Klasifikasi, Botani dan Syarat Tumbuh Tanaman Cabai
Tanaman cabai tergolong divisi Magnoliophyta, kelas Magnolipsida, ordo Solanales, famili Solanaceae, genus Capsicum dan spesies Capsicum annuum L. (Kusandriani 1996). Tanaman cabai merupakan tanaman tropika yang memiliki sifat menyerbuk sendiri dengan variasi penyerbukan silang yang tinggi tergantung genotipe dan lingkungan (Daskalov 1998). Persentase penyerbukan silang pada tanaman cabai sekitar 6 – 37 %, persentase penyerbukan silang dipengaruhi oleh posisi dan ukuran stigma. Stigma yang lebih tinggi dibandingkan dengan kotak sari akan menyebabkan penyerbukan silang dan sebaliknya, bunga tanaman cabai bersifat protogeny atau kepala putik telah siap diserbuki sebelum tepung sari masak (Permadi dan Kusandriani 1996). Tanaman cabai mempunyai jumlah kromosom somatik diploid dengan kromosom dasar x = 12. Jumlah kromosom normal cabai adalah 2n=2x=24 (Berke 2000). Menurut Rubatzky dan Yamaguchi (1997), tanaman cabai merupakan tanaman herba yang berkayu pada pangkal batangnya tetapi pada beberapa jenis menjadi semak. Batang utama tegak ber-kayu dan bercabang banyak dengan berkisar 0.5 – 1.5 m dan memiliki perakaran yang dangkal diawali dengan akar tunggang (akar primer) kemudian tumbuh akar rambut ke samping (akar lateral). Panjang akar primer berkisar 35 – 50 cm dan akar lateral berkisar 35 – 45 cm, perkembangan akar lateral yang cepat di dalam tanah dan menyebar pada kedalaman 10 – 15 cm.
Berke (2000) menyatakan bahwa bunga tanaman cabai termasuk lengkap berbentuk terompet terdiri dari kelopak bunga, mahkota bunga, benang sari dan putik. Kelopak bunga berjumlah 6 helai berwarna kehijauan atau ungu, mahkota bunga terdiri atas 5 – 7 petal berwarna putih atau ungu. Dalam satu bunga terdapat 1 putik dan 5 -7 benang sari. Menurut Greenleaf (1986), tanaman cabai mulai berbunga pada umur 23 – 31 hari setelah tanam (HST). Buah cabai mulai masak setelah 45 hari terjadi penyerbukan. Bunga cabai terletak pada setiap ruas, bentuk buah cabai bervariasi dari linear, kerucut atau bulat, warna buahnya bervariasi hijau, kuning atau ungu ketika muda, kemudian berubah menjadi merah, orange, kuning atau ungu pada matangnya. Biji tanaman cabai terletak dalam buah dan melekat sepanjang plasenta. Menurut Berke (2000) dalam satu gram buah cabai
terdapat 220 biji cabai. Rubatzky dan Yamaguchi (1996) menyatakan bahwa biji kultivar Capsicum annuum berbentuk pipih berwarna kuning pucat dan berbentuk bulat telur dengan panjang 3 – 5 mm.
Sumarni (1996) menyatakan bahwa tanaman cabai dapat tumbuh pada berbagai jenis tanah asalkan memiliki drainase dan aerasi yang baik. Tanaman cabai dapat dibudidayakan pada daerah dengan ketinggian tempat hingga 2000 m dpl. Keadaan pH tanah yang ideal untuk tanaman cabai adalah 6.0 – 6.5 dan mengandung bahan organik sekurang-kurangnya 1.5%. Keadaan pH tanah sangat penting karena erat kaitannya dengan ketersediaan unsur hara. Apabila ditanam pada tanah yang mempunyai pH lebih dari tujuh maka tanaman cabai akan menyebabkan tanaman menjadi kerdil dan gejala klorosis atau daun menguning yang disebabkan kekurangan unsur hara besi (Fe). Pada tanah yang mempunyai pH kurang dari lima tanaman cabai juga akan menjadi kerdil karena kekurangan unsur hara kalsium (Ca) dan magnesium (Mg) atau keracunan aluminium (Al) dan
mangan (Mn).
