STUDI KARAKTER FISIOLOGIS DAN SIFAT ALIRAN LATEKS
KLON KARET (Hevea brasiliensis Muell Arg.) IRR SERI 300
SKRIPSI
Oleh:
FAUZI KURNIA
050307023/PEMULIAAN TANAMAN
DEPARTEMEN BUDIDAYA PERTANIAN
FAKULTAS PERTANIAN
STUDI KARAKTER FISIOLOGIS DAN SIFAT ALIRAN LATEKS
KLON KARET (Hevea brasiliensis Muell Arg.) IRR SERI 300
SKRIPSI
Oleh:
FAUZI KURNIA
050307023/PEMULIAAN TANAMAN
Skripsi Sebagai Salah Satu Syarat Untuk Memperoleh Gelar Sarjana Pertanian, Fakultas Pertanian Universitas Sumatera Utara, Medan
Di Setujui Oleh:
Ketua Komisi Pembimbing (Ir. Yusuf Husni)
NIP : 19560821 1198603 1 001
DEPARTEMEN BUDIDAYA PERTANIAN
FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA
MEDAN
Disetujui Oleh:
(Dr. Ir. Lollie Agustina. P. Putri, MSi) Anggota Komisi Pembimbing
NIP : 19670821 199301 2 001
Disetujui Oleh:
(Dra. Sekar Woelan, MP) Komisi Pembimbing Lapangan
ABSTRACT
Character study of physiological and the flow of clones rubber latex (Hevea brasiliensis Muell Arg.) IRR 300 series. The purpose of the experimental was to determine the physiological character and the flow of latex rubber on IRR 300 series clones. The experimental uses Augmented Randomize Block Design with treatment of 24 clones and three replications. The treatment consist of IRR 300, IRR 301, IRR 302, IRR 303, IRR 304, IRR 305, IRR 306, IRR 307, IRR 308, IRR 309, IRR 310 , IRR 311, IRR 313, IRR 314, IRR 315, IRR 316, IRR 317, IRR 318, IRR 319, IRR 321, IRR 323, and BPM 24, PB 260, RRIC 100 clone as a comparators. The data were analyzed using ANOVA. The result showed that treatment of clones of physiological (sucrose, thiol, and inorganic phosphate) in general showed a significant influence with a third clone
comparators. This treatment of the flow of latex clone (production index, plugging index, the flow velocity of latex) showed significant effect with the three clones comparators except at long observation tapping grooves. In the observation of growth (girth and thick skin) generally are not significantly different from the third clone comparators. In the observation of skin anatomy (number of rows of latex vessels and vessel diameter latex is generally not significantly different from the third clone comparators. Treatment clones on the observation of production in general significantly different from the third clone comparators. Treatment clones on dry rubber content of the observations are not significantly different from the three clones comparators.
ABSTRAK
Studi karakter fisiologis dan sifat aliran lateks klon karet (Hevea brasiliensis Muell Arg.) IRR seri 300. Tujuan dari penelitian ini
adalah untuk mengetahui karakter fisiologis dan sifat aliran lateks pada klon karet IRR seri 300. Penelitian ini menggunakan Rancangan Acak Kelompok Augmented dengan 24 perlakuan klon dan tiga ulangan. Perlakuan klon terdiri IRR 300, IRR 301, IRR 302, IRR 303, IRR 304, IRR 305, IRR 306, IRR 307, IRR 308, IRR 309, IRR 310, IRR 311, IRR 313, IRR 314, IRR 315, IRR 316, IRR 317, IRR 318, IRR 319, IRR 321, IRR 323, dan BPM 24, PB 260, RRIC 100 sebagai klon pembanding. Data yang diperoleh dianalisis dengan menggunakan ANOVA. Hasil analisis menunjukkan bahwa perlakuan klon terhadap sifat fisiologis (sukrosa, tiol, dan fosfat anorganik) secara umum menunjukkan pengaruh yang sangat nyata dengan ketiga klon pembanding. Perlakuan klon terhadap sifat aliran lateks (indeks produksi, indeks penyumbatan, kecepatan aliran lateks) menunjukkan pengaruh yang nyata dengan ketiga klon pembanding kecuali pada pengamatan panjang alur sadap. Pada pengamatan pertumbuhan (lilit batang dan tebal kulit) secara umum tidak berbeda nyata dengan ketiga klon pembanding. Pada pengamatan anatomi kulit (jumlah baris pembuluh lateks dan diameter pembuluh lateks secara umum tidak berbeda nyata dengan ketiga klon pembanding. Perlakuan klon pada pengamatan produksi secara umum berbeda nyata dengan ketiga klon pembanding. Perlakuan klon pada pengamatan kadar karet kering tidak berbeda nyata dengan ketiga klon pembanding.
DAFTAR ISI
Pengambilan Sampel Kulit ... 18
Anatomi Kulit ... 18
Pengukuran Kadar Sukrosa ... 19
Pengukuran Kadar Tiol ... 20
Pengukuran Kadar Fosfat Anorganik ... 20
Pengamatan Parameter ... 20
Sifat Fisiologi ... 20
Sukrosa (mM) ... 20
Tiol (mM) ... 21
Fosfat Anorganik (mM) ... 21
Aliran Lateks ... 21
Indeks Produksi ... 21
Indeks Penyumbatan ... 21
Panjang Alur Sadap ... 21
Kecepatan Aliran Lateks ... 21
Pertumbuhan ... 22
Lilit Batang (cm) ... 22
Tebal Kulit (mm)... 22
Anatomi Kulit ... 22
Jumlah Baris Pembuluh Lateks (buah) ... 22
Diameter Pembuluh Lateks (mikron) ... 22
Produksi (g/p/s) ... 23
Kadar Karet Kering (%) ... 23
HASIL DAN PEMBAHASAN Hasil ... ... 24
Pembahasan ... ... 24
KESIMPULAN DAN SARAN Kesimpulan ... ... 45
Saran ... ... 46
DAFTAR PUSTAKA
DAFTAR TABEL
1. ... K ondisi Tanaman ... 17 2. ... H
asil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
Sukrosa (mM) ... 24 3. Hasil Rataan Yang Telah Dikoreksi Dan Yang Belum Pada Parameter
Tiol (mM) ... 26 4. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
Fosfat Anorganik (mM) ... 27 5. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
Indeks Produksi ... 29 6. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
Indeks Penyumbatan ... 30 7. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
Panjang Alur Sadap ... 32 8. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
Kecepatan Aliran Lateks ... 34 9. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
Lilit Batang (cm) ... 35 10. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
Tebal Kulit (mm) ... 37 11. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
Jumlah Baris Pembuluh Lateks (buah) ... 38 12. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
Diameter Pembuluh Lateks (micron) ... 40 13. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
Produksi (g/p/s) ... 41 14. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter
DAFTAR LAMPIRAN
1. Bagan Penelitian ... 49
2. Jadwal Kegiatan Penelitian ... 50
3. Data Pengamatan Sukrosa (mM) ... 51
4. Hasil Rataan Klon Pembanding Pada Pengamatan Sukrosa (mM) ... 51
5. Analisis Ragam Sukrosa (mM) ... 52
6. Data Pengamatan Tiol (mM) ... 53
7. Hasil Rataan Klon Pembanding Pada Pengamatan Tiol (mM) ... 53
8. Analisis Ragam Tiol (mM) ... 54
9. Data Pengamatan Fosfat (mM) ... 55
10. Hasil Rataan Klon Pembanding Pada Pengamatan Fosfat (mM) ... 55
11. Analisis Ragam Fosfat (mM) ... 56
12. Data Pengamatan Indeks Produksi ... 57
13. Hasil Rataan Klon Pembanding Pada Pengamatan Indeks Produksi ... 57
14. Analisis Ragam Indeks Produksi ... 58
