SEN YAW A KH I TI N D AN KAJI AN AKTI V I TAS EN ZI M M I KROBI AL PEN D EGRAD ASI N YA
YURN ALI ZA, S.Si., M .Si.
Fa k u lt a s M a t e m a t ik a D a n I lm u Pe n ge t a h u a n Ala m Pr ogr a m St u di Biologi
Un ive r sit a s Su m a t e r a Ut a r a
PEN D AH ULUAN
Khit in m erupakan hom opolim er dari β- 1,4 N- set il- D- glukosam in dan m erupak an polim er k e dua t erbany ak di alam set elah selulosa. Seny aw a ini dapat dit em uk an pada cangk ang udang k epit ing, Molusca, seranggga, Annelida, dan beberapa dinding sel j am ur dan alga. Meskipun sum ber k hit in di alam berm acam -m aca-m , na-m un sa-m pai saat ini su-m ber ut a-m a y ang prak t is dieksplorasi adalah cangkang udang yang secara ekonom is pot ensial.
Khit in m em iliki bent uk yang padat dan bersifat t idak larut dalam air at au pelarut organik biasa. Nam un k hit in dapat dim odifik asi secara k im iaw i m enj adi t urunan- t urunannya yang m em punyai sifat - sifat khas dan kegunaannya sendiri. Khit in dapat dihidrolisis secara enzim at is oleh enzim khit inase, m enghasilkan m onom er β- 1,4 N- set il- D- gluk osam in. Khit inase dapat dihasilkan oleh beberapa m acam bak t eri,, ak t inom iset es, j am ur dan t um buhan. Khit inase j uga disint esis oleh prot ozoa, saluran pencernaan nem at oda, polikhaet a dan Molusca. Khit inase j uga dit em ukan dalam lendir pencernaan burung- burung pem akan seranga, lendir pencernaan dan pangk reas ikan, am fibia dan rept il pem ak an serangga. Meskipun khit inase t ersebar luas di alam ,nam un khit inase m urni secara kom ersial diproduksi dari m ik roorganism e.
I . SEN YAW A KH I TI N
A. Se j a r a h pe n e m u a n Kh it in
Pada t ahun 1811, seorang ilm uan Prancis Hendri Baraconnot berhasil m engisolasi khit in unt uk pert am a kali dari j am ur, dan diberi nam a fu n gin e . Kem udian t ahun 1823, Odier m enem ukan senyaw a yang sam a dari kut ikula serangga dan diberi nam a khit in ( bahasa Yunani) yang art inya sam pul at au baj u ( Wint erow d & Sanford, 1995) . Odier m enduga bahw a pada m ushroom dan serangga m em iliki kom ponen st rukt ural yang sam a ( Richards, 1951) . Tahun 1859 Rouget m enem uk an m odifik asi k hit in y ang ak hirny a oleh Hoppe- Seiler ( 1894) diberi nam a khit osan. Sej ak saat it u penelit ian khit in dan khit osan berkem bang sam pai pert engahan abad 1900- an. Tahun 1930- an G.W. Rigby, karyaw an Dupont m enpat ent an khit in dan khit osan besert a cara isolasi dan preparasinya dari kulit udangg sert a pem anfaat anny a dalam indust ri.
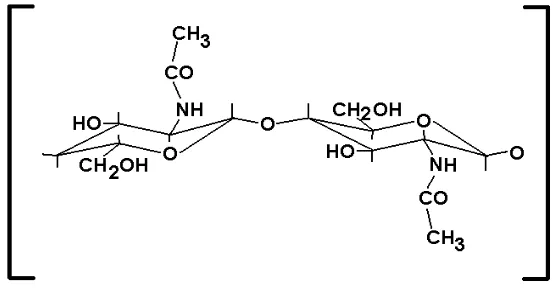
B. St r u k t u r da n Su m be r – Su m be r Kh it in di Ala m
paralel, sedangkan γ- k hit in fibrilny a m asing- m asing t ersusun dari t iga rant ai, dua rant ainy a t ersusun paralel dan rant ai k et iga ant iparalel ( Cabib, 1987) .
Gam bar 1. Unit k hit in ( Cabib, 1987) .
