HASIL
Posisi Geografis dan Administratif Lokasi Penelitian
Pengambilan sampel dilakukan pada empat stasiun yaitu: Desa Mandiangin Tebet (Mandiangin), Desa Sungai Bengkal (S. Bengkal), Desa Pelayangan dan Desa Simpang. Desa Mandiangin terletak pada ketinggian 29 mdpl dengan kordinat 02.01.923 LS dan 102.57.893 BT. Secara administratif Desa Mandiangin termasuk dalam Kecamatan Mandiangin, Kabupaten Sarolangun. Desa S. Bengkal terletak pada ketinggian 30 mdpl dengan kordinat 1.32.598 LS dan 102.43.232 BT. Secara administratif Desa S. Bengkal termasuk dalam Kecamatan Tebo Ilir, Kabupaten Muara Tebo. Desa Pelayangan terletak pada ketinggian 15 mdpl dengan kordinat 1.40.965 LS dan 103.5.851 BT. Secara administratif Desa Pelayangan termasuk dalam Kecamatan Muara Tembesi, Kabupaten Batang Hari. Desa Simpang terletak pada ketinggian 7 mdpl dengan kordinat 1.15.871 LS dan 104.05.550 BT. Secara administratif Desa Simpang termasuk dalam Kecamatan Berbak, Kabupaten Tanjung Jabung Timur.
Kondisi Perairan Lokasi Pengambilan Sampel
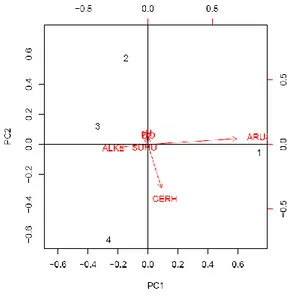
Analisis data parameter fisika-kimia lokasi pengambilan sampel dilakukan dengan metode Principal Component Analysis (PCA). Hasil analisis menunjukkan ke-empat stasiun pengambilan sampel terbagi dalam tiga kelompok, yaitu Mandiangin, Simpang dan gabungan S. Bengkal dengan Pelayangan (Gambar 2). Lokasi Mandiangin dicirikan dengan kecepatan arus dan kecerahan yang tinggi. Lokasi Simpang dicirikan dengan kecerahan yang tinggi. Tidak ada parameter fisika-kimia perairan yang dominan di lokasi S. Bengkal dan Pelayangan. Data parameter fisika-kimia lokasi pengambilan sampel ditampilkan pada lampiran 5.
Berdasarkan plotting data parameter fisika-kimia perairan dilakukan uji keterkaitan (korelasi) untuk melihat hubungan antarkarakter. Hasil uji korelasi Pearson menunjukkan bahwa kecepatan arus berbanding terbalik dengan tingkat alkalinitas (P = 0.02685), sedangkan pH berbanding terbalik dengan kecerahan (P= 0.02435). Korelasi antarkarakter lain seperti suhu dan DO tidak menunjukkan nilai korelasi yang signifikan.
Gambar 2 Biplot PCA parameter fisika-kimia perairan lokasi pengambilan sampel, 1=Mandiangin, 2=S. Bengkal, 3=Pelayangan, 4=Simpang.
Hasil uji t taraf 95% terhadap parameter fisika-kima perairan lokasi pengambilan sampel (Lampiran 6) menunjukkan keempat stasiun tidak memiliki parameter fisika-kimia yang berbeda nyata. Hal yang sama juga ditemukan ketika taraf uji t dinaikkan hingga 99%. Tidak ada lokasi yang memiliki parameter fisika-kimia yang berbeda nyata.
Keragaman Jenis dan Kelimpahan Ikan Lais di Setiap Lokasi Pengambilan Sampel
Ikan lais yang ditemukan dalam penelitian ini ada 4 jenis yaitu Ompok hypopthalmus, Kryptopterus micronema, K. limpok, dan K. bichirris (Tabel 3). Pengamatan selama periode pengambilan sampel menunjukkan bahwa O. hypopthalmus dan K. limpok merupakan jenis yang paling mudah ditemukan. Kedua jenis ini konsisten ditemukan setiap periode pengambilan sampel, yang berarti bahwa O. hypopthalmus dan K. limpok dapat ditemukan sepanjang tahun di DAS Batang hari. Diantara kedua spesies ini, O. hypopthalmus lebih sering ditemukan dan dengan jumlah individu tangkapan yang lebih banyak. Kryptopterus micronema hanya ditemukan dua kali dari tiga kali periode pengambilan sampel, yaitu saat air Sungai Batang Hari mulai naik (awal musim penghujan) dan saat air Sungai Batang Hari mulai surut (akhir musim penghujan). Kryptopterus bichirris hanya ditemukan saat air Sungai Batang Hari mulai surut
(pengambilan sampel ketiga), dan hanya ditemukan di stasiun Mandiangin dan Pelayangan.
Tabel 3 Jenis dan jumlah ikan lais yang ditemukan setiap periode pengambilan sampel
Nama jenis Periode pengambilan sampel Jumlah
Agustus 2009 Desember 2009 April 2010
A B C D A B C D A B C D O. hypopthalmus * - * - * - * * * * * * 40 K. limpok 3 * - - - * - - * * * * 40 K. micronema - - - 3 - 2 * * * * 40 K. bichirris - - - 4 - * - 14 Jumlah 134
*=Ditemukan dan dikoleksi dengan jumlah sampel maksimal 10 individu/jenis/lokasi, angka =hanya ditemukan sebanyak angka tersebut, -= tidak ditemukan, A= Mandiangin, B= S. Bengkal, C= Pelayangan, D= Simpang.
Berdasarkan periode pengambilan sampel ditemukan bahwa ikan lais dominan ditemukan saat akhir musim penghujan atau saat air Sungai Batang Hari mulai surut (April-Mei). Saat akhir musim penghujan disebut juga dengan musim puncak tangkapan ikan. Pada tahun 2010, musim puncak tangkapan ikan berlangsung pada akhir April hingga awal Mei. Menurut informasi nelayan di DAS Batang Hari, musim puncak tangkapan ikan pada tahun 2009 berlangsung satu bulan lebih cepat, yaitu pada pertengahan Maret. Musim puncak tangkapan ikan di daerah hulu (Mandiangin dan S. Bengkal) berlangsung lebih awal (lebih cepat) sekitar dua minggu dibandingkan dengan daerah hilir (Simpang).
Keragaman Struktur Morfologis
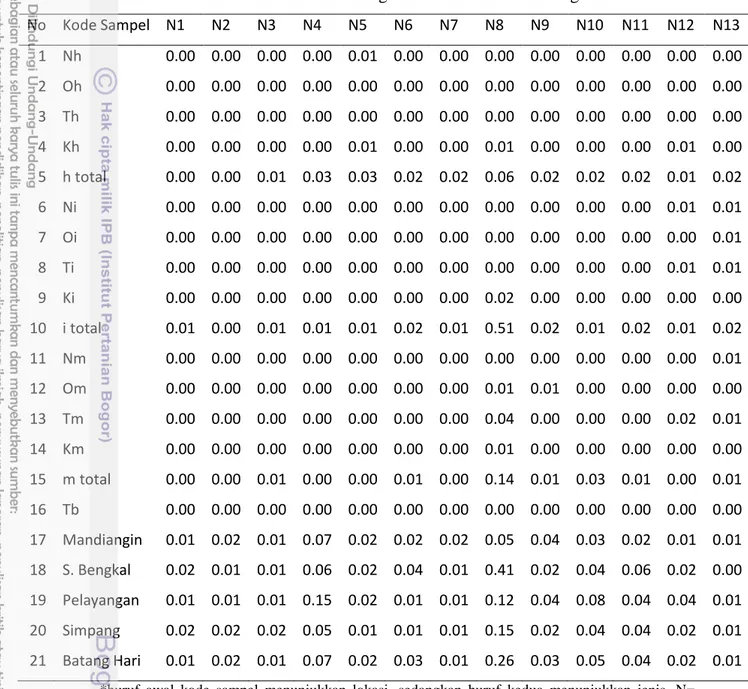
Pengukuran proporsi ukuran tubuh yang dikategorikan sebagai data nisbah morfometrik menunjukkan keragaman yang rendah pada intra dan antarpopulasi ikan lais di DAS Batang Hari. Nilai keragaman (varians) intra populasi pada 12 indeks struktur morfologis hanya berkisar 0.00–0.01 (χ = 0.13–1.20). Nilai varians antarpopulasi berkisar 0.01-0.51 (χ = 0.15–1.12) sedangkan varians antarspesies
pada seluruh sampel ikan lais dari DAS Batang Hari berkisar 0.01–0.26 (χ = 0.17– 1.01) (Tabel 4).
Tabel 4 Nilai varians struktur morfologis ikan lais di DAS Batang Hari
No Kode Sampel N1 N2 N3 N4 N5 N6 N7 N8 N9 N10 N11 N12 N13 1 Nh 0.00 0.00 0.00 0.00 0.01 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 2 Oh 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 3 Th 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 4 Kh 0.00 0.00 0.00 0.00 0.01 0.00 0.00 0.01 0.00 0.00 0.00 0.01 0.00 5 h total 0.00 0.00 0.01 0.03 0.03 0.02 0.02 0.06 0.02 0.02 0.02 0.01 0.02 6 Ni 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 0.01 7 Oi 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 8 Ti 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 0.01 9 Ki 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.02 0.00 0.00 0.00 0.00 0.00 10 i total 0.01 0.00 0.01 0.01 0.01 0.02 0.01 0.51 0.02 0.01 0.02 0.01 0.02 11 Nm 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 12 Om 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 0.01 0.00 0.00 0.00 0.00 13 Tm 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.04 0.00 0.00 0.00 0.02 0.01 14 Km 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.01 0.00 0.00 0.00 0.00 0.00 15 m total 0.00 0.00 0.01 0.00 0.00 0.01 0.00 0.14 0.01 0.03 0.01 0.00 0.01 16 Tb 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 0.00 17 Mandiangin 0.01 0.02 0.01 0.07 0.02 0.02 0.02 0.05 0.04 0.03 0.02 0.01 0.01 18 S. Bengkal 0.02 0.01 0.01 0.06 0.02 0.04 0.01 0.41 0.02 0.04 0.06 0.02 0.00 19 Pelayangan 0.01 0.01 0.01 0.15 0.02 0.01 0.01 0.12 0.04 0.08 0.04 0.04 0.01 20 Simpang 0.02 0.02 0.02 0.05 0.01 0.01 0.01 0.15 0.02 0.04 0.04 0.02 0.01 21 Batang Hari 0.01 0.02 0.01 0.07 0.02 0.03 0.01 0.26 0.03 0.05 0.04 0.02 0.01
*huruf awal kode sampel menunjukkan lokasi, sedangkan huruf kedua menunjukkan jenis. N= mandiangin, O= S. Bengkal, T= Pelayangan, K= Simpang, h = O. hypopthalmus, i = K. limpok, m = K. micronema, b = K. bichirris.