15. Data Pengamatan Indeks Penyumbatan ... 59
16. Hasil Rataan Klon Pembanding Pada Pengamatan Indeks Penyumbatan .... 59
17. Analisis Ragam Indeks Penyumbatan ... 60
18. Data Pengamatan Panjang Alur Sadap ... 61
19. Hasil Rataan Klon Pembanding Pada Pengamatan Panjang Alur Sadap ... 61
20. Analisis Ragam Panjang Alur Sadap ... 62
21. Data Pengamatan Kecepatan Aliran Lateks ... 63
22. Hasil Rataan Klon Pembanding Pada Pengamatan Kecepatan Aliran Lateks ... 63
23. Analisis Ragam Kecepatan Aliran Lateks ... 64
24. Data Pengamatan Lilit Batang (cm) ... 65
25. Hasil Rataan Klon Pembanding Pada Pengamatan Lilit Batang (cm) ... 65
26. Analisis Ragam Lilit Batang (cm) ... 66
27. Data Pengamatan Tebal Kulit (mm) ... 67
28. Hasil Rataan Klon Pembanding Pada Pengamatan Tebal Kulit (mm) ... 67
29. Analisis Ragam Tebal Kulit (mm) ... 68
30. Data Pengamatan Jumlah Baris Pembuluh Lateks (buah) ... 69
31. Hasil Rataan Klon Pembanding Pada Pengamatan Jumlah Baris Pembuluh Lateks (buah) ... 69
32. Analisis Ragam Jumlah Baris Pembuluh Lateks (buah) ... 70
33. Data Pengamaatan Diameter Pembuluh Lateks (mikron) ... 71
34. Hasil Rataan Klon Pembanding Pada Pengamatan Diameter Pembuluh Lateks (mikron) ... 71
35. Analisis Ragam Diameter Pembuluh Lateks (mikron) ... 72
36. Data Pengamatan Produksi (g/p/s) ... 73
37. Hasil Rataan Klon Pembanding Pada Pengamatan Produksi (g/p/s) ... 73
38. Analisis Ragam Produksi (g/p/s) ... 74
39. Data Pengamatan Kadar Karet Kering (%) ... 75
ABSTRACT
Character study of physiological and the flow of clones rubber latex (Hevea brasiliensis Muell Arg.) IRR 300 series. The purpose of the experimental was to determine the physiological character and the flow of latex rubber on IRR 300 series clones. The experimental uses Augmented Randomize Block Design with treatment of 24 clones and three replications. The treatment consist of IRR 300, IRR 301, IRR 302, IRR 303, IRR 304, IRR 305, IRR 306, IRR 307, IRR 308, IRR 309, IRR 310 , IRR 311, IRR 313, IRR 314, IRR 315, IRR 316, IRR 317, IRR 318, IRR 319, IRR 321, IRR 323, and BPM 24, PB 260, RRIC 100 clone as a comparators. The data were analyzed using ANOVA. The result showed that treatment of clones of physiological (sucrose, thiol, and inorganic phosphate) in general showed a significant influence with a third clone
comparators. This treatment of the flow of latex clone (production index, plugging index, the flow velocity of latex) showed significant effect with the three clones comparators except at long observation tapping grooves. In the observation of growth (girth and thick skin) generally are not significantly different from the third clone comparators. In the observation of skin anatomy (number of rows of latex vessels and vessel diameter latex is generally not significantly different from the third clone comparators. Treatment clones on the observation of production in general significantly different from the third clone comparators. Treatment clones on dry rubber content of the observations are not significantly different from the three clones comparators.
ABSTRAK
Studi karakter fisiologis dan sifat aliran lateks klon karet (Hevea brasiliensis Muell Arg.) IRR seri 300. Tujuan dari penelitian ini
adalah untuk mengetahui karakter fisiologis dan sifat aliran lateks pada klon karet IRR seri 300. Penelitian ini menggunakan Rancangan Acak Kelompok Augmented dengan 24 perlakuan klon dan tiga ulangan. Perlakuan klon terdiri IRR 300, IRR 301, IRR 302, IRR 303, IRR 304, IRR 305, IRR 306, IRR 307, IRR 308, IRR 309, IRR 310, IRR 311, IRR 313, IRR 314, IRR 315, IRR 316, IRR 317, IRR 318, IRR 319, IRR 321, IRR 323, dan BPM 24, PB 260, RRIC 100 sebagai klon pembanding. Data yang diperoleh dianalisis dengan menggunakan ANOVA. Hasil analisis menunjukkan bahwa perlakuan klon terhadap sifat fisiologis (sukrosa, tiol, dan fosfat anorganik) secara umum menunjukkan pengaruh yang sangat nyata dengan ketiga klon pembanding. Perlakuan klon terhadap sifat aliran lateks (indeks produksi, indeks penyumbatan, kecepatan aliran lateks) menunjukkan pengaruh yang nyata dengan ketiga klon pembanding kecuali pada pengamatan panjang alur sadap. Pada pengamatan pertumbuhan (lilit batang dan tebal kulit) secara umum tidak berbeda nyata dengan ketiga klon pembanding. Pada pengamatan anatomi kulit (jumlah baris pembuluh lateks dan diameter pembuluh lateks secara umum tidak berbeda nyata dengan ketiga klon pembanding. Perlakuan klon pada pengamatan produksi secara umum berbeda nyata dengan ketiga klon pembanding. Perlakuan klon pada pengamatan kadar karet kering tidak berbeda nyata dengan ketiga klon pembanding.
PENDAHULUAN
Latar Belakang
Perekonomian di Indonesia salah satunya dihasilkan dari pengembangan
kebun karet. Fungsi dari perkebunan karet tidak hanya sebagai sumber devisa,
sumber bahan baku industri, sumber pendapatan dan kesejahteraan masyarakat
tetapi sekaligus berperan dalam pelestarian fungsi lingkungan hidup.
Selama tiga dekade ini pengembangan karet di Indonesia mengalami
pertumbuhan yang sangat pesat. Di awal tahun 1968, luas areal karet baru 2,2 juta
ha dan pada tahun 2005 meningkat menjadi 3,2 juta ha atau meningkat menjadi
sekitar 50%. Hampir 85% pengusahaan karet diusahakan oleh rakyat dan sisanya
oleh perkebunan besar. Dari luasan tersebut, produksi yang dihasilkan mencapai
sebesar 2,2 juta ton dengan produktifitas rata-rata sebesar 840 kg/ha/tahun
(Ditjenbun, 2007).
Pengamatan produksi dan pertumbuhan selama umur ekonomis yang saat
ini diterapkan menyebabkan daur pemuliaan karet menjadi sangat panjang yakni
30 tahun per siklus. Masalah lamanya daur seleksi merupakan hambatan utama
terhadap kemajuan hasil pemuliaan karet. Kemajuan pemuliaan dalam 4 siklus
yang telah dilalui di Indonesia (90 tahun) telah berhasil meningkatkan
produktifitas tanaman karet dari 500 kg menjadi 3000 kg/ ha/ tahun
(Azwar, 1993).
Dalam sejarah perkembangan karet alam, pada mulanya perhatian hanya
ditujukan untuk menghasilkan karet sebesar-besarnya. Setelah penggunaan lateks
menyangkut komposisi, sifat dan mekanisme aliran lateks
(Gills dan Suharto, 1976).
Beberapa peneliti mencoba untuk memanfaatkan teknologi baru seperti
pengujian plot promosi untuk memperpendek siklus pemuliaan tanaman karet
(Tan, 1987). Upaya memperpendek siklus seleksi tanaman karet terus dilakukan
yaitu dengan mencari beberapa komponen produksi yang berkaitan dengan
produksi lateks. Menurut Narayanan, et al., (1973) bahwa pembuluh lateks, tebal
kulit batang dan lingkar batang saling berhubungan dan mempunyai peranan yang
besar terhadap pendugaaan produksi.