Khit in berbent uk padat , am orf, t idak berw arna, t idak larut dalam air, asam
encer, alk ohol dan sem ua pelarut organik lainny a, t et api k hit in dapat larut dalam fluoroalk ohol dan asam m ineral pek at ( Richards, 1951) . Koloidal k hit in adalah k hit in y ang bany ak digunak an sebagai subst rat dalam m edium ferm ent asi. Seny aw a ini diperoleh dengan m enghidrolisis secara parsial khit in dengan larut an asam klorida ( HCl) 10 N ( I nbar & Chet , 1991 ; Chernin et al., 1995 ; Haran et al., 1995) .
Khit in m erupakan polisakarida kedua t erbanyak di alam set elah selulosa dan di alam set iap t ahunnya dihasilkan sekit ar 108 t on khit in ( Muzzarelli, 1985) . Khit in dapat dit em uk an pada eksoskelet on serangga, m olusca, Coelent erat a, Nem at oda, Crust aceae dan dinding sel j am ur dan alga t ert ent u.
Khit in pada j am ur berbent uk fibril yang m em iliki panj ang yang berbeda t ergant ung pada spesies dan lok asi selny a. Pada Saccharom yces cereviceae, m ikrofibril khit innya m em iliki panj ang 60 nm dan t erdapat khusus pada sekat prim er ( Cabib, 1987) . Pada serangga lebih dari 80 % k om ponen k ut ik ulany a adalah k hit in ( Berk eley , 1979) . Pada Crust aceae, k hit in m elek at pada suat u m at riks dari CaCO3 dan fosfat . Pada serangga m at riksny a adalah prot einaceous yait u suat u prot ein yang sudah m engalam i pent aninnan ( Cabib, 1987) .Khit in pada alga t erut am a dit em uk an pada diat om ae laut y ait u Thallassiosira fluv iat ilis dan Cyclot ella crypt ica dengan k andungan 10 – 15 % berat k ering.
Pada bak t eri, ak t inom iset es dan j am ur k las Oom y cet es t idak dit em uk an khit in. Tet api khit in m erupakan kom ponen ut am a penyusun dinding sel j am ur klas Basidiom ycet es, Phycom ycet es, Ascom ycet es dan Lichens.
Meskipun sum ber khit in berm acam - m acam , nam un secara kom ersial khit in dieksplorasi dari cangkang udang- udangan dan Crst acea. Sebanyak 50 – 60 % dari lim bah udang, dihasilkan 25 % Khit in dari 32 % berat k ering lim bah t ersebut .
Kht in m erupakan bahan dasar uunt uk bahan- bahan kim ia yang diperlukan secara luas di berbagai bidang sepert i biokim ia, obat - obat an, pangan, gizi, enzim ologi, indust ri kert as, t ekst il dan film . Khit in j uga sebagai sum ber N-aset ilgluk osam in y ang dipak ai sebagai pengaw et dan ant ibiot ik. Khit osan sebagai derivat khit in dipakai dalam pengolahan lim bah dan pengikat an logam ( Muzarelli, 1985) .
C. Biosin t e sis Kh it in pa da Ja m u r
[image:2.612.135.414.147.291.2]k om ponen dinding sel. Langk ah- langk ah biosint esis k hit in diaw ali dengan perubahan glukosa- 6- fosfat m enj adi uridin difosfat N- aset ilglukosam in ( UDP- GlcNAc) sebagai prekursor k hit in ( Gam bar 2) . Enzim y ang berperan dalam m engubah UDP- GlcNAc m enj adi m olekul khit in adalah khit in sint et ase yang t erdapat dalam vesikel sit oplasm ik ( sit osom ) ( Carlile dan Wat kinson, 1994) .
Pada j am ur sebagaim ana organism e euk ariot ik lain, v esik el sit oplasm ik ny a berperan dalam pem bent uk an dinding sel. Vesik el ini m em baw a m at erial perm uk aan berupa prekursor, produk at au enzim y ang dibut uhk an unt uk pem bent uk an dinding. Ada dua v esik el y ang berperan pada pem bent uk an dinding sel j am ur y ait u 1) . Makrovesikel disebut j uga vesikel dinding at au vesikel sekret ori. Vesikel ini berperan dalam sint esis polim er polisakarida nonfibrilar. 2) . Mikrovesikel at au disebut j uga sit osom , berperan dalam sint esis polisakarida m ikrofibrl ( khit in) .
