SELEKSI DINI POHON INDUK TANAMAN KARET (Hevea brasiliensis Muell Arg.) DARI HASIL PERSILANGAN RRIM 600 X PN 1546 BERDASARKAN
PRODUKSI LATEKS DAN KAYU
KOKO MARDIANTO 070307020
PROGRAM STUDI PEMULIAAN TANAMAN DEPARTEMEN BUDIDAYA PERTANIAN
FAKULTAS PERTANIAN UNIVERSITAS SUMATERA UTARA
SELEKSI DINI POHON INDUK TANAMAN KARET (Hevea brasiliensis Muell Arg.) DARI HASIL PERSILANGAN RRIM 600 X PN 1546 BERDASARKAN
PRODUKSI LATEKS DAN KAYU
SKRIPSI
Oleh :
KOKO MARDIANTO
070307020
PROGRAM STUDI PEMULIAAN TANAMAN DEPARTEMEN BUDIDAYA PERTANIAN
FAKULTAS PERTANIAN UNIVERSITAS SUMATERA UTARA
SELEKSI DINI POHON INDUK TANAMAN KARET (Hevea brasiliensis Muell Arg.) DARI HASIL PERSILANGAN RRIM 600 X PN 1546 BERDASARKAN
PRODUKSI LATEKS DAN KAYU
SKRIPSI
Oleh :
KOKO MARDIANTO
070307020 / PEMULIAAN TANAMAN
Skripsi sebagai salah satu syarat untuk memperoleh gelar sarjana di Fakultas Pertanian
Universitas Sumatera Utara
PROGRAM STUDI PEMULIAAN TANAMAN DEPARTEMEN BUDIDAYA PERTANIAN
FAKULTAS PERTANIAN UNIVERSITAS SUMATERA UTARA
Judul Penelitian : Seleksi dini pohon induk tanaman karet (Hevea brasiliensis Muell Arg.) dari hasil persilangan RRIM 600 X PN 1546 berdasarkan produksi lateks dan kayu
Nama : Koko Mardianto
NIM : 070307020
Departemen : Budidaya Pertanian Program Studi : Pemuliaan Tanaman
Disetujui oleh, Komisi Pembimbing :
Ir. Yusuf Husni Prof. Dr. Ir. Rosmayati, MS Dra. Sekar Woelan, MP Ketua Anggota Anggota
Mengetahui,
Ir. T. Sabrina, M.Agr.Sc, PhD Ketua Departemen Agroekoteknologi
ABSTRAK
KOKO MARDIANTO: Seleksi Dini Pohon Induk Tanaman Karet (Hevea
brasiliensis Muell Arg.) dari Hasil Persilangan RRIM 600 X PN 1546
Berdasarkan Produksi Lateks dan Kayu, dibimbing oleh Ir. Yusuf Husni, Prof. DR. Ir. Rosmayati, MS dan Dra. Sekar Woelan, MP.
Perbaikan potensi genetik merupakan faktor yang penting dalam upaya meningkatkan produktivitas karet. Kemajuan pemuliaan karet sangat tergantung kepada potensi dan ketersediaan sumber keragaman genetik, yang diperoleh melalui kegiatan persilangan antara tetua yang memiliki hubungan kekerabatan jauh. Penelitian ini bertujuan untuk mengetahui keragaman genotipe, fenotipe, heritabilitas, dan kemajuan seleksi yang ditimbulkan dari hasil persilangan intraspesifik antara RRIM 600 x PN 1546 dan mendapatkan genotipe-genotipe yang berpotensi sebagai penghasil lateks dan kayu. Penelitian ini dilaksanakan di Kebun Percobaan tanaman F1 hasil okulasi (turunan) dari persilangan intraspesifik RRIM 600 x PN 1546 di Balai Penelitian Sungei Putih - Pusat Penelitian Karet, Kecamatan Galang, Kabupaten Deli Serdang-Sumatera Utara, dengan ketinggian tempat ± 54 m dpl pada bulan Januari 2011 sampai dengan Maret 2011 menggunakan rancangan acak kelompok non faktorial yang terdiri dari 25 genotipe dan 2 tetua, yaitu RRIM 600 dan PN 1546. Parameter yang diamati adalah tinggi tanaman, jumlah cabang primer, tinggi cabang pertama, lilit batang, tebal kulit, jumlah pembuluh lateks, diameter pembuluh lateks, produksi karet kering dan produksi kayu.
Hasil penelitian menunjukkan bahwa genotipe berbeda nyata terhadap parameter tinggi tanaman, lilit batang, tebal kulit, jumlah pembuluh lateks, produksi karet kering dan produksi kayu. Koefisien keragaman genetik (KKG) dan heritabilitas (h2) dari genotipe yang diteliti cukup tinggi untuk rata-rata produksi karet kering, tinggi tanaman, lilit batang, tebal kulit dan jumlah pembuluh lateks, dengan nilai KKG 7,84 - 40,49% dan nilai h2 antara 0,672 – 0,819 dan seleksi atas dasar karakter ini memberikan kemajuan genetik sebesar 0,542 – 6,355. Terdapat tiga genotipe yang memiliki potensi sebagai penghasil lateks yaitu genotipe no. 19, 37 dan 40, dan tiga genotipe yang memiliki potensi sebagai penghasil kayu yaitu genotipe no. 14, 16 dan 28.
ABSTRACT
KOKO MARDIANTO: Early Selection Mother Plant of Rubber (Hevea brasiliensis Muell Arg.) from the Result of RRIM 600 X PN 1546 Crossing Based on Latex and Timber Yield, supervised by Ir. Yusuf Husni, Prof. DR. Ir. Rosmayati, MS and Dra. Sekar Woelan, MP.
Improvement of genetic potential is an important factor in improving yield. Progress of rubber breeding is dependent on the potency and availability of sources of genetic diversity, which is obtained through crossing between parent who have distant relationship. This objective of reasearch to determine the diversity of genotype, phenotype, heritability, and selection progress arising from the results of intraspecific crosses between RRIM 600 x PN 1546 and obtain genotypes that have the potential as a producer of latex and timber. The research was conducted at the Experimental Garden results of grafting F1 plants (derived) from intraspecific crosses RRIM 600 x PN 1546 in Sungei Putih Research Center for Indonesian Rubber Research, Galang, Deli Serdang, North Sumatera, with ± 54 m altitude above sea level in January 2011 to March 2011. The design using non-factorial randomized block design consisting of 25 genotypes and 2 parents, namely RRIM 600 and PN 1546. The parameters observed were plant height, number of primary branches, first branch height, girth, bark thickness, number of latex vessels, diameter of latex vessel, dry rubber content and timber yield.
The results showed that genotype significantly increased the parameters plant height, girth, bark thickness, number of latex vessels, dry rubber content and timber yield. The coefficient of genetic variability (CGV) and heritability (h2) of the genotype is high for the average dry rubber content, plant height, girth, bark thickness and number of latex vessels, with CVG value from 7,84 to 40,49% and h2 values between 0,672 to 0,819 and selection on the basis of these characters provide the genetic progress of 0,542 to 6,355. There are three genotypes which have potential as a producer of latex were genotype No. 19, 37 and 40, and three genotypes that have potential as a producer of wood that is genotype No. 14, 16 and 28.
RIWAYAT HIDUP
Penulis dilahirkan di Medan pada tanggal 20 Maret 1989 dari ayah Ir. Usul
dan ibu Supiatmi. Penulis merupakan putra kedua dari empat bersaudara.
Tahun 2007 penulis lulus dari SMA Negeri 4, Medan dan pada tahun yang
sama masuk ke Fakultas Pertanian USU melalui jalur ujian tertulis Seleksi
Penerimaan Mahasiswa Baru. Penulis memilih program studi Pemuliaan
Tanaman, Departemen Budidaya Pertanian.
Penulis melaksakan Praktek Kerja Lapangan (PKL) di Balai Penelitian
Sungei Putih - Pusat Penelitian Karet, Kecamatan Galang, Kabupaten Deli
KATA PENGANTAR
Puji dan syukur penulis ucapkan kehadirat Allah SWT, atas segala rahmat
dan karunia-Nya sehingga penulis dapat menyelesaikan skripsi yang berjudul
”Seleksi Dini Pohon Induk Tanaman Karet (Hevea brasiliensis Muell Arg.) dari
Hasil Persilangan RRIM 600 X PN 1546 Berdasarkan Produksi Lateks dan
Kayu”.
Pada kesempatan ini penulis menghaturkan terima kasih sebesar-besarnya
kepada ayahanda Ir. Usul dan Ibunda Supiatmi yang telah memberikan kasih
sayang dan mendidik penulis selama ini. Penulis menyampaikan ucapan terima
kasih dan doa untuk Bapak Alm. Prof. DR. Ir. T.M. Hanafiah Oeliem, DAA
selaku ex. ketua komisi pembimbing, serta ucapan terima kasih kepada bapak
Ir. Yusuf Husni selaku ketua komisi pembimbing, ibu Prof. DR. Ir. Rosmayati, MS dan
ibu Dra. Sekar Woelan, MP selaku anggota komisi pembimbing yang telah
membimbing dan memberikan masukan berharga kepada penulis dalam
menyelesaikan skripsi ini.
Terima kasih kepada abangku Eka Bobby Febrianto serta adik-adikku
Indra Wahyu Wibawa dan Annisa Mardianti (Ubiet) yang telah menjadi
penyemangat selama masa perkuliahan. Terima kasih kepada sahabat terbaikku
Suci Apriani Harahap, Satriya, Bayu, Gusman, Ucup, Babenk, Baron, Adnan,
Budi, Ade, Ferdy, Nida, Fina, Nadya, Roza, dan seluruh teman-teman BDP 2007
serta pihak-pihak yang telah banyak membantu penulis dalam melaksanakan
Afrizal A.R. beserta keluarga dan Bang Sayurandi yang telah banyak memberikan
bantuan, para teknisi Pak Adi Mulyono, Bang Indra, Bang Adi, Kak Fina, Bang
Sahrul, Pak Surip dan Pak Zaiman yang membantu penulis di lapangan maupun di
laboratorium selama penelitian berlangsung, serta karyawan/i dan seluruh
keluarga besar Balai Penelitian Sungei Putih yang telah banyak membantu dan
memberikan dukungan sehingga penelitian ini dapat terlaksana.
Penulis menyadari bahwa skripsi ini masih jauh dari sempurna. Oleh
karena itu, penulis mengharapkan kritik dan saran yang bersifat membangun demi
kesempurnan skripsi ini. Semoga skripsi ini bermanfaat bagi seluruh pihak yang
memerlukan.
Medan, Juni 2011
DAFTAR ISI
Hal.
