang telah diberi pisang dan menutup sisi selang dengan spons.
Menunggu hingga pupa menjadi imago dan lalat yang sudah dewasa untuk disilangkan (siap untuk disilangkan).
3. Tahap persiapan Persilangan dan perlakuan (F1)
Menyiapkan botol yang sudah berisi medium dan ditutup dengan spons.
Menyilangkan D. melanogaster strain N♂ dengan strain e♀ beserta resiproknya dari ampulan yang sudah menetas (Usia lalat maksimal tiga hari setelah menetas dari selang ampul), kemudian memasukkan ke dalam botol selai baru yang berisi medium dengan masing-masing konsentrasi dilakukan minimal 3 kali ulangan.
Memberi label sesuai dengan jenis strain persilangan disertai dengan catatan ulangan, konsentrasi sodium siklamat pada medium dan tanggal persilangan.
Melepas jantan setelah 2 hari persilangan, kemudian menunggu sampai ada pupa dan memindahkan si betina ke medium berikutnya sampai si betina itu sendiri mati atau berhenti bertelur(minimal botol B).
Membiarkan sampai muncul anak, kemudian mengamati fenotipe yang muncul pada F1, menghitung jantan dan betina anak pada setiap strain, setiap ulangan, setiap generasi selama 7 hari (harike 0-6).
G. Teknik Analisis Data
Teknik analisa data yang digunakan adalah rekontruksi persilangan dan menghitung frekuensi nondisjunction yang muncul pada F1 dari persilangan Dhrosophila melanogasterstrain N♀ × e♂ beserta resiproknya . Persentase nondisjunction dihitung dengan rumus:
Frekuensi NDJ (%) = ΣTotal keturunan FΣ NDJ 1x100 , yang selanjutnya akan dianalisi dengan ANAVA 1 jalur.
Tabel Frekuensi NDJ
persilangan Frekuensi NDJ pada
ulangan
ke-∑
1 2 3 4
N♀ × e♂
N♂xe♀
Selanjutnya dianalisis secara statistik dengan langkah-langkah sebagai berikut: Faktor korelasi = FK=ΣX❑ 2 n JKtotal=Σ Xi2−FK JKperlakuan(kombinasi)=ΣXd t 2 −FK JKgalat=JKtotal−JKperlakuan
Tabel 4.5 Ringkasan Analisis Varian
SK Db JK KT Fhitung Ftabel 5%
Perlakua Jenis kelamin Galat
Jika F hitung jenis kelamin > Ftabel(0,05) maka H0 ditolak, ada perbedaan frekwensi fenomena gagal berpisah (nondisjunction) persilangan Droshopila melanogaster ♀N × ♂e beserta resiprok pada jenis kelamin yang berbeda.
BAB IV
DATA DAN ANALISA DATA
4.1 Data Hasil Pengamatan
1) Morfologi dari masing-masing strain Drosophila melanogaster
a. Drosophila melanogaster strain N
Gambar Keterangan
Sayap menutupi tubuh
tubuh kuning kecoklatan
Faset mata merah
Terdapat black spot pada abdomen posterior.
