BAB 2
TINJAUAN PUSTAKA
2.1 Limbah Udang
Pada tahun 1993 diperkirakan dunia dapat memperoleh kembali kitin dari invertebrata laut sebanyak 37.000 ton dan meningkat menjadi 80.000 ton pada tahun 2000 (Ogawa et al., 2002). Di Indonesia, sebaran ketersediaan kulit udang mencakup Pantura Jawa,
Sumatera Utara, Lampung, Sulawesi Selatan, Tenggara dan Tengah, dan Kalimantan Timur. Volume ekspor udang (kupas dan tanpa kepala) sekitar 135 ribu ton per tahunnya dengan limbah kulit udang sekitar 60 ribu ton. Dari hasil survei Badan Riset Kelautan dan Perikanan (BRKP) diketahui bahwa dari 100 ton kulit udang mengandung sekitar 13 ton kitin (Waltam, 2009).
Pabrik pembekuan udang (cold storage) yang mengolah udang untuk ekspor dalam bentuk udang beku tanpa kepala (headless) dan kulit (peeled) menghasilkan limbah berupa kulit keras (cangkang) sekitar 50-60% yang dibuang atau hanya digunakan sebagai campuran makanan ternak (Hedriyastuti et al., 2009; Xu et al.,
2008). Limbah udang mengandung protein sekitar 30 – 40%, kalsium karbonat 30– 50% dan kitin 20–30% (Kurita, 2006). Kulit udang juga mengandung karotinoid berupa astaxantin, daging dan sedikit lemak (Zhai dan Hawkins, 2002; Gimeno et al., 2007).
Tabel 2.1. Persentase Kandungan Kitin pada Cangkang Crustaceae.
Jenis Crustaceae Kitin (%)
Kepiting Biru 14,9
Kepiting Batu 18,1
Kepiting Merah 27,6
Kepiting Ladam 26,4
Rajungan 17,0
Udang 27,2
Sumber : Austin et. al, 1981
Limbah udang merupakan bahan yang mudah busuk. Proses degradasi dilakukan oleh mikroba pembusuk dengan menghasilkan enzim-enzim pendegradasi. Adanya kandungan kitin pada limbah udang menyebabkan munculnya bakteri penghasil enzim kitinase. Organisme ini biasanya memiliki berbagai macam gen penyandi enzim kitinase yang ekspresinya diinduksi oleh kitin dan derivatnya. Hasil pemecahan polimer kitin kemudian dimanfaatkan sebagai sumber karbon dan nitrogen (Donderski dan Brzezinska, 2003).
2.2 Kitin
Kitin merupakan homopolimer linear yang tersusun dari monomer
N-asetil-D-glukosamin dengan ikatan glikosidik β-(1,4) (Gooday, 1990). Monomer N-asetil-D-glukosamin pada polimer kitin dihubungkan dengan ikatan hidrogen yang sangat kuat antara gugus –NH dari satu monomer dan gugus C=O dari monomer yang berdekatan, sehingga membentuk formasi fibril yang bersifat stabil dan kaku (Gambar 2.2). Kitin bersifat tidak larut air dan hanya larut dalam pelarut asam mineral pekat seperti HCl
(Herdyastuti et al., 2009). Berdasarkan penyusun rantai polimernya, kitin fibril
Gambar 2.2 Struktur kimia kitin
Setelah selulosa, kitin merupakan biopolimer yang paling banyak ditemukan di alam dan terdistribusi di lingkungan biosfer. Kitin merupakan polimer linier yang tersusun dari 2.000–3.000 monomer N-asetil-D-glukosamin yang dihubungkan
dengan ikatan β-1,4-glikosida. Monomer dari kitin dimanfaatkan oleh mikroorganisme di alam, antara lain bakteri, sebagai sumber karbon dan nitrogen untuk nutrisi (Donderski dan Brzezinska, 2003). Kitin merupakan komponen utama pada kutikula serangga, dinding sel jamur, yeast dan alga hijau (Enibu dan Varum, 2008; Sato et al.,1998; Yoshihiro et al., 2008). Penyumbang kitin terbesar di tanah adalah jamur dengan jumlah sekitar 500-5000 kg/ha (Shahidi dan Abuzaytoun, 2005). Sedangkan pada lingkungan perairan, kitin banyak ditemukan pada kulit kepiting dan udang (Wang dan Xing, 2007). Sumber komersial kitin terbesar didapat dari kulit crustaceae seperti udang, kepiting, lobster dan lainnya yang berasal dari proses pengolahan udang (Arbia et al., 2012).