Kemajuan bidang fisiologis dan biokimia membentuk lateks oleh tanaman
karet pada saat itu telah memungkinkan menentukan karakteristik tanaman muda
yang bakal memilki produktifitas tinggi (Jacob et al., 1989). Apabila parameter
fisiologi dan biokimia itu dirumuskan secara tepat dan dilaksanakan dengan
akurat, maka profil dari klon dapat digambarkan pada potensi produksinya
diketahui secara dini (Bricard dan Nicolas, 1989).
Belakangan ini peneliti menemukan peluang untuk membedakan
karakteristik fisiologi lateks dari jenis klon yang berbeda secara mantap tetap
banyak dipengaruhi oleh faktor lingkungan dan umur tanaman (Jacob et al, 1989).
Klon IRR (Indonesian Rubber Research) seri 300 merupakan hasil seleksi
Pusat Penelitian Karet, khususnya Balai Penelitian Sungei Putih. Perakitan klon
tersebut dilakukan pada tahun 1991 di Kebun Persilangan Balai Penelitian Sungei
Putih dengan pohon yang diperpendek. Jumlah persilangan yang dilakukan untuk
menghasilkan klon IRR seri 300 sebanyak 25 388 bunga betina dan sebanyak 397
dan produksi. Genotipe-genotipe yang terseleksi tersebut akan digunakan sebagai
materi genetik klon IRR seri 300 (Azwar et al, 1998).
Berdasarkan uraian diatas maka diperlukan suatu pengkajian terhadap
karakter fisiologis dan sifat aliran klon karet pada IRR seri 300. Pada penelitian
ini digunakan 24 genotipe.
Tujuan Penelitian
Untuk mengetahui karakter fisiologis dan sifat aliran lateks pada klon
karet IRR seri 300.
Hipotesis
Ada variasi karakter fisiologis dan aliran lateks diantara klon karet
IRR seri 300.
Kegunaan Penelitian
1. Sebagai salah satu syarat untuk memperoleh gelar sarjana
di Fakultas Pertanian Universitas Sumatera Utara, Medan
TINJAUAN PUSTAKA
Botani Tanaman
Menurut (Setiawan dan Andoko, 2005) dalam taksonomi tumbuhan,
tanaman karet termasuk dalam kelas dicotyledonae, ordo euphorbiales, famili
euphorbiaceae, genus hevea dan spesies Hevea brasiliensis.
Syarat Tumbuh
Iklim
Tanaman karet adalah tanaman daerah tropis daerah tropis yang ditanami
karet yakni terletak pada 150 LU-100 LS, dengan suhu harian yang diinginkan
rata-rata 250 C-300
Tanaman karet tumbuh dengan baik di daerah tropis. Daerah yang cocok
untuk tanaman karet adalah pada zone antara 15° LS dan 15° LU. Bila ditanam di
luar zone tersebut, pertumbuhannya agak lambat, sehingga memulai produksinya
pun lebih lambat. Tanaman karet tumbuh optimal di dataran rendah, yakni pada
ketinggian sampai 200 meter di atas permukaan laut. Makin tinggi letak tempat
pertumbuhannya makin lambat dan hasilnya lebih rendah. Ketinggian lebih dari
600 meter dari permukaan laut tidak cocok lagi untuk tanaman karet
C (Nazaruddin dan Paimin, 1992).
Secara garis besar tanaman karet dapat tumbuh baik pada kondisi iklim
sebagai berikut : suhu rata-rata harian 28° C (dengan kisaran 25-35o C) dan curah
hujan tahunan rata-rata antara 2.500 – 4.000 mm dengan hari hujan mencapai 150
mempengaruhi kegiatan penyadapan. Daerah yang sering mengalami hujan pada
pagi hari produksinya akan kurang. Keadaan daerah di Indonesia yang cocok
untuk pertanaman karet adalah daerah-daerah Indonesia bagian barat, yaitu
Sumatera, Jawa dan Kalimantan, sebab iklimnya lebih basah
Tanah
Tanaman karet tidak terlalu menuntut kesuburan tanah yang tinggi, bisa
saja ditanami di lahan yang kurang subur dibandingkan dengan tanaman
perkebunan lainnya (kopi, kakao, teh, tembakau), tanaman karet adalah tanaman
yang paling toleran terhadap tanah yang kesuburannya rendah. Untuk membantu
pertumbuhan dapat dilakukan dengan penambahan pupuk
(Nazaruddin dan Paimin, 1992).
Berbagai jenis tanah mempunyai sifat yang berbeda baik dalam sifat fisik
maupun kesuburan kimiawi dan keadaan lingkungannya. Budidaya tanaman karet
mengandalkan perawatan tertentu untuk pengusahaan secara komersil selain dari
iklim juga bentuk wilayah/ fisiografi keadaan tanah dan lingkungannya
(Rasjidin, 1989).
Tanah yang pH nya mendekati normal cocok untuk ditanami karet. Derajat
keasaman yang paling cocok adalah 5-6. Batas optimum toleransi pH tanah bagi
tanaman karet adalah 4-8. Tanah yang agak masam masih lebih baik dari pada
Lateks dan Pembentukannya
Lateks adalah cairan putih dari pohon karet yang diambil dari tanaman
pada proses penyadapan. Lateks berguna bagi tanaman sebagai bahan pengawet
(preservative). Lateks dibentuk dalam pembuluh lateks. Pembuluh ini terdiri dari
2 macam. Pertama pembuluh lateks yang berasal dari 1 sel yang kemudian
bercabang-cabang membentuk suatu pembuluh seperti amuba. Pembuluh lateks
seperti ini disebut pembuluh lateks simple, misalnya terdapat pada biji. Kedua
pembuluh lateks yang berasal dari deretan sel-sel dimana dinding-dinding sel
kearah tegak lurus masing-masing melebur membentuk suatu pembuluh.
Pembuluh lateks ini disebut pembuluh kompoun dan inilah yang terdapat pada
tanaman karet yaitu pada kulit lunak dan kulit keras (Lukman, 1984).
Pembuluh Lateks
Pembuluh lateks mengandung pembuluh dengan dinding yang permanen
dan elastis. Sebelum melakukan penyadapan tekanan didalam pembuluh lateks
tinggi. Pengaliran lateks disebabkan karena tekanan dalam pembuluh serta
pergerakan cairan lateks akibat perbedaan konsentrasi setelah pohon disadap.
Pada mikroskop elektron dapat dilihat partikel lateks yang rusak akan
mengeluarkan lateks (Southorn, 1961).
Jika penampang melintang tanaman karet dipelajari, bagian tengah
terdapat jaringan kayu (xylem) yang dilapisi oleh kambium. Pada bagian luar
dijumpai kulit lunak yang menyusul kulit keras pada kulit luar sel gabus sebagai
lapisan terakhir. Di dalam kulit lunak tersebut terdapat sederetan pembuluh tapis
Menurut Southorn (1961), lateks merupakan suatu sistem pembuluh
berupa pipa saluran di dalam jaringan floem yang halus dari karet. Pembuluh ini
berada dekat dengan kambium, pertama-tama membentuk sel tunggal lalu
membentuk suatu jaringan pembuluh melalui anatomisis. Gills dan Suharto (1976)
menyatakan bahwa semakin dekat dengan kambium maka aliran pembuluh
semakin kecil dengan ukuran 30 mikron.
Baik ketebalan asli maupun jumlah baris pembuluh lateks yang ada di
dalam semakin meningkat dan bertambahnya usia tanaman. Jumlah baris
pembuluh lateks pada prinsipnya merupakan cirri khas suatu klon tetapi
perkembangannya tergantung pada tingkat pertumbuhan tanaman yang
dipengaruhi oleh faktor-faktor seperti kepadatan tanaman dan status hara dan juga
oleh klon (Webster dan Baulkwill, 1989).
