PATOGENISITAS JAMUR ENTOMOPATOGEN
Beauveria bassiana (DEUTEROMYCOTINA: HYPHOMYCETES) TERHADAP TELUR Crocidolomia pavonana (LEPIDOPTERA: PYRALIDAE)
Pathogenicity of Entomopathogenic Fungi Beauveria bassiana (Deuteromycotina:
Hyphomycetes) on Crocidolomia pavonana (Lepidoptera: Pyralidae) egg
Oleh:
Trizelia1), T. Santoso2), S. Sosromarsono2), A. Rauf2) dan L.I. Sudirman3)
1)
Jurusan Hama dan Penyakit Tumbuhan Faperta, Universitas Andalas
2)
Departemen Proteksi Tanaman, Faperta, Institut Pertanian Bogor
3)
Departemen Biologi, FMIPA, Institut Pertanian Bogor
ABSTRAK
Penelitian ini bertujuan untuk mempelajari patogenisitas jamur entomopatogen Beauveria bassiana (Bals.) Vuill. (Deuteromycotina: Hyphomycetes) yang diisolasi dari berbagai lokasi dan jenis inang di Indonesia terhadap telur Crocidolomia pavonana (F.) (Lepidoptera: Pyralidae). Pada percobaan ini digunakan kelompok telur dan 13 isolat B. bassiana, yaitu La1, La2, La3, La4, Hh1, Bb-Hh2, Bb-Cp, Bb-Thr, Bb-Sl, Bb-Nl, Bb-725, Bb-Cc, Bb-Rl. Aplikasi jamur dilakukan dengan cara perendaman kelompok telur C. Pavonana ke dalam suspensi konidia selama 60 detik. Hasil penelitian menunjukkan bahwa tidak ada pengaruh aplikasi seluruh isolat B. bassiana terhadap mortalitas telur, akan tetapi beberapa larva instar I mati 3 hari setelah keluar dari telur. Mortalitas larva instar I maksimal 91.00%. Kata kunci: Beauveria bassiana, patogenesitas, isolat, telur Crocidolomia pavonana
ABSTRACT
The purpose of this research was to study the pathogenicity of Beauveria bassiana (Bals.) Vuill. (Deuteromycotina: Hyphomycetes) originated from different geographical locations and host ranges in Indonesia to eggs of Crocidolomia pavonana (F.) (Lepidoptera: Pyralidae). Egg clusters and thirteen isolates, i.e., Bb-La1, Bb-La2, Bb-La3, Bb-La4, Bb-Hh1, Bb-Hh2, Bb-Cp, Bb-Thr, Bb-Sl, Bb-Nl, Bb-725, Bb-Cc, Bb-Rl were used in the experiment. Egg cluster directly inoculated with conidial suspensions using dipping methods in 60 seconds. The results of experiment showed that there was no effect of all isolates on egg mortality, but that some first instar larvae died 3 days after eclosion. The maximum mortality of first instar larvae was 91.00%.
Key words: Beauveria bassiana, pathogenicity, isolates, Crocidolomia pavonana eggs
PENDAHULUAN
Di Indonesia, Crocidolomia
pavonana (F.) (Lepidoptera: Pyralidae) merupakan salah satu jenis hama yang
menimbulkan masalah penting pada pertanaman sayuran Brassicaceae, seperti kubis, brokoli, kubis bunga, sawi dan lobak (Kalshoven, 1981; Shepard et al., 1997). Kerusakan ditimbulkannya dengan
cara memakan daun, terutama daun yang masih muda dan menuju ke bagian titik tumbuh, sehingga titik tumbuh habis dan tanaman dapat mati (Kalshoven ,1981).
Pada saat ini, pengendalian hama C. pavonana masih sangat tergantung kepada pestisida sintetis, karena cara ini mudah dilaksanakan, cepat menurunkan populasi hama dan belum ditemukan pilihan pengendalian lainnya yang cukup efektif. Aplikasi pestisida dilakukan secara intensif, seminggu sekali, bahkan 2-3 hari sekali (Rauf, 1996).