Kitin memiliki sifat yang unik, yaitu; biodegradable, biocompatible dan tidak toksik. Hal ini menyebabkan kitin dan derivatnya banyak digunakan dalam berbagai macam aplikasi dalam industri dan biomedik sehingga kitin dan derivatnya memiliki nilai ekonomi yang besar (Synowiecki dan Al-Khateeb, 2000; Wang et. al, 2010; Arbia et al., 2012). Misalnya digunakan dalam bioremediasi logam berat (Sirait, 2002),
2.3 Enzim Kitinase dan Pemanfaatannya
Kitinase dapat dihasilkan oleh beberapa mikroorganisme dan mempunyai peran
penting pada fisiologi dan ekologi. Semua enzim yang dapat mendegradasi kitin disebut sebagai kitinase total atau kitinase non spesifik. Enzim kitinase dibagi menjadi tiga, yaitu: (i) Eksokitinase atau kitobiosidase, mengkatalisis pembebasan
N-asetil-glukosamin atau unit dimmer kitobiosa (β-1,4-N-asetil-glukosamin); (ii) Endokitinase (EC.3.2.1.14) enzim yang mendegradasi kitin secara acak dari dalam menghasilkan
oligomer pendek N-asetil-glukosamin; (iii) N-asetilglukosaminidase (EC.3.2.1.30) bekerja pada pemutusan diasetilkitobiosa menghasilkan N-asetil-glukosamin (Tronsmo dan Harman, 1993).
Keberadaan kitin di alam yang sangat melimpah ini dengan cepat terdegradasi, karena banyaknya bakteri dan fungi yang mempunyai enzim kitinase yang mampu
mendegradasi kitin. Degradasi kitin secara enzimatis memiliki dua alternatif lintasan degradasi yaitu dengan mekanisme kitinase dan mekanisme kitosanase. Lintasan
dengan kitinase mekanismenya dengan menghidrolisis ikatan glikosidik β-(1,4) dan mekanisme ini sering disebut sebagai kitinolitik. Lintasan degradasi kitin yang kedua yaitu mekanisme pengubahan kitin oleh deasetilase kitin menjadi kitosan. Pada
mekanisme ini enzim kitosanase akan menghidrolisis ikatan glikosida β-(1,4) pada kitosan dan menghasilkan diasetilkitobiosa (kitobiosa) yang kemudian dihidrolisis
kembali oleh β-Nasetilglukosaminidase menjadi glukosamin (Gooday, 1990).
Kitinase dapat diperoleh dari mikroorganisme perairan, misalnya dari danau, laut dan kolam. Pada lingkungan biosfer juga tersedia kitinase yang melimpah yang dihasilkan dari berbagai macam organisme (Herdyastuti et al., 2009). Limbah pengolahan pabrik udang juga merupakan sumber kitinase karena banyak didapatkan kitin sebagai substratnya. Limbah udang mempunyai kandungan jumlah jamur lebih sedikit daripada bakteri, tetapi jamur dapat menghasilkan kitinase lebih banyak daripada bakteri (Brzezinska et al., 2007).
manfaatnya. Enzim kitinase berperan penting dalam kontrol jamur patogen tanaman
secara mikroparasitisme (Nugroho et al., 2003). Kitinase banyak dimanfaatkan sebagai agen biokontrol terutama bagi tanaman yang terserang infeksi jamur. Hal ini berhubungan dengan komponen utama dinding sel jamur yang tersusun atas kitin. Kitin pada dinding sel jamur patogen dapat didegradasi oleh enzim kitinase dan menghasilkan produk yang ramah lingkungan dibandingkan penggunaan zat kimia (Herdyastuti et al., 2009). Natsir et al. (2012) melaporkan bahwa produksi kitinase ekstraseluler dari Bacillus licheniformis HSA3-1a dapat menghidrolisis limbah udang dan dinding sel jamur Ganoderma sp. penyebab busuk batang pada kelapa sawit. Selain itu kitinase juga berperan dalam proses pembuatan senyawa kitosan melalui proses deasetilase secara termokimia. Kitinase juga dapat dimanfaatkan dalam penanganan limbah terutama limbah yang mengandung kitin berasal pabrik pembekuan udang. Pabrik tersebut menghasilkan limbah cangkang udang yang bila dibiarkan dapat menyebabkan pencemaran lingkungan sehingga meningkatkan BOD dan COD (Herdyastuti et al., 2009). Dibandingkan kitin, aplikasi kitosan lebih luas dan dapat ditemukan pada berbagai bidang seperti industri pangan, pengolahan limbah, kesehatan, bioteknologi, pertanian, kosmetik, dan industri kertas (Rahayu et al., 1999).