Struktur Lateks
Lateks merupakan suatu sistem koloid yang bermuatan negatif berupa
serum yang berisi protein anionik yang membentuk suatu badan yang dikelilingi
oleh membran (lutoid) yang merupakan suatu sistem koloid kedua yang
mengandung asam yang kebanyakan cation serum (Southorn dan Yip, 1968).
Menurut Subronto dan Napitupulu (1978), menayatakan bahwa lateks
mengalir karena adanya proses pengenceran sebelum disadap tugor tanaman
adalah tinggi akan tetapi setelah disadap menjadi penurunan tugor terutama dalam
sel pembuluh lateks. Semakin tinggi tugor antara sel sekitar pembuluh maka
proses pengenceran semakin lama.
Dijkman (1951), melaporkan bahwa lateks yang keluar dari organ muda
kulit batang tanaman yang berumur 5-10 tahun, tetapi proses penggumpalan lateks
lebih lama terjadi pada lateks yang keluar dari organ muda, sebab partikel dari
organ ini sangat sedikit dan viskositas lateksnya lebih rendah.
Aliran Lateks
Pembuluh lateks adalah sel-sel hidup yang mengandung larutan seperti
gula, protein dan garam mineral yang dapat menyimpan air dari jaringan yang
berada disekitarnya. Ketika tanaman karet disadap lateks berhenti beberapa saat.
Adapun faktor yang berhubungan dengan aliran lateks, yaitu :
Fisiologi Aliran Lateks
Sifat-sifat fisiologi aliran lateks antara lain dicirikan oleh indeks
penyumbatan, kecepatan aliran lateks, indeks produksi, kadar karet kering, total
solud konten serta anatomi kulit yang meliputi jumlah, diameter dan kerapatan
pembuluh lateks (Rasjidin, 1989).
Proses Pengaliran Lateks
Apabila suatu alur sadap dibuka maka keluarlah lateks oleh tekanan dari
dalam. Pengurangan terjadi secara berlanjutan sepanjang pembuluh lateks
sehingga mengalirnya lateks menuju bagian yang dipotong. Pada saat yang sama
akibat menurunnya tekanan dalam sel pembuluh lateks maka mengalirlah air ke
dalam pembuluh dari sel sekelilingnya sehingga mengencerkan lateks
(Rasjidin, 1989).
Daerah Aliran Lateks
Penelitian fisiologi tentang luasnya daerah pengaliran lateks yang secara
efektif turut serta mengalirkan lateks selama penyadapan dilakukan oleh Frey
daerah aliran lateks hampir seluruhnya terdapat dibawah alur sadap hanya
sebagian kecil dari samping alur sadap, luasnya tergantung kapasitas produksi
pohon yang berproduksi tinggi daerah pengaliran pengaliran vertikal mencapai
171 cm (Rasjidin, 1989).
Indeks Penyumbatan
Indeks penyumbatan dan panjang alur sadap sewaktu penyadapan juga
menentukan pola aliran lateks. Semakin panjang alur sadapan, indeks
penyumbatan semakin kecil sehingga lateks yang mengalir lebih lama. Sebaliknya
semakin pendek alur sadap, indeks penyumbatan semakin besar. Sebab utama
terjadinya penyumbatan pembuluh lateks adalah pecahnya butir lutoid yang
terdapat dalam lateks akibat gesekan yang terjadi ketika lateks mengalir.
Terjadinya penyempitan pada pembuluh lateks kemungkinan dapat mengganggu
aliran lateks sehingga menyebabkan pola aliran lateks untuk setiap klon berbeda
(Boerhendy, 1988).
Indeks penyumbatan merupakan sifat khas yang tidak dipengaruhi oleh
umur tanaman, tetapi sedikit dipengaruhi oleh faktor lingkungan akibat terjadinya
variasi produksi antara pohon dan variasi harian (Subronto dan Napitupulu, 1978).
Kecepatan Aliran Lateks
Pengamatan kecepatan aliran lateks dimaksudkan untuk mengetahui pola
aliran lateks. Pada awalnya aliran lateks mengalir cepat, kemudian lambat dan
akhirnya berhenti. Lambat cepatnya aliran lateks sewaktu disadap berpengaruh
terhadap tinggi rendahnya produksi. Semakin cepat dan lama lateks mengalir,
maka hasil lateksnya semakin tinggi. Dari hasil penelitian yang telah dilakukan,
memungkinkan disebabkan oleh banyaknya pembuluh lateks yang terpotong.
Selain itu, komposisi pembuluh lateks juga berbeda. Berdasarkan hasil itu maka
pola aliran lateks berbeda untuk setiap klon sehingga hasil juga berbeda
(Boerhendy, 1988).
Subronto dan Harris (1977), menyatakan bahwa kecepatan aliran akan
menggambarkan aliran lateks per satuan waktu per panjang alur sadap yang
dilalui. Kecepatan aliran lateks berkorelasi positif dengan produksi.
Indeks Produksi
Indeks produksi merupakan suatu perbandingan antara produksi dengan
lilit batang yang menggambarkan kemampuan berproduksi tanaman. Indeks ini
juga menggambarkan produksi kulit. Indeks produksi dipengaruhi faktor anatomis
dan fisiologis tanaman. Oleh sebab itu, indeks produksi nilainya dipengaruhi oleh
umur tanaman (Subronto dan Napitulu, 1979).
Subronto dan Napitupulu (1978) menyatakan, indeks produksi dari
tanaman yang distimulan umumnya lebih besar dari pada tanaman yang tidak
distimulan, tetapi cenderung menurun pada tahun berikutnya. Hal ini terjadi
karena produksi tanaman distimulan tinggi sedangkan lilit batangnya relatif kecil.
Sebelumnya Napitupulu (1977) menjelaskan, bahwa rata-rata indeks produksi
lebih tinggi dihasilkan oleh intensitas penyadapan ½ S d/3.
Kadar Karet Kering
Kadar karet kering cenderung lebih tinggi pada tanaman yang memiliki
lilit batang yang kecil dibanding dengan tanaman yang memiliki lilit batang lebih
Kadar karet kering yang tinggi terutama disebabkan oleh viskositas lateks
yang tinggi, yang menyebabkan proses penyumbatan berjalan lebih cepat dan
lateks yang dihasilkan menurun (Subronto dan Harris, 1977).
Lilit Batang
Pendugaan produksi pohon karet dapat dilakukan dengan mengukur
besarnya lilit batang dan tebal kulit yang dipakai untuk mengetahui kemampuan
produksi maksimum untuk menghasilkan lateks sebanyak mungkin. Maka besar
lilit batang dan tebal kulitnya diharapkan produksinya semakin tinggi.
Pertumbuhan lilit batang tiap tahun sebelum penyadapan berkisar antara
6,52–10,44 cm dengan nilai rata-rata 9,08/ tahun. Pertambahan lilit batang
sesudah tanaman disadap berkisar 1,82-6,64 cm/ tahun dengan nilai rata-rata
3,04 cm/ tahun (Danimihardja, 1988).
Tebal Kulit
Pertumbuhan tebal kulit merupakan karakteristik pada klon tertentu,
namun tebalnya kulit dapat terpengaruh oleh faktor lingkungan. Dalam seleksi
tebal kulit dinilai dengan memperbandingkan dengan tebal kulit klon. Pada
umumnya kulit yang tipis karena kemungkinan terjadinya luka ketika penyadapan
lebih kecil (Lukman, 1983).
Anatomi Kulit
Kulit perawan atau asli dapat dibedakan 3 lapisan konsentrasi yaitu lunak
yang paling dekat dengan kambium terdiri dari silinder-silinder laticiper yang
lebih tipis. Lapisan kedua adalah kulit keras juga mengandung tabung-tabung
pembuluh floem dan laticifer tetapi keduanya tidak teratur dan tidak berfungsi
periderm terdiri dari penutup luar sel gabus (fellogen) yang membentuk sel-sel
gabus pada sisi bagian luar dan feloderm yaitu suatu jaringan yang mirip dengan
perenchym korteks pada sisi bagian dalam (Webster dan Baulkwill, 1989).