Kadang-kadang petani masih melakukan
penyemprotan pada tanaman yang siap
dipanen tanpa memperhatikan
dampaknya terhadap konsumen.
Penggunaan insektisida kimia yang
sangat intensif ini dapat mengganggu
kehidupan bahkan mematikan
sumberdaya alam hayati dan mencemari
lingkungan hidup. Hal ini sangat
disayangkan mengingat Indonesia sedang menuju era pembangunan pertanian yang
berwawasan lingkungan, sehingga
penggunaan pestisida kimia sintetis harus digunakan seminimum mungkin.
Untuk menjawab dilema tersebut,
konsep pengendalian hama terpadu
(PHT) merupakan pilihan yang tepat,
karena PHT bertujuan membatasi
penggunaan pestisida sesedikit mungkin tetapi sasaran kualitas dan kuantitas produksi pertanian masih dapat dicapai
(Sastrosiswojo dan Oka, 1997).
Pengurangan masukan pestisida sekaligus juga akan menurunkan residu pestisida, sehingga produk yang dihasilkan dapat lebih bersaing di pasar.
Didalam PHT, pemberdayaan
musuh alami dan potensi hayati lainnya merupakan komponen utama, karena musuh alami mempunyai peranan penting dalam penekanan populasi hama dan
menjaga keseimbangan ekosistem. Oleh karena itu, musuh alami yang sudah ada perlu dijaga kelestariannya dan upaya untuk meningkatkan peranannya dalam pengendalian hama juga perlu dilakukan. Di antara musuh alami yang dapat digunakan untuk pengendalian hama C. pavonana secara hayati adalah jamur
entomopatogen Beauveria bassiana
(Bals.) Vuill. (Deuteromycotina:
Hyphomycetes). Pemanfaatan B.
bassiana untuk pengendalian hama C. pavonana belum banyak dilaporkan.
Hasil penelitian pendahuluan
menunjukkan bahwa B. bassiana yang
diisolasi dari hama Hypothenemus
hampei (Ferr.) (Coleoptera: Scolytidae) efektif dalam mengendalikan hama C. pavonana (Trizelia dan Arneti, 1996; Trizelia, 1997), tetapi tidak efektif dalam mengendalikan Spodoptera litura (F.) (Lepidoptera: Noctuidae) (Yunisman et al., 1997).
Salah satu keuntungan penggunaan jamur B. bassiana untuk pengendalian hayati adalah dapat digunakan untuk
mengendalikan berbagai tingkat
perkembangan serangga hama mulai dari telur, larva, pupa dan imago. Hasil penelitian Prayogo (2004) menunjukkan bahwa B. bassiana dapat menginfeksi
telur Riptortus linearis (Linn.)
(Hemiptera: Alydidae) sehingga jumlah nimfa yang terbentuk rendah. Hal ini berarti bahwa kematian serangga hama yang terjadi pada stadium awal ini
sangat menguntungkan dalam
pengendalian karena populasi serangga hama dapat ditekan lebih dini.
Penelitian ini bertujuan untuk mempelajari patogenisitas B. bassiana terhadap telur C. pavonana.
METODE PENELITIAN
Percobaan ini dilaksanakan di Laboratorium Patologi Serangga Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian IPB Bogor dari bulan April 2003 sampai dengan Juli 2003.