2.4 Mikroorganisme Penghasil Kitinase
Mikroorganisme kitinolitik adalah mikroorganisme yang dapat mendegradasi kitin dengan menggunakan enzim kitinase. Mikroorganisme ini dapat diperoleh dari berbagai sumber seperti rizosfer, filosfer, tanah atau dari lingkungan air seperti laut, danau, kolam, tambak udang atau limbah udang dan sebagainya (Ilmi, 2007; Suryanto, 2011; Asril, 2011; Dewi, 2011; Fauziah dan Herdyastuti, 2013). Selain lingkungan mesofil, mikroorganisme kitinolitik juga dapat diisolasi dari lingkungan termofilik seperti sumber air panas, daerah geotermal dan lain-lain (Dewi, 2008; Herdyastuti et al., 2009) dan lingkungan yang asam (Natsir, 2002).
laporan diketahui bakteri yang mampu menghasilkan kitinase diantaranya adalah dari
genus Bacillus (B. pumilus, B. firmus, B. cereus) Streptomyces hygroscopicus, Streptomyces griceus, Serattia marcescens, Vibrio spp., Flavobacterium sp.
Alcaligenes denitrificans, Actinomycetes, Pseudomonas stutzeri, Escherechia coli dan
genus Sanguibacter (Roberts dan Selitrennikoff, 1987; Yong et al., 2005; Ilmi, 2007; Herdyastuti et al., 2009; Muharni dan Widjajanti, 2011; Priya et al., 2011; Suryanto et al., 2011).
Mikroorganisme kitinolitik dapat diseleksi keberadaannya dengan mendegradasi media agar kitin yang dapat dideteksi dengan adanya zona bening di sekitar koloni bakteri. Penggunaan media yang mengandung kitin, misalnya koloidal kitin dapat menginduksi kitinase pada bakteri, jamur dan aktinomisetes. Substrat ini
mampu menginduksi enzim hidrolitik seperti β-1,4-N-asetilglukosaminidase, endokitinase dan kitobiosidase (Inbar dan Chet, 1991).
Bakteri kitinolitik memiliki berbagai peran di alam, antara lain mempertahankan siklus karbon pada lingkungan yang kaya kitin, seperti ekosistem perairan (Donderski dan Trzebiatowska, 1999; Donderski dan Brzezinska, 2001; Metcalfe et al.,2002). Selain itu bakteri kitinolitik juga menyebabkan penyakit pada crustaceae, terutama udang dan kepiting. Bakteri tersebut mendegradasi eksoskeleton crustaceae sehingga terjadi kerusakan dan mempermudah terjadinya infeksi pada jaringan tubuh yang berada di bawah eksoskeleton (Guzman dan Valle, 2000; Vogan
et al., 2002).
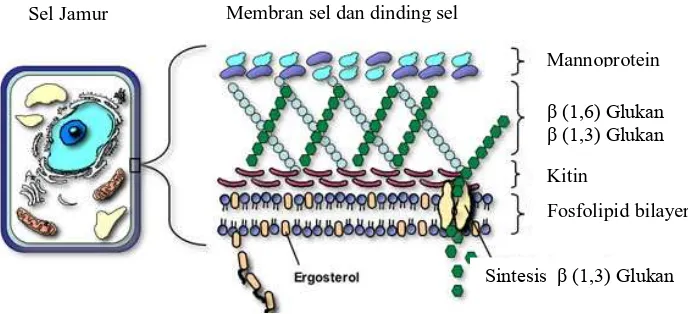
2.5 Dinding Sel Jamur
Gambar 2.5 Komponen Dinding dan Membran Sel Jamur
Sekitar 80% penyusun dinding sel jamur terdiri atas polisakarida. Sedangkan protein hanya memiliki bagian yang kecil dari seluruh komponen penyusun dinding sel jamur yaitu sekitar 20% dan biasanya dalam bentuk glikoprotein. Namun, dalam proses mating, modifikasi hifa dan pengambilan nutrisi melibatkan protein yang terikat pada dinding sel. Lipid juga terdapat dalam jumlah yang sangat sedikit pada dinding sel jamur yang berfungsi untuk mengontrol perpindahan jamur pada air khususnya dalam mencegah terjadinya kerusakan sel. Dinding sel jamur juga mengandung pigmen (misalnya melanin) dan garam-garam, namun hanya dalam jumlah yang sangat sedikit. Melanin berperan penting dalam melindungi hifa dan spora dari sinar UV terutama untuk patogenesis dan berhubungan dalam memunculkan hifa baru dari spora (Gow dan Gadd, 1995).