Suhu udara yang baik bagi pertumbuhan tanaman cabai adalah 24 – 270 C
dengan suhu udara yang paling cocok untuk pertumbuhan cabai adalah 160C pada
malam hari dan 230C pada siang hari. Perbedaan suhu udara yang terlalu besar
akan menyebabkan kegagalan proses pembungaan dan pembuahan tanaman cabai. Curah hujan yang sesuai bagi pertumbuhan tanaman berkisar 600 mm – 1250 mm. Curah hujan yang terlalu rendah dapat menghambat pertumbuhan tanaman. Me-nurut Pitojo (2003), curah hujan terlalu tinggi dapat menyebabkan kelembaban udara meningkat dan mendorong pertumbuhan penyakit tanaman. Agar cabai berproduksi optimal disarankan lokasi penanaman pada tempat ruang terbuka dan tidak ternaungi. Tanaman cabai sangat cocok dibudidayakan di sawah atau tegal. Tanaman cabai juga dapat hidup di perkarangan dan mendapat sedikit naungan dari tanaman lain. Tanaman cabai bukan merupakan tanaman hari panjang di-perlukan sinar matahari selama 9 jam per hari.
Interaksi Genotipe x Lingkungan dan Stabilitas
Pemuliaan tanaman bertujuan untuk memperbaiki karakter tanaman sesuai dengan kebutuhan manusia. Perbaikan karakter dilakukan dengan pemanfaatan potensi genetik dan interaksi genotipe x lingkungan. Interaksi genotipe x ling-kungan dapat dipergunakan oleh pemulia tanaman untuk mengembangkan va-rietas unggul baru yang spesifik lingkungan atau vava-rietas yang beradaptasi secara luas. Pemulia tanaman memiliki tugas yang relatif mudah apabila semua ke-ragaman fenotipe hanya dihasilkan oleh genetik saja, proses pemulian tanaman hanya akan dibatasi pada mengidentifikasi nilai tambah genetik dari efek dominan serta akumulasi alel-alel yang menguntungkan ke dalam populasi tanaman. Alberts (2004) menyatakan bahwa karakter tanaman adalah hasil akhir dari genetik yang hampir semuanya bersifat kualitatif serta kebanyakan karakter agronomi yang penting seperti daya hasil bersifat kuantitatif dan dipengaruhi oleh lingkungan. Sangat tidak realistis apabila suatu genotipe unggul di satu lokasi satu musim akan menjadi unggul di semua lokasi yang lain serta unggul di semua musim.
Indonesia merupakan negara kepulauan yang memiliki banyak perbedaan kondisi iklim dan jenis tanah, hal ini akan lebih banyak menimbulkan kesulitan dalam interaksi genotipe x lingkungan. Untuk mengatasi permasalahan ini, pe-mulia tanaman harus membuat percobaan pada beberapa lingkungan (lokasi) dan beberapa musim agar menyakinkan telah memilih genotipe yang memiliki daya hasil yang tinggi dan penampilan yang stabil. Gauch (1992) dan De Lacy et al. (1996) menyatakan bahwa pengaruh lingkungan terhadap genotipe dan inte-raksinya lebih banyak berperan pada uji multi lokasi. Menurut Falconer (1952) dan Fernandez (1991) kedua faktor tersebut tidak selalu berinteraksi positif bahkan seringkali negatif. Untuk mendapatkan hasil interaksi genetik dan kungan yang signifikan diperlukan pengujian multi lokasi yang beragam ling-kungannya. Peto (1982) membagi dua bentuk dari interaksi genotipe x lingkung-an sebagai kualitatif (perubahlingkung-an rlingkung-anking) dlingkung-an kulingkung-antitatif (perbedalingkung-an absolut lingkung-antar genotipe). Hill (1975) dan Yau (1995) menyatakan bahwa interaksi genotipe x lingkungan membuat tidak mudah untuk menyeleksi penampilan terbaik. Genotipe paling stabil dan hal ini merupakan pertimbangan yang paling penting
dalam program pemuliaan tanaman karena mengurangi kemajuan dari seleksi pada satu lingkungan.