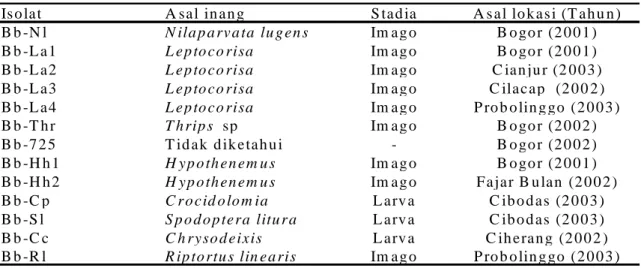
A. Koleksi dan Perbanyakan Isolat Isolat B. bassiana yang digunakan dalam penelitian ini merupakan koleksi Laboratorium Patologi Serangga Jurusan
HPT IPB, Balitro Bogor, koleksi
langsung dari serangga yang terinfeksi di lapangan dari lokasi yang berbeda (Tabel 1). Seluruh isolat ditumbuhkan pada
medium Sabauraud dextrose agar
(SDAY) (dekstrosa 10 g, pepton 2.5 g, ekstrak khamir 2.5 g, agar 20 g, kloramfenikol 0.5 g dan akuades 1 l) (Samuels et al., 2002).
Isolasi B. bassiana dari serangga yang telah terinfeksi oleh B. bassiana
dilakukan dengan cara mengambil
miselium atau konidiumnya dan
ditumbuhkan pada medium SDAY, kemudian dilakukan pemurnian pada
media yang sama. Semua isolat
diinkubasikan pada suhu 25C selama 15 hari. Untuk mempertahankan kevirulenan dari isolat yang diuji, semua isolat diinokulasikan kembali pada larva C. pavonana. Dari larva C. pavonana yang
terinfeksi diisolasi kembali dan
dimurnikan pada media SDAY. B. Penyediaan Tanaman Kubis
Tanaman kubis yang digunakan sebagai pakan larva C. pavonana ditanam dalam polibeg kapasitas 5 kg. Benih kubis yang digunakan adalah Cabbage F1 Asia Cross merek Tropica. Benih
dibibitkan dalam kotak persemaian
sampai berumur satu bulan dan
kemudian dipindahkan ke dalam polibeg yang berisi campuran tanah dan pupuk kandang. Tanaman dipupuk dengan pupuk dasar NPK (15:15:15) seminggu setelah tanam sebanyak 0.5 g /polibeg. Pemupukan dilakukan kembali pada waktu tanaman telah berumur satu dan dua bulan. Tanaman disiram setiap hari dan tanaman tidak disemprot dengan pestisida.
Tabel 1. Sumber isolat B. bassiana yang digunakan
Is o la t A sa l in a n g S ta d ia A s a l lo k a s i (T a h u n ) B b -N l N ila p a rv a ta lu g e n s Im a g o B o go r (2 0 0 1 ) B b -L a 1 L e p to c o risa Im a g o B o go r (2 0 0 1 ) B b -L a 2 L e p to c o risa Im a g o C ia n ju r (2 0 0 3 ) B b -L a 3 L e p to c o risa Im a g o C ila c a p (2 0 0 2 ) B b -L a 4 L e p to c o risa Im a g o P ro b o lin g go (2 0 0 3 ) B b -T h r T h rip s sp Im a g o B o go r (2 0 0 2 ) B b -7 2 5 T id a k d ik e ta h u i - B o go r (2 0 0 2 ) B b -H h 1 H y p o th e n e m u s Im a g o B o go r (2 0 0 1 ) B b -H h 2 H y p o th e n e m u s Im a g o F a ja r B u la n (2 0 0 2 ) B b -C p C r o c id o lo m ia L a rv a C ib o d a s (2 0 0 3 ) B b -S l S p o d o p te ra litu r a L a rv a C ib o d a s (2 0 0 3 ) B b -C c C h ry so d e ix is L a rv a C ih e ra n g (2 0 0 2 ) B b -R l R ip to rtu s lin e a ris Im a g o P ro b o lin g go (2 0 0 3 )
C. Perbanyakan C. pavonana
Hama C. pavonana dikumpulkan dari pertanaman kubis di lapangan. Larva dipelihara dalam kotak plastik dan diberi makanan berupa daun kubis yang masih segar. Makanan larva diganti setelah habis atau sudah tidak segar lagi.
Pada waktu larva akan berpupa, di dasar kotak diberi serbuk gergaji. Semua imago yang keluar dari pupa dipelihara secara massal dalam kurungan serangga yang telah diberi daun kubis segar sebagai tempat peletakan telur. Sebagai makanan imago digunakan madu dengan konsentrasi 10%. Kelompok telur yang diletakkan pada daun kubis digunakan untuk pengujian.