Indeks N8 (tinggi moncong/tinggi pangkal kepala), N4 (tinggi ekor/panjang baku), dan N10 (tinggi ekor/tinggi badan) memiliki varian tertinggi. Indeks N1 (panjang kepala/panjang baku), N3 (tinggi badan/panjang baku), dan N7 (panjang
hidung/panjang kepala) memiliki varian terendah. Hal ini berarti struktur tinggi merupakan karakter yang paling bervariasi diantara sampel ikan lais yang didapatkan, sedangkan karakter panjang kepala dan panjang baku relatif tidak berbeda. Berdasarkan nilai varian dapat dinyatakan bahwa struktur morfologis antarlokasi dan antarspesies ikan lais di DAS Batang Hari tidak beragam. Data lengkap proporsi ukuran tubuh (nisbah morfometrik) seluruh sampel ditampilkan pada lampiran 7.
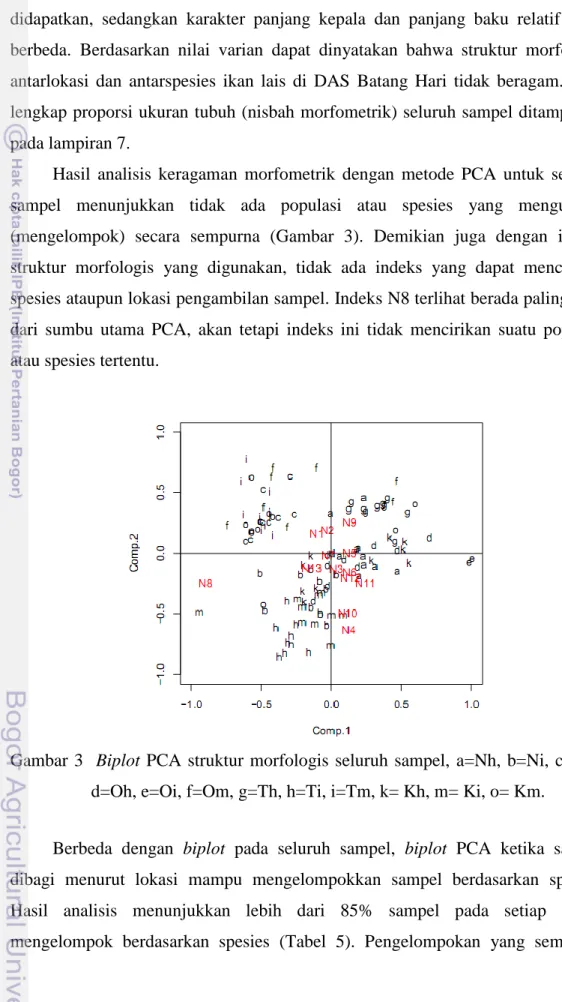
Hasil analisis keragaman morfometrik dengan metode PCA untuk seluruh sampel menunjukkan tidak ada populasi atau spesies yang mengumpul (mengelompok) secara sempurna (Gambar 3). Demikian juga dengan indeks struktur morfologis yang digunakan, tidak ada indeks yang dapat mencirikan spesies ataupun lokasi pengambilan sampel. Indeks N8 terlihat berada paling jauh dari sumbu utama PCA, akan tetapi indeks ini tidak mencirikan suatu populasi atau spesies tertentu.
Gambar 3 Biplot PCA struktur morfologis seluruh sampel, a=Nh, b=Ni, c=Nm, d=Oh, e=Oi, f=Om, g=Th, h=Ti, i=Tm, k= Kh, m= Ki, o= Km.
Berbeda dengan biplot pada seluruh sampel, biplot PCA ketika sampel dibagi menurut lokasi mampu mengelompokkan sampel berdasarkan spesies. Hasil analisis menunjukkan lebih dari 85% sampel pada setiap lokasi mengelompok berdasarkan spesies (Tabel 5). Pengelompokan yang sempurna
(100%) ditemukan pada sampel ikan lais yang berasal dari Pelayangan (Gambar 4). Individu ke-8 dari spesies K. micronema (nomor 28) walaupun terlihat terpisah dari kumpulan individu lain, akan tetapi masih berada dalam kuadran yang sama. Diagram biplot PCA dari lokasi Mandiangin, S. Bengkal dan Simpang ditampilkan pada lampiran 8.
Tabel 5 Persentase pengelompokan spesies pada setiap lokasi pengambilan sampel
Nama jenis Lokasi
Mandiangin S. Bengkal Pelayangan Simpang
O. hypopthalmus 90.00 80.00 100.00 50.00
K. limpok 80.00 100.00 100.00 100.00
K. micronema 100.00 100.00 100.00 80.00
K. bichirris 100.00
Rerata 90.00 93.33 100.00 76.67
Gambar 4 Biplot PCA sampel ikan lais dari Pelayangan, 1-10 = O. hypopthalmus, 11-20 = K. limpok, 21-30 = K. micronema, 31-40 = K. bichirris.
Pengelompokan sampel berdasarkan lokasi juga mampu menunjukkan karakter spesifik sebagai penciri spesies dalam kelompok ikan lais (Gambar 4, Lampiran 8). Ompok hypopthalmus dicirikan dengan indeks N9 (tinggi pangkal kepala/tinggi badan). Kryptopterus limpok dicirikan dengan indeks N8 (tinggi moncong/tinggi pangkal kepala). Kryptopterus bichirris dicirikan dengan indeks
N11 (diameter mata/panjang kepala) dan N12 (panjang sirip dada/panjang kepala). Kryptopterus micronema walaupun terletak mengelompok, akan tetapi tidak ada indeks morfometrik penciri jenis ini.
Analisis biplot PCA antarlokasi menunjukkan hasil yang berbeda pada K. limpok dan O. hypopthalmus. Ompok hypopthalmus dapat dikelompokkan berdasarkan lokasi (Gambar 5), sedangkan K. limpok hanya dapat dikelompokkan di lokasi S. Bengkal (Lampiran 9). Biplot PCA pada O. hypopthalmus menunjukkan 75% individu pada spesies ini mengelompok berdasarkan lokasi. Pengelompokan tertinggi ditemukan pada O. hypopthalmus yang berasal dari Pelayangan. 90% individu O. hypopthalmus dari lokasi ini mengelompok di kuadran II. Pengelompokan terendah ditemukan pada O. hypopthalmus dari lokasi Simpang. 50% individu O. hypopthalmus di lokasi Simpang berada di kuadran I, sedangkan 50% yang lain berada di kuadran III.
Gambar 5 Biplot PCA O. hypopthalmus, 1-10 = Mandiangin, 11-20 = S. Bengkal, 21-30 = Pelayangan, 31-40 = Simpang.
Analisis biplot PCA pada O. hypopthalmus juga mampu menunjukkan indeks morfometrik penciri lokasi. Ompok hypopthalmus di Mandiangin dicirikan dengan indeks N4 (tinggi ekor/panjang baku) dan N10 (tinggi ekor/tinggi badan). Hal ini berarti O. hypopthalmus di Mandiangin memiliki struktur ekor yang lebih tinggi/tebal dibanding O. hypopthalmus dari lokasi lain. Ompok hypopthalmus di Pelayangan dicirikan dengan N6 (panjang rahang atas/panjang kepala), sedangkan O. hypopthalmus di Simpang dicirikan dengan N8 (tinggi ‟moncong‟/tinggi pangkal kepala). Tidak ada penciri O. hypopthalmus dari S. Bengkal.
Keragaman Gen Cytochrome b Berdasarkan Runutan Nukleotida
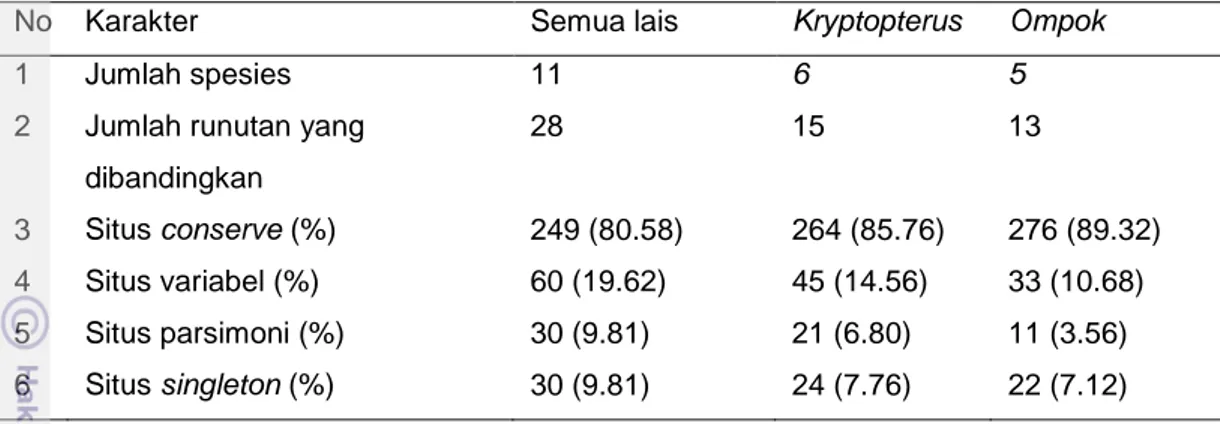
Penjajaran (alignment) gen cyt b semua sampel dari Batang Hari dengan gen cyt b semua jenis ikan lais di genbank dan Kampar (Elvyra 2009) menghasilkan runutan nukleotida sepanjang 927 bp (Lampiran 10). Nukleotida yang berbeda (variabel) pada genus Kryptopterus dan Ompok tidak menunjukkan perbedaan yang signifikan (33.01% berbanding 28.91%) (Tabel 6).