Sepotong kulit bagian dalamnya yang dekat dengan kambium adalah
floem, blast merupakan kulit lunak yang utamanya terdiri dari baris-baris sel yang
hampir vertikal dengan dinding-dinding melintang berperforasi (tabung-tabung
ayakan) yang mengantarkan bahan-bahan makanan, sel-sel bulat yang lebih kecil
(parenchym) yang tersusun terutama berkaitan dengan simpanan bahan makanan,
dan baris-baris pembuluh lateks yang merupakan sel-sel penunjang hampir
vertikal dimana dinding melintang tak beraturan (Edgar, 1958).
Kulit bentukan baru lebih tipis dari kulit semula, disebabkan berkurangnya
lapisan-lapisan gabus kulit dan tidak adanya sel-sel batu. Oleh karena itu sebagian
lebih besar dari pembuluh-pembulu lateks fungsional pada kulit bentukan baru
dan ini bertanggung jawab terhadap hasil yang lebih tinggi yang ada kalanya
dapat diperoleh dari kulit bentukan baru. Sebagian kasus, ini dapat mengakibatkan
pengeringan prematur bila bentukan baru disadap, disebabkan menipisnya
BAHAN DAN METODE PENELITIAN
Tempat dan Waktu Penelitian
Penelitian ini dilaksanakan di kebun Percobaan Pusat Penelitian Karet
Sungai Putih, Kecamatan Galang, Kabupaten Deli Serdang dengan ketinggian
54 m dpl. Penelitian ini akan dilaksanakan pada bulan Maret 2011 sampai dengan
bulan Juni 2011.
Bahan dan Alat
Bahan yang digunakan dalam penelitian ini adalah tanaman klon karet
IRR seri 300 umur 11 tahun terdiri dari 24 klon, larutan FAA (campuran dari 10
ml Formalin 37-40% + 5 ml Acetic Acid + 70 ml Ethanol absolut dan 15 ml
aquadest), KOH 15%, HNO3
Alat yang digunakan dalam penelitian ini adalah cork borrer, magnetic
stirrer, pisau silet, object glass, cover glass, mikroskop, mikro pipet, pisau sadap,
ember plastik, gelas ukur, timbangan Metler, alat tulis dan alat-alat lainnya yang
dapat membantu penelitian ini.
32,5 %, Alkohol 70%, Sudan III, Acetol, Glyserin,
H2SO4, TCA 2,5%, larutan standar Pi (KH2PO4 10 mM), Ammoniak
heptamolibdat 10 g, DTNB 79,3 g, EDTA 148,8 g, Tris 05 ml, Glutation reduce
Metode Penelitian
Penelitian ini menggunakan Rancangan Acak Kelompok (RAK)
Augmented dengan 24 perlakuan klon dan 3 ulangan.
Perlakuan terdiri dari klon-klon sebagai berikut :
K1 = IRR 300 K6 = IRR 305 K11 = IRR 310 K16 = IRR 316
K2 = IRR 301 K7 = IRR 306 K12 = IRR 311 K17 = IRR 317
K3 = IRR 302 K8 = IRR 307 K13 = IRR 313 K18 = IRR 318
K4 = IRR 303 K9 = IRR 308 K14 = IRR 314 K19 = IRR 319
K5 = IRR 304 K10 = IRR 309 K15 = IRR 315 K20 = IRR 321
K21 = IRR 323 K22 = RRIC 100 (Klon Pembanding)
K23 = PB 260 (Klon Pembanding) K24 = BPM 24 (Klon Pembanding)
Jumlah ulangan : 3 ulangan (5 tanaman/ ulangan)
Jumlah plot : 24 plot
Jarak tanam : 5 x 4 meter
Luas Percobaan : ± 1,5 ha
Jumlah tanaman/plot : 15 tanaman ( ½ S d/3)
Jumlah sampel/plot : 15 tanaman
Jumlah seluruh tanaman : 360 tanaman.
Analisis Data
Data hasil penelitian di analisis dengan sidik ragam dengan model linear
aditif sebagai berikut:
Yij = µ + ρi+ τj + ε
i=1,2,3 j=1,2,3,. . .,24
Dimana:
Yij
µ = Efek nilai tengah
= Hasil pengamatan blok ke- i dan perlakuan ke- j
ρi
τ
= Efek dari blok ke- i
j ε
= Efek dari perlakuan klon pembanding pada taraf ke- j
ij
Nilai rataan hasil di koreksi dari pengaruh blok (rj) dengan rumus : = Efek error pada blok ke- i dan taraf ke-j.
rj = Bj
Dimana :
– M
Bj
M = rataan umum klon pembanding
= rataan semua varietas check dalam blok ke-j (klon pembanding)
Dengan syarat Σrj
Untuk membandingkan nilai rata-rata hasil digunakan standard error yang
diperoleh dari analisis sidik ragam klon pembanding : = 0
SY KTG(b+1)(c+1)/bc
Dimana : b = jumlah blok
c = jumlah klon pembanding (Sutjihno, 2002).
Untuk melihat hubungan antar parameter yang diamati, maka dilakukan
pengujian berdasarkan analisa korelasi sebagai berikut :
n xiyi – (Σxi) (Σyi)/n
√(Σx2
– (Σx)2(Σy2 – (Σy)2 n n
PELAKSANAAN PENELITIAN
Persiapan Areal
Penelitian ini merupakan penelitian lanjut dari penelitian sebelumnya.
Oleh karena itu areal yang digunakan adalah areal yang telah ada sebelumnya.
Populasi yang digunakan tanaman okulasi hasil dari IRR seri 300 di Pusat
Penelitian Karet Sungei Putih.
Sensus Tanaman
Untuk mengetahui jumlah populasi yang ada, maka dilakukan sensus
tanaman yang meliputi jumlah tanaman yang ada dan yang diamati, tanaman
kerdil dan terserang penyakit.
Penentuan Batas Plot
Penentuan batas plot dilakukan sebelum melaksanakan penelitian.
Tujuannya agar lebih memudahkan dalam masing-masing plot. Batas antara satu
plot dengan plot yang lain diberi tanda nomor plot.
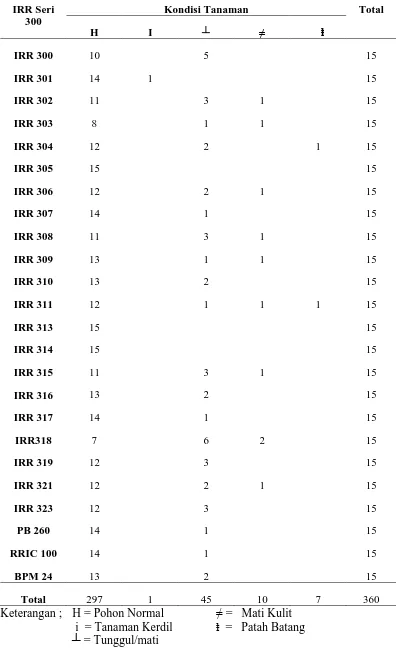
Kondisi Tanaman
Pengamatan kondisi tanaman dilakukan melalui sensus tanaman, yaitu
dengan mengamati keadaan tanaman dilapangan satu persatu dalam setiap progeni
untuk mengetahui keadaan tanaman yang tumbuh normal, kerdil, tunggul, patah
batang dan titik tanam tinggal lubang.
Untuk mengetahui keadaan tanaman yang hidup normal dapat dilihat pada
Tabel 1. Diatas menjelaskan bahwa tidak semua tanaman yang ada
dilapangan tumbuh normal. Jumlah keseluruhan tanaman yang normal adalah
297 tanaman (82,5%). Yang terbanyak berasal dari IRR 314, IRR 313, IRR 305
(4,2%), dan yang paling sedikit berasal dari IRR 318 (1,9%). Sedangkan tanaman
yang tidak normal seperti kerdil, mati kulit, tunggul dan patah batang jumlahnya
63 tanaman (17,5%).