Lin dan Binns (1988a) menyatakan bahwa pengamatan penampilan geno-tipe dalam percobaan genogeno-tipe x lokasi x tahun seringkali bermasalah karena ke-hadiran interaksi lokasi x tahun atau yang disebut pengaruh lingkungan. Crossa (1990) menyatakan bahwa data yang dikoleksi dari percobaan multilokasi akan memiliki tiga aspek fundamental yaitu: (1) pola data yang terstruktur, dimana jumlah genotipe yang berespon terhadap lingkungan tertentu sudah tersistematik, signifikan dan bisa terukur; (2) data yang tidak terstruktur, dimana respon bisa tidak terprediksi dan terukur. Fungsi dari disain percobaan dan analisis statistik dari multilokasi adalah menghilangkan dan kemungkinan membuang data yang tidak bisa dijelaskan; (3) hubungan antar genotipe, antar lingkungan dan interaksi genotipe dan lingkungan.
Tanaman sangat dipengaruhi oleh lingkungan tempat tumbuhnya baik se-cara fisik, kimia maupun biologis. Comstock dan Moll (1963) membagi ling-kungan menjadi dua kategori, yaitu: (1) Lingling-kungan mikro, suatu lingling-kungan di-mana satu tanaman bersaing dengan tanaman lain yang tumbuh bersamaan wak-tunya dan tempat. Hal ini termasuk sifat fisik dan kimia seperti jenis tanah, perbedaan cuaca, radiasi matahari, hama dan penyakit yang ada pada lingkungan tanaman tersebut tumbuh. Menurut Roy (2000), lingkungan mikro memberikan dampak variasi galat pada analisis statistik. Kategori (2) Lingkungan makro, lingkungan yang berhubungan skala lokasi atau area pada satuan periode. Lingkungan makro merupakan kumpulan dari lingkungan mikro, dimana setiap lingkungan mikro memberikan dampak yang berbeda pada lingkungan makronya. Dengan kata lain lingkungan makro merujuk pada kondisi iklim, tanah, serta manajemen penanamannya (pemupukan, pengairan, kerapatan tanaman, tanggal tanam, curah hujan dan lain-lain).
Roy (2000) membagi empat klasifikasi genotipe berkaitan dengan kemampuan genotipe beradaptasi dengan lingkungannya, yaitu: (1) Genotipe tidak responsif, dimana tidak menunjukkan dalam perbedaan penampilan dibandingkan dengan genotipe lain walaupun lingkungan sudah diperbaiki seperti penambahan pemupukan, air dan lain-lain; (2) Genotipe toleran, dimana menunjukkan sedikit
atau tidak ada perubahan dibandingkan dengan genotipe lain apabila lingkungan dibuat menjadi lebih jelek, seperti kekurangan nutrisi, air dan lain-lain; (3) Genotipe stabil, dimana menunjukkan sedikit atau tidak ada perubahan diban-dingkan dengan genotipe lain walaupun lingkungan berubah dratis dan tidak bisa dikontrol seperti perbedaan antar musim pada wilayah agroklimat yang sama. Secara singkat stabilitas dikategorikan sebagai mengurangi variasi antar musim; (4) Genotipe adaptasi luas atau fleksibel, genotipe yang tidak atau sedikit menun-jukkan perbedaan dibandingkan dengan genotipe lain ketika ditanam pada wilayah agroklimat yang berbeda. Kemampuan adaptasi dapat didefinisikan berkurangnya variasi dalam semua lingkungan.
Berdasarkan respon terhadap perubahan Roy (2000) juga membagi dua kategori genotipe, yaitu: (1) Homeostatis: kemampuan genotipe apabila ditanam pada lingkungan berbeda, tanaman secara menyeluruh (ukuran, bentuk waktu berbunga atau waktu panen) seragam dan stabil seperti yang dideskripsikan; (2) Stabilitas berkembang (developmental stability): kemampuan genotipe memper-cepat tahap pertumbuhan baik secara fisiologi maupun morfologi dalam meng-hadapi perubahan lingkungan dibandingkan dengan genotipe lain.
Menurut Alberts (2004), pemulia tanaman setuju akan pentingnya sta-bilitas hasil, tetapi sedikit sekali yang menjelaskan definisi stasta-bilitas serta sedikit metode untuk mengukur dan meningkatkan stabilitas hasil. Stabilitas suatu ge-notipe adalah kemampuan gege-notipe untuk hidup pada berbagai lingkungan yang beragam, sehingga fenotipenya tidak banyak mengalami perubahan pada ling-kungan lain. Penyebab stabilitas adalah adanya mekanisme penyangga individu dan penyangga populasi, genotipe dengan hasil tinggi dan stabil akan berpe-nampilan baik pada semua lingkungan. Stabilitas fenotipe disebabkan oleh ke-mampuan tanaman untuk dapat menyesuaikan dirinya terhadap lingkungan yang beragam sehingga tanaman tidak banyak mengalami perubahan sifat fenotipenya.