D. Penyiapan Suspensi Konidium
Seluruh isolat diperbanyak pada medium SDAY dalam cawan Petri pada suhu 25 o C selama 15 hari. Konidium
jamur dipanen dengan cara
menambahkan 5 ml akuades steril dan 0.05% Triton X100 sebagai bahan perata ke dalam cawan Petri dan konidia dilepas dari medium dengan kuas halus. Suspensi disaring dan konsentrasi konidia dihitung menggunakan hemositometer. E. Aplikasi Konidium B. bassiana
Terhadap Telur C. pavonana
Konsentrasi konidium
masing-masing isolat yang digunakan untuk pengujian patogenisitas B. bassiana terhadap stadia telur C. pavonana adalah 108 konidium/ml.
Telur uji terdiri atas dua kelompok telur C. pavonana yang diletakkan pada daun kubis dan berumur satu hari. Perlakuan berupa perendaman kelompok telur dalam suspensi konidium selama 60
detik, kemudian kelompok telur
dipindahkan ke dalam cawan Petri yang telah dialas dengan kertas saring lembap. Kelompok telur yang telah diperlakukan dipelihara sampai menetas. Percobaan diulang tiga kali dan percobaan disusun dalam rancangan acak lengkap (RAL) dengan 13 taraf perlakuan (isolat). Data hasil percobaan diolah dengan sidik ragam dan dilanjutkan dengan pengujian nilai tengah menggunakan uji Tukey (HSD) pada taraf nyata 5%. Variabel pengamatan adalah persentase telur yang menetas dan persentase larva instar I yang terinfeksi. Data dianalisis dengan
menggunakan program Statistical
Analysis System (SAS) versi 6.12.
HASIL DAN PEMBAHASAN
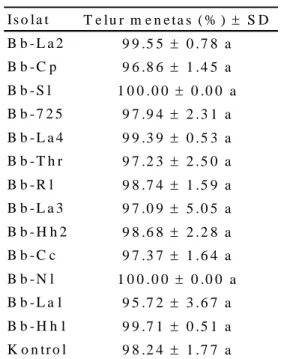
Hasil uji patogenisitas B. bassiana terhadap telur C. pavonana menunjukkan bahwa seluruh isolat B. bassiana yang diuji tidak memengaruhi perkembangan telur C. pavonana. Analisis statistika menunjukkan bahwa tidak ada perbedaan yang nyata antara telur C. pavonana yang diaplikasi dengan suspensi konidium B. bassiana dengan kontrol (F=1.11; db=13, 28; P<0.3885). Hampir 100% telur C. pavonana masih mampu menetas walaupun telah diberi perlakuan dengan suspensi konidium B. bassiana (Tabel 2). Semua isolat yang diuji tidak dapat menginfeksi telur C. pavonana dan telur menetas secara normal.
Hasil pengamatan makroskopis
menunjukkan bahwa telur C. pavonana yang telah diaplikasi dengan B. bassiana tidak memperlihatkan adanya gejala infeksi B. bassiana. Hal ini menunjukkan
bahwa B. bassiana tidak bisa
menginfeksi telur C. pavonana. Tidak terjadinya infeksi pada telur mungkin
disebabkan oleh sifat fisik dan kimia telur C. pavonana yang diduga menghalangi tabung kecambah konidium menembus ke bagian dalam telur.