ABSTRAK ... i
ABSTRACT ... ii
RIWAYAT HIDUP ... iii
KATA PENGANTAR ... iv
DAFTAR ISI ... vi
DAFTAR TABEL ... viii
DAFTAR LAMPIRAN ... ix
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 4
Hipotesis Penelitian ... 4
Kegunaan Penelitian ... 4
TINJAUAN PUSTAKA... 5
Botani Tanaman ... 5
Syarat Tumbuh ... 8
Iklim ... 8
Tanah ... 9
Peningkatan Produktivitas Melalui Pemuliaan Tanaman Karet ... 10
Keragaman Genotipe dan Fenotipe... 15
Heritabilitas ... 16
Kemajuan Genetik ... 18
METODE PENELITIAN ... 20
Tempat dan Waktu ... 20
Bahan dan Alat ... 20
Metode Penelitian ... 21
PELAKSANAAN PENELITIAN ... 25
Persiapan Areal ... 25
Sensus Tanaman ... 25
Membuat Batas Tinggi Penyadapan ... 25
Menyiapkan Penampungan Lateks ... 25
Penyadapan ... 25
Pengamatan Parameter ... 26
Tinggi Tanaman (m) ... 26
Jumlah Cabang Primer (cabang) ... 26
Tinggi Cabang Pertama (m) ... 26
Lilit Batang (cm) ... 26
Tebal Kulit (mm)... 26
Jumlah dan Diameter Pembuluh Lateks ... 27
Produksi Kayu (cm3/pohon)... 29
HASIL DAN PEMBAHASAN ... 30
Hasil ... 30
Tinggi Tanaman (cm) ... 30
Jumlah Cabang Primer (cabang) ... 31
Tinggi Cabang Pertama (m) ... 31
Lilit Batang (cm) ... 32
Tebal Kulit (mm)... 33
Jumlah Pembuluh Lateks ... 34
Diameter Pembuluh Lateks (mµ) ... 35
Produksi Karet Kering (g/p/s) ... 36
Produksi Kayu (cm3/pohon)... 37
Pendugaan Parameter Genetik ... 38
Hubungan Karakter Agronomi Terhadap Produksi Karet Kering ... 39
Hubungan Karakter Agronomi Terhadap Produksi Kayu ... 40
Seleksi Genotipe Berdasarkan Potensi Produksi Karet Kering dan Kayu .... 40
Pembahasan ... 41
KESIMPULAN ... 46
Kesimpulan ... 46
DAFTAR PUSTAKA ... 47
DAFTAR TABEL
No. Hal.
1. Analisis ragam dan pendugaan komponen ragam ... 22
2. Rataan tinggi tanaman dari 25 genotipe dan tetua ... 30
3. Rataan jumlah cabang primer dari 25 genotipe dan tetua ... 31
4. Rataan tinggi cabang pertama dari 25 genotipe dan tetua ... 32
5. Rataan lilit batang dari 25 genotipe dan tetua ... 33
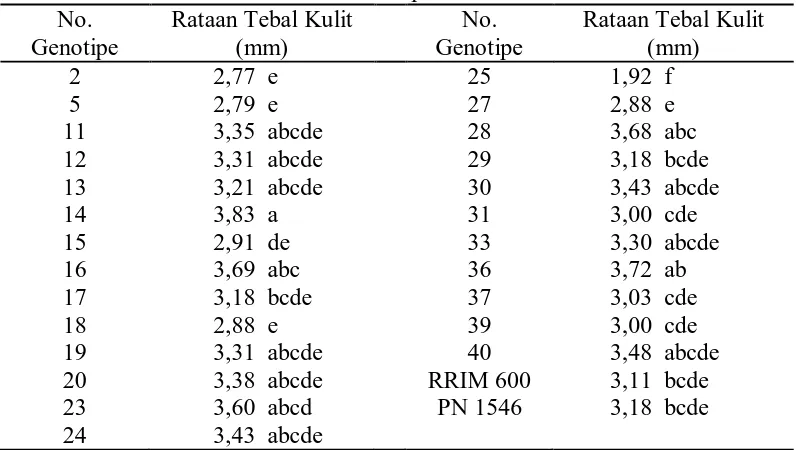
6. Rataan tebal kulit dari 25 genotipe dan tetua ... 34
7. Rataan jumlah pembuluh lateks dari 25 genotipe dan tetua ... 35
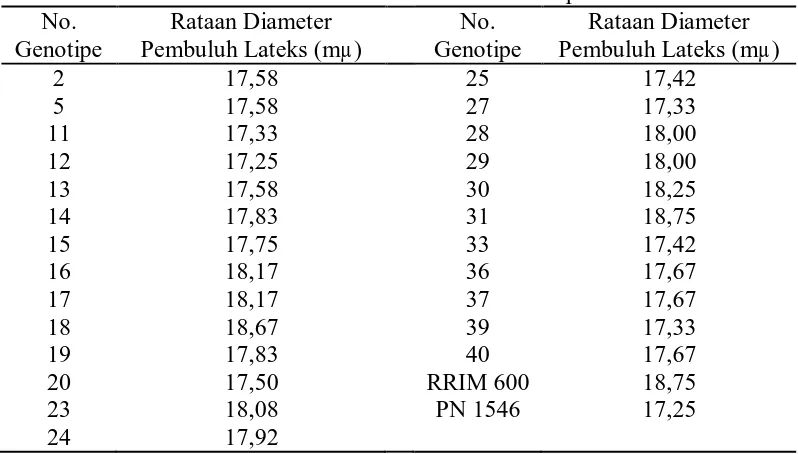
8. Rataan diameter pembuluh lateks dari 25 genotipe dan tetua ... 36
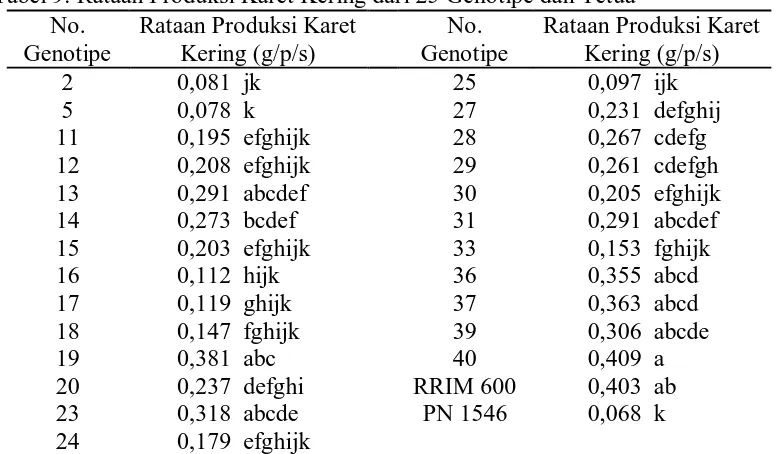
9. Rataan produksi karet kering dari 25 genotipe dan tetua ... 37
10. Rataan produksi kayu dari 25 genotipe dan tetua ... 38
11. Nilai pendugaan komponen ragam genotipe (σ2g), ragam fenotipe (σ2p), koefisien keragaman genetik (KKG), koefisien keragaman fenotipe (KKF), heritabilitas (h2) dan harapan kemajuan genetik (HKG) ... 39
12. Matriks korelasi karakter agronomi terhadap produksi karet kering ... 39
13. Matriks korelasi karakter agronomi terhadap produksi kayu ... 40
DAFTAR LAMPIRAN
No. Hal.
1. Deskripsi tetua karet... 51
2. Bagan penelitian ... 53
3. Jadwal kegiatan ... 54
4. Data pengamatan tinggi tanaman (m) ... 55
5. Daftar sidik ragam tinggi tanaman ... 55
6. Data pengamatan jumlah cabang primer (cabang) ... 56
7. Daftar sidik ragam jumlah cabang primer ... 56
8. Data pengamatan tinggi cabang pertama (m) ... 57
9. Daftar sidik ragam tinggi cabang pertama ... 57
10. Data pengamatan lilit batang (cm) ... 58
11. Daftar sidik ragam lilit batang ... 58
12. Data pengamatan tebal kulit (mm) ... 59
13. Daftar sidik ragam tebal kulit ... 59
14. Data pengamatan jumlah pembuluh lateks ... 60
15. Daftar sidik ragam jumlah pembuluh lateks ... 60
16. Data pengamatan diameter pembuluh lateks (mµ) ... 61
17. Daftar sidik ragam diameter pembuluh lateks ... 61
18. Data pengamatan produksi karet kering (g/p/s) ... 62
19. Daftar sidik ragam produksi karet kering ... 62
20. Data pengamatan produksi kayu (cm3/pohon)... 63
22. Nilai pendugaan komponen ragam genotipe (σ2g), ragam fenotipe (σ2p),
koefisien keragaman genetik (KKG), koefisien keragaman fenotipe (KKF),
heritabilitas (h2) dan harapan kemajuan genetik (HKG) ... 64
23. Matriks korelasi karakter agronomi terhadap produksi karet kering ... 64
ABSTRAK
KOKO MARDIANTO: Seleksi Dini Pohon Induk Tanaman Karet (Hevea
brasiliensis Muell Arg.) dari Hasil Persilangan RRIM 600 X PN 1546
Berdasarkan Produksi Lateks dan Kayu, dibimbing oleh Ir. Yusuf Husni, Prof. DR. Ir. Rosmayati, MS dan Dra. Sekar Woelan, MP.
Perbaikan potensi genetik merupakan faktor yang penting dalam upaya meningkatkan produktivitas karet. Kemajuan pemuliaan karet sangat tergantung kepada potensi dan ketersediaan sumber keragaman genetik, yang diperoleh melalui kegiatan persilangan antara tetua yang memiliki hubungan kekerabatan jauh. Penelitian ini bertujuan untuk mengetahui keragaman genotipe, fenotipe, heritabilitas, dan kemajuan seleksi yang ditimbulkan dari hasil persilangan intraspesifik antara RRIM 600 x PN 1546 dan mendapatkan genotipe-genotipe yang berpotensi sebagai penghasil lateks dan kayu. Penelitian ini dilaksanakan di Kebun Percobaan tanaman F1 hasil okulasi (turunan) dari persilangan intraspesifik RRIM 600 x PN 1546 di Balai Penelitian Sungei Putih - Pusat Penelitian Karet, Kecamatan Galang, Kabupaten Deli Serdang-Sumatera Utara, dengan ketinggian tempat ± 54 m dpl pada bulan Januari 2011 sampai dengan Maret 2011 menggunakan rancangan acak kelompok non faktorial yang terdiri dari 25 genotipe dan 2 tetua, yaitu RRIM 600 dan PN 1546. Parameter yang diamati adalah tinggi tanaman, jumlah cabang primer, tinggi cabang pertama, lilit batang, tebal kulit, jumlah pembuluh lateks, diameter pembuluh lateks, produksi karet kering dan produksi kayu.