Tabel 6 Karakterisasi nukleotida pada beberapa perbandingan ikan lais
No Karakter Semua lais Kryptopterus Ompok
1 Jumlah spesies 11 6 5
2 Jumlah runutan 28 15 13
3 Situs conserve (%) 551 (59.44) 621 (66.99) 659 (71.09)
4 Situs variabel(%) 376 (40.56) 306 (33.01) 268 (28.91)
5 Variabel di kodon pertama (%) 74 (7.98) 63 (6.80) 44 (4.75)
6 Variabel di kodon kedua (%) 22 (2.37) 12 (1.29) 12 (1.29)
7 Variabel di kodon ketiga (%) 280 (30.20) 147 (15.86) 212 (22.87)
a. Substitusi transisi (%) 147 (15.86) 119 (12.84) 111 (11.97) b. Substitusi transversi (%) 133 (14.35) 28 (3.02) 101 (10.90) 8 Situs parsimoni (%) 274 (29.56) 198 (21.36) 154 (16.61) 9 Situs singleton (%) 102 (11.00) 108 (11.65) 114 (12.30) 10 Persentase timin 26.20 27.20 25.30 11 Persentase sitosin 32.30 31.60 32.00 12 Persentase adenin 27.40 26.90 27.90 13 Persentase guanin 14.10 14.30 13.80
*situs parsimoni=ditemukan minimal dua jenis nukleotida, setiap jenis nukleotida dimiliki oleh minimal dua runutan, situs singleton=nukleotida yang berbeda hanya ditemukan pada satu runutan.
Berdasarkan posisi nukleotida pada triplet kodon, keragaman nukleotida terbesar secara berturut-turut adalah pada nukleotida ketiga, kesatu dan kedua. Berdasarkan jumlah keragaman nukleotida pada kodon pertama dan kedua, dapat dinyatakan bahwa keragaman asam amino pada genus Kryptopterus minimal sebesar 8.09%, sedangkan pada Ompok minimal sebesar 6.04%. Komposisi basa nitrogen terbesar penyusun runutan adalah sitosin, sedangkan guanin memiliki komposisi terkecil. Ditemukan satu situs nukleotida pembeda Kryptopterus dan Ompok. Nukleotida pembeda ini terletak pada situs ke-39 dari runutan gen cyt b
parsial (ke-132 gen cyt b utuh). Situs ke-39 pada Kryptopterus adalah timin, sedangkan pada Ompok adalah sitosin.
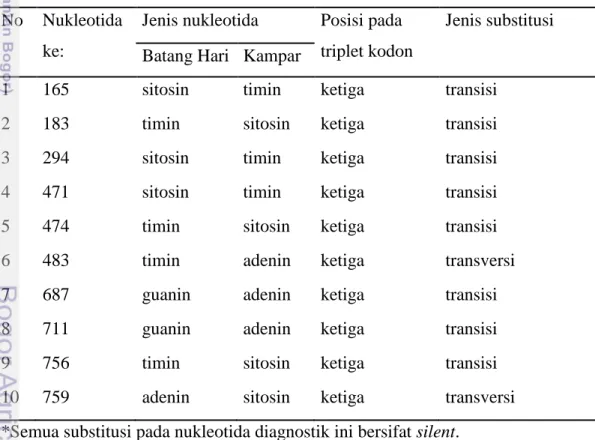
Terdapat 20 situs nukleotida pembeda Kryptopterus dan Ompok dari Sumatera (Batang Hari dan Kampar) (Lampiran 11). Satu diantara 20 situs pembeda ini bersifat substitusi nonsilent, sehingga menghasilkan asam amino pembeda Kryptopterus dengan Ompok dari Sumatera. Asam amino ke-124 dari gen cyt b parsial (ke-155 dari gen cyt b utuh) pada Kryptopterus adalah valina sedangkan pada Ompok adalah isoleusina.
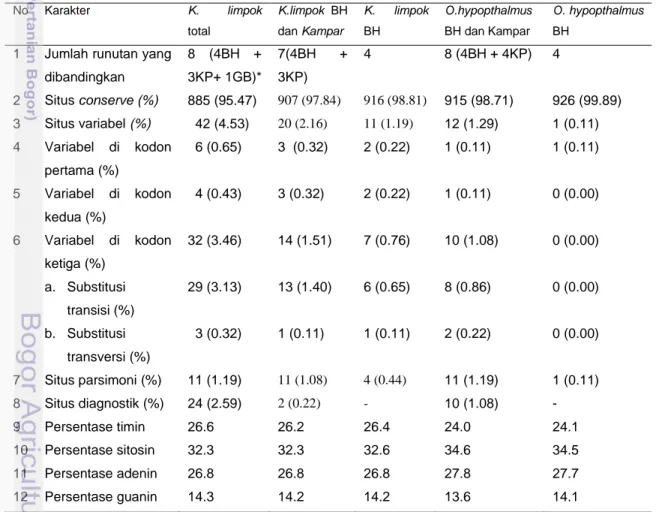
Hasil alignment intraspesies di DAS Batang Hari menunjukkan keragaman yang lebih tinggi pada K. limpok dibanding pada O. hypopthalmus (Tabel 7). Nukleotida yang berbeda pada K. limpok sebanyak 11 situs (1.19 %), sedangkan pada Ompok hanya 1 situs (0.11%).
Tabel 7 Karakterisasi nukleotida pada K. limpok dan O. hypopthalmus
No Karakter K. limpok total K.limpok BH dan Kampar K. limpok BH O.hypopthalmus BH dan Kampar O. hypopthalmus BH
1 Jumlah runutan yang
dibandingkan 8 (4BH + 3KP+ 1GB)* 7(4BH + 3KP) 4 8 (4BH + 4KP) 4 2 Situs conserve (%) 885 (95.47) 907 (97.84) 916 (98.81) 915 (98.71) 926 (99.89) 3 Situs variabel (%) 42 (4.53) 20 (2.16) 11 (1.19) 12 (1.29) 1 (0.11) 4 Variabel di kodon pertama (%) 6 (0.65) 3 (0.32) 2 (0.22) 1 (0.11) 1 (0.11) 5 Variabel di kodon kedua (%) 4 (0.43) 3 (0.32) 2 (0.22) 1 (0.11) 0 (0.00) 6 Variabel di kodon ketiga (%) 32 (3.46) 14 (1.51) 7 (0.76) 10 (1.08) 0 (0.00) a. Substitusi transisi (%) 29 (3.13) 13 (1.40) 6 (0.65) 8 (0.86) 0 (0.00) b. Substitusi transversi (%) 3 (0.32) 1 (0.11) 1 (0.11) 2 (0.22) 0 (0.00) 7 Situs parsimoni (%) 11 (1.19) 11 (1.08) 4 (0.44) 11 (1.19) 1 (0.11) 8 Situs diagnostik (%) 24 (2.59) 2 (0.22) - 10 (1.08) - 9 Persentase timin 26.6 26.2 26.4 24.0 24.1 10 Persentase sitosin 32.3 32.3 32.6 34.6 34.5 11 Persentase adenin 26.8 26.8 26.8 27.8 27.7 12 Persentase guanin 14.3 14.2 14.2 13.6 14.1
*BH = Batang Hari, KP = Kampar (Elvyra 2009), GB = genbank, situs diagnostik= situs pembeda antar populasi, - = tidak cukup data.
Keragaman K. limpok Sumatera (Batang Hari dan Kampar) juga lebih tinggi dibanding keragaman O. hypopthalmus Sumatera (2.16% berbanding 1.29%). Keragaman K. limpok total (Batang Hari, Kampar dan genbank) dua kali lebih besar dibanding keragaman K. limpok Sumatera. Ditemukan dua situs nukleotida diagnostik (barcoding) K. limpok dari DAS Batang Hari dan 10 situs nukleotida diagnostik O. hypopthalmus dari DAS Batang Hari dengan DAS Kampar (Tabel 8). Dua situs nukleotida barcodingK. limpok Batang Hari terletak pada nukleotida ke-30 dan ke-891 dari runutan gen cyt b parsial. Nukleotida ke-30 pada K. limpok Batang Hari adalah sitosin, sedangkan pada K. limpok genbank dan Kampar adalah timin. Nukleotida ke-891 pada K. limpok Batang Hari adalah guanin, sedangkan pada K. limpok genbank dan Kampar adalah adenin. Kedua substitusi ini bersifat silent.
Tabel 8 Daftar situs diagnostik O. hypopthalmus dari DAS Batang Hari dengan DAS Kampar.
No Nukleotida ke:
Jenis nukleotida Posisi pada triplet kodon
Jenis substitusi Batang Hari Kampar
1 165 sitosin timin ketiga transisi 2 183 timin sitosin ketiga transisi 3 294 sitosin timin ketiga transisi 4 471 sitosin timin ketiga transisi 5 474 timin sitosin ketiga transisi 6 483 timin adenin ketiga transversi 7 687 guanin adenin ketiga transisi 8 711 guanin adenin ketiga transisi 9 756 timin sitosin ketiga transisi 10 759 adenin sitosin ketiga transversi *Semua substitusi pada nukleotida diagnostik ini bersifat silent.