Tabel 2. Rata-rata persentase telur C. pavonana menetas setelah aplikasi berbagai isolat B. bassiana Is o l a t T e l u r m e n e t a s ( % ) S D B b - L a 2 9 9 .5 5 0 . 7 8 a B b - C p 9 6 .8 6 1 . 4 5 a B b - S l 1 0 0 . 0 0 0 .0 0 a B b - 7 2 5 9 7 .9 4 2 . 3 1 a B b - L a 4 9 9 .3 9 0 . 5 3 a B b - T h r 9 7 .2 3 2 . 5 0 a B b - R l 9 8 .7 4 1 . 5 9 a B b - L a 3 9 7 .0 9 5 . 0 5 a B b - H h 2 9 8 .6 8 2 . 2 8 a B b - C c 9 7 .3 7 1 . 6 4 a B b - N l 1 0 0 . 0 0 0 .0 0 a B b - L a 1 9 5 .7 2 3 . 6 7 a B b - H h 1 9 9 .7 1 0 . 5 1 a K o n t r o l 9 8 .2 4 1 . 7 7 a
Keterangan: Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata menurut uji Tukey (HSD) pada taraf nyata 5%.
Menurut Inglis et al. (2001)
kemampuan patogen untuk bisa
menimbulkan infeksi pada serangga ditentukan oleh tiga faktor yaitu patogen, inang dan lingkungan. Dari segi patogen,
dosis dan cara aplikasi akan
mempengaruhi mortalitas serangga,
sedangkan dari segi inang, berbagai faktor fisiologi dan morfologi inang
mempengaruhi kerentanan serangga
terhadap jamur entomopatogen.
Dos Santos dan Gregorio (2003) mengemukakan bahwa lapisan kulit telur famili Pyralidae tersusun oleh membran vitellin, korion dan lapisan lilin. Lapisan
ini mempunyai banyak fungsi di
antaranya melindungi embrio dari
serangan mikroba (Margaritis 1985, dalam Dos Santos dan Gregorio, 2003). James et al. (2003) mengemukakan bahwa lapisan lilin pada serangga dapat menghambat perkecambahan konidia B. bassiana. Selanjutnya Smith dan Grula (1982) mengemukakan bahwa beberapa jenis asam lemak seperti caprylic acid yang ada pada permukaan tubuh serangga
dapat menghambat perkecambahan
konidium B. bassiana.
James et al. (2003) juga
melaporkan bahwa telur Bemisia
argentifolii Bellows & Perring (Homoptera: Aleyrodidae) tahan terhadap infeksi B. Bassiana. Hasil pengamatan mikroskop elektron menunjukkan bahwa hanya 13.0% konidium B. bassiana yang berkecambah pada telur. Di alam juga belum pernah dilaporkan adanya jamur entomopatogen yang menginfeksi telur B. argentifolii (Lacey et al. 1996, dalam James et al. 2003).
Walaupun aplikasi B. bassiana tidak memengaruhi persentase telur yang menetas, akan tetapi aplikasi B. bassiana pada telur berpengaruh nyata terhadap kelangsungan hidup larva instar I ( F=195.04; db= 13, 28; P<0.0001). Aplikasi konidium B. bassiana pada
konsentrasi 108 konidium per ml
menghasilkan mortalitas larva instar I maksimum sebesar 91.223 % (Bb-La2) dan minimum 5.537 % (Bb-Hh1). Isolat Bb-Hh1 dianggap tidak bersifat patogen terhadap larva instar I yang baru keluar dari telur, karena persentase mortalitas
larvanya tidak berbeda nyata dengan kontrol (Tabel 3).
Terjadinya kematian pada larva instar I disebabkan oleh larva yang baru keluar dari telur memakan kulit telur dan diduga konidium yang menempel pada kulit telur juga termakan oleh larva dan infeksi terjadi melalui saluran pencernaan (Gambar 5.1). Hasil penelitian Broome et al. (1976) serta Kramm dan West (1982) menunjukkan bahwa selain melalui integumen, infeksi B. bassiana pada serangga juga dapat melalui saluran pencernaan. Infeksi B. bassiana pada larva C. pavonana instar I dapat melalui kontak antara konidium yang ada pada kulit telur dengan bagian ventral tubuh larva, kaki dan alat mulut sewaktu larva keluar dari kulit telur. Keberhasilan proses infeksi sangat dipengaruhi oleh kemampuan konidium dari masing-masing isolat bertahan pada permukaan kulit telur.