Khit in dibuat in- sit u y ait u pebent uk anny a t erj adi di luar sit oplasm a oleh enzim y ang dibaw a oleh sit osom . Khit in sint et ase y ang ada dalam sit osom adalah enzim y ang t idak ak t iv ( zim ogen) dan prot ease y ang ada di perm uk aan sel ( periplasm a) akan m engakt ivkan zim ogen ini dan dim ulai lagi pem bent ukan m ikrofibril khit in ( Bart nick i- Garcia, 1989) .
Gam bar 2. Jalur sint esis khit in ( ) dan t eori siklus N- aset ilglukosam in ( - - > ) 1) Enzim fosfo- gluko- isom erase; 2) glut am ine- frukt ose- 6- fosfat - am
ino-t ransferase; 3) gluk osam ine- fosfaino-t e- aseino-t il- ino-t ransferase; 4) aset il- glucosam ine fosfom ut ase; 5) UDP- aset il- gluk osam ine
[image:4.612.128.440.82.539.2]I I . EN ZI M KH I TI N ASE
Khit inase [ Poli { 1,4-β ( 2 aset am ido–2–deok si–D- gluk osam inide) } glikano hidrolase] adalah enzim y ang m enghidrolisis ikat an β- 1,4- aset am ido–2- deoksi- D- glikosida dari khit in dan khit odekst rin ( Bielka et al., 1984) .
A. Ta t a n a m a e n z im k h it in a se .
Sist im t at a nam a enzim khit inase m asih banyak m enim bulkan kerancuan. Harm an et al. ( 1993) sert a Sahai dan Manocha ( 1993) m em bagi k hit inase dalam t iga t ipe y ait u :
a. Endok hit inase ( EC. 3.2.1.14) y ait u : enzim y ang m em ot ong secara acak ikat an β- 1,4 bagian int ernal m ikrofibril khit in ( Gam bar 3) . Produk akhir yang t erbent uk berupa oligom er pendek N- aset ilgluk osam in ( GlcNAc) y ang m em puny ai berat m olek ul rendah sepert i k hit ot et raose, k hit ot riose dengan didom inasi oleh di-aset ilkhit obiose. Produk y ang dihasilkan bersifat m udah larut .
b. Eksokhit inase ( belum m em iliki nom or ent ry dalam Enzy m e Nom enclat ure) dinam ak an j uga k hit obiosidase at au k hit in–1,4-β- khit obiosidase, yait u enzim yang m engkat alisis secara akt iv pem bebasan unit unit diaset ilkhit obiose t anpa ada unit -unit m onosakarida at au oligosakarida yang dibent uk ( Gam bar 3) . Pem ot ongan hanya t erj adi pada uj ung non reduk si m ik rofibril k hit in dan t idak secara acak .
Ak t iv it a s e n z im
Gam bar 3. Mekanism e kerj a enzim eksokhit inase dan endokhit inase ( Sahai & Manocha, 1993)
c. β- 1,4–N aset ilglukosam idase ( EC. 3.2.1.30) adalah suat u enzim khit inolit ik y ang bek erj a pada pem ut usan diaset ilkhit obiose, k hit ot riose dan k hit ot et raose dengan m enghasilkan m onom er- m onom er GlcNAc.
B. M e t ode pe m e k a t a n ca ir a n e n z im da n pe n gu k u r a n a k t ivit a s k h it in a se
[image:5.612.166.403.335.571.2]k ult ivasi. Pem ek at an dapat dilak uk an dengan beberapa cara y ait u : ult rafilt rasi, liofiliasi dan m engendapk an prot ein dengan am m onium sulfat , aset on, et anol, at au poliet ilen glikol ( PEG) ( Scopes, 1994) .
Pem ek at an enzim k hit inase dari St rept om y ces dengan am m onium sulfat pada kej enuhan 70 % dan et anol dingin, dapat m eningkat kan kem urnian enzim bert urut - t urut 5 dan 3,5 kali dibanding enzim kasarnya ( Lloyd et al., 1965) . Singh et al. ( 1999) m eny at ak an bahw a prot ein k hit inase dari St rept om y ces sp. 385 y ang dipek at k an dengan poliet ilen glikol ( PEG) , k em urnianny a m eningk at sebany ak 11,9 k ali dibanding enzim k asarny a.