Untukbetina tidak ada
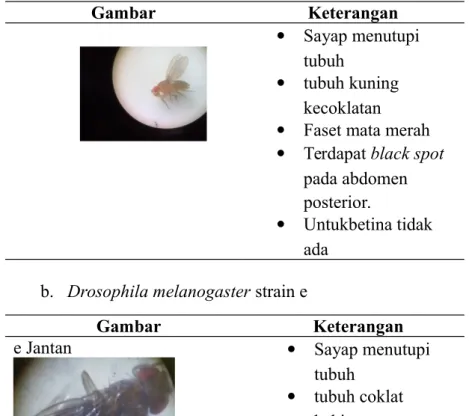
b. Drosophila melanogaster strain e
Gambar Keterangan
e Jantan Sayap menutupi
tubuh
tubuh coklat kehitaman
Faset mata merah
Terdapat black spot pada abdomen posterior
pada betina tidak ada
4.2. Rekontruksi Persilangan
Di bawah ini merupakan rekontruksi persilangan Drosophila melanogaster strain ♀ N >< ♂ e beserta resiproknya. 1. Persilangan ♀ N >< ♂ e
Rekontruksi persilangan yang tidak mengalami nondisjungtion (NDJ)
P1 : ♀ N >< ♂ e Genotip : e+ >< e e+ e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ Gamet : e+ ; e F1
Fenotip yang muncul adalah N ♀, N ♂
Rekontruksi persilangan yang
mengalami nondisjungtion (NDJ) P1 : N ♀ >< e ♂ Genotip : e+ >< e e+ e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ Gamet : e+ , e+ e+ , 0 ; e +¿,0 +¿w¿ +¿, w¿ −¿,¬ , w¿ w¿ F1 ♀ ♂ e + e+ e+ 0 e e + e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ e+ e+ e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ e 0 ♀ ♂ e+ e+ e e+ w−¿ w+¿ ¿ ¿ e (N) e+ w−¿ w+¿ ¿ ¿ e w−¿ w+¿ ¿ ¿ (N) e e+ w−¿ w+¿ ¿ ¿ e w+¿ ¬ ¿ ( N) w+¿ ¬ ¿ e w−¿ w+¿ ¿ ¿ + e (N)
(N) (N super) w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ (e) e e + e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ (N) e+ e+ e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ (N super) e 0 w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ (e) Fenotip yang muncul adalah N, N (super), dan e
2. Persilangan ♂N >< ♀ e
Rekontruksi persilangan yang tidak mengalami nondisjungtion (NDJ) P1 : ♀ N >< ♂ e Genotip : e+ >< e e+ e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ Gamet : e+ ; e F1 19 ♀ e+ e+ e e+ w−¿ w+¿ ¿ ¿ e (N) e+ w−¿ w+¿ ¿ ¿ e w−¿ w+¿ ¿ ¿ (N) e e+ w−¿ w+¿ ¿ ¿ e w+¿ ¬ ¿ w+¿ ¬ ¿ e w−¿ w+¿ ¿ ¿
Rekontruksi persilangan yang mengalami nondisjunction (NDJ) P1: ♂N >< ♀e Genotip : e+ >< e e+ e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ Gamet : e+ , e+ e+ , 0 ; e +¿,0 +¿w¿ +¿, w¿ −¿,¬ , w¿ w¿ F1 ♀ ♂ e+ e+ e+ 0 e e + e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ (N) e+ e+ e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ (N super) e 0 w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ (e) e e + e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ (N) e+ e+ e w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ (N super) e 0 w+¿ w+¿ ¿ w−¿ ¬ >¿ ¿ ¿ (e)
Fenotip yang muncul adalah N, N (super), dan e 4.3 Analisis ANAVA tunggal
Persilangan N♀ × ♂e beserta resiproknya.
Ulangan Total Jenis Kelamin 1 2 3 ♂ 0 10,2 1,75 11,95 ♀ 0 14,2 1,5 15,7 total 27,65 FK=27,65 6 = 764,5225 6 =127,42 Jk total = (0)2 + (10,2) 2 + (1,75) 2 ...- FK 310,9925 – 127,42 = 183,572 JK perlakuan=(11,95)+(15,7) 3 −127,42=¿ 2,344 JK galat=183,572−2,344=181,228 SK db JK KT Fhitung F5% Perlakuan 1 2,344 2,344 0,052 7,71 galat 4 181,228 45,307 total 5 Rujukan,
1. F hitung perlakuan 0,052< F tabel0,05 maka, H0 diterima, dan Hp ditolak. Jadi tidak ada perbedaan frekwensi fenomena gagal berpisah (nondisjunction) persilangan Droshopila melanogaster ♀N × ♂e beserta resiprok pada jenis kelamin yang berbeda.