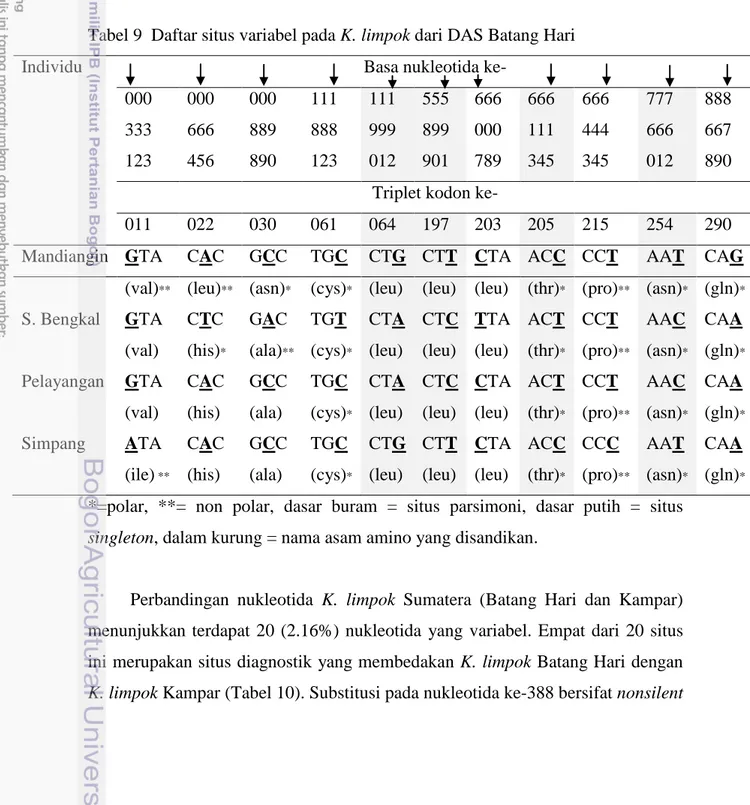
Nukleotida yang variabel pada K. limpok Batang Hari terdiri dari empat situs parsimoni dan tujuh situs singleton (Tabel 9). Empat situs parsimoni yang
ditemukan konsisten membedakan K. limpok Mandiangin-Simpang dengan K. limpok S. Bengkal-Pelayangan. Keempat situs nukleotida parsimoni ini merupakan substitusi transisi, tiga terjadi pada basa pirimidin (timin menjadi sitosin atau sebaliknya) dan satu terjadi pada basa purin (guanin menjadi adenin atau sebaliknya). Selain bersifat transisi, substitusi pada keempat situs parsimoni ini terjadi pada kodon ketiga, sehingga tidak ada yang mengakibatkan perubahan asam amino yang disandikan (substitusi silent). Tujuh situs singleton pada K. limpok Batang hari terdiri empat situs di S. Bengkal, dua situs di Simpang dan satu situs di Mandiangin.
Tabel 9 Daftar situs variabel pada K. limpok dari DAS Batang Hari
Individu Basa nukleotida ke-
000 333 123 000 666 456 000 889 890 111 888 123 111 999 012 555 899 901 666 000 789 666 111 345 666 444 345 777 666 012 888 667 890 Triplet kodon ke-
011 022 030 061 064 197 203 205 215 254 290 Mandiangin GTA CAC GCC TGC CTG CTT CTA ACC CCT AAT CAG
(val)** (leu)** (asn)* (cys)* (leu) (leu) (leu) (thr)* (pro)** (asn)* (gln)* S. Bengkal GTA CTC GAC TGT CTA CTC TTA ACT CCT AAC CAA
(val) (his)* (ala)** (cys)* (leu) (leu) (leu) (thr)* (pro)** (asn)* (gln)* Pelayangan GTA CAC GCC TGC CTA CTC CTA ACT CCT AAC CAA
(val) (his) (ala) (cys)* (leu) (leu) (leu) (thr)* (pro)** (asn)* (gln)* Simpang ATA CAC GCC TGC CTG CTT CTA ACC CCC AAT CAA
(ile) ** (his) (ala) (cys)* (leu) (leu) (leu) (thr)* (pro)** (asn)* (gln)* *=polar, **= non polar, dasar buram = situs parsimoni, dasar putih = situs
singleton, dalam kurung = nama asam amino yang disandikan.
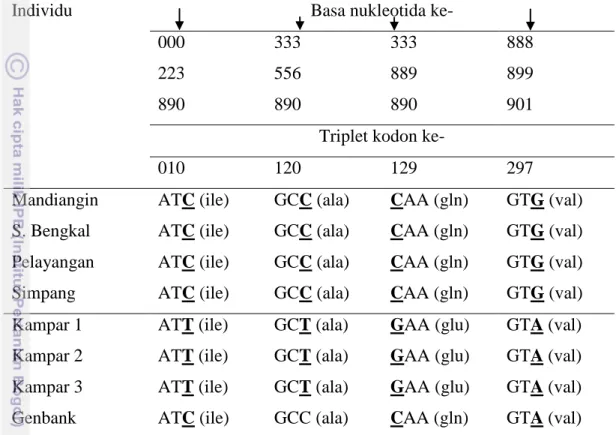
Perbandingan nukleotida K. limpok Sumatera (Batang Hari dan Kampar) menunjukkan terdapat 20 (2.16%) nukleotida yang variabel. Empat dari 20 situs ini merupakan situs diagnostik yang membedakan K. limpok Batang Hari dengan K. limpok Kampar (Tabel 10). Substitusi pada nukleotida ke-388 bersifat nonsilent
sehingga dapat digunakan sebagai situs asam amino diagnostik pembeda K. limpok Batang Hari dengan K. limpok Kampar.
Tabel 10 Daftar situs nukleotida pembeda K. limpok Batang Hari dengan Kampar
Individu Basa nukleotida ke-
000 223 890 333 556 890 333 889 890 888 899 901 Triplet kodon ke-
010 120 129 297
Mandiangin ATC (ile) GCC (ala) CAA (gln) GTG (val) S. Bengkal ATC (ile) GCC (ala) CAA (gln) GTG (val) Pelayangan ATC (ile) GCC (ala) CAA (gln) GTG (val) Simpang ATC (ile) GCC (ala) CAA (gln) GTG (val) Kampar 1 ATT (ile) GCT (ala) GAA (glu) GTA (val) Kampar 2 ATT (ile) GCT (ala) GAA (glu) GTA (val) Kampar 3 ATT (ile) GCT (ala) GAA (glu) GTA (val) Genbank ATC (ile) GCC (ala) CAA (gln) GTA (val) *semua asam amino yang disandikan bersifat polar.
Perbandingan nukleotida pada K. limpok total (Batang Hari, Kampar dan genbank) menunjukkan terdapat 42 (4.53%) nukleotida yang variabel. 21 dari 42 nukleotida yang variabel ini merupakan situs diagnostik pembeda K. limpok Sumatera dengan genbank (Lampiran 10). Empat situs (situs ke-499, 571, 616 dan 884) dari 21 nukleotida yang variabel pada K. limpok total merupakan substitusi nukleotida pada kodon pertama dan kedua, sedangkan 17 lainnya terjadi pada kodon ketiga. Satu situs diantara 17 substitusi yang terjadi pada kodon ketiga (situs ke-786) bersifat transversi. Lima situs ini (situs ke-499, 571, 616, 786 dan 884) menghasilkan lima situs asam amino diagnostik pembeda K. limpok Sumatera dengan genbank (Lampiran 12).
Hanya terdapat satu nukleotida yang berbeda pada seluruh runutan O. hypopthalmus DAS Batang Hari. Nukleotida tersebut terdapat pada nukleotida
ke-883 dari runutan gen cyt b parsial atau nukleotida ke-976 dari gen cyt b utuh. Nukleotida pada O. hypopthalmus dari Simpang dan S. Bengkal adalah guanin, sedangkan pada O. hypopthalmus dari Mandiangin dan Pelayangan adalah adenin. Posisi substitusi nukleotida yang terletak pada kodon pertama mengakibatkan terjadinya perbedaan asam amino ke-295 dari gen cyt b parsial pada jenis O. hypopthalmus. Asam amino ke-295 pada O. hypopthalmus dari Simpang-S. Bengkal adalah alanina (A) dengan triplet kodon GCC, sedangkan pada O. hypopthalmus dari Mandiangin-Pelayangan adalah treonina (T) dengan triplet kodon ACC. Alanina bersifat non polar (hidrofobi), sedangkan treonina bersifat polar.
Perbandingan nukleotida O. hypopthalmus Sumatera (Batang Hari dan Kampar) menunjukkan terdapat 12 (1.29%) nukleotida yang variabel. 10 dari 12 situs ini merupakan situs nukleotida diagnostik yang membedakan O. hypopthalmus Batang Hari dengan O. hypopthalmus Kampar (Tabel 8). Ke-10 situs nukleotida diagnostik ini bersifat substitusi silent. Dua situs variabel lain adalah nukleotida singleton pada situs nukleotida ke-257 gen cyt b parsial pada individu Kampar 2 dan situs ke-883 pada individu S. Bengkal-Simpang.
Terdapat keragaman runutan nukleotida gen cyt b yang sangat besar antara K. bichirris Batang Hari dengan K. bichirris genbank. Jumlah perbedaan nukleotida gen cyt b pada kedua populasi ini sebesar 148 situs (15.97%), padahal runutan yang dibandingkan hanya ada dua. Runutan nukleotida K. bichirris genbank sangat mirip dengan K.minor genbank (AY458895). Jumlah perbedaan nukleotida antara kedua spesies ini hanya 11 situs (1.19%).