Hasil penelitian menunjukkan bahwa genotipe berbeda nyata terhadap parameter tinggi tanaman, lilit batang, tebal kulit, jumlah pembuluh lateks, produksi karet kering dan produksi kayu. Koefisien keragaman genetik (KKG) dan heritabilitas (h2) dari genotipe yang diteliti cukup tinggi untuk rata-rata produksi karet kering, tinggi tanaman, lilit batang, tebal kulit dan jumlah pembuluh lateks, dengan nilai KKG 7,84 - 40,49% dan nilai h2 antara 0,672 – 0,819 dan seleksi atas dasar karakter ini memberikan kemajuan genetik sebesar 0,542 – 6,355. Terdapat tiga genotipe yang memiliki potensi sebagai penghasil lateks yaitu genotipe no. 19, 37 dan 40, dan tiga genotipe yang memiliki potensi sebagai penghasil kayu yaitu genotipe no. 14, 16 dan 28.
ABSTRACT
KOKO MARDIANTO: Early Selection Mother Plant of Rubber (Hevea brasiliensis Muell Arg.) from the Result of RRIM 600 X PN 1546 Crossing Based on Latex and Timber Yield, supervised by Ir. Yusuf Husni, Prof. DR. Ir. Rosmayati, MS and Dra. Sekar Woelan, MP.
Improvement of genetic potential is an important factor in improving yield. Progress of rubber breeding is dependent on the potency and availability of sources of genetic diversity, which is obtained through crossing between parent who have distant relationship. This objective of reasearch to determine the diversity of genotype, phenotype, heritability, and selection progress arising from the results of intraspecific crosses between RRIM 600 x PN 1546 and obtain genotypes that have the potential as a producer of latex and timber. The research was conducted at the Experimental Garden results of grafting F1 plants (derived) from intraspecific crosses RRIM 600 x PN 1546 in Sungei Putih Research Center for Indonesian Rubber Research, Galang, Deli Serdang, North Sumatera, with ± 54 m altitude above sea level in January 2011 to March 2011. The design using non-factorial randomized block design consisting of 25 genotypes and 2 parents, namely RRIM 600 and PN 1546. The parameters observed were plant height, number of primary branches, first branch height, girth, bark thickness, number of latex vessels, diameter of latex vessel, dry rubber content and timber yield.
The results showed that genotype significantly increased the parameters plant height, girth, bark thickness, number of latex vessels, dry rubber content and timber yield. The coefficient of genetic variability (CGV) and heritability (h2) of the genotype is high for the average dry rubber content, plant height, girth, bark thickness and number of latex vessels, with CVG value from 7,84 to 40,49% and h2 values between 0,672 to 0,819 and selection on the basis of these characters provide the genetic progress of 0,542 to 6,355. There are three genotypes which have potential as a producer of latex were genotype No. 19, 37 and 40, and three genotypes that have potential as a producer of wood that is genotype No. 14, 16 and 28.
PENDAHULUAN
Latar Belakang
Perekonomian di Indonesia salah satunya dihasilkan dari pengembangan
kebun karet. Fungsi dari perkebunan karet tidak hanya sebagai sumber devisa,
sumber bahan baku industri, sumber pendapatan dan kesejahteraan masyarakat
tetapi sekaligus berperan dalam pelestarian fungsi lingkungan hidup.
Selama tiga dekade ini pengembangan karet di Indonesia mengalami
pertumbuhan yang sangat pesat. Di awal tahun 1968, luas areal karet baru 2,2 juta
ha dan pada tahun 2005 meningkat menjadi 3,2 juta ha atau meningkat menjadi
sekitar 50%. Hampir 85% pengusahaan karet diusahakan oleh rakyat dan sisanya
oleh perkebunan besar. Produksi yang dihasilkan dari luasan tersebut mencapai
sebesar 2,2 juta ton dengan produktivitas rata-rata sebesar 840 kg/ha/tahun
(Dirjenbun, 2006).
Tanaman karet dikenal mempunyai daya adaptasi yang cukup luas.
Tanaman ini mampu tumbuh pada berbagai jenis tanah dan kondisi iklim. Tetapi
walaupun demikian keunggulan yang dimiliki akan terealisasi secara maksimal
apabila ditanam pada kondisi agroekosistem yang sesuai.
Kemajuan produktivitas yang telah dicapai selama 10 tahun penyadapan
dari generasi-1 ke generasi-4 adalah sebagai berikut: rata-rata produktivitas dari
seedling terseleksi pada generasi-1 adalah 20,9 g/p/s, dan klon primer dihasilkan
pada generasi-2 sekitar 35,6 g/p/s (70% lebih baik dari generasi-1). Adapun klon
produktivitas sebesar 44,9 g/p/s (26% lebih baik dari generasi-2). Peningkatan
produktivitas pada generasi-3 agak lambat dibanding peningkatan produktivitas
pada generasi-1. Hal ini kemungkinan disebabkan oleh sempitnya keragaman
genetik dari turunan Wickham 1876. Beberapa peneliti mengemukakan bahwa
peningkatan produktivitas masih dapat dicapai, karena karet bersifat heterozygous
(Liu, 1998; Aidi-Daslin, et al., 2000).
Salah satu upaya yang telah dilakukan untuk memperbesar keragaman
genetik tanaman karet di Indonesia yaitu dengan memanfaatkan plasma nutfah
yang telah dikonservasi sejak tahun 1984-1989. Plasma nutfah tanaman karet
merupakan hasil ekspedisi IRRDB pada tahun 1981 di Lembah Amazone, Brazil.
Peluang untuk mendapatkan genotipe unggul baru akan lebih besar apabila
dilakukan penggabungan genetik antara Wickham 1876 x Plasma Nutfah IRRDB 1981.
Evaluasi terhadap potensi produksi awal dapat dilakukan lebih awal
dengan menggunakan penyadapan testateks pada saat tanaman berumur 1,5 tahun
sebagai seleksi awal (dini). Penyadapan dilakukan dengan menggunakan alat
ciptaan Cramer yang berbentuk huruf V. Jarak antara pisau masing-masing 2 cm.
Pisau ini ditekan ke kulit batang tanaman pada ketinggian 50 cm di atas
permukaan tanah dan lateks akan mengalir keluar dari luka (Danimihardja, 1986).
Karakter agronomi suatu tanaman merupakan komponen yang menentukan
besarnya produksi, sehingga perlu dikaji hubungan antar beberapa karakter
dengan produksinya. Khusus untuk tanaman karet, pemuliaan tanaman tidak
hanya bertujuan menghasilkan klon penghasil lateks saja tetapi juga diharapkan
menghasilkan kayu yang tinggi. Sehingga pengkajian terhadap karakter agronomi
pembuluh lateks, diameter pembuluh lateks), fisiologi lateks, dan hasil lateks
perlu dikaji, sebab korelasi diantara karakter tanaman merupakan dasar dalam
program dan perencanaan pemuliaan yang lebih efisien. Hal ini menunjukkan
bahwa antara sifat-sifat tersebut terdapat hubungan, baik secara positif maupun
negatif. Berdasarkan beberapa hasil percobaan dengan melihat komponen hasil
tidak ada varietas yang superior dalam semua sifat. Keunggulan yang dihasilkan
merupakan hasil gabungan antara berbagai komponen hasil yang dapat diperoleh
melalui persilangan dan setiap komponen hasil bersifat poligenik dalam
keturunannya (Lasminingsih, 1993).
Paradigma baru bahwa tanaman karet tidak hanya menghasilkan lateks
tetapi juga diharapkan kayu karetnya, maka seleksi juga diarahkan kepada
klon-klon yang berpotensi sebagai penghasil kayu. Sejak dari tahap awal seleksi sampai
dengan pengujian klon, kedua peubah tersebut terus dievaluasi. Pada seleksi F1
yang merupakan tahap awal di dalam siklus pemuliaan tanaman karet, potensi
genotipe penghasil lateks dan kayu menjadi fokus seleksi (Suhendry, 2002).
Berdasarkan uraian tersebut di atas maka diperlukan suatu pengkajian
terhadap keragaman hasil persilangan intraspesifik antara klon RRIM 600 x PN
1546.
Tujuan Penelitian
Untuk mengetahui keragaman genotipe, fenotipe, heritabilitas, dan
kemajuan seleksi yang ditimbulkan dari hasil persilangan intraspesifik antara
RRIM 600 x PN 1546 dan mendapatkan genotipe-genotipe yang berpotensi
Hipotesis Penelitian
1. Adanya keragaman genotipe, fenotipe, heritabilitas, dan kemajuan genetik dari
hasil persilangan intraspesifik antara RRIM 600 x PN 1546.
2. Adanya genotipe yang terseleksi sebagai penghasil lateks dan kayu.
Kegunaan Penelitian
1. Sebagai bahan penulisan skripsi yang merupakan salah satu syarat untuk
memperoleh gelar sarjana di Fakultas Pertanian, Universitas Sumatera Utara,
Medan.
TINJAUAN PUSTAKA
Botani Tanaman
Menurut (Kartasapoetra, 1988) tanaman karet (Hevea brasiliensis Muell
Arg.) memiliki sistematika sebagai berikut:
Divisio : Spermatophyta
Subdivisio : Angiospermae
Class : Dicotyledoneae
Ordo : Euphorbiales
Family : Euphorbiaceae
Genus : Hevea
Species : Hevea brasiliensis Muell Arg.
Tanaman karet adalah anggota famili Euphorbiaceae. Berbentuk pohon,
tinggi 10-20 m, bercabang dan mengandung banyak getah susu. Daun
berselang-seling, tangkai daun panjang, 3 anak daun yang licin bertangkai, petiola pendek,
hijau dan memiliki panjang 3,5-30,0 cm. Helaian anak daun bertangkai pendek
dan berbentuk elips atau bulat telur, pangkal sempit dan tegang, ujung runcing,
sisi atas daun hijau tua dan sisi bawah agak cerah, panjangnya 5-35 cm dan lebar
2,5-12,5 cm (Sianturi, 1996).
Daun karet berwarna hijau dan terdiri dari tangkai daun utama dan tangkai
anak daun. Tanaman karet adalah tanaman berumah satu. Pada satu tangkai bunga
yang terbentuk bunga majemuk terdapat bunga betina dan bunga jantan
Buah tanaman ini beruang tiga dan jarang beruang empat atau enam,
diameter buah 3-5 cm dan terpisah 3, 4 dan 6 cocci berkatup dua. Pericarp
berbatok dan endocarp berkayu (Sianturi, 1996).
Buah jadi (fruit set) merupakan produk dari keberhasilan pesilangan secara
alami maupun secara buatan. Satu buah karet biasanya mengandung tiga butir biji
tetapi kadang-kadang ada yang empat biji. Biji karet dilindungi oleh epicarp
(lapisan luar) dan endocarp (lapisan dalam). Epicarp berwarna hijau muda
sedangkan endocarp berwarna putih pudar dan apabila buah telah masak fisiologis
epicarp akan berwarna hijau tua dan endocarp akan mengeras dan mengayu. Jika
epicarp kering buah akan pecah dan melepaskan biji (Dijkman, 1951).