BAB V PEMBAHASAN
5.1 Perbedaan frekuensi fenomena gagal berpisah (nondisjunction) persilangan Droshopila melanogaster ♀N × ♂e beserta resiprok pada jenis kelamin yang berbeda
Hasil analisis data pada penelitian ini menunjukkan tidak adanya perbedaan yang signifikan frekuensi fenomena gagal berpisah (nondisjunction) antara Droshopila melanogaster strain N jantan dengan betina. penelitian ini sejalan dengan penelitian yang dilakukan oleh Volders et al, (2010) tentang perbedaan respon jenis kelamin terhadap suatu mutagen yang menggunakan model hewan mamalia. Menurut temuan Volders et al (2010) bahwa dari analisis kolam gen (CAs) tidak ditemukan perbedaan utama mengenai jenis kelamin., tidak ada bukti yang jelas mengenai perbedaan respon jenis kelamin terhadap pajanan genotoksik. Penelian ini juga sejalan dengan penelitian yang
dilakukan oleh (Fauzi, 2016) yang sudah menggunakan model hewan Drosophila melanogaster tentang perbandingan jenis kelamin terhadap jumlah anakan pada Drosophila yang telah terpapar radiasi. Banyaknya dari EMF tidak menunjukkan indikasi bahwa EMF berpengaruh terhadap jenis kelamin. Hal ini menunjukkan tidak ada kelangsungan hidup yang bervariasi antara lalat jantan dan lalat betina yang terpapar oleh EMF. Dari hasil ini, dapat dikatakan radiosensitivity dari lalat jantan terhadap EMF memiliki tingkat yang sama dengan lalat betina. Kemungkinan EMF tampaknya tidak menunjukkan bahwa EMF mungkin memberikan pengaruh yang berhubungan dengan jenis kelamin pada lalat. Tetapi hasil ini tidak konsistent dengan beberapa penelitain yang lain yang menggunakan model hewan lain seperti yang telah dilaporkan oleh Bonassi ,et al (1995) yang menyatakan perbedaan antara jenis kelamin ada pada fekwensi pertukaran kromatit, dengan hasil yang lebih tinggi terjadi pada wanita dari pada pria. Hal tersebut juga dipengaruhi oleh keadaan system regulasi hormone yang ada pada kedua jenis kelamin. Dimungkinkan bahwa sistem hormonal mempunyai peranan penting dalam hal proses fisiologi (Alsbeih et al, 2015). Tidak banyak peneliti yang meneliti tentang respon jenis kelamin terhadap suatu mutagen, maka dari itu sangat diperlukan penelitian lebih lajut.
BAB V1 KESIMPULAN 6.1 Kesimpulan
Kesimpulan dari penelitian ini adalah tidak adanya perbedaan yang signifikan frekuensi fenomena gagal berpisah (nondisjunction) antara Droshopila melanogaster strain N jantan dengan betina. Hal ini menunjukkan bahwa tidak ada bukti yang jelas mengenai perbedaan respon jenis kelamin terhadap pajanan genotoksik.
6.2 Saran
1) Peneliti hendaknya mengamati fenotipe dari masing-masing strain terlebih dahulu sebelum melakukan pengamatan.
2) Peneliti harus memanfaatkan waktu seefektif mungkin untuk melakukan pengamatan.
DAFTAR RUJUKAN
Balqis.1995. Pengaruh Suhu Terhadap Frekuensi Gagal Berpisah Sekunder Kromosom Kelamin X antara D.melanogaster. skripsi tidak diterbitkan. Malang: IKIP Malang
Bonassi, S., Bolognesi, C., Abbondandolo, A. et al. (1995) Influence of sex on cytogenetic end points: evidence from a large human sample and review of the literature. Cancer Epidemiol. Biomarkers Prev., 4, 671–679
Corebima, A.D. 2003. Genetika Mendel. Surabaya: Airlangga University Press. Fauzi, A. (2016). The Comparison of Males and Females Number of Drosophila
melanogaster that Exposed by Mobile Phone in Multiple Generations. Makalah disajikan pada seminar Intrernational diselenggarakan Institut Teknologi Sepuluh Nopember (ITS), 15 October 2016
Felix, R. and M.E. de la Rosa.1971. Cytogenetic studies with' sod:lum cyclamate in D. melanogaster femåles. Mexico: Genetics and Radiobiology Program of the National Commission, of Nuclear Energy.
Gardner, Eldon, J. Dan Snustad, D.P.1991. Principle Of Genetics Edition. New York: John Wiley and Sons, Inc.
Indranata, Rudy. 2012. Mekanisme Perbaikan DNA, Mutasi Dan Adaptasi, Aplikasi Praktis Mutasi, Serta Sakit Genetic Menusia Yang Ditimbulkan Oleh Kesalahan Replikasi DNA Dan Perbaikan DNA. (online) (http://rudy-indranatan.blogspot.com/2012/04/mekanisme-perbaikan-dna-mutasi-dan.html, diakses pada tanggal 7 november 2016)