Pengambilan Sampel Kulit
Pengambilan sampel kulit dilakukan dari 3 tanaman per plot. Sampel kulit
yang diambil sebanyak 3 buah per plot pada ketinggian 150 cm dari pertautan
okulasi dengan menggunakan cork borrer.
Pelaksanaan di Laboratorium
Anatomi Kulit
a. Sampel kulit yang diambil, langsung dimasukkan ke dalam larutan tambahan
FAA (Formalin Acetic Acid) yang telah disiapkan. Larutan FAA adalah
campuran dari 10 ml Formalin 37-40% + 5 ml Acetic Acid + 70 ml Ethanol
absolut dan 15 ml akuades.
b. Contoh kulit yang telah difiksasi tersebut selanjutnya dibawa ke laboratorium
dan dipindahkan ke dalam larutan KOH 15% selama 1 jam. Larutan KOH
berguna untuk mematikan sel-sel gabus atau melarutkan lateks yang masih
melekat pada kulit.
c. Dari dalam larutan KOH 15 % contoh kulit dipindahkan lagi kedalam larutan
d. Selanjutnya dipindahkan ke dalam larutan alkohol 70% selama 15 menit untuk
membersihkan sisa-sisa larutan HNO3
e. Agar preparat yang dihasilkan baik dan jelas, contoh kulit yang direndam
dalam larutan pewarna yang terdiri dari 0,5 g Sudan III + 50 ml Alkohol 70%
+ 50 ml Acetol. Campuran zat pewarna ini diaduk merata dengan alat
Magnetic Stirer, lalu disaring.
yang masih tertinggal pada kulit.
Catatan : pemindahan kulit dari larutan ke dalam larutan yang lain terlebih
dahulu harus dibilas di bawah air mengalir selama 5 menit dan dikeringkan
dengan kertas penghisap.
f. Preparat dibuat setelah contoh kulit yang telah diberi perlakuan diiris setipis
mungkin, yakni dengan menyayat kulit secara membujur dan melintang.
Kemudian sayatan tersebut diletakkan pada objec glass yang sebelumnya telah
ditetesi glyserin dan ditutup dengan cover glass. Selanjutnya pengamatan
dilakukan dengan menggunakan mikroskop.
Pengukuran Kadar Sukrosa
a. Sampel diambil ± 150 µL dan ditambah TCA 2,5 % hingga volume total
500 µL.
b. Ditambah peraksi Anthrone 3 ml (Anthrone 0,1 g ditambahkan ke dalam
larutan sulfat 100 ml) dan divortex.
c. Dipanaskan dengan merendam pada air mendidih selama 15 menit, lalu
didinginkan dengan merendam dalam air.
Pengukuran Kadar Tiol
Larutan Standar Glutation :
Stok I (10 mM) : Glutation reduce 47,9 mg + 15,58 TCA 2,5%.
Stok II (1 mM) : Stok I 1 ml + 9 ml TCA 2,5% (GSH 1 mM).
a. Sampel diambil ± 1,5 ml dan ditambah TCA 2,5 % hingga volume 1,5 ml
kemudian ditambahkan pereaksi DTNB 10 mM 75 µL (DTNB 79,3 g +
EDTA 148,8 g + Tris 0,5 M 5ml dan Akuades 5 ml diaduk da pH ditepatkan
6,5 dengan TCA 2,5% dan volume ditepatkan 20 ml).
b. Ditambah 1,5 ml Bufer Tris 0,5 (30,3 g Tris dilarutkan dalam 500 M Akuades)
dan divortex.
c. Didiamkan pada suhu kamar selama 30 menit.
d. Absorbansi diukur pada λ 412 nm.
Pengukuran Kadar Fosfat Anorganik (Pi)
a. Sampel diambil ± 0,3 ml dan ditambah TCA 2,5% hingga volume 1,5 ml
(faktor pengenceran 1,5 : 0,3 ml).
b. Ditambah 1 ml pereaksi campur (FeSO4 5 g + 50 ml Aquades + larutan stok
Molibdat 10 ml dan ditera hingga 100 ml) dan divortex.
c. Didiamkan 10 menit pada suhu kamar.
d. Kemudian absorbansi diukur pada λ 750 nm.
Pengamatan Parameter
1. Sifat Fisiologi
Sukrosa (mM)
Sukrosa juga dapat diamati dengan jumlah pengamatan sebanyak 1 kali
Tiol (mM)
Tiol dapat diamati dengan jumlah pengamatan sebanyak 1 kali selama
pengamatan dilakukan.
Fosfat Anorganik (mM)
Fosfat anorganik (Pi) juga dapat diamati dengan jumlah pengamatan
sebanyak 1 kali selama pengamatan dilakukan.
2. Aliran Lateks
Indeks Produksi
Indeks produksi dapat dilakukan dan diamati dengan jumlah pengamatan
sebanyak 1 kali dalam 1 bulan. Rumus yang digunakan adalah :
IPr =
x
100Indeks Penyumbatan
Pengamatan indeks penyumbatan dilakukan 1 kali dalam 1 bulan
pengamatan sebelum dilakukan stimulan. Adapun rumus yang digunakan adalah :
IP =
x
100Panjang Alur Sadap
Panjang alur sadap diperoleh dari alat jarum tusuk, dimana jarak jarum
1 dengan lainnya 1 cm. Kalau 10 jarum maka panjang alur sadapnya 10 cm.
Pengamatan ini dilakukan 1 kali selama pengamatan.
Kecepatan Aliran Lateks
Pengamatan kecepatan aliran lateks dilakukan sebanyak 1 kali dalam
KA = x 50
3. Pertumbuhan
Lilit Batang (cm)
Lilit batang diukur 1 kali selama penelitian dan dilakukan pada awal
pengamatan pada ketinggian 50 cm diatas pertautan okulasi.
Tebal Kulit (mm)
Pengamatan tebal kulit dilakukan 1 kali selama pengamatan setelah
dilakukan penyadapan terlebih dahulu.
Anatomi Kulit
Jumlah Baris Pembuluh Lateks (buah)
Perhitungan jumlah baris pembuluh lateks dilakukan dengan cara
pembuatan preparat setipis mungkin secara membujur. Perhitungan dilakukan
dibawah mikroskop dengan pembesaran 4x10.
Diameter Pembuluh Lateks (mikron)
Diameter pembuluh lateks diamati dengan cara pembuatan preparatnya
setipis mungkin secara melintang. Diamati dibawah mikroskop dengan
pembesaran 4x10. Pengukuran dilakukan dengan 2 arah yang saling tegak lurus
kemudian dirata-ratakan dan dikali dengan 2,5 (skala pengukuran lensa okuler
mikroskop).
Produksi adalah produksi lateks yang dihasilkan oleh tanaman pada hari
pengamatan dalam gr/ pohon/ sadap. Pengamatan ini dilakukan 1 kali dalam
3 hari.
Kadar Karet Kering (%)
Kadar karet kering diamati 1 kali dalam 1 bulan. Adapun rumus yang
digunakan adalah :
KKK = x 100 %
HASIL DAN PEMBAHASAN
1. Sifat Fisiologi
Pengamatan Sukrosa (mM)
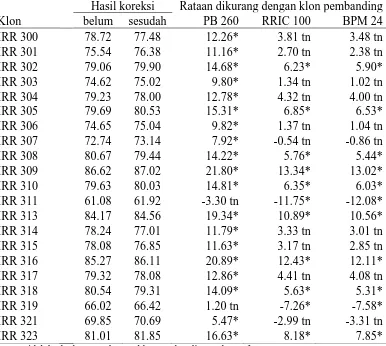
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 2. Dapat diketahui bahwa pada parameter pengamatan sukrosa secara
umum menunjukkan perbedaan yang nyata dengan ketiga klon pembanding. Ada
yang menunjukkan perbedaan ke arah yang lebih tinggi dan berbeda nyata seperti
IRR 309, IRR 307, IRR 319, IRR 323, dan IRR 305 jika dibandingkan dengan
klon PB 260, dan yang menunjukkan perbedaan yang nyata tetapi ke arah yang
lebih rendah terlihat pada klon IRR 315, IRR 308, IRR 313, IRR 311, IRR 316
dan IRR 302. Seperti klon IRR 311, IRR 316 dan IRR 318 tidak berbeda nyata
dengan klon PB 260, klon IRR 300 dan IRR 301 , menunjukkan perbedaan yang
tidak nyata dengan klon pembanding BPM 24.