Tabel 3. Rerata persentase mortalitas larva instar I C. pavonana setelah aplikasi berbagai isolat B. Bassiana
Isolat Mortalitas larva instar I
(%) SD Bb-La2 91.22 1.18 a Bb-Cp 88.25 3.05 a Bb-Sl 85.00 9.26 a Bb-725 84.08 4.47 a Bb-La4 83.43 2.89 a Bb-Thr 83.10 2.07 a Bb-Rl 80.49 2.33 a Bb-La3 59.21 4.65 b Bb-Hh2 48.27 5.56 bc Bb-Cc 40.44 6.61 c Bb-Nl 17.51 2.16 d Bb-La1 16.00 2.19 d Bb-Hh1 5.54 2.68 de Kontrol 1.19 0.55 e
Ketarangan: Angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata menurut uji Tukey (HSD) pada taraf nyata 5%.
Isolat Bb-La2, Bb-Cp, Bb-Sl, Bb725, Bb-La4, Bb-Thr, dan Bb-Rl merupakan isolat yang sangat virulen, karena mampu menyebabkan kematian pada larva instar I lebih dari 80 %. Long et al. (1998) juga melaporkan bahwa
aplikasi B. bassiana pada telur
Leptinotarsa decemlineata (Say)
(Coleoptera: Chrysomelidae) tidak
berpengaruh terhadap mortalitas telur,
tetapi berpengaruh nyata terhadap
mortalitas larva instar I yang baru keluar dari telur.
Hasil penelitian ini berbeda dengan hasil penelitian yang dilaporkan oleh
beberapa peneliti lain yang
mengemukakan bahwa B. bassiana dapat
menginfeksi telur serangga. Pada
konsentrasi 5x108 konidium/ml,
mortalitas telur Neochetina bruchi Hustache (Coleoptera: Curculionidae) akibat infeksi B.bassiana pada hari ke-12 adalah 54.8%. Nilai LC50 B. bassiana
pada stadium telur adalah 9.5 x 107 konidium/ml. Stadium telur merupakan stadium yang paling tahan dibandingkan dengan stadium larva (Chikwenhere dan Vestergaard, 2001). Pada telur Blissus antillus (Leonard) (Hemiptera: Lygaeidae), mortalitas telur akibat infeksi B. bassiana bervariasi antara isolat.
Isolat B. bassiana CG24 mampu
menimbulkan infeksi telur sebesar 43.3% sedangkan isolat CG04 dan ARSEF792 hanya 7.8%. Hasil pengamatan dengan mikroskop pendar menunjukkan bahwa
adanya perbedaan kevirulenan ini
disebabkan adanya perbedaan
kemampuan konidia jamur menempel pada permukaan telur dan selanjutnya menembus korion (Samuels et al., 2002). Selanjutnya Prayogo (2004) melaporkan bahwa lima jenis jamur entomopatogen, yaitu Nomuraea rileyi (Farl.) Sams,
Verticillium lecanii (Zimmermann), Metarhizium anisopliae (Metschn.)
Sorokin, Beauveria bassiana, dan
Paecilomyces fumosoroseus (Wize) Brown and Smith) mampu menginfeksi telur Riptortus linearis (L.) (Hemiptera: Alydidae), sehingga persentase telur yang menetas menjadi nimfa sangat rendah.
DAFTAR PUSTAKA
Broome J.R, Sikorowski P.P, Norment B.R. 1976. A mechanism of pathogenicity of B. bassiana on larvae of the imported fire ant Solenopsis richteri. J. Invertebr. Pathol. (28): 87-91.
Chikwenhere G.P, Vestergaard S. 2001. Potential effects of Beauveria bassiana (Balsamo) Vuillemin on Neochetina bruchi Hustache (Coleoptera: Curculionidae), a biological control agent of water hyacinth. Biol.Contr. (21): 105-110. Dos Santos D.C, Gregorio E.A. 2003.