Proses pemasakan buah berlangsung selama 5-6 bulan. Musim panen biji
berlangsung pendek, hanya sekitar 1,5 bulan. Sedangkan daya kecambah biji
sangat cepat berkurang, terutama bila penanganannya kurang baik
(Setyamidjadja, 1993).
Biji karet memiliki bentuk dan ukuran yang bervariasi bergantung pada
masing-masing tetua. Biasanya biji berbentuk bulat lonjong (ellips), panjang
14-25 mm dan berat rata-rata 3,5 gram sampai 6 gram. Bentuk permukaan perut
(ventral) biji agak rata dan punggung (dorsal) agak menonjol. Kulit biji biasanya
keras, berkilat, dan berwarna cokelat atau cokelat keabu-abuan dengan banyak
batik (mosaik) pada permukaan punggung tetapi sedikit atau tidak ada pada bagian
perut (Webster dan Baulkwill, 1989).
Tanaman karet merupakan tanaman berumah satu (monoceous) yang
bersifat unisexual yaitu, pada satu tanaman terdapat bunga betina (femineus) dan
Bunga karet termasuk bunga majemuk tidak terbatas yang berbentuk
rangkaian (inflorecentia) yang tangkai utamanya (pedenculus) bercabang terdiri
dari atas beberapa malai (panicula) yang berbentuk piramida atau kerucut
(Dijkman, 1951; Darjanto dan Satifah, 1982).
Bunga betina tumbuh diujung tangkai dan cabangnya. Sedangkan bunga
jantan tumbuh disetiap tangkai bunga yang tersusun atas tiga bunga (trifolia).
Kedua bunga ini memiliki tangkai pendek, berbau harum, berwarna kuning untuk
bunga jantan, dan kuning kehijauan untuk bunga betina. Ukuran bunga betina 8
mm dan umumnya lebih besar dari bunga jantan yang ukurannya sekitar 5-6 mm.
Bunga betina terdiri atas dasar bunga, tenda bunga dan bakal buah. Dasar bunga
berwarna hijau, tenda bunga terdiri atas lima helai daun bunga yang saling
berlekatan pada bagian bawah dan terbelah, sedangkan pada bagian ujung
membelah. Bunga jantan terdiri atas tangkai sari (filamen) dan kepala sari
(anther). Kepala sari melekat pada tangkai sari tersusun dalam dua lingkaran yang
masing-masing terdiri atas lima kepala sari (Dijkman, 1951).
Karakteristik bunga betina pada beberapa tetua karet bervariasi antara 5-16
bunga per tangkai dengan rata-rata 11 bunga per tangkai dan 49-130 bunga per
karangan dengan rata-rata 98 bunga per karangan. Ukuran rata-rata bunga betina
adalah 8-9 mm dengan rata-rata panjang tangkai putik 3,5 mm. Pembuahan dan
pembentukan biji yang terbaik diperoleh apabila bunga memiliki tangkai putik
yang pendek (Syarifah dan Woelan, 2007; Darjanto dan Satifah, 1982).
Karakteristik bunga jantan pada beberapa tetua karet cukup bervariasi,
yaitu 295-500 bunga per tangkai dengan rata-rata 383,4 per tangkai dan
Masing-masing bunga jantan dari setiap tetua tumbuh di setiap tangkai utama dan
cabang-cabangnya, untuk satu tangkai bunga tersusun atas tiga bunga jantan (trifolia)
yang berwarna kuning (Syarifah dan Woelan, 2007).
Syarat Tumbuh Iklim
Tanaman karet tumbuh di dataran rendah, yang paling ideal adalah pada
ketinggian 0-200 m dari permukaan laut. Pada ketinggian lebih dari 200 m dpl
rataan pertumbuhan batang lebih lambat, penyebaran perkebunan karet di
Indonesia terbanyak adalah hingga tinggi 400 m dpl. Pada ketinggian 400-600 m
masih mungkin mengusahakan tanaman karet, lebih dari 600 m tidak dianjurkan
untuk ditanami karet (Dijkman, 1951; Sianturi, 1996 ).
Tanaman karet adalah tanaman tropis, kebanyakan perkebunan karet
diusahakan pada kawasan dengan letak lintang antara 150 LU hingga 100 LS.
Vegetasi yang sesuai untuk kondisi lintang tersebut adalah hutan hujan tropis
yang disertai dengan suhu panas dan kelembaban tinggi. Sekalipun demikian,
pada umumnya produksi maksimum lateks dapat tercapai apabila ditanam pada
lokasi yang semakin mendekati garis khatulistiwa (5-60 LU - LS)
(Syamsulbahri, 1996).
Lama penyinaran dan intensitas cahaya matahari sangat menentukan
produktivitas tanaman. Di dataran yang kurang air hujan menjadi faktor pembatas
adalah kurang air, sebaliknya di daerah yang terlampau banyak hujan, cahaya
menjadi faktor pembatas (Sianturi, 1996).
Tanaman karet tumbuh baik di daerah yang mempunyai curah hujan
mungkin ditanam dengan karet, asal curah hujan turun merata sepanjang tahun.
Pada daerah yang mempunyai curah hujan 5000-6000 mm tanaman karet dapat
tumbuh baik, tetapi hari hujan yang terlalu banyak menyulitkan dalam
pelaksanaan penyadapan dan pencucian tanah sangat efektif, akibatnya akan
menyulitkan pengelolaan produksi pertumbuhan dan banyak kehilangan produksi.
Rata-rata jumlah hari hujan yang diperlukan adalah 100-150 hari hujan per tahun
(Sianturi, 1996).
Tanaman karet dapat tumbuh pada suhu di antara 250C hingga 350C. Suhu
sangat erat kaitannya dengan tinggi tempat. Setiap naik 100 m tinggi tempat dari
permukaan laut maka suhu akan turun 0,50C. Rata-rata suhu di dataran rendah
sekitar 280C, kondisi suhu yang demikian terbaik untuk pertumbuhan tanaman
karet. Dengan penurunan suhu 10C menyebabkan tanaman karet lebih lambat
disadap 3 sampai dengan 6 bulan.
Tanah
Tanaman karet dapat tumbuh pada berbagai jenis tanah, baik pada tanah
vulkanis muda maupun vulkanis tua. Jenis tanah alluvial adalah jenis tanah yang
cukup baik untuk pertumbuhan tanaman karet atau juga tanah gambut.
Tanah-tanah alluvial umumnya memiliki sifat kimia yang cukup baik, tetapi sifat fisisnya
terutama airase dan drainasenya kurang baik, pembuatan saluran drainase akan
menolong perbaikan tanah ini. Begitu juga dengan tanah vulkanis mempunyai
sifat fisika yang cukup baik terutama struktur, tekstur, solum, kedalaman air
tanah, airase dan drainasenya, tetapi sifat kimianya secara umum kurang baik
Sebagian besar perkebunan karet di pulau Jawa terletak pada ketinggian
sekitar 400 m dan di luar pulau Jawa, sebagian besar perkebunan karet pada
ketinggian 0-400 m di atas permukaan laut (Dijkman, 1951).
Menurut Setyamidjaja (1993) sifat-sifat tanah yang cocok untuk tanaman
karet sebagai berkut:
- Solum cukup dalam, sampai 100 cm atau lebih, tidak terdapat batu-batuan.
- Airase dan drainase baik
- Remah, porus dan dapat menyimpan air
- Tekstur tanah terdiri dari atas 35 % liat dan 30 % pasir
- Tidak bergambut, dan jika ada tidak lebih tebal dari 20 cm
- Kandungan unsur hara N, P dan K cukup dan tidak berkurang unsur mikro
- pH 4,5-6,5
- kemiringan tidak lebih dari 16 %
Peningkatan Produktivitas Melalui Pemuliaan Tanaman Karet
Tahapan awal pada pemuliaan tanaman karet adalah memilih tanaman
terbaik di pembibitan atau Seedling Evaluation Trial (SET). Seleksi dilakukan
terhadap peubah amatan yang utama yaitu potensi produksi (lateks dan kayu) dan
pertumbuhan seperti lilit batang, tinggi tanaman, jumlah payung, tebal kulit,
anatomi kulit (jumlah dan diameter pembuluh lateks), indeks penyumbatan, dan
DRC (Woelan dan Azwar, 1990; Annamma Varghese, et al., 1993). Tinggi
tanaman, diameter batang, jumlah payung daun, dan jumlah tangkai daun pada
umumnya diamati sampai dengan umur 2 tahun. Lilit batang dan hasil lateks
(dengan metode testateks) yang pengamatannya dilakukan pada umur 2 s/d 5
tanaman berumur 5 tahun. Peubah amatan untuk pertumbuhan umumnya kurang
berkorelasi positif dengan hasil lateks, pengamatan pertumbuhan pada umumnya
dilakukan untuk menentukan lamanya masa Tanaman Belum Menghasilkan
(TBM). Beberapa hasil penelitian sebelumnya menunjukkan bahwa karakter
jumlah dan diameter pembuluh lateks, indeks penyumbatan, kadar sukrosa lateks
dan kadar tiol merupakan peubah yang berhubungan erat dengan potensi produksi
lateks (Gomez et al., 1972; Ho, 1976; Millford et al., 1969; Premakumari et al.,
1996).
Genotipe yang terseleksi diperbanyak secara vegetatif dan kemudian
dievaluasi pada beberapa tahapan yaitu: pengujian pendahuluan, pengujian
lanjutan, dan pengujian adaptasi. Dengan demikian, sebagai tahapan dari kegiatan
pemuliaan maka kegiatan pengujian potensi produksi sejak awal pengujian sampai
pengujian adaptasi klon harapan perlu dilakukan secara sistematis dan
berkesinambungan (Tan, 1987; Simmonds, 1989).
Berdasarkan aktivitas pemuliaan dan seleksi tanaman karet, maka klon
unggul yang telah dihasilkan dibagi menjadi empat generasi, yaitu:
- Generasi-1 (1910-1935) : Seedling Selected
- Generasi-2 (1935-1960) : Tjir 1, PR 107, GT 1, AVROS 2037
- Generasi-3 (1960-1985) : BPM 1, BPM 107, PR 255, TM 2
- Generasi-4 (1985-2010) : IRR 104, IRR 112, IRR 118, IRR 208, IRR 220
Lamanya siklus pemuliaan tanaman karet yang mencapai 25-30 tahun
merupakan suatu kendala yang secara terus-menerus dihadapi. Beberapa peneliti
mencoba untuk memanfaatkan teknologi baru seperti pengujian plot promosi
memperpendek siklus seleksi tanaman karet terus dilakukan yaitu dengan mencari
beberapa komponen produksi yang berkaitan dengan produksi lateks. Menurut
Narayanan, et al., (1973) bahwa pembuluh lateks, tebal kulit batang, dan lingkar
batang saling berhubungan dan mempunyai peranan yang besar terhadap
pendugaan produksi.