Pengendapan prot ein dengan am m onium sulfat adalah cara yang paling bany ak digunak an. Hal ini disebabk an k arena am m onium sulfat m udah didapat k an, hargany a relat if m urah, bersifat m enst abilkan enzim sert a dapat m encegah ak t iv it as enzim prot eolit ik. Garam am m onium sulfat k onsent rasi 2 - 3 M dapat m enst abilkan enzim selam a beberapa t ahun. Kelem ahannya adalah t idak dapat m engendapkan seluruh prot ein y ang t elah larut dan bila m engandung logam m ak a ak an dapat m erusak enzim ( Scopes, 1994) .
Penguk uran ak t iv it as k hit inase dalam m em ecah k hit in dapat dilak uk an dengan beberapa cara sepert i y ang disebut k an dalam Jeaniaux ( 1966) dan Cabib ( 1987) yait u :
a. Berdasark an pengurangan subst rat .
1) . Met ode viskosim et ri yait u akt ivit as khit inase t erhadap khit osan, glikol khit in at au karboksim et ilkhit in yang dit unj ukkan oleh t erj adinya pengurangan v iskosit as subst rat .
2) . Met ode t urbidim et ri ( nephelom et ri) yait u m engukur variasi t urbidit as suspensi koloidal khit in selam a khit inolisis. Pengukuran ini bersifat cepat dan ak urat t api t idak cocok unt uk enzim dengan ak t iv it as rendah. Cont oh pada penguk uran ak t iv it as enzin endok hit inase. Unit ak t iv it as enzim endok hit inase diuk ur sebagai persen pengurangan k erapat an at au t urbidit as relat if dari suspensi yang sam a ant ara yang berisi enzim dengan ak uades. Sat u unit endok hit inase didefinisik an sebagai j um lah enzim yang dibut uhkan unt uk m ereduksi t urbidit as suspensi khit in 5 % ( Harm an et al., 1993 ; Chernin et al., 1995) .
b. Berdasark an pem bent uk an produk ak hir y ait u GlcNAc ( Met ode Reissig, 1955) .
GlcNAc yang dibebaskan dari khit in dit ent ukan secara kolorim et rik dengan p- dim et ilam inobenzaldehida. Sat u unit ak t iv it as k hit inase diny at ak an sebagai
µm ol GlcNAc y ang dibebaskan selam a 1 j am dalam k ondisi y ang dit et apk an. c. Spect rom et er Assay yait u m enggunakan krom ogen 3,4, dinit rophenil t et ra
N- aset ilkhit ot et raose. d. Radiom et er Assay.
I I I . M I KROORGAN I SM E PEN GH ASI L KH I TI N ASE
Sebagian besar m ik roorganism e t anah dan air adalah pendegradsai k hit in yang baik. Di dalam t anah set iap gram nya m engandung 106 m ikr oor ganism e yang m am pu m engolah khit in. Sebanyak 90- 99 % dari organism e t ersebut adalah ak t inom iset es dan selebihny a bak t eri dan j am ur. Genus ak t inom iset es y ang dapat m em anfaat kan khit in sebagai sum ber karbon dan nit rogennya yait u : St rept om yces, Nocardia, St rept osporangium , Microm onospora dan Act inoplanes ( Alex ander, 1977) .
ant ibiot icus ( Jeaniaux , 1966) , S. alboflav us, S. albus, S. rubriret iculi, S. griseus, S. scabies ( Wak sm an, 1967) , S. orient alis ( Tom inanga & Tsuj isak a, 1976) , S. liv idans ( Miyashit a et al., 1991) , S. oliv aceov iridis ( Schrem pf, 1995) dan S. t endae ( Borm ann, 1999) .
Jenis bakt eri dan j am ur penghasil khit inase dapat dilihat pada Tabel 1 .
Ta be l 1 . M ik r oor ga n ism e pe n gh a sil k h it in a se
Ba k t e r i Ja m u r
Vibrio parahaem alut icus Vibrio alginoly t icus
Flavobact erium indolt hecium Serrat ia m arcencens
Ent erobact er liquefaciens Bacillus cereus
Klebsiella sp
Micrococcus colpogenes Bacillus cereus
Clost ridium sp Pseudom onas sp Aerom onas sp
Mort ierella sp Mucor subt illisim um Aspergillus fum igt us Trichoderm a viridae
Pada j am ur berfilam en dan k am ir, k hit inase m erupak an bagian int egral dalam proses m orfogenesis dinding sel. Selain it u khit inase j uga m engaw ali perist iw a int erak si inang- parasit dari m ik oparasit biot rop dan nek rt ot rop, j am ur ent om opat ogen dan j am ur Vesik ular Arbuskular Mik orhiza ( VAM) .