Keragaman Gen Cytochrome b Berdasarkan Runutan Asam Amino
Hasil alignment runutan nukleotida seluruh sampel yang dianalisis sepanjang 927 nukleotida ditranslasi menjadi 309 asam amino (Lampiran 12). Asam amino yang variabel pada genus Kryptopterus lebih besar dibanding Ompok (14.56% dengan 6 spesies berbanding 10.68% dengan 5 spesies) (Tabel 11). Situs asam amino yang parsimoni dan singleton pada Kryptopterus lebih tinggi dibanding Ompok.
Tabel 11 Karakterisasi asam amino pada berbagai perbandingan jenis ikan lais
No Karakter Semua lais Kryptopterus Ompok
1 Jumlah spesies 11 6 5
2 Jumlah runutan yang
dibandingkan 28 15 13 3 Situs conserve (%) 249 (80.58) 264 (85.76) 276 (89.32) 4 Situs variabel (%) 60 (19.62) 45 (14.56) 33 (10.68) 5 Situs parsimoni (%) 30 (9.81) 21 (6.80) 11 (3.56) 6 Situs singleton (%) 30 (9.81) 24 (7.76) 22 (7.12)
Perbandingan asam amino hasil translasi gen cyt b parsial pada K. limpok Sumatera (Batang Hari dan Kampar) menunjukkan terdapat 5 (1.62%) asam amino yang variabel (Tabel 12). Satu dari lima situs ini merupakan situs diagnostik, sedangkan empat lainnya merupakan situs singleton. Situs diagnostik pembeda K. limpok Batang Hari dengan K. limpok Kampar ini terletak pada asam amino ke-130 dari translasi nukleotida gen cyt b parsial. Asam amino pada K. limpok Batang Hari adalah glutamina (CAA), sedangkan pada K. limpok Kampar adalah asam glutamat (GAA). Glutamina dan asam glutamat sama-sama bersifat polar. Empat situs singleton pada K. limpok Sumatera terdiri dari tiga situs singleton di Batang Hari (ke-11 di Simpang, ke-22 dan 30 di S. Bengkal) dan satu situs singleton di Kampar (ke-89 di KP1).
Tabel 12 Karakterisasi asam amino pada K. limpok dan O. hypopthalmus
No Karakter K. limpok total K.limpok Sumatera K. limpok BH O.hypopthalmus Sumatera O. hypopthalmus BH 1 Jumlah runutan yang dibandingkan 8 (4BH + 3KP + 1GB) 7 (4Bh + 3 KP) 4 8 (4BH + 4 KP) 4 2 Situs conserve (%) 299 (96.76) 304 (98.38) 306 (99.03) 307(99.35) 308 (99.68) 3 Situs variabel (%) 10 (3.24) 5 (1.62) 3 (0.97) 2 (0.65) 1 (0.32) 4 Situs parsimoni (%) 7 (2.27) 1 (0.32) 0 (0.00) 1 (0.32) 1 (0.32) 5 Situs diagnostik (%) 5 (1.62) 1 (0.32) - 0 (0.00) -
*BH = Batang Hari, KP = Kampar, GB = genbank, - = tidak cukup data.
Perbandingan asam amino pada K. limpok total (Batang Hari, Kampar dan genbank) menunjukkan terdapat 10 (3.24%) asam amino yang variabel. Lima dari 10 asam amino yang variabel ini merupakan situs asam amino diagnostik
pembeda K. limpok Sumatera dengan genbank (Lampiran 12). Lima asam amino ini terletak pada situs ke-167, 179, 191, 206 dan 295 dari asam amino hasil translasi nukleotida gen cyt b parsial.
Perbandingan asam amino hasil translasi gen cyt b parsial pada O. hypopthalmus Sumatera (Batang Hari dan Kampar) menunjukkan terdapat 2 (0.65%) asam amino yang variabel. Dua situs ini merupakan situs singleton, yaitu situs ke-86 pada O. hypopthalmus KP2 dan situs ke-295 pada O. hypopthalmus S.Bengkal-Simpang. Situs ke-86 pada sebagian besar O. hypopthalmus adalah valina (GTC) sedangkan pada O. hypopthalmus KP2 adalah asam aspartat (GAC). Valina bersifat non polar, sedangkan asam aspartat bersifat polar. Situs ke-295 pada sebagian besar O. hypopthalmus adalah treonina (ACC) sedangkan pada O. hypopthalmus S. Bengkal-Simpang adalah alanina (GCC). Treonina bersifat polar, sedangkan alanina bersifat non polar.
Perbandingan asam amino hasil translasi gen cyt b parsial pada K. bichirris Batang Hari dengan genbank menunjukkan terdapat 15 (4.85%) asam amino yang variabel. Tiga dari 15 situs asam amino yang variabel ini merupakan situs singleton dari genus Kryptopterus secara umum. Dua dari tiga situs singleton (situs ke-177 dan 187) ini ditemukan pada K. bichirris Batang Hari, sedangkan satu situs (ke-282) ditemukan pada K. bichirris genbank (Lampiran 13).
Jarak Genetik dan Filogeni
Jarak genetik diasumsikan berdasarkan jumlah perbedaan nukleotida dan nilai p-distance. Jumlah perbedaan nukleotida pada sampel intra Batang Hari berkisar dari 1 hingga 130 (Tabel 13). Jarak genetik antarspesies terbesar ditemukan antara K. micronema dengan K. bichirris, sedangkan jarak genetik antarspesies terkecil ditemukan antara K. limpok dengan O. hypopthalmus. K. limpok intra Sumatera (Batang Hari dan Kampar) konsisten memiliki jarak genetik yang lebih kecil dibandingkan dengan K. limpok genbank.
Tabel 13 Jarak genetik ikan lais dari Batang Hari berdasarkan runutan gen cyt b parsial 927 bp dengan spesies pembanding dari genbank dan Kampar
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 1 - 0.01 0.01 0.00 0.01 0.01 0.01 0.03 0.12 0.13 0.15 0.11 0.11 0.11 0.11 0.15 2 9 - 0.00 0.01 0.01 0.02 0.01 0.03 0.12 0.14 0.15 0.11 0.11 0.11 0.11 0.15 3 5 4 - 0.01 0.01 0.01 0.01 0.03 0.12 0.13 0.15 0.11 0.11 0.11 0.11 0.15 4 3 10 6 - 0.01 0.01 0.01 0.03 0.12 0.13 0.15 0.11 0.11 0.10 0.11 0.15 5 8 11 7 9 - 0.01 0.00 0.03 0.12 0.14 0.15 0.12 0.11 0.11 0.11 0.15 6 12 14 11 13 6 - 0.00 0.03 0.12 0.14 0.15 0.12 0.12 0.11 0.11 0.15 7 10 12 9 11 2 4 - 0.03 0.12 0.14 0.15 0.12 0.12 0.11 0.11 0.15 8 26 31 27 27 26 30 28 - 0.12 0.13 0.16 0.12 0.12 0.11 0.11 0.16 9 110 113 109 110 110 112 112 110 - 0.14 0.16 0.11 0.12 0.11 0.11 0.16 10 125 126 122 123 127 129 129 124 130 - 0.16 0.12 0.12 0.12 0.12 0.16 11 141 137 137 141 139 142 141 144 144 148 - 0.14 0.14 0.14 0.14 0.01 12 103 106 104 101 107 109 109 108 106 115 129 - 0.00 0.01 0.01 0.14 13 102 105 103 100 106 108 108 107 107 114 128 1 - 0.01 0.01 0.14 14 99 104 100 97 103 105 105 104 100 112 130 10 11 - 0.00 0.14 15 100 105 101 98 104 106 106 105 101 113 130 11 12 1 - 0.14 16 142 138 138 142 140 143 142 144 144 148 11 132 131 130 131 -
*1=K. limpok Mandiangin, 2=K.limpok S. Bengkal, 3=K. limpok Pelayangan, 4=K.limpok
Simpang, 5 =K. limpok KP1, 6=K. limpok KP2, 7=K. limpok KP3, 8=K. limpok genbank,
9=K. micronema Pelayangan, 10=K. bichirris Pelayangan, 11=K. bichirris genbank, 12=O. hypopthalmus Mandiangin-Pelayangan, 13=O. hypopthalmus S. Bengkal-Simpang, 14=O. hypopthalmus KP 1,3 dan 4, 15=O. hypopthalmus KP 2, 16=K. minor
genbank, nilai dibawah diagonal=jumlah perbedaan nukleotida, diatas diagonal=nilai p-distance.
Pengujian jarak genetik dengan metode Kimura 2 parameter menunjukkan nilai distance yang hampir sama dengan nilai p-distance. Nilai genetic distance metode Kimura 2 parameter pada sampel ikan lais intra Batang Hari berkisar dari 0.00 hingga 0.16 (Lampiran 14). Perbandingan jarak genetik terkecil dan terbesar antar runutan, baik pada intra maupun antarspesies sama dengan perbandingan jarak genetik terkecil dan terbesar pada p-distance.
Konstruksi filogeni antara semua sampel ikan lais berdasarkan nilai p-distance dan Kimura distance menunjukkan topologi yang sama, hanya berbeda dalam angka bootstrap (Gambar 6, Lampiran 15). Genus Kryptopterus dan Ompok dapat dibedakan menjadi cluster yang terpisah. O. hypopthalmus
membentuk cluster sendiri yang terpisah dari cluster Kryptopterus maupun Ompok secara umum.
Gambar 6 Konstruksi filogeni seluruh ikan lais yang dianalisis berdasarkan nilai p-distance runutan nukleotida gen cyt b parsial 927 bp dengan metode neighbour joining, bootstrap 1000.
Populasi O. hypopthalmus intra DAS Batang Hari mengelompok sempurna, akan tetapi populasi K.limpok Batang Hari bergabung dengan K. limpok kampar. K. limpok dan O. hypopthalmus Sumatera mengelompok pada kedua topologi dengan nilai bootstrap yang hampir sempurna (99 dan 100). Populasi K. limpok total (Batang Hari, Kampar dan genbank) juga mengelompok dengan nilai bootstrap yang sempurna (100).