Pemuliaan, seleksi, dan perbanyakan klon dari tanaman unggul merupakan
tahapan awal yang sangat penting yang digunakan untuk mengestimasi beberapa
perbaikan karakter seperti pertumbuhan, produksi dan sifat-sifat sekunder
(Simmonds, 1989). Produksi merupakan objek paling dominan dalam program
perbaikan genetik melalui persilangan. Produksi diukur sebagai berat kadar karet
per unit area per satuan waktu (Simmonds, 1989). Sedangkan karakteristik
pertumbuhan yang dapat mendukung produksi diantaranya kejaguran yang
hubungannya terhadap ketahanan terhadap angin, morfologi kulit, jumlah
pembuluh lateks, dan ketahanan terhadap penyakit. Keempat karakter
pertumbuhan tersebut mempunyai peranan yang sangat besar sekali terhadap
produksi suatu klon (Simmonds, 1989).
Tetua unggul adalah tetua atau kultivar terbaik yang dianjurkan memiliki
produksi tinggi dan sifat sekunder yang lebih baik. Namun keungulan tetua karet
terbukti sering tidak berlaku umum pada semua lokasi atau lingkungan. Karakter
produksi dipengaruhi oleh sifat genetik dan juga oleh adanya interaksi lingkungan
(Sumarmadji, et al., 2005).
Pemanfaatan tetua unggul sebagai komponen teknologi, memberikan
proporsi yang besar dalam upaya meningkatkan efisiensi melalui peningkatan
produktifitas kebun dapat mencapai 1400-2000 kg/ha/th dibanding tanaman asal
biji (semaian) yang hanya 400-500 kg/ha/th. Kendala yang dihadapi bahwa
optimasi potensi produksi tetua di pertanaman komersial dapat sangat bervariasi.
Yang disebabkan produksi tetua unggul per satuan luas sangat tergantung kepada
faktor lingkungan yaitu lingkungan fisik, biologi maupun manajemen kebun.
Karena itu penanaman tetua yang sesuai dengan lingkungan tumbuhnya serta
dengan manajemen yang tepat, akan menghasilkan produktifitas yang optimal
(Aidi-Daslin, et al., 1995).
Paradigma baru yang telah disepakati bersama dalam pembangunan kebun
karet ialah menanam karet tidak hanya untuk menghasilkan lateks, tetapi juga
kayu yang bertujuan untuk meningkatkan produktivitas lahan, meningkatkan
pendapatan, dan pada gilirannya meningkatkan daya saing. Optimalisasi hasil
lateks dan kayu karet, dapat dilakukan terhadap perbaikan teknik budidayanya
yaitu melalui sistem tanam dengan meningkatkan populasi tanaman per hektar dan
jenis tetua. Volume kayu karet yang diperoleh pada saat peremajaan, dengan
populasi awal ± 500 ph/ha yaitu sebesar 180-200 m3/ha. volume kayu masih dapat
ditingkatkan sampai menjadi ± 350 m3/ha tanpa mengurangi hasil lateks dengan
pengaturan sistem tanam, mempertinggi populasi awal, dan dengan menggunakan
tetua anjuran lateks kayu (Siagian dan Aidi-Daslin, 2003).
Selama tiga generasi pemuliaan karet (1910-1985) telah dihasilkan
sejumlah tetua unggul yang memiliki potensi karet kering dari mulai rata-rata 500
kg/ha/th menjadi 2500 kg/ha/th. Pada saat ini, paradigma berkebun karet telah
berubah dari menghasilkan lateks menjadi menghasilkan lateks-kayu, karena kayu
Karena itu sasaran program pemuliaan pada generasi keempat (1985-2010)
yang sedang berjalan sampai saat ini, selain bertujuan untuk menghasilkan
tetua-tetua unggul sebagai penghasil lateks juga lateks-kayu
(Aidi-Daslin dan Lasminingsih, 2001).
Tetua IRR 5, IRR 21, IRR 32, IRR 39, IRR 42 dan IRR 118 merupakan
tetua karet unggul terbaru seri IRR sebagai penghasil lateks dan kayu untuk
anjuran penanaman komersial. Tetua-tetua tersebut di atas memiliki pertumbuhan
awal yang cepat, sehingga dengan tingkat rata-rata pertumbuhan yang normal
dapat disadap pada umur kurang dari 5 tahun (Siagian dan Aidi-Daslin, 2003).
Tetua penghasil lateks dikategorikan sebagai tetua penghasil awal cepat
(quick starter) dengan pola produksi awal tinggi dan rataan pertumbuhan batang
sedang, baik pada masa TBM maupun TM. Tetua tipe ini umumnya kurang
respons terhadap stimulan dan pada umumnya agak rentan terhadap kepatahan
batang (Azwar dan Suhendry, 1998).
Volume kayu karet sangat ditentukan oleh besaran lilit batang dan tinggi
tanaman maka volume kayu karet yang dihasilkan semakin besar dan sebaliknya
semakin kecil lilit batang dan ketinggian tanaman maka volume kayu yang
dihasilkan semakin kecil. Demikian halnya dengan semakin tinggi cabang primer
dan tebal kulit maka kayu log yang dihasilkan semakin besar (Wan Razali Mohd
et al, 1983).
Tetua penghasil lateks-kayu dikategorikan sebagai tetua penghasil awal
yang moderat dengan pola produksi lanjutan mendatar atau meninggi, dan dengan
pertumbuhan yang sedang, tipe tetua ini memiliki produksi kayu yang tidak terlalu
yang rendah (slow starter) dengan pola produksi lanjutan rendah sampai sedang,
tetapi tetua tipe ini memiliki rataan pertumbuhan yang cepat baik pada masa TBM
dan TM sehingga volume kayu yang dihasilkan pada saat peremajaan cukup tinggi
(Aidi-Daslin, 2005).
Keragaman Genotipe dan Fenotipe
Perbedaan kondisi lingkungan memberikan kemungkinan munculnya
variasi yang akan menentukan penampilan akhir tanaman tersebut. Bila ada
variasi yang timbul atau tampak pada populasi tanaman yang ditanam pada
kondisi lingkungan yang sama maka variasi tersebut merupakan variasi atau
perbedaan yang berasal dari genotip individu anggota populasi
(Mangoendidjojo, 2003).
Keragaman yang sering ditunjukkan oleh tanaman sering dikaitkan dengan
aspek negatif. Hal ini sering tidak diperhatikan oleh peneliti yang menganggap
bahwa susunan genetik dari bahan tanaman digunakan adalah sama karena berasal
dari varietas yang sama. Keragaman penampilan tanaman akibat perbedaan
susunan genetik selalu mungkin terjadi sekalipun bahan tanaman yang digunakan
berasal dari jenis tanaman yang sama. Jika ada dua jenis tanaman yang sama
ditanam pada lingkungan yang berbeda, dan timbul variasi yang sama dari kedua
tanaman tersebut maka hal ini dapat disebabkan oleh genetik dari tanaman yang
bersangkutan (Sitompul dan Guritno, 1995).
Keragaman genetik alami merupakan sumber bagi setiap program
pemuliaan tanaman. Variasi ini dapat dimanfaatkan, seperti semula dilakukan
manusia, dengan cara melakukan introduksi sederhana dan tehnik seleksi atau
kombinasi genetik yang baru. Jika perbedaan dua individu yang mempunyai
faktor lingkungan yang sama dapat diukur, maka perbedaan ini berasal dari
genotipe kedua tanaman tersebut. Keragaman genetik menjadi perhatian utama
para pemulia tanaman, karena melalui pengelolaan yang tepat dapat menghasilkan
varietas baru yang lebih baik (Welsh, 2005).
Gen-gen tidak dapat menyebabkan berkembangnya karakter terkecuali jika
mereka berada lingkungan yang sesuai, dan sebaliknya tidak ada pengaruh
terhadap perkembangnya karakteristik dengan mengubah tingkat keadaan
lingkungan terkecuali jika gen yang diperlukan ada. Namun, harus disadari bahwa
keragaman yang diamati terhadap sifat-sifat yang terutama disebabkan oleh
perbedaan gen yang dibawa oleh individu yang berlainan dan terhadap variabilitas
di dalam sifat yang lain, pertama-tama disebabkan oleh perbedaan lingkungan
dimana individu berada (Allard, 2005).
Variasi yang ditimbulkan ada yang dapat langsung dilihat, misalnya
adanya perbedaan warna bunga, daun dan bentuk biji (ada yang berkerut, ada
yang tidak), ini yang disebut variasi sifat yang kualitatif. Namun ada pula variasi
yang memerlukan pengamatan dengan pengukuran, misal tingkat produksi,
jumlah anakan, tinggi tanaman, dan lainnya (Mangoendidjojo, 2003).
Heritabilitas
Heritabilitas adalah salah satu alat ukur dalam sistem seleksi yang efisien
yang dapat menggambarkan efektivitas seleksi genotipe berdasarkan penampilan
fenotipenya. Nilai heritabilitas yang tinggi untuk suatu karakter menggambarkan
karakter tersebut lebih ditentukan oleh faktor genetik, karakter yang demikian
korelasi antar karakter fenotipe diperlukan dalam seleksi tanaman, untuk
mengetahui karakter yang dapat dijadikan petunjuk seleksi terhadap produktivitas
yang tinggi (Suharsono et al., 2006; Wirnas et al., 2006).
Heritabilitas adalah hubungan antara ragam genotipe dengan ragam
fenotipenya. Hubungan ini menggambarkan seberapa jauh fenotipe yang tampak
merupakan refleksi dari genotipe. Pada dasarnya seleksi terhadap populasi
bersegregasi dilakukan melalui nilai-nilai besaran karakter fenotipenya. Dalam
kaitan ini, penting diketahui peluang terseleksinya individu yang secara fenotipe
menghasilkan turunan yang sama miripnya dengan individu terseleksi tadi.
Misalkan dalam suatu populasi dijumpai ragam genetik tinggi untuk suatu
karakter dan ragam fenotipenya rendah, maka dapat diramalkan bahwa turunan
individu terseleksi akan mirip dengan dirinya untuk karakter tersebut; dan
sebaliknya. Heritabilitas biasanya dinyatakan dalam persen (%). Heritabilitas
dikatakan tinggi bila nilai H > 50%, sedang apabila nilai H terletak antara
20%-50% dan dikatakan rendah bila nilai H < 20% (Mangoendidjojo, 2003).
Variasi keseluruhan dalam suatu populasi merupakan hasil kombinasi
genotipe dan pengaruh lingkungan. Proporsi variasi merupakan sumber yang
penting dalam program pemuliaan karena dari jumlah variasi genetik ini
diharapkan terjadi kombinasi genetik yang baru. Proporsi dari seluruh variasi yang
disebabkan oleh perubahan genetik disebut heritabilitas. Heritabilitas dalam arti
yang luas adalah semua aksi gen termasuk sifat dominan, aditif, dan epistasis.
Nilai heritabilitas secara teoritis berkisar dari 0 sampai 1. Nilai 0 ialah bila seluruh
seluruh variasi disebabkan oleh faktor genetik. Dengan demikian nilai heritabilitas
akan terletak antara kedua nilai ekstrim tersebut (Welsh, 2005).