Khit inase yang dihasilkan m ikroorganism em em iliki berat m olekull yang berkisar ant ara 20.000 – 120.000. Pada bak t eri berat m olek ulny a ant ara 60.000 – 110.000, sedangk an ak t inom iset es y ait u 30.000 at au lebih rendah. Pada j am ur berat m olek ulny a lebih t inggi dari 30.000.
A. Biosin t e sis En z im Kh it in a se Pa da M ik r oor ga n ism e
Pengat uran biosint esis enzim khit inase m elalui sist em represor- induser. Khit in dan produk hasil degradasiny a ( oligom er/ m onom er) berperan sebagai induser sedangk an subst rat sepert i selulosa, x ilan, pek t in, lignin dan sebagainy a t idak dapat m enginduk si k hit inase ( Sahai & Manocha, 1993) . Gluk osam in dapat m enginduk si k hit inase k arena pada k hit osan ( k hit in y ang m engalam i deaset ilasi) m asih t erdapat sek it ar 10 – 20 % residu aset il ( Sahai & Manocha, 1993) .
Pengat uran sint esis khit inase dipengaruhi j uga oleh produk akhir ( kat abolit ) berupa GlcNAc dan gluk osa. Kedua j enis gula sederhana ini m erupak an represor k at abolit bagi spesies Trichoderm a harzianum ( Ulhoa & Peberdy , 1993) dan St achy bot ry s elegans ( Tw eddell et al., 1994) . Pada St rept om y ces liv idans kehadiran glukosa dalam m edium m engham bat sint esis enzim khit inase ( Sait o et al., 1998) .
Khit in yang dipreparasi dengan hidrolisis parsial dengan HCl 10 N akan m enghasilkan koloidal khit in yang m am pu m enginduksi khit inase kom pleks sepert i N- aset ilglukosam inidase, endokhit inase dan khit obiosidase pada Aerom onas caviae ( I nbar & Chet , 1991) , Ent erobact er agglom erans ( Chernin et al., 1995) dan Trichoderm a harzianum ( Haran et al., 1995) .
gluk osam in dan GlcNAc relat if y ang dibebaskan selam a degradasi k hit in. Kehadiran k om ponen lain sepert i prot ein y ang berik at an k ov alen dengan part ikular k hit in dapat dikenal secara langsung oleh sensor prot ein m em bran paling luar. Unt uk m em isahkan enzim prot eolit ik ini, S. oliv aceov iridis m enghasilkan khit inase dengan dom ain enzim at ik t am bahan sepert i prot ease. S. oliv aceov iridis m enghasilkan 59 kDa prot ein dan t erdiri at as 47 kDa khit inase dan 12 kDa prot einase. Jadi efisiensi penggunaan k hit in j uga m em but uhk an beberapa enzim selain k hit inase ( Schrem pf, 1995) .
B. Sist im Kh it in olit ik Ak t in om ise t e s
Sist im khit inolit ik akt inom iset es t erut am a pada St rept om yces t erdiri at as beberapa enzim yang m em iliki sifat yang berbeda. Pada S. liv idans t elah diident ifik asi sebany ak 4 m acam k hit inase ( k hit inase A, B, C dan D) dengan berat m olekul m asing- m asing 36, 46, 65 dan 41 kDa. Produksi keem pat enzim ini diinduksi oleh adany a k hit in dan diham bat oleh adany a gluk osa dan gliserol ( Miyashit a et al., 1991) . Khit inase A, B, C dan D m asing m asing m em iliki pH opt im al pada pH 3 ; 5 ; 6,5 dan 6,5.
Jeaniaux ( 1966) m eny at ak an bahw a k hit inase pada St rept om y ces ak t iv pada suasana asam , yait u pada pH ant ara 5 – 6, sedangkan suhu opt im al unt uk akt ivit as khit inase berada pada suhu 40 0C ( Kam el et al.,1993) .