Konstruksi filogeni berdasarkan asam amino hasil translasi nukleotida gen cyt b parsial (Gambar 7), tidak dapat memisahkan populasi O. hypopthalmus intra
K.limpok KP2 K.limpok KP3 K.limpok KP1 Oi7 K.limpok Ti1 K.limpok Ni4 K.limpok Ki3 K.limpok K.limpok GB K.kryptopterus GB K.schilbeides GB K.bichirris GB K.minor GB Wilcox K.minor GB Hardman Tm2 K.micronema Tb8 K.bichirris Nh2 O.hypopthalmus Th6 O.hypopthalmus Oh3 O.hypopthalmus Kh2 O.hypopthalmus O.hypopthalmus KP2 O.hypopthalmus KP4 O.hypopthalmus KP1 O.hypopthalmus KP3 O.pabo GB O.bimaculatus GB Lakra O.pabda GB O.bimaculatus GB Hardman O.miostoma GB 97 100 100 58 97 92 63 44 99 100 98 59 85 57 20 10 16 24 72 71 99 100 100 0.02
Batang Hari. Populasi K. limpok total dan O. hypopthalmus Sumatera mengelompok sempurna dengan nilai bootstrap yang lebih kecil dibanding konstruksi filogeni berdasarkan nukleotida. Konstruksi filogeni berdasarkan asam amino hasil translasi nukleotida gen cyt b parsial juga tidak dapat memisahkan cluster Kryptopterus dengan Ompok.
Gambar 7 Konstruksi filogeni seluruh ikan lais yang dianalisis berdasarkan nilai p-distance asam amino hasil translasi runutan gen cyt b parsial 927 bp dengan metode Neighbour joining, bootstrap 1000.
Berdasarkan perbandingan topologi filogeni yang dihasilkan, filogeni berdasarkan runutan nukleotida lebih mendekati sistematika ikan lais yang dikemukakan oleh ahli taksonomi morfologis. Konstruksi filogeni berdasarkan runutan asam amino tidak mampu memisahkan cluster Kryptopterus dengan Ompok. Konstruksi filogeni berdasarkan runutan asam amino juga tidak mampu memisahkan populasi O. hypopthalmus Batang Hari dengan O. hypopthalmus Kampar. Padahal filogeni berdasarkan nukleotida menunjukkan populasi O.
O.hypopthalmus KP1 O.hypopthalmus KP4 O.hypopthalmus KP3 Nh2 O.hypopthalmus Th6 O.hypopthalmus O.hypopthalmus KP2 Oh3 O.hypopthalmus Kh2 O.hypopthalmus O.pabo GB Tb8 K.bichirris O.pabda GB O.bimaculatus GB Hardman O.miostoma GB Tm2 K.micronema O.bimaculatus GB Lakra K.minor GB Wilcox K.bichirris GB K.minor GB Hardman K.schilbeides GB K.limpok GB Ki3 K.limpok Ni4 K.limpok Ti1 K.limpok Oi7 K.limpok K.limpok KP1 K.limpok KP2 K.limpok KP3 K.kryptopterus GB 99 73 100 54 46 41 61 66 94 52 31 22 25 9 13 11 23 67 70 58 45 0.01
hypopthalmus Batang Hari dengan O. hypopthalmus Kampar terpisah dengan nilai bootstrap yang hampir sempurna (99). Selain itu, nilai bootstrap yang konsisten lebih tinggi pada filogeni berdasarkan nukleotida menunjukkan bahwa filogeni berdasarkan nukleotida memiliki nilai kepercayaan yang lebih tinggi.
PEMBAHASAN
Kondisi Perairan Lokasi Pengambilan Sampel
Hasil analisis biplot PCA pada data parameter fisika-kima perairan lokasi pengambilan sampel sedikit berbeda dengan asumsi awal lokasi pengambilan sampel. Asumsi awal lokasi pengambilan sampel adalah Mandiangin dan Sungai Bengkal (S. Bengkal) sebagai lokasi perwakilan bagian hulu, Simpang perwakilan lokasi bagian hilir dan Pelayangan sebagai lokasi antara. Mandiangin dan S. Bengkal yang sama-sama sebagai perwakilan daerah hulu seharusnya memiliki parameter fisika-kimia yang lebih mirip. Akan tetapi, hasil analisis PCA menunjukkan lokasi S. Bengkal dan Pelayangan memiliki parameter fisika-kimia yang lebih mirip dibanding dengan S. Bengkal dan Mandiangin. Perbedaan debit sungai yang besar antara lokasi Mandiangin dan S. Bengkal kemungkinan menjadi penyebab perbedaan ini. Berdasarkan hasil pengamatan pada lebar sungai, debit sungai yang berasal dari S. Bengkal memiliki volume yang lebih besar. Debit sungai yang lebih besar dari S. Bengkal membuat lokasi Pelayangan lebih terpengaruh dengan parameter fisika-kimia dari S. Bengkal dibanding menghasilkan parameter fisika-kimia gabungan Mandiangin dan S. Bengkal.
Kecepatan arus yang merupakan ciri parameter fisika daerah hulu sesuai dengan hasil penelitian ini. Kecepatan arus tertinggi ditemukan di Mandiangin sebesar 0.42 m/dtk, kemudian di S. Bengkal sebesar 0.20 m/dtk. Pelayangan dan Simpang memiliki kecepatan arus yang sama (0.17 m/dtk), namun arah arus berbeda antara Pelayangan dan Simpang. Arah arus di Pelayangan tetap dari hulu ke hilir, sedangkan arah arus di Simpang sudah terpengaruh dengan pasang surut air laut. Arah arus pagi dan sore hari di Simpang begerak dari hilir ke hulu. Hanya arus pada siang hari yang bergerak dari hulu ke hilir. Pasang laut mempengaruhi pergerakan arus di Simpang antara pukul 17.00 hingga 09.00. Kecepatan arus di Simpang dihitung sebagai rata-rata pagi, siang dan sore, tanpa memperhitungkan arah arus.
Parameter kecerahan yang juga biasanya menjadi ciri parameter fisika di daerah hulu ternyata berbeda dengan hasil penelitian ini. Kecerahan tertinggi justru didapatkan di daerah Simpang, yang merupakan lokasi pengambilan sampel
paling hilir (muara). Hal ini mungkin diakibatkan banyaknya aktivitas penambangan emas liar (dompeng) di daerah Sarolangun, Muara Bungo dan Muara Tebo. Aktivitas dompeng mengangkat endapan pasir dan lumpur dalam volume yang sangat besar setiap hari. Hal ini menyebabkan sungai di daerah Sarolangun, Muara Tebo dan Muara Tembesi sangat keruh. Kecerahan yang tinggi di Simpang diduga karena telah terjadi pengendapan lumpur di sepanjang aliran sungai Batang Hari dari Muara Tembesi hingga Simpang. Sepanjang aliran sungai Batang Hari dari Muara Tembesi hingga Simpang bayak terdapat cerukan-cerukan (danau-danau) kecil yang membuat pengendapan lumpur lebih mudah. Selain itu daerah Simpang telah terpengaruh oleh pasang surut air laut, sehingga kualitas air (kecerahan) telah dipengaruhi oleh kecerahan air laut.
Nilai oksigen terlarut (DO) yang ditemukan berada dalam kisaran normal untuk DO perairan mengalir. Menurut Wetzel (2001), nilai DO untuk air mengalir yang tidak mengalami eutrofikasi berfluktuasi dari 9 ppm hingga 11 ppm. Nilai DO terendah akan ditemukan sesaat sebelum matahari terbit yaitu sekitar 9 ppm. Nilai DO tertinggi akan ditemukan sesaat setelah matahari tenggelam mencapai angka diatas 11 ppm. Nilai DO pada perairan yang mengalami eutrofikasi memiliki fluktuasi yang lebih besar yaitu dari 3 ppm sesaat sebelum matahari terbit hingga 13 ppm sesaat setelah matahari tenggelam. Nilai DO di perairan dipengaruhi oleh aktivitas fotosintesis fitoplankton, respirasi biota air, laju difusi oksigen antara udara dengan air dan intensitas panas matahari di permukaan perairan (Culberson & Piedrahita 1996). Berdasarkan nilai DO yang ditemukan dapat dinyatakan bahwa DAS Batang Hari tidak mengalami eutrofikasi.
Hubungan nilai pH dengan alkalinitas dari penelitian yang dilakukan sedikit berbeda dengan yang dinyatakan pada literatur. Nilai pH yang didapatkan berkisar dari 7.2 hingga 7.9, sedangkan nilai alkalinitas berkisar dari 43-59 ppm. Menurut Wetzel (2001), jika pH berkisar dari 7 hingga 8 maka nilai alkalinitas akan berkisar dari 78 hingga 100 ppm. Jika alkalinitas berada pada angka 43-59 ppm, maka pH seharusnya berada di angka 6.3-6.5. Perbedaan nilai yang ditemukan dengan literatur kemungkinan disebabkan karena nilai alkalinitas mengalami fluktuasi yang besar di perairan. Alkalinitas tidak hanya dipengaruhi pH, tetapi juga material organik terlarut dan aktivitas mikroba perairan. Selain itu, perbedaan
nilai ini kemungkinan disebabkan oleh error pengukuran alkalinitas. Pengukuran alkalinitas dilakukan dengan metode titrasi asam basa yang sangat tergantung dengan ketajaman mata mengamati perubahan warna.
Keragaman Jenis dan Kelimpahan Ikan Lais di Setiap Lokasi Pengambilan Sampel
Berdasarkan hasil penelitian ditemukan bahwa stasiun yang memiliki jenis dan jumlah sampel ikan lais terbanyak adalah stasiun Pelayangan. Jumlah jenis yang ditemukan di Pelayangan sama dengan yang ditemukan di Mandiangin, akan tetapi jumlah individu K. bichirris di Mandingin tidak mencapai target jumlah individu yang direncanakan. Hal ini menunjukkan Pelayangan sebagai lokasi antara lebih kondusif sebagai habitat ikan lais.