Variasi genetik akan membantu dalam mengefisienkan kegiatan seleksi.
Apabila variasi genetik dalam suatu populasi besar, ini menunjukkan individu
dalam populasi beragam sehingga peluang untuk memperoleh genotip yang
diharapkan akan besar (Bahar dan Zein, 1993). Sedangkan pendugaan nilai
heritabilitas tinggi menunjukkan bahwa faktor pengaruh genetik lebih besar
terhadap penampilan fenotip bila dibandingkan dengan lingkungan. Untuk itu
informasi sifat tersebut lebih diperankan oleh faktor genetik atau faktor
lingkungan, sehingga dapat diketahui sejauh mana sifat tersebut dapat diturunkan
pada generasi berikutnya.
Dalam Hanson (1963) menyatakan nilai heritabilitas dalam arti luas
menunjukkan genetik total dalam kaitannya keragaman genotip, sedangkan
menurut Poespodarsono (1988), bahwa makin tinggi nilai heritabilitas satu sifat
makin besar pengaruh genetiknya dibanding lingkungan.
Kemajuan Genetik
Selain menggunakan nilai heritabilitas yang tinggi, juga menggunakan
parameter yang lainnya, yaitu nilai duga kemajuan genetik yang tinggi, sebab nilai
heritabilitas itu sendiri kurang memberikan gambaran sebenarnya mengenai
kemajuan yang diharapkan terhadap genetik. Dengan nilai heritabilitas dan
kemajuan genetik akan didapatkan gambaran terbaik mengenai kemajuan yang
diharapkan dari seleksi (Rachmadi, dkk, 1990).
Menurut Dudley and Moll (1976), nilai heritabilitas dapat memberikan
suatu populasi, sehingga apabila nilai heritabilitas dipadukan dengan nilai
kemajuan genetik dari seleksi maka akan lebih bermanfaat dalam meramalkan
hasil akhir untuk melakukan seleksi sifat individu yang baik.
Seleksi akan menunjukkan kemajuan genetik yang tinggi jika sifat yang
dilibatkan dalam seleksi mempunyai variasi genetik dan heritabilitas yang tinggi.
Jika nilai heritabilitas tinggi, sebagian besar variasi fenotipe disebabkan oleh
variasi genetik, maka seleksi akan memperoleh kemajuan genetik (Zen, 1995).
Dalam Knight (1979) menyatakan informasi mengenai variasi genetik dan
heritabilitas berguna untuk menentukan kemajuan genetik yang diperoleh dari
seleksi. Hayward (1990) menyatakan bahwa sifat-sifat yang dikendalikan oleh
gen-gen bukan aditif menyebabkan kemajuan genetik yang rendah. Hal ini
disebabkan pengaruh tindak gen bukan aditif tidak diwariskan dan akan lenyap
semasa seleksi (Suprapto dan Kairuddin, 2007).
Menurut Burton (1952) menyatakan bahwa pemulia lebih banyak
mempertimbangkan dugaan kemajuan genetik dalam persen di atas nilai rata-rata
populasi. Oleh karena itu sesuai rumus yang disajikan Singh and Chaudhary
(1977) tergambar bahwa KG (%) merupakan produk dari nilai-nilai diferensial
seleksi, heritabilitas yang menentukan efisiensi sistem seleksi sehingga seleksi
akan efektif bila nilai kemajuan genetik tinggi ditunjang oleh salah satu nilai KVG
METODE PENELITIAN
Tempat dan Waktu
Penelitian ini dilaksanakan di Kebun Percobaan tanaman F1 hasil okulasi
(turunan) dari persilangan intraspesifik RRIM 600 x PN 1546 di Balai Penelitian
Sungei Putih-Pusat Penelitian Karet, Kecamatan Galang, Kabupaten Deli
Serdang-Sumatera Utara, dengan ketinggian tempat ± 54 meter di atas permukaan
laut. Pelaksanaan penelitian ini dilakukan pada bulan Januari 2011 sampai dengan
Maret 2011.
Bahan dan Alat
Bahan penelitian yang digunakan adalah genotipe (tanaman F1) hasil
persilangan intraspesifik dari klon RRIM 600 x PN 1546 sebanyak 25 genotipe
dan 2 tetua (RRIM 600 sebagai tetua betina dan PN 1546 sebagai tetua jantan).
Genotipe yang digunakan tersebut ditanam pada tahun 2008 dan ditanam dengan
jarak 5 m x 4 m. Bahan kimia alkohol, sudan III, FAA (Formalin Acetic Acid),
KOH 15%, HNO3, kertas saring serta bahan-bahan lain yang mendukung
penelitian ini.
Alat yang digunakan dalam penelitian ini yaitu : meteran kain, alat ukur
tinggi tanaman, scliper, alat bor kulit, timbangan analitik, pisau silet, mikroskop,
deck glass, cover glass, gelas ukur, tabung gelas, sendok, stirer, oven, dan alat-alat
Metode Penelitian
Penelitian ini menggunakan rancangan acak kelompok (RAK) nonfaktorial
yang terdiri dari 25 genotipe dan 2 tetua.
Jumlah blok (ulangan) = 3 blok
Jumlah plot / blok = 27 plot
Jumlah tanaman per plot = 5 tanaman
Jumlah sampel per plot = 5 tanaman
Jumlah tanaman sampel = 405 tanaman
Jumlah tanaman seluruhnya = 405 tanaman
Data hasil penelitian dianalisis dengan sidik ragam dengan model linier
sebagai berikut:
Yij = µ+ ρi+ δj + εij
Dimana:
Yij : nilai pengamatan pada blok ke-i dalam genotipe ke-j
µ : nilai tengah (nilai rata-rata umum)
ρi : pengaruh blok ke-i
δj : pengaruh genotipe ke-j
εij : pengaruh galat terhadap blok ke-i pada genotipe ke-j
Data hasil penelitian pada genotipe yang berbeda nyata dilanjutkan dengan
uji beda rataan menurut Uji Jarak Berganda Duncan atau duncan multiple range
Tabel 1. Analisis Ragam dan Pendugaan Komponen Ragam
1. Keragaman Genotipe dan Fenotipe
Keragaman sifat dihitung melalui analisis sidik ragam yang dikemukakan
oleh Singh and Chaudary (1977) dalam Tempake dan Luntungan (2002) adalah
sebagai berikut :
x = Rataan populasi
KKG = Koefisien Keragaman Genetik KKF = Koefisien Keragaman Fenotipe
Kriteria keragaman menurut Murdaningsih, dkk (1990) dalam Tempake
dan Luntungan (2002) adalah :
Rendah = 0 – 25% dari Koefisien Keragaman Genetik (KKG) tertinggi
Sedang = 25 – 50% dari Koefisien Keragaman Genetik (KKG) tertinggi
Tinggi = 50 – 75% dari Koefisien Keragaman Genetik (KKG) tertinggi
Sangat Tinggi = 75 – 100% dari Koefisien Keragaman Genetik (KKG) tertinggi
2. Heritabilitas
Heritabilitas dari seluruh sampel dihitung dengan rumus :
e
Menurut Stansfield (1991) kriteria heritabilitas adalah sebagai berikut :
Heritabilitas tinggi > 0,5
Heritabilitas sedang = 0,2 – 0,5
Heritabilitas rendah < 0,2
3. Kemajuan Genetik
Harapan Kemajuan Genetik (HKG) dapat dihitung dan diduga menurut
cara sebagai berikut :
)
K = Konstanta 2,06 untuk intensitas seleksi 0,05
4. Seleksi Genotipe Berdasarkan Produksi Lateks dan Kayu
Secara umum telah diketahui bahwa populasi semaian F1 menyebar secara
normal, sehingga intensitas seleksi yang dipergunakan adalah areal yang terletak
pada 10% dan 1% di sebelah kanan kurva normal. Pendugaan tersebut dilakukan
dengan memanfaatkan rumus sebaran Z sebagai berikut:
SD X Z = −µ
Keterangan:
Z = konstanta 1,32 untuk seleksi 10% dan 2,48 untuk seleksi 1% pada 25 genotipe yang akan diseleksi
X = nilai minimum untuk parameter seleksi µ = nilai rata-rata populasi
SD = simpangan baku
Hasil seleksi 10% genotipe terbaik akan masuk dalam Pengujian
Pendahuluan dan 1% genotipe terbaik masuk dalam Pengujian Plot Promosi.
5. Analisis Korelasi Antar Peubah Amatan Terhadap Produksi
Nilai korelasi (rij) dapat dicari dengan menggunakan rumus sebagai
berikut:
Penafsiran nilai rij dapat juga dilakukan dengan menggunakan program
PELAKSANAAN PENELITIAN
Persiapan Areal
Membersihkan areal pertanaman dari gulma-gulma agar mudah melakukan
kegiatan penelitian. Bagan penelitian terlampir pada lampiran 2.
Sensus Tanaman
Untuk mengetahui jumlah dan kondisi seluruh populasi tanaman di
lapangan, dilakukan sensus tanaman dengan cara menomori masing-masing
tanaman.
Membuat Batas Tinggi Penyadapan
Untuk menentukan batas tinggi penyadapan, terlebih dahulu dibuat batas
tinggi penyadapan dengan melingkari batang tanaman pada ketinggian 50 cm dari
permukaan tanah. Penandaan ini dibuat dengan menggunakan spidol atau cat.
Menyiapkan Penampungan Lateks
Penampungan lateks dibuat dengan kertas saring yang berukuran 7,5 cm x
10 cm yang direkatkan dengan lakban (selotip) pada batang tanaman dengan
ketinggian 50 cm dari permukaan tanah (sesuai ketinggian penyadapan).
Penyadapan
Penyadapan dilakukan pada pagi hari (pukul 07.00 - 11.00) dengan
metode testateks yang menggunakan pisau penusuk kulit yang dibuat mirip
dengan alat ciptaan Cramer. Jarak antara pisau masing-masing 2 cm. Pisau ini
ditekan ke kulit batang tanaman sehingga lateks akan mengalir keluar dari luka.
Lateks ditampung kemudian dibawa ke laboratorium untuk ditimbang berat
Pengamatan Parameter Tinggi Tanaman (m)
Tinggi tanaman diukur dengan menggunakan alat ukur berskala yang
diukur dari permukaan tanah sampai ke titik tumbuh ujung tanaman. Pengamatan
dilakukan 1 kali selama penelitian.
Jumlah Cabang Primer (cabang)
Jumlah cabang primer dihitung pada batang utama untuk setiap tanaman.
Pengamatan dilakukan 1 kali selama penelitian.
Tinggi Cabang Pertama (m)
Tinggi cabang pertama diukur dengan menggunakan alat ukur berskala
yang diukur dari permukaan tanah sampai ke titik pangkal cabang pertama
tanaman. Pengamatan dilakukan 1 kali selama penelitian.