Dalam sist im khit inolit ik St rept om y ces dihasilkan prot ein lain yang bukan enzim yait u prot ein pengikat khit in ( chit in- binding prot ein) yang m am pu berikat an k uat dengan st ruk t ur α- khit in, khit osan dan dinding sel j am ur ( Borm ann et al., 1999) . Prot ein ini t elah diisolasi dari St rept om y ces yait u CHB1 dan CHB2 ( Schnellm ann et al., 1994) dan AFP1 ( Borm ann et al., 1999) . CHB dihasilkan oleh St rept om yces. olivaceoviridis, berat m olek ulny a 12 k Da dan t idak bersifat ant ij am ur. Sedangkan AFP1 yang dihasilkan S. t endae bersifat ant ij am ur, dengan berat m olek ul 9,8 k Da.
C. Sist im Kh it in olit ik Pa da Ja m u r
Khit inase j am ur t erdapat ekst raseluler dalam ruang periplam ik dan m em bran plasm a. Enzim ini dit em uk an m endegradasi k hit in y ang baru t erbent uk secepat pem bent ukan khit in it u sendiri oleh enzim khit in sint et ase. Akt ivit as khit inase dist im ulasi dengan m em berikan prot ease t ripsin pada m ik rosom dan inak t iv asi t erj adi set elah perlakuan dengan phospolipid. Hal ini m enunj ukkan bahw a enzim berint erak si dengan phospolipid k om ersialunt uk ak t iv it asny a.
Ak t iv it as k hit inase j am ur dit em uk an pada frak si subselular y ang berbeda dibandingk an dengan pem bent uk an m ik rosom . Pada Neurrospora crassa ak t iv it as k hit inase t erj adi pada frak si m em bran t api k ebany ak an ak t iv it as didist ribusik an k e dinding sel at au frak si sit osool lainny a sebany ak 30 % dan 50 % dan sebalikny a. Khit inase j uga t erdapat pada dinding sel Aspergillus nidulans, dalam sit osol dan frak si m em bran dari C. cucurbit arum dan P. art iculosus, dalam frak si m em bran, sit osol dan dinding sel dari M. rouxii dan periplasm ik dan v ak uola sert a dinding sel dari S. cereviceae.
Khit inase j am ur bersifat akt iv pada pH asam , m em iliki t em perat ur opt im al y ang t inggi, t ingk at k est abilan y ang t inggi, dan m em puny ai ak t iv it as endok hit inase dan eksok hit inase. Ak t iv it as enzim ini diham bat oleh t em baga dan garam m erkuri, t idak m em iliki kofakt or ionik dan secara kom pet it if diham bat oleh allosam in. Pengham bat an kom pet it if dari seluruh khit inase disebabkan oleh chit obionolact one oxim e.
k hit inase m ak sim al t erj adi pada saat k ony ugasi dengan ak t iv it as k hit in sint et ase m aksim al selam a pert um buhan.
Ak t iv it as k hit inolit ik dari beberapa ent om opat ogen pent ing unt uk pert um buhan dan secara pot ensial dibut uhk an unt uk penet rasi. Pada t iga j enis ent om opat ogen Beauv eria bassiana, Met arhizium anisopliae dan Vert illissin lecanii y ang t um buh pada k ut ik ula belalang sebagai sum ber k arbon m em berikan ak t iv it as endokhit inase, ß- 1,4 N- Aset ilglukosam inidase dan enzim hidrolit ik lain.
Khit inase dan enzim lit ik lainnya dilibat kan dalam kolonisasi ant ara t um buhan inang dengan j am ur m ik orhiza v esik ular arbuskular ( VAM) , ordo Endogonales k las Zygom ycet es. Walaupun j am ur VAM bersifat sim bion obligat , j am ur ini m enghasilkan perok sidase dan k hit inase pada ak ar m ik orhizal dari Allium porrum .
Mik oparasit daei beberapa st rain Trichoderm a spp. berkorelasi dengan kehadiran enzim lit ik sepert i khit inase dan ß- 1,3 glukanase. Kedua enzim ini diinduksi oleh st rain yang berbeda dari Trichoderm a harzianum pada m edia pert um buhan yang diberikan dinding sel Rhizoct onia solani dan Sclerot ium rolfsii sebagai sum ber k arbon t unggal. Pengham bat an perkecam bahan spora dan pem anj angan kecam bah dari 9 j enis j am ur yang m em iliki khit in disebabkan oleh ak t iv it as endok hit inase dan eksok hit inase dari T. harzianum st rain P1. Peningkat an secara sinergis dari ak t iv it as ant ij am ur dihasilkan j ik a dua enzim dik om binasikan. Endok hit inase lainny a dipurifik asi dari Gliocladium v irens yang m em iliki akt ivit as ant ij am ur t erhadap B. cinerea w alaupun dengan kem am puan yang rendah.