Dua jenis Kryptopterus yang ditemukan dalam penelitian ini sama dengan yang ditemukan Tan dan Kottelat (2009). Kedua jenis tersebut adalah K. limpok dan K. bichirris. Lima jenis lain yang ditemukan Tan dan Kottelat (2009), K. kryptopterus, K. eugeneiatus, K. macrocephalus, K. palembangensis dan K. schilbeides tidak ditemukan pada penelitian ini. Penelitian ini justru menemukan satu jenis lain yaitu K. micronema yang tidak ditemukan Tan dan Kottelat (2009). Keberadaan K. micronema di DAS Batang Hari telah dilaporkan Weber dan de Beaufort (1965).
Tidak satupun jenis Ompok yang dikemukakan Tan dan Kottelat (2009) yang ditemukan dalam penelitian ini. Tiga jenis Ompok yang dikemukakan Tan dan Kottelat (2009) adalah O. fumidus, O. leiachantus dan O. rhadinurus. Penelitian ini hanya menemukan satu jenis Ompok yaitu O. hypopthalmus. Ompok hypopthalmus tidak ditemukan oleh Tan dan Kottelat (2009), namun keberadaan O. hypopthalmus di DAS Batang Hari juga telah dilaporkan Tan dan Ng (2000). Ompok hypopthalmus merupakan jenis ikan lais yang paling dominan di DAS Batang Hari dan dapat ditemukan sepanjang tahun, walaupun kelimpahannya tetap dipengaruhi musim.
Perbedan jenis ikan lais yang didapatkan antara penelitian ini dengan Tan dan Kottelat (2009), kemungkinan disebabkan luas area dan periode pengambilan sampel. Sebagian besar sampel yang digunakan Tan dan Kottelat (2009), dan Tan
dan Ng (2000) merupakan koleksi MuseumRaffles, Singapura (ZRC). Publikasi Tan dan Ng (2000) merupakan hasil ekspedisi Sumatera tahun 1994-1997, ditambah dengan koleksi beberapa museum zoologi, seperti USNM (national museum of natural history, Washington) dan ZMA (instituut voor systematik and populatiebiologie, Amsterdam). Publikasi Tan dan Kottelat (2009) merupakan hasil ekspedisi Sumatera tahun 1998-2003. Ekspedisi Tan dan Kottelat (2009) dan Tan dan Ng (2000) kemungkinan mencakup area yang lebih luas dan waktu yang lebih lama dibanding pengambilan sampel penelitian ini. Lokasi pengambilan sampel Tan dan Kottelat (2009) lebih banyak dilakukan pada rawa-rawa dataran rendah yang terdapat di DAS Batang Hari. Selain itu, waktu pengambilan sampel Tan dan Kottelat (2009) dilakukan tujuh tahun lalu (1998-2003). Rentang waktu tujuh tahun (2003-2010), memungkinkan telah terjadi perbedaan pada komposisi jenis ikan lais di DAS Batang Hari.
K. bichirris merupakan jumlah sampel yang paling sedikit dan hanya ditemukan di Pelayangan dan Mandiangin. Jenis ini disebut dengan lais kaca karena memiliki tubuh yang transparan saat masih hidup (Kottelat et al. 1993). Selain berdasarkan jumlah jari-jari sirip anal, jenis ini mudah dibedakan dengan jenis Kryptopterus lain secara kasat mata karena memiliki tubuh yang relatif lebih tinggi (punggung yang lebih cembung). Karakter tubuh yang lebih tinggi ini ternyata muncul dalam analisis biplot PCA.
Keragaman Struktur Morfologis
Pengukuran proporsi ukuran tubuh yang dikategorikan sebagai data nisbah morfometrik menunjukkan nilai keragaman yang rendah dalam dan antar populasi ikan lais di DAS Batang Hari. Keragaman yang hanya berkisar dari 0.01 hingga 0.51 (χ = 0.15–1.12) pada spesies yang sama antar lokasi menunjukkan bahwa belum terjadi pembentukan subpopulasi. Hal ini didukung dengan hasil uji t yang menunjukkan bahwa parameter lingkungan antar lokasi pengambilan sampel tidak berbeda nyata.
Hasil biplot PCA terhadap indeks morfometrik menunjukkan sebagian besar jenis ikan lais memiliki struktur morfometrik yang khusus. Hal ini berarti bahwa, selain berdasarkan ciri meristik yang digunakan Kottelat et al. (1993), jenis ikan
lais dapat juga dibedakan berdasarkan ciri morfometrik. Indeks N9 (tinggi pangkal kepala/tinggi badan) yang ditemukan pada O. hypopthalmus dan indeks N11 (diameter mata/panjang kepala) pada K. bichirris telah digunakan Ng (2003;Ng 2004) sebagai kunci identifikasi jenis-jenis Kryptopterus. Indeks N12 (panjang sirip dada/panjang kepala) pada K. bichirris juga telah digunakan untuk kunci identifikasi Kryptopterus oleh Kottelat et al. (1993). Indeks N8 (tinggi moncong/tinggi pangkal kepala) pada K. limpok belum pernah digunakan untuk kunci identifikasi jenis-jenis Kryptopterus dan Ompok. Indeks N8 dapat diajukan sebagai salah satu karakter penciri K. limpok. K. micronema tidak memiliki ciri khusus pada struktur morfometriknya sehingga penciri jenis ini harus tetap didasarkan pada karakter meristik yang dikemukakan Weber dan de Beafort (1965; Kottelat et al. 1993).
Ciri batang ekor yang lebih besar pada O. hypopthalmus di Mandiangin kemungkinan berhubungan dengan arus yang kuat di lokasi ini. Kondisi arus yang kuat membuat pergerakan ekor menjadi lebih aktif. Pergerakan yang lebih aktif membuat struktur batang ekor menjadi lebih besar. Rahang atas yang lebih panjang pada O. hypopthalmus di Pelayangan kemungkinan berhubungan dengan jenis makanan yang lebih bervariasi di lokasi ini. Jenis makanan yang bervariasi akan membuat bukaan mulut menjadi lebih lebar yang ditandai rahang atas yang lebih panjang.
Ompok hypopthalmus yang terletak mengelompok berdasarkan lokasi dan pengelompokan individu K. limpok dari Pelayangan menunjukkan bahwa telah mulai terjadi perbedaan struktur morfologis antar lokasi di DAS Batang Hari. Parameter fisika-kimia perairan yang diukur antar lokasi pengambilan sampel memang tidak menunjukkan perbedaan yang signifikan. Akan tetapi, penambahan parameter lain terutama parameter kimia yang berhubungan dengan pencemaran akibat dompeng mungkin akan memberikan hasil yang berbeda.
Keragaman Gen Cytochrome b
Nilai keragaman runutan gen parsial cyt b yang ditemukan dalam penelitian ini sedikit lebih tinggi dibanding nilai keragaman pada beberapa literatur. Nilai keragaman nukleotida seluruh sampel ikan lais yang dibandingkan adalah sebesar
40.56%. Keragaman ini mencakup dua genus, 11 spesies dan 28 runutan sepanjang 927 bp. Slechtova et al. (2006), menemukan nilai keragaman nukleotida gen cyt b Botiidae sebesar 44.73%. Nilai keragaman ini mencakup 7 genus, 34 spesies dan 96 runutan sepanjang 1111 bp. Menurut Tang et al. (2006), keragaman nukleotida gen cyt b antar genus dalam famili yang sama berkisar 7.79-14.80%, walaupun pada beberapa takson dapat lebih tinggi.
Nilai keragaman nukleotida yang lebih besar dari nilai rata-rata keragaman gen cyt b kemungkinan besar karena asal populasi sampel yang sangat beragam. Runutan gen cyt b ikan lais pembanding yang diperoleh dari genbank berasal dari tiga penelitian yang berbeda yaitu Wilcox (2004), Hardman (2005) dan Lakra et al. (unpublished data). Wilcox (2004), menggunakan ikan lais akuarium sebagai sumber sampel DNA sehingga asal populasi tidak diketahui. Hardman (2005), menggunakan sampel ikan lais dari museum zoologi sehingga asal populasi juga tidak dapat dipastikan. Lakra et al. (unpublished data) kemungkinan besar menggunakan sampel ikan lais dari sungai Gangga di India. Penelitian Slechtova et al. (2006), walaupun menggunakan 34 spesies dari 7 genus, akan tetapi semua sampel yang digunakan hanya berasal dari kawasan Asia Tenggara.
Nilai keragaman untuk genus Kryptopterus dan Ompok juga lebih tinggi dibanding dengan beberapa literatur pembanding. Keragaman untuk genus Kryptopterus adalah sebesar 33.01% untuk enam spesies dengan 15 runutan sepanjang 927 bp. Keragaman untuk genus Ompok adalah sebesar 28.91% untuk lima spesies dengan 13 runutan sepanjang 927 bp. Slechtova et al. (2006) menemukan nilai keragaman antarspesies dalam genus yang sama (intragenus) hanya berkisar dari 7.24% hingga 11.55%. Slechtova et al. (2006) rata-rata menggunakan lima spesies dengan 16 runutan sepanjang 1111 bp dalam pengujian nilai keragaman intragenus.
Posisi situs asam amino pembeda Kryptopterus dengan Ompok intra Sumatera yang ditemukan dalam penelitian ini sama dengan yang ditemukan Elvyra (2009) di DAS Kampar. Situs pembeda ini terletak pada asam amino ke-124 dari runutan gen cyt b parsial atau ke-155 dari runutan gen cyt b utuh. Penambahan runutan dari lokasi lain di Sumatera selain Batang Hari dan Kampar akan membantu memverifikasi dan mengukuhkan situs asam amino pembeda ini.