Lilit Batang (cm)
Lilit batang diukur pada ketinggian 50 cm di atas permukaan tanah
menggunakan meteran kain (ukuran 100 cm). Semua genotipe F1 hasil okulasi
(turunan) hasil persilangan RRIM 600 x PN 1546 diukur untuk mengetahui
besaran lilit batang yang dimiliki. Pengamatan dilakukan 1 kali selama penelitian.
Tebal Kulit (mm)
Kulit diambil dengan menggunakan alat pelubang kulit yang terbuat dari
besi dengan diameter 1 cm. Sedangkan alat yang digunakan untuk mengukur
ketebalan kulit adalah scliper. Contoh kulit diambil 5 cm di atas batas bidang
penyadapan. Untuk menghitung tebal kulit yang sebenarnya, skala yang
yang diambil untuk masing-masing genotipe berasal dari 2 tanaman sampel yang
dipilih berdasarkan kondisi fisik (fenotipik) tanaman.
Jumlah dan Diameter Pembuluh Lateks
Untuk menghitung jumlah dan diameter pembuluh lateks dilakukan
menurut metode Gomez et al. (1972). Pembuatan preparat pembuluh lateks
dilakukan dilaboratorium Fisiologi dan Penyakit Balai Penelitian Sungei Putih.
Contoh kulit yang telah diambil untuk pengukuran tebal kulit sekaligus digunakan
untuk pengukuran jumlah dan diameter pembuluh lateks. Contoh kulit yang
diambil langsung dari lapangan difiksasi dalam larutan FAA (Formalin Acetic
Acid) yang ditempatkan pada botol plastik atau pirex. Contoh kulit yang diambil
diberi tanda dengan pensil 2B sesuai nomor masing-masing genotipe. Larutan
FAA merupakan campuran dari 10 ml formalin 40%, 5 ml asam asetat glasial, 70
ml alkohol absolut (95%) dan 15 ml aquades. Selanjutnya kulit dibawa ke
laboratorium untuk langsung dibuat preparat atau disimpan terlebih dahulu.
Pembuatan preparat semi permanen, kulit yang telah difiksasi dipindahkan
dan dimasukkan ke dalam larutan KOH 15% selama 1 jam, kemudian dibilas
dengan aquades atau air mengalir selama 5 menit dan dikeringkan dengan kertas
tisue. Selanjutnya direndam ke dalam larutan HNO3 selama 2 jam dan dibilas lagi
dengan aquades atau air mengalir selama 5 menit dan dikeringkan dengan kertas
tisue. Untuk tahapan berikutnya atau proses selanjutnya direndam dalam larutan
alkohol 70% selama 15 menit, dibilas kembali dengan aquades atau air mengalir
dan dikeringkan dengan dihisap menggunakan kertas tisue agar tidak ada
kandungan airnya, dan terakhir untuk dapat dilihat di bawah mikroskop diberikan
campuran 15 gram KOH ditambahkan 85 ml aquades. Sedangkan untuk larutan
HNO3, dibuat perbandingan volume antara HNO3 pekat dan aquades 1:2 dan
larutan alkohol 70% adalah campuran dari 70 ml alkohol absolut dan 30 ml
aquades. Setelah itu dilakukan pewarnaan, kemudian preparat diiris dengan
menggunakan pisau silet yang tajam secara membujur untuk melihat jumlah
pembuluh, sedangkan untuk melihat diameter pembuluh kulit dipotong melintang.
Masing-masing irisan tipis dari pereparat tersebut di letakkan di gelas objek dan
ditutup dengan gelas penutup (cover glass) yang sebelumnya diberikan gliserin
untuk menjaga peraparat itu tidak mengering. Setelah itu dilakukan pengamatan
dibawah mikroskop dan diamati dengan pembesaran 10 x. Dengan pemberian
warna Sudan III, maka jaringan pembuluh lateks akan berwarna merah cerah yang
dapat dilihat perbedaan dengan jaringan lain yang tidak mengalami perubahan
warna. Pengamatan terhadap jumlah dan diameter pembuluh lateks dilakukan
secara bersamaan. Diameter pembuluh lateks diukur dengan menggunakan skala
okuler. Besaran skala diukur mulai dari nol sampai dengan skala yang berimpit
dan satu ukuran skala di mikroskop setara dengan satu mikron, dengan demikian
ukuran diameter pembuluh lateks dapat diketahui. Perbesaran yang digunakan
adalah 40 x.
Produksi Karet Kering (g/p/s)
Produksi karet kering diukur dengan cara menimbang berat kering lateks
yang telah ditampung pada kertas saring. Berat kering lateks ditimbang setelah
lateks dikeringkan di dalam oven pada suhu 700C selama 24 jam.
Produksi kayu diukur dengan menggunakan rumus yang dikembangkan
oleh Wan Razali Mohd et al, (1983) yaitu sebagai berikut:
(
)
{
LB}
TTVp =0,0435485+ 0,00005031× /π 2 ×
Keterangan:
Vp = volume pohon LB = lilit batang TT = tinggi tanaman
HASIL DAN PEMBAHASAN
Hasil
Tinggi Tanaman (m)
Data pengamatan serta daftar analisis sidik ragam tinggi tanaman dapat
dilihat pada Lampiran 4 dan 5. Hasil analisis statistika dengan metode sidik ragam
tersebut menunjukkan bahwa genotipe berbeda nyata terhadap tinggi tanaman.
Hasil uji beda rataan 25 genotipe dan tetua terhadap tinggi tanaman dapat
dilihat pada Tabel 2 di bawah ini .
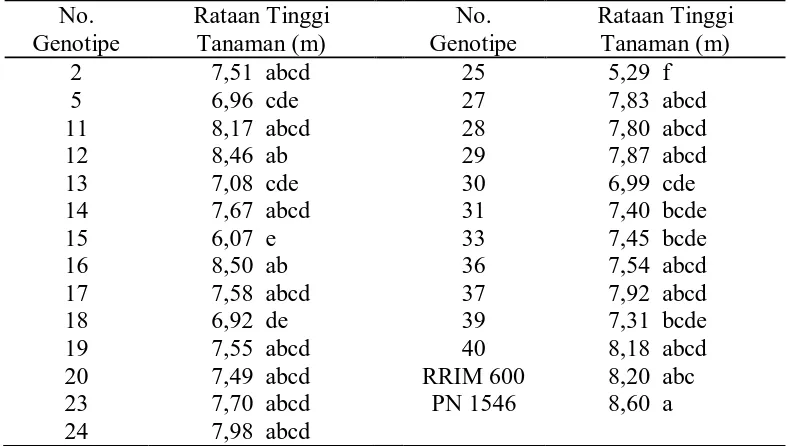
Tabel 2. Rataan Tinggi Tanaman dari 25 Genotipe dan Tetua No.
Keterangan : Angka-angka yang diikuti oleh notasi yang sama tidak berbeda nyata menurut Uji Duncan Multiple Range Test (DMRT) pada taraf 5%.
Dari tabel di atas terlihat bahwa tanaman paling tinggi terdapat pada klon
primer PN 1546 yaitu 8,60 m yang berbeda nyata terhadap genotipe no. 5, 13, 15,
18, 25, 30, 31, 33, dan 39, dan tanaman terendah terdapat pada genotipe no. 25
Jumlah Cabang Primer (cabang)
Data pengamatan serta daftar analisis sidik ragam jumlah cabang primer
dapat dilihat pada Lampiran 6 dan 7. Hasil analisis statistika dengan metode sidik
ragam tersebut menunjukkan bahwa genotipe tidak berbeda nyata terhadap jumlah
cabang primer.
Rataan 25 genotipe dan tetua terhadap jumlah cabang primer dapat dilihat
pada Tabel 3 di bawah ini.
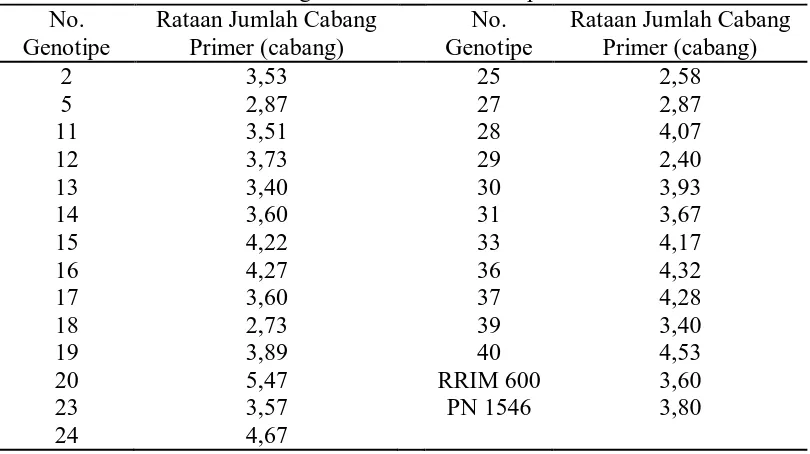
Tabel 3. Rataan Jumlah Cabang Primer dari 25 Genotipe dan Tetua No.
Dari Tabel 3 dapat dilihat bahwa genotipe yang memiliki jumlah cabang
primer tertinggi pada genotipe no. 20 yaitu 5,47 cabang dan terendah pada
genotipe no. 29 yaitu 2,4 cabang.
Tinggi Cabang Pertama (m)
Data pengamatan serta daftar analisis sidik ragam tinggi cabang pertama
ragam tersebut menunjukkan bahwa genotipe tidak berbeda nyata terhadap tinggi
cabang pertama.
Rataan 25 genotipe dan tetua terhadap tinggi cabang pertama dapat dilihat
pada Tabel 4 di bawah ini.
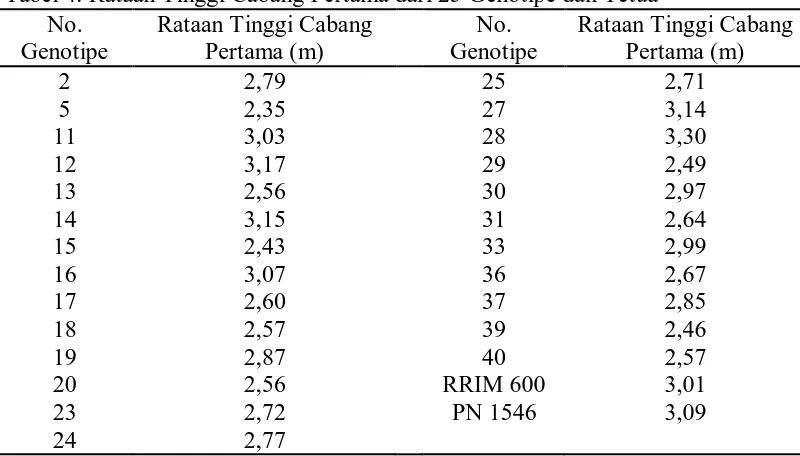
Tabel 4. Rataan Tinggi Cabang Pertama dari 25 Genotipe dan Tetua No.