Lin et al. (1986) mengajukan tiga tipe konsep stabilitas, yaitu : Konsep stabilitas tipe 1, suatu genotipe cenderung stabil apabila ragam antar lingkung-annya kecil. Becker dan Leon (1988) menyebutnya stabilitas statik atau konsep stabilitas biologis. Genotipe ini sangat stabil walaupun berada pada berbagai ma-cam lingkungan. Konsep ini sangat berguna untuk data kualitas, ketahanan hama
dan penyakit atau stres lingkungan. Parameter stabilitas yang bisa mengambarkan
ini adalah koefisien ragam (CVi) (Francis dan Kannenburg 1978) pada setiap
genotipe dan ragam genotipe pada seluruh lingkungan (S2i).
Konsep stabilitas Tipe 2, suatu genotipe cenderung stabil apabila respon terhadap lingkungannya adalah sejajar dengan respon daya hasil untuk semua genotipe. Becker dan Leon (1988) menyatakan sebagai stabilitas dinamis atau stabilitas agronomis. Suatu genotipe stabil apabila tidak memiliki perbedaan secara umum respon terhadap lingkungannya dan bisa diprediksikan responnya
terhadap lingkungan yang lain. Koefisien regresi (bi) (Finlay dan Wilkinson
1963), komponen ragam nilai tengah terhadap interaksi genotipe x lingkungan (i)
(Plasteid dan Peterson 1959), komponen ragam dari interaksi genotipe x
lingkungan ( (i)) (Plaisteid (960), ecovalen (W 2i) (Wricke 1962) dan ragam
stabilitas (2i) (Shukla 1972) dapat digunakan untuk mengukur stabilitas tipe ini.
Konsep stabilitas Tipe 3, suatu genotipe cenderung stabil apabila residu kuadrat tengah (MS) dari model regresi terhadap indeks lingkungannya kecil. Indeks lingkungan digambarkan dari nilai tengah semua genotipe dari setiap lokasi dikurangi total nilai tengah semua genotipe pada semua lokasi. Tipe 3 ini juga bagian dari stabilitas dinamis atau agronomis menurut Becker dan Leon (1988). Metode yang menjelaskan stabilitas tipe 3 adalah metode Eberthart dan Russell (1966), Perkins dan Jinks (1968) dan Tai (1971). Becker dan Leon (1988) menyatakan bahwa semua prosedur stabilitas yang berdasarkan kuantitatif pengaruh interaksi genotipe x lingkungan termasuk kedalam konsep stabilitas dinamis. Lin et al. (1986) mendefinisikan empat grup stabilitas hasil yang didasarkan dari deviasi dari pengaruh rata-rata genotipe (DG) dan pola interaksi genotipe x lingkungan (GE).
Lin dan Binns (1988a) mengajukan konsep stabilitas tipe 4 yang berdasarkan variasi bukan genetik yang bisa diprediksi dan tidak diprediksi. Komponen yang bisa diprediksi berkaitan dengan lokasi sedangkan komponen yang tidak bisa diprediksi berkaitan dengan tahun. Lin dan Binns (1988a) menyarankan untuk menggunakan pendekatan regresi pada bagian yang bisa diprediksi dan kuadrat tengah (MS) dari tahun x lokasi untuk setiap genotipe sebagai perhitungan ragam yang tidak bisa diprediksi. Menurut Annicchiarico
(2002) konsep stabilitas tipe 4 memiliki pengertian yang sama dengan konsep stabilitas statis. Simmonds (1991) menyatakan bahwa stabilitas statis akan lebih banyak berguna dibandingkan dengan stabilitas dinamis pada semua kondisi, terutama untuk negara berkembang.