Deposition of the eggshell layers in the sugarcane borer (Lepidoptera: Pyralidae): ultrastructural aspects. Acta microscopica 12(1): 37-41. Inglis G.D, Goettel M.S, Butt T.M,
Strasser H. 2001. Use of
hyphomycetous fungi for managing insect pests. in: Butt T.M, Jackson C.W dan Magan N. (Eds). Fungi as Biocontrol Agents, Progress, Problems and Potential. CABI Publishing, London.
James R.R, Buckner J.S, Freeman T.P. 2003. Cuticular lipids and silverleaf whitefly stage affect conidial germination of Beauveria bassiana
and Paecilomyces fumosoroseus. J. Invertebr. Pathol. (84): 67-74. Kalshoven, L.G.E. 1981. The Pests of
Crops in Indonesia. Laan PA van der. penerjemah. Jakarta: Ichtiar Baru-Van Hoeve. Revisi dari : De Plagen van de Cultuurgewassen in Indonesie.
Kramm KR, West DF. 1982. Termite pathogens: effects of ingested Metarhizium, beauveria and Gliocladium conidia on worker termites (Reticulitermes sp.). J. Invertebr. Pathol. (40): 7-11. Long D.W, Drummond F.A, Groden E.,
1998. Susceptibility of Colorado
potato beetle (Leptinotarsa
decemlineata) eggs to Beauveria bassiana. J. Invertebr. Pathol. (71): 182-183.
Prayogo Y. 2004. Keefektifan Lima Jenis Cendawan Entomopatogen
untuk Mengendalikan Hama
Pengisap Polong Kedelai. Riptortus linearis L. (Hemiptera: Alydidae) dan Dampaknya Terhadap Predator Oxyopes javanus (Araneidae: Oxyopidae). Tesis. IPB, Bogor Rauf, A. 1996. PHT mereguk manfaat
dari globalisasi pasar. Disampaikan
dalam Seminar dan Rapat
Koordinasi Wilayah II. Himpunan Mahasiswa Perlindungan Tanaman Indonesia, 22-24 Desember 1996. Samuels R.I, Coracini D.L.A, dos Santos
C.A.M, Gava C.A.T. 2002.
Infection of Blissus antillus (Hemiptera: Lygaeidae) eggs by
entomopathogenic fungi M.
anisopliae and B. bassiana. Biol. Contr. (23): 269-273.
Sastrosiswodjo S, Oka I.N. 1997. Implementasi pengelolaan serangga secara berkelanjutan. Makalah disajikan pada Kongres ke V dan Simposium Entomologi. PEI. 24-26 Juni 1997, Bandung.
Shepard M, Shepard EF, Carner GR, Hammig MD, Rauf A, Turnipseed SG dan Samsudin. 1997. Prospect for IPM in secondary food crops. Makalah disajikan pada Kongres V dan Simposium Entomologi, PEI, 24-26 Juni 1997, Bandung.
Smith RJ, Grula E.A. 1982. Toxic components on the larval surface of the Corn Earworm (Heliothis zea) and their effects on germination and growth of Beauveria bassiana. J. Invertebr. Pathol. (39):15-22.
Trizelia, A. 1996. Kemampuan Jamur Beauveria bassiana untuk Pengendalian Hama Crocidolomia binotalis Z. pada Tanaman Kubis. Laporan Penelitian. Lembaga Penelitian Unand, Padang.
Trizelia, A. 1997. Pengaruh Infeksi Beauveria bassiana terhadap
Biologi Hama Crocidolomia
binotalis Z. pada Tanaman Kubis. Laporan Penelitian. Lembaga Penelitian Unand, Padang
Yunisman, Rusli R, Busnia M,
Yaherwandi, Kiman Z.B. 1997. Patogenisitas Cendawan Beauveria bassiana terhadap Hama Perusak Daun Kubis, Spodoptera Litura F. (Lepidoptera: Noctuidae). Laporan Penelitian. Lembaga Penelitian Unand, Padang.