Dinding sel jamur mempunyai struktur yang dinamis dan mengalami perubahan secara konstan yang dipengaruhi oleh berbagai kondisi dan tempat tumbuhnya (Bowman dan Free, 2006; Adams, 2004). Misalnya selama perbesaran dan pembelahan pada yeast, pembentukan spora, percabangan hifa dan pembentukan septat pada jamur berfilamen. Percabangan dan cross-linking dari dinding sel jamur, dan juga pengaturan plastisitas dinding sel selama morfogenesis dipengaruhi oleh aktivitas enzim hidrolitik yang ditemukan pada dinding sel (Adams, 2004).
Kitin merupakan homopolimer dari β-1,4-N-asetilglukosamin yang merupakan komponen penting dalam dinding sel jamur. Namun kandungannya hanya sekitar
1-Sel Jamur Membran sel dan dinding sel
Mannoprotein
β (1,6) Glukan β (1,3) Glukan
Kitin
Fosfolipid bilayer
2% pada yeast dan 10-20% pada jamur berfilamen, misalnya Neurospora sp. dan
Aspergillus sp. Kitin pada dinding sel jamur berbentuk mikrofibril yang terbentuk dari
ikatan hidrogen antar rantai. Polimer kristalin ini memiliki kekuatan tarik yang sangat besar dan secara signifikan berkontribusi pada integritas keseluruhan dinding sel. Ketika sintesis kitin terganggu, dinding sel menjadi tidak teratur yang menyebabkan bentuk sel jamur menjadi tidak normal dan kondisi osmotiknya tidak stabil (Bowman dan Free, 2006). Pada kelompok Oomycetes, dinding selnya sebagian besar terdiri atas
polimer β-1,3 glukan dan selulosa. Tidak seperti dinding sel jamur yang lainnya, kandungan kitin pada kelompok ini sangat sedikit. Meskipun demikian, gen kitin sintase menyebar secara meluas pada jenis-jenis Oomycetes dan kitin sintase ini menjadi penghambat polyoxin D yang menyebabkan penurunan pertumbuhan Saprolegnia secara signifikan. Hal ini menunjukkan bahwa kitin merupakan
komponen yang penting pada dinding sel Oomycetes walaupun jumlanya sangat sedikit (Kamoun, 2003). Komponen penyusun dinding sel dari masing-masing divisi jamur dapat dilihat pada Tabel 2.5.
Tabel 2.5 Komponen Penyusun Dinding Sel Jamur (Gow dan Gadd, 1995)
Divisi Fibrous Polimer
Basidiomycetes Kitin
Glukan merupakan polisakarida struktural utama dari sebagian besar jamur. Jumlah glukan pada dinding sel jamur sekitar 50-60% dari berat keringnya. Polimer gluka n terdiri atas kumpulan residu glukosa yang membentuk rantai melalui berbagai ikatan kimia. Secara umum, antara 65-90% dari glukan yang ditemukan pada dinding
Semua dinding sel jamur memiliki komponen protein yang berikatan kuat
dalam kitin dan glukan yang tesusun dalam struktur matriks. Kandungan protein pada dinding sel S. cerevisiae dan C. albicans dinding sel sekitar 30-50% dari berat kering. Jumlah protein dalam dinding sel jamur berfilamen telah diperkirakan mewakili sekitar 20-30% dari total dinding sel. Perhitungan secara empiris pada dinding sel hifa Neurospora crassa didapatkan kandungan protein sekitar 15% dari total berat
keringnya (Bowman et al., 2006).