Tabel 1 Pembagian grup stabilitas (Lin et al. 1986)
Grup Dasar Perhitungan Sumber Ragam
Grup A DG (pengaruh rata-rata genotipe) jumlah kuadrat (SS)
Grup B GE (pola interaksi Genetipe X Lingkungan) jumlah kuadrat (SS)
Grup C DG atau GE koefisien regresi
Grup D DG atau GE deviasi regresi
Analisis Stabilitas Parametrik
Beragam metode telah banyak diajukan untuk menganalisis interaksi genotipe x lingkungan dari pengujian multi lokasi. Apabila terdapat interaksi dan terbukti nyata maka dilanjutkan dengan menganalisis stabilitas dari genotipe untuk mendapatkan genotipe yang berdaya hasil tinggi dan stabil. Alberts (2004) menyatakan bahwa secara umum metode yang tersedia untuk menganalisis interaksi genotipe x lingkungan dapat dibagi menjadi empat grup, yaitu; (1) analisis komponen ragam atau anova; (2) analisis stabilitas parametrik; (3) metode peubah ganda (multivariate); (4) metode kualitatif atau stabilitas non parametrik.
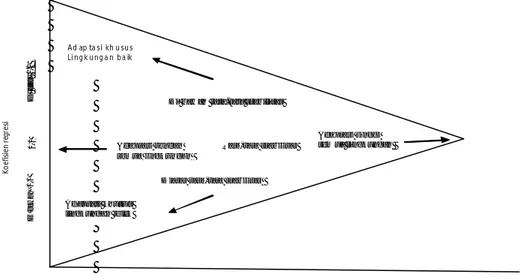
Analisis stabilitas parametrik pertama kali diajukan oleh Yates dan Cochran (1938), yang menyatakan bahwa derajat hubungan antara perbedaan varietas dan nilai tengah semua varietas dapat dijelaskan dengan menghitung regresi dari hasil satu varietas dengan nilai tengah hasil dari semua varietas. Yates dan Cochran (1938) menunjukkan regersi tersebut pada percobaan barley, tetapi ide mereka tidak teramati sampai Finlay dan Wilkinson (1963) menemukan ulang metode yang sama dan digunakan pada analisis adaptasi pada percobaan 277 varietas barley di tujuh lingkungan. Metode Finlay – Wilkinson mengukur stabilitas dan adaptasi tanaman berdasarkan regresi linear untuk setiap lokasi dan musim. Perhitungan regresi ini dijadikan sebagai dasar untuk menentukan tingkat derajat kelinearan yang timbul. Finlay dan Wilkinson (1963) menggunakan
rata-rata stabilitas. Penambahan nilai koefisien terhadap 1.0 berarti meningkatkan kepekaan adaptasi terhadap perubahan lingkungan. Penurunan koefisien berarti peningkatan adaptasi terhadap perubahan lingkungan (Gambar 2). Suatu genotipe
dikatakan sangat stabil apabila nilai koefisien regresinya (bi) = 0.
Gambar 2 Interpretasi umum dari pola populasi genotipe yang didapat ketika koefisien regresi genotipe diplot terhadap nilai tengah (rata-rata hasil) genotipe (Finlay dan Wilkinson 1963).
Eberhart dan Russell (1966) mengajukan pengabungan jumlah kuadrat dari lingkungan (E) dan interaksi genotipe x lingkungan (GE) serta membaginya ke dalam pengaruh linear antar lingkungan (derajat bebas = 1) dan pengaruh linear dari genotipe x lingkungan (derajat bebas E = 2). Pengaruh dari residual kuadrat tengah dari model regresi antar lingkungan digunakan sebagai indeks stabilitas. Suatu genotipe dikatakan stabil apabila memiliki nilai deviasi regresi kuadrat
tengah (S2di) = 0 dan memiliki nilai koefisien regresi (bi) = 1.
Perkins dan Jinks (1968) mengajukan model koefisien regresi yang mirip dengan Finlay dan Wilkinson (1963) tetapi data yang diamati disesuaikan terlebih dahulu dengan pengaruh lingkungan sebelum dibuat regresi. Menurut Roy (2000), model Perkins dan Jinks menjelaskan bahwa genotipe yang sangat stabil apabila
memiliki nilai βi = 0 dan genotipe ini tidak memiliki interaksi genotipe x
lingkungan. Genotipe dengan nilai βi > 0.0 tidak terlalu sensitif dengan
lingkungan, tetapi genotipe ini direkomendasikan khusus untuk lingkungan yang
Ad ap ta si kh u su s Li n g k u n g a n b a ik Ad ap t asi k h u su s lin g k u n g a n je lek A d a p ta si r en d a h s em u a lin g k u n g an Ad a p ta si ti n g g i sem u a li n g k u n g an R a ta- r ata sta b ilit as
D iat as r at a- r ata s ta b ilit as D i b aw ah ra ta -r at a s ta b ili ta s K o e fi si e n r eg re si 1 .0 D i b aw a h 1 .0 D i a ta s 1 .0 Ni lai ten g a h v ar ie ta s
optimal. Genotipe dengan nilai βi < 0.0 atau negatif akan lebih sedikit
perbedaannya antar lingkungan dan sangat cocok untuk ditanam pada semua lingkungan.