Adapun klon yang telah di uji memiliki sukrosa paling tinggi adalah
klon IRR 319 (3,17), sedangkan sukrosa yang paling rendah adalah klon IRR 315
(0,60). Nilai rataan klon pembanding dapat dilihat pada Lampiran 4.
Hal ini menjelaskan bahwa adanya variasi yang dimiliki dari klon-klon
IRR seri 300 dan klon pembanding RRIC 100, BPM 24, dan PB 260. Apabila
suatu tanaman memiliki kandungan sukrosa yang tinggi maka lateks yang
dihasilkan semakin banyak atau sebaliknya. Karena sukrosa merupakan bahan
dasar untuk pembentukkan lateks.
Berdasarkan hasil nilai evaluasi sukrosa pada umumnya rendah. Hasil
penelitian Sumarmadji, dkk (2006), menyatakan bahwa kadar sukrosa rendah
(< 5 mM) memberikan indikasi penggunaan bahan asimilat menjadi lateks sangat
intensif atau dapat dikatakan tanaman mengalami lelah fisiologi dengan
kemampuan produksi semakin menurun.
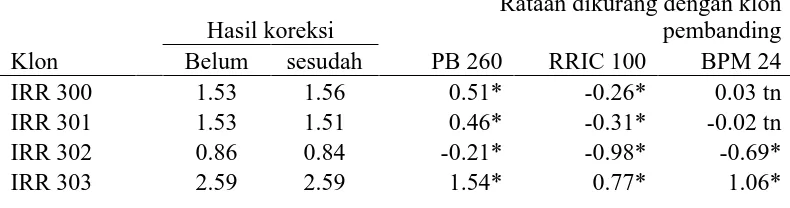
Tabel 2. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Sukrosa (mM)
Klon
Hasil koreksi
IRR 304 2.11 2.14 1.09* 0.32* 0.61* Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
* = berbeda nyata dengan klon pembanding pada taraf 5%
Pengamatan Tiol (mM)
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 3. Dapat diketahui bahwa pada parameter pengamatan tiol secara umum
menunjukkan perbedaan yang nyata dengan ketiga klon pembanding. Ada yang
menunjukkan perbedaan ke arah yang lebih tinggi seperti klon IRR 300, IRR 318,
IRR 306, IRR 309, IRR 307 dan klon lain yang bernilai positif dan yang
menunjukkan perbedaan yang nyata tetapi ke arah yang lebih rendah seperti
terlihat pada klon IRR 317, IRR 315, IRR 303, dan IRR 301. Adapun klon yang
telah di uji memiliki tiol yang paling tinggi adalah klon IRR 323 (0,50),
sedangkan yang paling rendah terdapat pada IRR 303 (0,18). Nilai rataan dari
klon pembanding dapat dilihat pada Lampiran 7. Dari beberapa klon yang diamati
terdapat beberapa klon yang menunjukkan perbedaan yang tidak nyata dengan
dengan klon RRIC 100 dan klon IRR 308, IRR 310, IRR 302 dan IRR 306 tidak
berbeda nyata dengan klon BPM 24 sebagai klon pembanding.
Hal ini menjelaskan bahwa adanya variasi yang dimiliki dari klon-klon
IRR seri 300 dan klon pembanding RRIC 100, BPM 24, dan PB 260. Berdasarkan
hasil nilai evaluasi tiol pada umumnya rendah. Hasil penelitian Sumarmadji, dkk
(2006), menyatakan bahwa kadar sukrosa rendah (< 5 mM) memberikan indikasi
penggunaan bahan asimilat menjadi lateks sangat intensif atau dapat dikatakan
tanaman mengalami lelah fisiologi dengan kemampuan produksi semakin
menurun. Demikian juga ditunjukkan dengan kadar tiol yang umumnya rendah
Tabel 3. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
* = berbeda nyata dengan klon pembanding pada taraf 5%
Pengamatan Fosfat (mM)
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 4. Dapat diketahui bahwa pada parameter pengamatan fosfat secara umum
menunjukkan perbedaan yang nyata dengan ketiga klon pembanding. Ada yang
menunjukkan perbedaan ke arah yang lebih tinggi seperti klon IRR 300 jika
dibandingkan dengan klon PB 260, dan yang menunjukkan perbedaan yang nyata
tetapi ke arah yang lebih rendah terlihat pada seluruh klon kecuali IRR 300.
Adapun klon yang telah di uji yang memiliki fosfat paling tinggi adalah klon
Nilai rataan dari klon pembanding dapat dilihat pada lampiran 10. Dari beberapa
klon yang diamati terdapat beberapa klon yang menunjukkan perbedaan yang
tidak nyata dengan klon pembanding BPM 24 seperti pada klon IRR 311, IRR 314
dan IRR 302.
Hal ini diduga muatan negatif fosfat akan membantu dalam menstabilkan
partikel karet sehingga koagulasi lateks akan terjadi lebih lambat dan lateks akan
lebih lama mengalir. Selain itu diduga bahwa pada tanaman dengan kadar fosfat
tinggi akan mampu mendukung berlangsungnya proses metabolisme terutama
yang berkaitan dengan biosintesis lateks.
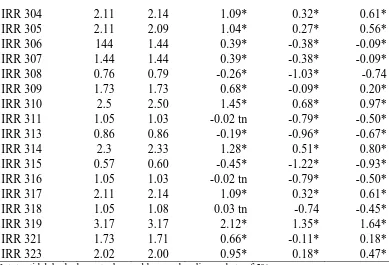
Tabel 4. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Fosfat (mM)
Klon
Hasil koreksi Rataan dikurang dengan klon pembanding belum sesudah PB 260 RRIC 100 BPM 24 Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
2. Aliran Indeks
Indeks Produksi
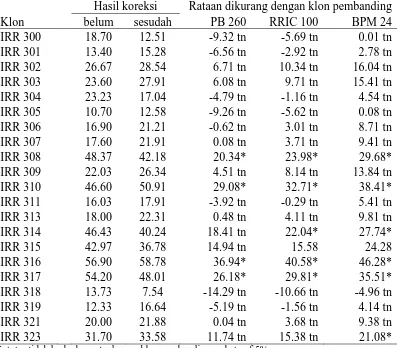
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 5. Dapat diketahui bahwa pada parameter indeks produksi menunjukkan
perbedaan yang nyata ke arah yang lebih tinggi dengan klon pembanding hampir
pada semua klon yang diuji kecuali pada klon IRR 319 dan IRR 311, jika
dibandingkan dengan klon PB 260 sebagai klon pembanding. Jika dibandingkan
dengan klon RRIC 100 dan BPM 24 terdapat beberapa klon yang memiliki
perbedaan yang nyata pada parameter indeks produksi, dan perbedaan tersebut
terjadi ke arah yang lebih tinggi atau positif, yaitu pada klon IRR 318, IRR 308,
IRR 310, IRR 309 IRR 313, IRR 323, IRR 305, IRR 316 dan IRR 302. Selain
klon tersebut seluruhnya menunjukkan perbedaan yang tidak nyata dengan klon
pembanding. Indeks produksi tertinggi terdapat pada klon IRR 309 (86,62) dan
yang terendah terdapat pada klon IRR 311 (61,08). Nilai rataan dari klon
pembanding pada parameter indeks produksi dapat dilihat pada Lampiran 13.