Ak t iv it as lit ik dari Trichoderm a- Rhizoct onia dan Trichoderm a- Sclerot ium berkorelasi dengan t ingk at pengendalianny a secara invivo. Siv an dan Chet ( 1989)
m endem onst rasikan bahw a ß- 1,3- glukanase dan khit inase dihasilkan oleh T. Harzianum . Khit inase ekst raseluler dari Trichoderm a sp. m em bebask an oligom er
dari N- aset ilglukosam in dari F. oxysporum f. sp. radicis- lycopersici k e dalam m edium , sej ak 48 j am set elah inokulasi.
Ekspresi gen k hit inase pada j am ur dan m ik roorganism e lainny a dik ont rol oleh sist im represor- induser. Khit in dan produk hasil degradasi ( oligom er) berperan sebagai induser. Gluk osa m erupak an represor pada A. album , B.bassiana dan M. anisoplae. Ak t iv it as k hit inase y ang t inggi hany a dit em uk an pada k ult ur y ang diberi k hit in sebagai sum ber k arbon, t et api buk an selulosa, k hit osan at au k hit obiose. Sebalikny a dit em uk an bahw a GlcNAc t idak m eningk at k an produk si enzim m pada T.harzianum m , GlcNAc dan glukosa m engham bat sint esis khit inase dan ß- 1,4 N-aset ilglukosam inidase.
I V. PEN UTUP
Khit in sebagai polim er yang t erdapat m elim pah di t anah, m em iliki st rukt ur dan karakt erist ik yang unik. Banyak hew an dan m ikroorganism e ( sepert i j am ur, dan alga) m enj adi penyum bang ket ersedian khit in di dalam t anah. Keberadaan senyaw a ini di t anah m erupakan sesuat u hal yang m enarik. Dalam indust ri khit in m em iliki banyak kegunaan yang dapat dim anfaat kan oleh m anusia.
D AFTAR PUSTAKA
Alexander, M. 1977. I nt roduct ion t o Soil Microbiology. Second edit ion. John Willey and Sons, New York.
Bart nicki- Garcia, S. 1989. The Biocheical Cy t ology of Chit in and Chit osan Sy nt hesis in Fungi, Dalam G. Sk j ak , B. T. Ant honsen and P.A. Sanford ( Eds) . Procedings of t he 4t h I nt ernat ional Conferenceon Chit in and Chit osan. Elsevier. Applied Science, Barking- UK.
Berkeley, R. C. W. 1979. Chit in, Chit osan and Their Degradat ive Enzy m es. Dalam R. C. W. Berkeley, G. W. Gooday and D.C. Ellw ood ( Eds.) Microbial Polysaccharides and Polysaccharases pp. 205 – 236. Academ ic Press, London.
Bielka, H., H. B. F. Dixon, P. Karlson, C. Liebeeg, N. Sharon, F. J. Van Lent en, S. F. Velix, J. F. G. Vliegenhart and E. C. Webb. 1984. Enzy m e Nom enclat ure. Academ ic Press, I nc. New york.
Borm ann, C., D. Baier, I . Horr, C. Raps, J. Berger, G. Jung and H. Schw arz. 1999. Charact erizat ion of a Nov el, Ant ifungal, Chit in- binding Prot ein from
St rept om y ces t endae TU 901 t hat I nt erferes w it h Grow t h Polarit y . J. Bact eriol. 181 : 7421 – 7429.
Cabib, E. 1987. The Sy nt hesis and Degradat ion of Chit in. Dalam A. Meist er ( Ed) Adv ances in Enzy m ology. Vol. 59, pp. 59 – 101. An I nt erscience Publicat ion John Willey and Sons I nc., New York.
Carlile, M. J. and S.C. Wat kinson. 1994. The Fungi. Academ is Press, Harcourt Brace and Com pany Publishers, London.
Chernin, L., Z. I sm ailo, S. Haran and I . Chet . 1995. Chit inoly t ic Ent erobact er agglom erans Ant agonist ic t o Fungal Plant Pat hogens. Appl. Env iron. Microbiol. 61 : 1720 – 1726.