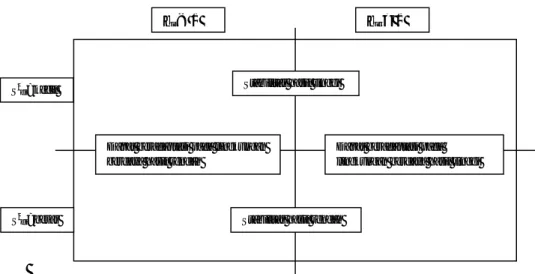
Pendekatan regresi telah menjadi alat yang berguna untuk pemulia ta-naman, tetapi ada beberapa penulis yang menunjukkan keterbatasan regresi baik secara statistik maupun biologis. Keterbatasan pertama seperti dikemukakan oleh Freeman dan Perkins (1971) serta Freeman (1973) adalah secara statistik nilai tengah genotipe tidaklah bebas dari nilai marginal lingkungan. Membuat regresi satu set peubah terhadap peubah yang lain tidak akan bebas dari saling mem-pengaruhi satu dengan yang lain. Masalah ini akan menjadi lebih besar apabila menggunakan jumlah genotipe yang banyak. Keterbatasan secara statistik yang kedua seperti yang dikemukan oleh Crossa (1990) adalah galat berhubungan dengan slope dari genotipe dan tidak secara statistik bebas, karena deviasi jumlah kuadrat dengan derajat bebas (G-1)(E-2) dapat tidak terbagi secara orthogonal antara G genotipe. Permasalahan yang ketiga seperti yang dikemukakan oleh Mungomery et al. (1974) dan Wescott (1986) adalah hubungan linear antara in-teraksi dan nilai lingkungan. Ketika asumsi ini tidak dipenuhi maka keefektifan dari analisis akan berkurang dan menyebabkan salah pengertian dari hasil analisis. Permasalahan secara biologi seperti dikemukan oleh Westcott (1986) dan Crossa (1990) adalah jika lingkungan yang diikutkan hanya memiliki sedikit lokasi yang optimal atau marjinal. Genotipe terpilih akan dijelaskan secara umum hanya cocok pada lingkungan ekstrem. Hal ini dapat menyebabkan salah pe-ngertian oleh karena itu analisis regresi dipergunakan dengan hati-hati apabila set data mengikutsertakan hasil dari sedikit lokasi yang rendah atau tinggi. Becker dan Leon (1988) mencatat ketika mempelajari metode biometrical utama, bahwa
pendekatan regresi jarang digunakan jika koefisien regresi (bi) dimasukkan ke
dalam definisi stabilitas. Dengan alasan ini, koefisien regresi (bi) oleh
keba-nyakan penulis dipandang bukan sebagai ukuran kestabilan tetapi hanya sebagai tambahan informasi dari respon rata-rata genotipe terhadap perubahan kondisi lingkungan. Skema informasi ini ditampilkan pada Gambar 3.
Gambar 3 Interpretasi parameter bi dan S2di dari pendekatan regresi.
Wricke (1962) menggunakan interaksi genotipe x lingkungan pada setiap genotipe sebagai ukuran kestabilan. Ukuran kestabilan Wricke (1962) disebut
ecovalance (W2i), yang merupakan jumlah kuadrat yang disumbangkan oleh satu
genotipe kepada interaksi genotipe x lingkungannya. Ukuran perbedaan kestabilan merupakan nilai konsistensi dari suatu genotipe pada semua lingkungan.
Geno-tipe yang memiliki nilai ecovalance (W2i) terkecil merupakan genotipe yang
pa-ling stabil.