Dari tabel 4 dapat dilihat bahwa tinggi cabang pertama tertinggi terdapat
pada genotipe no. 28 yaitu 3,3 m dan terendah terdapat pada genotipe no. 5 yaitu
2,35 m.
Lilit Batang (cm)
Data pengamatan serta daftar analisis sidik ragam lilit batang dapat dilihat
pada Lampiran 10 dan 11. Hasil analisis statistika dengan metode sidik ragam
tersebut menunjukkan bahwa genotipe berbeda nyata terhadap lilit batang.
Hasil uji beda rataan 25 genotipe dan tetua terhadap lilit batang dapat
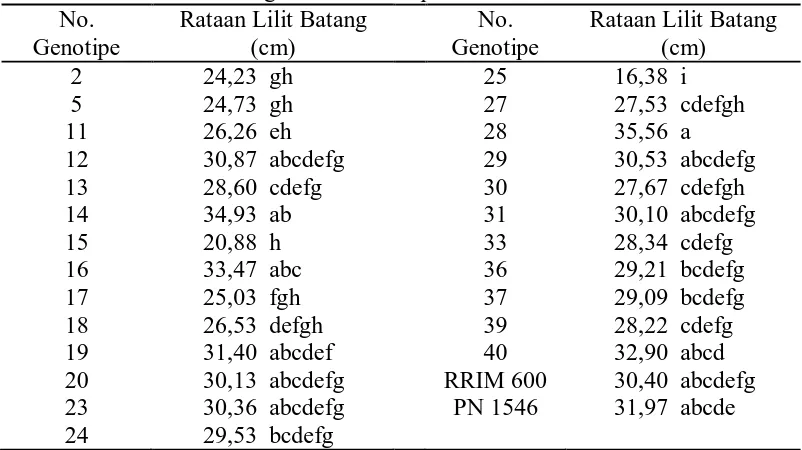
Tabel 5. Rataan Lilit Batang dari 25 Genotipe dan Tetua
Keterangan : Angka-angka yang diikuti oleh notasi yang sama tidak berbeda nyata menurut Uji Duncan Multiple Range Test (DMRT) pada taraf 5%.
Dari tabel 5 dapat dilihat bahwa lilit batang tertinggi terdapat pada
genotipe no. 28 yaitu 35,56 cm yang berbeda nyata terhadap genotipe no. 2, 5, 11,
13, 15, 17, 18, 24, 25, 27, 30, 33, 36, 37 dan 39, dan lilit batang terendah terdapat
pada genotipe no. 25 yaitu 16,38 cm yang berbeda nyata terhadap genotipe
lainnya.
Tebal Kulit (mm)
Data pengamatan serta daftar analisis sidik ragam tebal kulit dapat dilihat
pada Lampiran 12 dan 13. Hasil analisis statistika dengan metode sidik ragam
tersebut menunjukkan bahwa genotipe berbeda nyata terhadap tebal kulit.
Hasil uji beda rataan 25 genotipe dan tetua terhadap tebal kulit dapat
Tabel 6. Rataan Tebal Kulit dari 25 Genotipe dan Tetua
Keterangan : Angka-angka yang diikuti oleh notasi yang sama tidak berbeda nyata menurut Uji Duncan Multiple Range Test (DMRT) pada taraf 5%.
Dari tabel di atas terlihat bahwa tebal kulit tertinggi terdapat pada genotipe
no. 14 yaitu 3,83 mm yang berbeda nyata terhadap genotipe no. 2, 5, 15, 17, 18,
25, 27, 29, 31, 37, 39, klon RRIM 600 dan PN 1546, dan tebal kulit terendah
terdapat pada genotipe no. 25 yaitu 1,92 mm yang berbeda nyata terhadap
genotipe lainnya.
Jumlah Pembuluh Lateks
Data pengamatan serta daftar analisis sidik ragam jumlah pembuluh lateks
dapat dilihat pada Lampiran 14 dan 15. Hasil analisis statistika dengan metode
sidik ragam tersebut menunjukkan bahwa genotipe berbeda nyata terhadap jumlah
pembuluh lateks.
Hasil uji beda rataan 25 genotipe dan tetua terhadap jumlah pembuluh
Tabel 7. Rataan Jumlah Pembuluh Lateks dari 25 Genotipe dan Tetua No.
Genotipe
Rataan Jumlah Pembuluh Lateks
No. Genotipe
Rataan Jumlah Pembuluh Lateks
2 2,50 ef 25 3,08 cdef
5 2,17 f 27 3,83 bcde
11 4,08 abcd 28 5,17 a
12 3,67 bcdef 29 4,92 ab
13 3,50 cdef 30 3,42 cdef
14 4,42 abc 31 5,00 ab
15 3,42 abc 33 2,75 def
16 3,83 bcde 36 3,08 cdef
17 4,33 abc 37 4,00 abcde
18 3,00 def 39 4,08 abcd
19 4,50 abc 40 4,50 abc
20 3,83 bcde RRIM 600 3,50 cdef
23 4,33 abc PN 1546 2,17 f
24 4,08 abcd
Keterangan : Angka-angka yang diikuti oleh notasi yang sama tidak berbeda nyata menurut Uji Duncan Multiple Range Test (DMRT) pada taraf 5%.
Gambar 1. Sayatan Longitudinal Pembuluh Lateks
Dari tabel 7 dapat dilihat bahwa jumlah pembuluh lateks tertinggi terdapat
pada genotipe no. 28 yaitu 5,17 yang berbeda nyata terhadap genotipe no. 2, 5, 12,
13, 16, 18, 20, 25, 27, 30, 33, 36, klon RRIM 600 dan PN 1546, dan jumlah
2,17 yang berbeda nyata terhadap genotipe no. 11, 14, 15, 16, 17, 19, 20, 23, 24,
27, 28, 29, 31, 37, 39 dan 40.
Gambar 2. Sayatan Transversal Pembuluh Lateks
Diameter Pembuluh Lateks (mµ)
Data pengamatan serta daftar analisis sidik ragam diameter pembuluh
lateks dapat dilihat pada Lampiran 16 dan 17. Hasil analisis statistika dengan
metode sidik ragam tersebut menunjukkan bahwa genotipe tidak berbeda nyata
terhadap diameter pembuluh lateks.
Rataan 25 genotipe dan tetua terhadap diameter pembuluh lateks dapat
Tabel 8. Rataan Diameter Pembuluh Lateks dari 25 Genotipe dan Tetua
Dari tabel 8 dapat dilihat bahwa diameter pembuluh lateks tertinggi
terdapat pada genotipe no. 31 dan klon RRIM 600 yaitu 18,75 mµ dan terendah
terdapat pada genotipe no. 12 dan klon PN 1546 yaitu 17,25 mµ.
Produksi Karet Kering (g/p/s)
Data pengamatan serta daftar analisis sidik ragam produksi karet kering
dapat dilihat pada Lampiran 18 dan 19. Hasil analisis statistika dengan metode
sidik ragam tersebut menunjukkan bahwa genotipe berbeda nyata terhadap
produksi karet kering.
Hasil uji beda rataan 25 genotipe dan tetua terhadap produksi karet kering
Tabel 9. Rataan Produksi Karet Kering dari 25 Genotipe dan Tetua
Keterangan : Angka-angka yang diikuti oleh notasi yang sama tidak berbeda nyata menurut Uji Duncan Multiple Range Test (DMRT) pada taraf 5%.
Dari tabel 9 dapat dilihat bahwa produksi karet kering tertinggi terdapat
pada genotipe no. 40 yaitu 0,409 g/p/s yang berbeda nyata terhadap genotipe no.
2, 5, 11, 12, 14, 15, 16, 17, 18, 20, 24, 25, 27, 28, 29, 30, 33 dan klon PN 1546,
dan produksi karet kering terendah terdapat pada klon PN 1546 yaitu 0,068 g/p/s
yang berbeda nyata terhadap genotipe no. 13, 14, 19, 20, 23, 27, 28, 29, 31, 36,
37, 39, 40 dan klon RRIM 600.
Produksi Kayu (cm3/pohon)
Data pengamatan serta daftar analisis sidik ragam produksi karet kering
dapat dilihat pada Lampiran 20 dan 21. Hasil analisis statistika dengan metode
sidik ragam tersebut menunjukkan bahwa genotipe berbeda nyata terhadap
produksi kayu.
Hasil uji beda rataan 25 genotipe dan tetua terhadap produksi kayu dapat
Tabel 10. Rataan Produksi Kayu dari 25 Genotipe dan Tetua
Keterangan : Angka-angka yang diikuti oleh notasi yang sama tidak berbeda nyata menurut Uji Duncan Multiple Range Test (DMRT) pada taraf 5%.
Dari tabel 10 dapat dilihat bahwa produksi kayu tertinggi terdapat pada
genotipe no. 28 yaitu 0,04380571 cm3/pohon yang berbeda nyata terhadap
genotipe no. 2, 5, 11, 13, 15, 17, 18, 19, 20, 23, 24, 25, 27, 29, 30, 31, 33, 36, 37,
39 dan klon RRIM 600 dan produksi kayu terendah terdapat pada genotipe no. 25
yaitu 0,04358837 cm3/pohon yang berbeda nyata terhadap genotipe lainnya.
Pendugaan Parameter Genetik
Hasil perhitungan nilai pendugaan komponen ragam genotipe (σ2g) dan
ragam fenotipe (σ2p), koefisien keragaman genetik (KKG), heritabilitas (h2) serta
nilai harapan kemajuan genetik (HKG) disajikan pada Tabel 11. Pentingnya nilai
komponen ragam genotipe adalah untuk menunjukkan tingkat penampilan
genotipe/klon tanaman dan besarnya akan mempengaruhi KKG maupun nilai h2.
Nilai KKG berkisar antara 1,33% - 40,49%, nilai KKF 2,47% - 44,74%
Tabel 11. Nilai pendugaan komponen ragam genotipe (σ2g), ragam fenotipe (σ2p), koefisien keragaman genetik (KKG), koefisien keragaman fenotipe (KKF), heritabilitas (h2) dan harapan kemajuan genetik (HKG)
Karakter σ2g σ2p KKG (%) KKF (%) h2 HKG
Tabel 11 menunjukkan nilai duga heritabilitas berkisar antara 0,230 –
0,819. Berdasarkan kriteria heritabilitas diperoleh dua parameter yang mempunyai
nilai duga heritabilitas sedang yaitu pada parameter tinggi cabang pertama (0,23)
dan diameter pembuluh lateks (0,291), enam parameter mempunyai nilai duga
heritabilitas tinggi yaitu pada parameter produksi karet kering (0,819), tinggi
tanaman (0,687), lilit batang (0,751), tebal kulit (0,672), jumlah pembuluh lateks
(0,682) dan produksi kayu (0,675).
Hubungan Karakter Agronomi Terhadap Produksi Karet Kering
Tabel 12. Matriks Korelasi Karakter Agronomi Terhadap Produksi Karet Kering
Karakter Lilit