Keragaman gen cyt b K. limpok di DAS Batang Hari (1.19%) lebih tinggi dibanding K. limpok Kampar. Keragaman gen cyt b K. limpok di DAS Kampar adalah sebesar 0.68% (Elvyra 2009). Panjang runutan yang digunakan dalam penelitian ini (927 bp) sama dengan yang digunakan Elvyra (2009), akan tetapi jumlah yang digunakan Elvyra (2009) hanya ada tiga. Perbedaan jumlah runutan yang digunakan memang berpengaruh terhadap perbedaan nilai keragaman, akan tetapi keragaman K. limpok di DAS Batang Hari hampir dua kali lebih besar dibanding DAS Kampar. Aboim et al. (2005), menemukan keragaman runutan gen cyt b intra populasi pada Helicolenus dactylopterus berkisar 0.2-0.7%. Nilai keragaman ini untuk runutan gen cyt b sepanjang 423 bp dengan 40 individu.
Nilai keragaman antar populasi K. limpok Batang Hari dengan K. limpok genbank adalah sebesar 3.02%. Nilai yang hampir sama dengan yang ditemukan Elvyra (2009) saat membandingkan runutan K. limpok dari Kampar dengan genbank, yaitu sebesar 3.13%. Keragaman gen cyt b parsial antar populasi K. limpok Batang Hari, Kampar dan genbank sebesar 3.88% (36 situs). Nilai ini masuk dalam kisaran yang dikemukakan oleh Tang et al. (2006). Tang et al. (2006), menyatakan bahwa nilai keragaman dalam spesies yang sama harus lebih kecil dari 6.90%.
Penelitian ini menemukan bahwa telah terjadi perbedaan gen cyt b yang signifikan antar populasi K. limpok di DAS Batang Hari. Empat situs nukleotida parsimoni pada K. limpok seperti membagi daerah dengan kecerahan rendah (S. Bengkal-Pelayangan) dengan daerah yang relatif memiliki kecerahan tinggi (Mandiangin-Simpang). Penelitian lanjutan dengan menganalisis parameter fisika-kimia perairan yang lebih lengkap antar lokasi pengambilan sampel dibutukan untuk menjelaskan penyebab adanya perbedaan gen cyt b K. limpok di DAS Batang Hari. Selain analisis parameter fisika-kimia perairan, perlu dilakukan penelitian lanjutan tentang ukuran populasi, intensitas penangkapan dan rasio jantan betina dalam populasi K. limpok untuk menganalisis adanya perbedaan ini. Menurut Yamaguchi et al. (2010) intensitas penangkapan yang tinggi akan menimbulkan keragaman gen yang tinggi antar populasi dan menurunkan keragaman intrapopulasi. Ukuran populasi yang kecil akan memacu inbreeding
sehingga akan meningkatkan keragaman gen antar populasi (Reed & Frankham 2003).
Berbeda dengan K. limpok, runutan gen cyt b pada O. hypopthalmus sangat conserve. Seperti halnya penelitian ini, Elvyra (2009) hanya menemukan satu nukleotida yang variabel dari 4 runutan sepanjang 927 bp yang dibandingkan. Akan tetapi posisi nukleotida yang variabel berbeda antara O. hypopthalmus Kampar dengan O. hypopthalmus Batang Hari. Nukleotida yang variabel pada O. hypopthalmus Kampar terletak pada situs ke-257, sedangkan pada O. hypopthalmus Batang Hari terletak pada situs ke-883. Substitusi pada situs ke-257 O. hypopthalmus Kampar adalah substitusi timin (T) menjadi adenin (A) pada individu Kampar 2. Substitusi ini terletak pada kodon ke-2 sehingga bersifat nonsilent. Asam amino pada situs ke-86 pada individu Kampar 2 adalah asparagina (N), sedangkan pada individu Kampar 1 dan 3 adalah valina. Substitusi pada situs ke-883 pada O. hypopthalmus Batang Hari adalah substitusi guanin (G) menjadi adenin (A) atau sebaliknya. Substitusi ini terletak pada kodon pertama dan bersifat nonsilent. Asam amino ke-295 pada O. hypopthalmus dari Simpang-S. Bengkal adalah alanina (A), sedangkan pada O. hypopthalmus dari Mandiangin-Pelayangan adalah treonina (T). Hal ini menunjukkan bahwa gen cyt b pada K. limpok lebih beragam dibanding dengan O. hypopthalmus.
Nilai keragaman gen cyt b yang sangat besar antara K. bichirris Batang Hari dan K. bichirris genbank merupakan hal yang membutuhkan penelitian lebih lanjut. Nilai keragaman nukleotida gen cyt b antar kedua populasi ini (15.45%) melebihi dua kali lipat nilai keragaman intraspesies Tang et al. (2006). Sebaliknya, nilai keragaman nukleotida gen cyt b antarspesies K. bichirris genbank dengan K. minor genbank (1.19%) masuk dalam nilai keragaman intraspesies Tang et al. (2006). Dibutuhkan penambahan jumlah sampel dari lokasi lain, selain Batang Hari untuk membantu menjelaskan keanehan nilai keragaman ini.
Persentase keragaman berdasarkan posisi kodon pada triplet kodon penyusun asam amino juga sama dengan yang ditemukan Farias et al. (2001; Slechtova et al. 2006; Doadrio & Perdices 2006). Penelitian ini menemukan bahwa rata-rata situs nukleotida yang paling bervariasi secara beturut-turut adalah
nukleotida pada posisi ketiga, pertama dan kedua dari triplet kodon penyusun asam amino. Farias et al. (2001; Slechtova et al. 2006; Doadrio & Perdices 2006) menyatakan bahwa nukleotida yang paling bervariasi adalah pada kodon ketiga, sedangkan nukleotida yang paling conserve adalah nukleotida pada kodon kedua.
Nilai situs informasi parsimoni hasil penelitian ini lebih rendah dari yang dikemukakan Farias et al. (2001). Nilai situs informasi parsimoni diartikan sebagai situs-situs yang memiliki minimal dua jenis nukleotida. Salah satu jenis nukleotida harus dimiliki oleh minimal dua runutan dari total runutan yang dibandingkan (Tamura et al. 2007). Nilai situs informasi parsimoni pada penelitian ini adalah sebesar 29.34%, sedangkan yang ditemukan oleh Farias et al. (2001) adalah sebesar 52.64%. Hal ini berarti bahwa takson yang dianalisis Farias et al. (2001) lebih beragam dibanding dengan takson yang dianalisis dalam penelitian ini.
Nilai p-distance antar hasil penelitian juga tidak berbeda jauh dengan literatur pembanding. Hasil pengujian John dan Avise (1998), pada seluruh runutan gen cyt b ikan yang terdapat di genbank hingga 9 0ktober 1997 menunjukkan jarak genetik tingkat genus pada ikan berkisar dari 0 hingga 0.36, dengan 86.25% jarak genetik tingkat genus berada di bawah angka 0.20. Hasil penelitian ini menunjukkan jarak genetik pada genus Kryptopterus berkisar dari 0 hingga 0.18, sedangkan jarak genetik pada Ompok berkisar 0.13 hingga 0.16.
Konstruksi pohon filogeni hasil penelitian ini mampu memisahkan genus Kryptopterus dengan Ompok. Kedua genus ini terpisah tanpa angka bootstrap, walaupun telah digunakan outgroup dari anggota Siluriformes lain. Konstruksi pohon filogeni yang ditemukan berbeda dengan yang ditemukan oleh Elvyra (2009). Penelitian Elvyra (2009), dengan menggunakan gen yang sama pada lokasi penelitian yang berbeda, tidak dapat memisahkan genus Kryptopterus dan Ompok. Penelitian Elvyra (2009), menemukan tiga cluster hasil konstruksi filogeni ikan lais di genbank dengan ikan lais di sungai Kampar, Riau. Cluster pertama diisi oleh K. minor genbank, O. bimaculatus genbank, K. schilbeides genbank, K. schilbeides kampar dan O. hypopthalmus Kampar. Cluster kedua hanya diisi oleh K. kryptopterus genbank. Cluster ketiga diisi oleh K. limpok genbank, K. limpok Kampar, K apogon Kampar dan O. eugeniatus Kampar.
Adanya cluster tersendiri pada O. hypopthalmus yang terpisah dari kelompok Ompok secara umum dan struktur gen cyt b yang sangat conserve merupakan hal yang membutuhkan penelitian lebih lanjut. Dibutuhkan penambahan jumlah sampel baik dari populasi yang sama ataupun dari populasi yang berbeda. Selain penambahan sampel, penggunaan marka genetik lain mungkin akan membantu menjelaskan sistematika berdasarkan data genetik pada O. hypopthalmus. Marka genetik yang disarankan adalah gen cytochrome oxidase 1 (CO1) atau 16S DNA mitokondria.
Gen CO1 mtDNA disarankan karena gen ini telah umum digunakan dan terbukti akurat untuk sistematika berbagai tingkatan hewan (Vences et al. 1995; Hebert & Gregory 2005; Frezal & Leblois 2008). Selain itu, berdasarkan data genbank hingga 19 Desember 2010, runutan pembanding untuk gen ikan lais yang tersedia digenbank hanya gen cyt b dan CO1. Populasi pembanding untuk gen CO1 adalah hasil penelitian Lakra et al. (unpublished data) yang kemungkinan berasal dari India. Selain gen CO1, gen lain yang disarankan untuk sistematika organisme eukariot, selain tumbuhan, adalah tiga situs mtDNA (16S, 12S dan Cyt b) dan dua situs pada genom inti (28S dan ITS) (Vences et al. 1995). Hasil pengujian Cummings et al. (1995) pada 15 situs mtDNA menunjukkan situs 16S memiliki pola topologi yang sama dengan runutan total gen mtDNA pada metode neighbour joining dan maximum likelihood. Hal inilah yang mendasari pemilihan gen CO1 dan 16S mtDNA untuk membantu menjelaskan sistematika O. hypopthalmus.