Tai mengajukan dua parameter stabilitas, yaitu alpha () dan lamdha (), yang menyerupai koefisien regresi dan ragam regresi, tapi didapat dengan cara melanjutkan analisis ragam dan mengunakan prinsip hubungan struktural (principle of structural relationships) (Kendall dan Stuart 1979). Mempartisikan interaksi genotipe x lingkungan kedalam regresi jumlah kuadrat dan deviasi jumlah kuadrat dari regresi bisa dilakukan apabila pengaruh lingkungan dapat diukur tanpa melakukan kesalahan. Tai (1971) menggunakan metode alternatif ini dikarenakan pengaruh lingkungan tidak dapat diukur tanpa melakukan kesalahan. Berdasarkan asumsi bahwa pengaruh lingkungan dan pengaruh interaksi genotipe x lingkungan berdistribusi normal, Tai (1971) melakukan hubungan struktural menurut metode Kendall dan Stuart (1979) untuk menemukan estimasi mak-simum dari dan dari komponen yang didapat langsung dari anova. Genotipe yang paling stabil akan tidak berubah penampilannya dari lingkungan satu ke
bi < 1 bi > 1
Stabilitas hasil tinggi
Stabilitas hasil rendah Dapat beradaptasi pada lingkungan
berdaya hasil rendah
Dapat beradaptasi pada lingkungan berdaya hasil tinggi
S2di=besar S2
lingkungan lainnya. Parameter = -1 dan = 1 merupakan parameter yang nunjukkan genotipe yang paling stabil sedangkan parameter = 0 dan = 1 me-nunjukkan genotipe yang memiliki stabilitas rata-rata.
Shukla (1972) mendefinisikan ragam stabilitas genotipe sebagai ragam seluruh lingkungan setelah pengaruh utama dari nilai lingkungan dihilangkan. Karena pengaruh utama genotipe telah stabil, ragam stabilitas didasarkan pada residual matrik interaksi genotipe x lingkungan dan galat sebagai klasifikasi dua
arah. Stabilitas Shukla dinamakan sebagai ragam stabilitas (σ2i). Suatu genotipe
dikatakan stabil apabila ragam stabilitas (σ2i) adalah sama dengan ragam
ling-kungan (σ2e) dimana nilai (σ2i) = 0. Nilai (σ2i) relatif besar menunjukkan
keti-dakstabilan dari genotipe. Karena ragam stabilitas adalah perbedaan antara dua
jumlah kuadrat, maka bisa bernilai negatif. Tetapi estimasi (σ2i) negatif tidak
akan menjadi masalah pada komponen ragam karena estimasi negatif dari ragam
stabilitas (σ2i) dapat dianggap sebagai nol. Pinthus (1973) mengajukan pengunaan
koefisien determinasi (ri2) dari setiap genotipe sebagai parameter stabilitas. Suatu
genotipe dikatakan stabil apabila memiliki nilai koefisien determinasi (ri2) = 1.
Francis dan Kannenberg (1978) menggunakan ragam lingkungan (S2i) dan
koefisien ragam (CVi) untuk menentukan kestabilan suatu genotipe. Suatu
geno-tipe dikatakan stabil apabila memiliki nilai ragam lingkungan (S2i) dan koefisien
ragam (CV i) kecil serta memiliki hasil yang optimal.
Lin dan Binns (1988b) mengajukan stabilitas berdasarkan ukuran
su-perioritas genotipe (Pi) atau yang disebut sebagai stabilitas tipe 4. Superioritas
genotipe (Pi), merupakan parameter stabilitas yang sangat spesifik. Suatu
geno-tipe dikatakan stabil apabila memiliki nilai (Pi) terkecil. Menurut Lin dan Binns
(1988), stabilitas tipe 4 lebih konsisten dibandingkan dengan stabilitas tipe 3 ka-rena mengunakan ekspresi nyata dari prilaku genetik suatu genotipe. Oleh kaka-rena
itu suatu genotipe yang memiliki nilai (Pi) terkecil akan memiliki sedikit
Analisis Peubah Ganda
Menurut Crossa (1990), analisis peubah ganda memiliki tiga tujuan utama yaitu: (1) menghilangkan gangguan pola data, seperti membedakan ragam sis-tematik dengan non sissis-tematik; (2) menyimpulkan data; (3) mengungkap struktur data. Berbeda dengan metode statistik biasa, fungsi dari analisis peubah ganda adalah mengurai struktur internal data dari hipotesis yang dapat dijabarkan dan kemudian diujikan dengan metode statistik. Alberts (2004) menyimpulkan bahwa analisis peubah ganda mengutamakan analisis matrik dua arah dari genotipe dan lingkungan. Respon dari setiap genotipe pada lingkungan tertentu dapat dianggap sebagai pola ruang dimensi lingkungan, dengan koordinat masing-masing hasil dari genotipe dalam satu lingkungan. Menurut Gauch (1982) dan Crossa (1990), terdapat dua grup teknik peubah ganda yang bisa digunakan untuk mengurai struktur internal interaksi genotipe x lingkungan, yaitu :
1. Teknik ordinat, seperti principal component analysis (PCA), principal
coordinate’s analysis dan analisis faktor. Asumsi yang diperlukan adalah
data merupakan satu kesatuan. Tehnik ini menempatkan genotipe yang terpilih dan kaitannya dengan lingkungan adalah sangat mungkin berada pada ruang dimensi yang dekat.