Griffin, D. H. 1981. Fungal Phy siology . John Wiley and Sons, New York.
Haran, S., H. Schickler, A. Oppenheim and I . Chet . 1995. New Com ponent s of Chit inolit ic Syst em of Trichoderm a harzianum . Mycol. Res. 94 : 441- 446.
Harm an, G. E., C.K. Hayes, M. Lorit o, R. M. Broadw ay, A. Di Piet ro, C. Pet erbauer and A. Tronsm o. 1993. Chit inoly t ic Enzy m es of Trichoderm a harzianum : Purificat ion of Chit obiosidase and Endochit inase. Phyt opat hology 83: 313- 318.
I nbar J. and Chet I . 1991. Evidence That Chit inase Produced By Aer om onas caviae I s I nvolved I n The Biological Cont rol Of Soil- Borne Plant Pat hogens By This Bact erium . Soil. Biol. Biochem . 23 : 973- 978.
Jeaniaux , C. 1966. Chit inases, Dalam E, F. Neufeld and V. Ginburg ( Eds.) Com plex Carbohydrat es, Met hods in Enzy m ology . Vol. VI I , pp. 644 – 650. Academ ic Press, New York.
Lloyd, A. B., R. L. Nov eroske and J. L. Lockw ood. 1965. Lysis of Fungal My celium by St rept om y ces spp. and Their Chit inase Syst em s. Phyt opat hology 55 : 871– 875.
Miy ashit a, K., T. Fuj ii and Y. Saw ada. 1991. Molecular Cloning and Charact erizat ion of Chit inase Genes from St rept om yces lividans 66. J. Gen. Microbiol. 137 : 2065 - 2072.
Muzzarelli, R. A. A. 1985. Chit in, Dalam G. O. Aspinal ( Ed.) The Polysaccharides. Vol. 3, pp. 417 – 450. Academ ic Press I nc., New York.
Raj arat hanam , S., M. N. J. Shashirek ha and Z. Bano. 1998. Biodegradat iv e and Biosint het ic Capacit ies of Mushroom s : Present and Fut ure St rat egies. Crit . Rev. in Biot echnol. 18 : 91 – 236.
Richards, A. G. 1951. The I nt egum ent of Art hropods. The Chem ical Com ponent s and Their Propert ies : The Anat om y and Developm ent and Perm eabilit y. Universit y of Minnesot a Press, Minneapolis.
Sahai , A. S. and M. S. Manocha. 1993. Chit inases of Fungi and Plant s : Their I nv olvem ent in Morphogenesis and Host - Parasit e I nt eract ion. FEMS Microbiol. Rev . 11 : 317 – 338.
Sait o, A., T. Fuj ii, T, Yoney am a and K. Miy ashit a. 1998. glk A is I nv olved in Glucose Repression of Chit inase Product ion in St rept om y ces liv idanns. J. Bact eriol. 180: 2911- 2914.
Schrem pf, H. 1995. The Chit inoly t ic Sy st em of St rept om yces olivaceoviridis. Dalam S. B Pet ersen., B. Sv enson and S. Pedersen ( Eds.) . Progress I n Biot echnology. Carbohydrat e Bioenginering. Vol. 10, pp. 71 – 75. Elsev ier Science. B. V. Am st erdam .
Schnellm ann, J., A. Zelt ins, H. Blaak and H. Schrem pf. 1994. The Nov el Lect in- like CHB 1 is Encoded by a Chit in- inducible St rept om y ces oliv aceov iridis gene and Binds Specifically t o Cryst alline a- Chit in of Fungi and Ot her Organism . Mol. Microbiol. 13 : 807 – 819.
Scopes, R. K. 1994. Pr ot ein Pur ificat ion, Principles and Pract ice. Third edit ion. Springer- Verlag, New York.
Singh, P.P., Y. C. Shin, C. S. Park and Y. R. Chung. 1999. Biological Cont rol of Fusarium Wilt of Cucum ber by Chit inolyt ic Bact eria. Phyt opat hology 89: 92-99.
Siv an, A and I . Chet . 1989. Degradat ion of Fungal Cell Walls by Lyt ic Enzy m es of Trichoderm a harzianum . J. Gen. Microbiol. 135 : 675 – 682.
Wak sm an, S. A. 1967. The Act inom y cet es, A Sum m ary of Current Know ledge. The Ronald Press Com pany, New York.