2.6Mekanisme Kerja Agen Biokontrol terhadap Patogen
Agen biokontrol yang biasa digunakan dalam mengatasi pertumbuhan patogen adalah jamur dan bakteri. Ada tiga tipe antagonisme yang dilakukan oleh jamur dalam mengatasi patogen, yaitu (1) antagonisme secara langsung, (2) antagonisme tidak langsung dan (3) kombinasi antara antagonisme langsung dan tidak langsung. Pada antagonisme secara langsung jamur mampu memparasiti dan membunuh patogen. Jamur agen biokontrol mampu berpenetrasi dan merusak spora patogen. Antagonisme tidak langsung, tidak ada kontak fisik antara jamur dengan patogen. Jamur agen biokontrol akan meningkatkan level resistensi dengan mengaktifkan mekanisme pertahanan dari inangnya. Sebagian besar jamur sudah menunjukkan kemampuannya dalam menginduksi resistensi inangnya dalam melawan patogen. Selain itu persaingan nutrisi juga merupakan salah satu cara yang dilakukan oleh agen biokontrol untuk melindungi inangnya dari serangan patogen. Kombinasi antagonisme langsung dan tidak langsung yaitu dengan cara menghasilkan enzim, antibiotik ataupun metabolit toksik lainnya untuk menghambat patogen (Narayanasamy, 2013).
Penelitian jamur sebagai agen biokontrol telah banyak dilakukan diantaranya oleh El-Katatny et al. (2000) yang menggunakan Trichoderma harzianum dalam mengontrol pertumbuhan Sclerotium rolfsii penyebab penyakit busuh batang dan akar
Gambar 2.6.1 Aktivitas Mikoparasit oleh T.virens terhadap R. solani. (a) Hifa T. virens membentuk haustoria dan (b) hifa R. solani (Howell, 2003)
Bakteri sebagai agen biokontrol memiliki mekanisme antagonisme yang tidak jauh berbeda dengan jamur sebagai agen biokontrol. Bakteri agen biokontrol mengatasi patogen melalui dua cara yaitu secara langsung dan tidak langsung dengan mekanisme yang berbeda seperti antibiosis, kompetisi nutrisi, kolonisasi pada tempat yang spesifik untuk mengatasi infeksi patogen dan menginduksi resistensi inangnya dari patogen dengan cara mengaktifkan sistem pertahanan inangnya. Sebagai contoh, Plant-growth promoting rhizobacteria (PGPR) yang telah diketahui dapat
meningkatkan pertumbuhan tanaman dan juga melindungi tanaman dari serangan mikroba patogen tanaman. Pada mekanisme biokontrol ada tiga komponen yang berperan yaitu tanaman, bakteri sebagai agen biokontrol dan patogen tanaman (Narayanasamy, 2013).
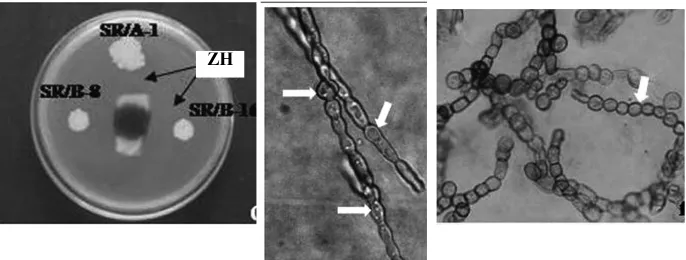
Bacillus subtilis yang digunakan untuk menghambat pertumbuhan Curvularia
gudauskasii penyebab penyakit pada bibit tebu menunjukkan adanya mekanisme
penghambatan dengan metabolit yang dikeluarkan oleh B. subtilis. Mekanisme ini dapat dilihat dari adanya zona hambat yang terbentuk pada uji in vitro dan adanya keabnormalan bentuk hifa pada pengamatan mikroskopi. Hifa mengalami pembengkokan dan membengkak (Gambar2.6.2) (Raton et al., 2012). Penggunaan bakteri sebagai agen biokontrol juga telah dilakukan untuk menghambat pertumbuhan Fusarium oxysporum f. sp. cucumerinum, Phytophthora capsici, Botrytis cinerea,
Scopulariopsis sp. dan Ganoderma boninense (Chae et al., 2006; Robert et al., 2007;
Singh, 2008; Suryanto et al., 2012 dan Novitasari, 2013). a
Gambar 2.6.2 Mekanisme Penghambatan C. gudauskasii oleh B.subtilis. : Hifa C. gudauskasii membengkak dan membengkok ZH : Zona Hambat (Raton et al., 2012)