2. Teknik klasifikasi, seperti analisis gerombol dan analisis diskriminasi.
Metode ini melibatkan kelompok yang mirip pada satu gerombol dan efektif untuk menyimpulkan kelebihan pada data.
Menurut Crossa (1990), principal component analysis sering digunakan pada metode peubah ganda (multivariate). Tujuannya adalah untuk mentrans-formasikan satu set koordinat axis ke yang lain dengan segala kemungkinannya. Konfigurasi asli dari set utama dari struktur data ditempatkan pada komponen axis utama. Principal coordinates analysis adalah generalisasi dari analisis PCA yang mana setiap ukuran kemiripan antara individu dapat digunakan. Analisis faktor masih terkait dengan PCA, faktor yang terbentuk menjadi mirip seperti principal
component. Jumlah besar peubah yang berkolerasi akan berkurang menjadi
jum-lah kecil faktor utama. Keragaman dijelaskan sebagai pola faktor umum untuk se-mua peubah dan dibuat pola dari faktor keunikan untuk setiap peubah.
Menurut De Lacy et al. (1996), analisis gerombol adalah teknik klasifikasi numerik yang mendefinisikan grup dari gerombol individu-individu. Tujuan utama dari analisis gerombol adalah untuk mengklasifikasikan obyek baik geno-tipe maupun lingkungan ke dalam grup-grup dimana telah dibuat minimal kera-gaman dalam grup, sementara kerakera-gaman antar grup dimaksimalkan. Banyak cara untuk membuat skala dan standarisasi data melalui pemusatan pada lingkungan, standarisasi lingkungan, pembobotan heritabilitas lingkungan, atau membuat ranking pada lingkungan. Beragam metode gerombol kadang kala membuat per-bedaan hasil, walaupun data yang digunakan sama. Dasar dari semua metode ge-rombol adalah mengunakan ukuran kemiripan (proximity) atau ketidakmiripan (distance) untuk mengklasifikasikan obyek ke dalam grup.
Analisis biplot AMMI dapat menjelaskan interaksi genotipe x lingkungan, menampilkan pola sebaran posisi relatif genotipe pada lingkungan. Hasil pengu-raian nilai singular diplotkan antara satu komponen genotipe dengan komponen lingkungan secara simultan. Biplot AMMI dapat meringkas pola hubungan antar genotipe, antar lingkungan, dan interaksi genotipe x lingkungan. Biplot menam-pilkan nilai komponen utama pertama dan nilai tengah. Interpretasi biplot nilai komponen pertama dan nilai tengah respon, dibuat jarak titik amatan yang ber-dasarkan sumbu datar dimana titik amatan menunjukkan perbedaan pengaruh utama amatan-amatan tersebut (Mattjik 2005).
Sumertajaya (1998) menyatakan bahwa jarak titik-titik amatan berdasar-kan sumbu tegak menunjukberdasar-kan perbedaan pengaruh interaksinya atau perbedaan tingkat sensitifitas terhadap lokasi. Sedangkan interpretasi untuk titik sejenis yang diperoleh dari biplot nilai komponen utama kedua dan nilai komponen utama pertama merupakan jarak titik-titik amatan yang menunjukkan perbedaan in-teraksi. Interpretasi titik-titik amatan yang berlainan jenis biplot nilai komponen utama kedua dan nilai komponen utama pertama menunjukkan jenis interaksi antar titik-titik amatan. Titik-titik amatan yang mempunyai arah yang sama me-nunjukkan berinteraksi positif (saling menguatkan) dan titik-titik yang berbeda arah menunjukkan berinteraksi negatif.