Hal ini menjelaskan bahwa adanya variasi yang dimiliki dari klon-klon
IRR seri 300 dan klon pembanding RRIC 100, BPM 24, dan PB 260. Perbedaan
indeks produksi dalam penelitian ini disebabkan oleh perbedaan volume lateks
yang dihasilkan tanaman. Klon ideal adalah klon yang memiliki indeks produksi
yang tinggi seperti pada klon IRR 309 (86,62). Indeks ini juga menggambarkan
produksi kulit. Indeks produksi dipengaruhi faktor anatomis dan fisiologis
tanaman. Oleh sebab itu, indeks produksi nilainya dipengaruhi oleh umur tanaman
Tabel 5. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Indeks Produksi
Klon
Hasil koreksi Rataan dikurang dengan klon pembanding belum sesudah PB 260 RRIC 100 BPM 24 Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
* = berbeda nyata dengan klon pembanding pada taraf 5%
Indeks Penyumbatan
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 6. Dapat diketahui bahwa pada parameter pengamatan indeks penyumbatan
menunjukkan perbedaan yang nyata dengan ketiga klon pembanding pada klon
IRR 317, IRR 308, IRR 310 dan IRR 316. Dari klon yang diuji, indeks
penyumbatan yang paling rendah dijumpai pada klon IRR 318 (7,54), tetapi nilai
ini tidak berbeda nyata dengan klon pembanding. Nilai rataan dari klon
pembanding pada parameter indeks penyumbatan dapat dilihat pada Lampiran 16.
Hasil pengamatan terdahulu yang dilakukan oleh Boatman (1966),
Sothorn dan Gomez (1970), menunjukkan proses penyumbatan tidak sama untuk
setiap klon, sehingga indeks penyumbatan dapat digunakan sebagai ciri spesifik
dari masing-masing klon.
Milford et al (1969), membuktikan bahwa produksi sangat efektif
dihasilkan oleh tanaman yang memiliki indeks penyumbatan yang rendah.
Menurut Ho (1975), klon yang demikian akan memperlihatkan waktu aliran yang
lebih lama dan ini merupakan ciri spesifik yang dimiliki oleh klon-klon tertentu.
Tabel 6. Tabel Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Indeks Penyumbatan
Klon
Hasil koreksi Rataan dikurang dengan klon pembanding belum sesudah PB 260 RRIC 100 BPM 24 Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
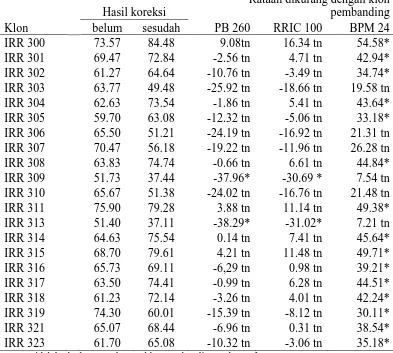
Panjang Alur Sadap
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 7. Dapat diketahui bahwa pada parameter panjang alur sadap menunjukkan
perbedaan yang nyata dengan klon pembanding yaitu pada klon IRR 318 dan
IRR 319 berbeda nyata dengan klon PB 260, klon IRR 309 dan IRR 302 berbeda
nyata dengan klon RRIC 100 dan klon IRR 318 menunjukkan perbedaan yang
nyata dengan klon BPM 24. Selain klon tersebut seluruhnya menunjukkan
perbedaan yang tidak nyata dengan klon pembanding. Panjang alur sadap tertinggi
terdapat pada klon IRR 318 (46,63 cm) dan yang terendah terdapat pada klon
IRR 302 (31,93 cm). Nilai rataan dari klon pembanding pada parameter
panjang alur sadap dapat dilihat pada Lampiran 19.
Hal ini menjelaskan bahwa adanya variasi yang dimiliki dari klon-klon
IRR seri 300 dan klon pembanding RRIC 100, BPM 24, dan PB 260. Hal ini
berarti bahwa semakin panjang alur sadap, maka semakin banyak jumlah
pembuluh lateks yang terpotong dan sebaliknya indeks penyumbatan semakin
kecil dan aliran lateksnya akan semakin lama mengalir
(Subronto dan Harris, 1977).
Adapun klon yang ideal adalah klon yang memiliki lilit batang besar
Tabel 7. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
* = berbeda nyata dengan klon pembanding pada taraf 5%
Kecepatan Aliran Lateks
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 8. Dapat diketahui bahwa pada parameter kecepatan aliran lateks
menunjukkan perbedaan yang nyata dengan ketiga klon pembanding yaitu pada
klon IRR 315, IRR 314, IRR 309, IRR 311, IRR 321, IRR 316 dan IRR 302
menunjukkan perbedaan ke arah yang lebih baik sedangkan klon IRR 306
menunjukkan perbedaan ke arah yang lebih rendah jika dibandingkan dengan klon
PB 260. Dari klon yang diuji , kecepatan aliran lateks yang paling tinggi dijumpai
pembanding. Sedangkan kecepatan aliran lateks yang paling rendah dijumpai
pada klon IRR 306 (6,06 g/mnt/cm) dan nilai ini tidak berbeda nyata jika
dibandingkan dengan klon RRIC 100. Nilai rataan dari klon pembanding pada
parameter kecepatan aliran lateks dapat dilihat pada Lampiran 22.
Kecepatan aliran lateks menunjukkan per satuan waktu per panjang alur
sadap yang dilalui. Artinya semakin cepat dan lama lateks mengalir maka
produksi semakin tinggi (Subronto dan Harris, 1977). Lambat cepatnya aliran
lateks sewaktu disadap berpengaruh terhadap tinggi rendahnya produksi. Semakin
cepat dan lama lateks mengalir, maka hasil lateksnya semakin tinggi. Dari hasil
penelitian yang telah dilakukan, ternyata pola aliran lateks itu berbeda-beda setiap
klon. Perbedaan aliran lateks ini memungkinkan disebabkan oleh banyaknya
pembuluh lateks yang terpotong. Selain itu, komposisi pembuluh lateks juga
berbeda. Berdasarkan hasil itu maka pola aliran lateks berbeda untuk setiap klon
Tabel 8. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
* = berbeda nyata dengan klon pembanding pada taraf 5%
3. Pertumbuhan
Lilit Batang
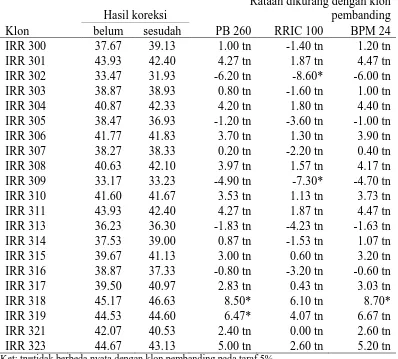
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 9. Dapat diketahui bahwa klon dengan lilit batang terbesar dijumpai pada
klon IRR 300, namun nilai ini tidak berbeda nyata dengan klon pembanding PB
260 dan RRIC 100 tetapi berbeda nyata dengan klon pembanding BPM 24.
Rataan nilai klon pembanding dapat dilihat pada Lampiran 25. Dari semua klon
yang diamati, lilit batang yang terkecil terdapat pada IRR 309 dan IRR 313, dan
dengan klon pembanding PB 260 dan RRIC 100 tetapi tidak berbeda nyata dengan
klon BPM 24. Berdasarkan hasil ini dapat dibuat suatu kesimpulan bahwa jika lilit
batang dijadikan sebagai acuan untuk penentuan permulaan sadap maka ada
kemungkinan klon-klon selain IRR 309 dan klon IRR 313 akan memiliki umur
untuk dapat disadap sama dengan klon PB 260 dan RRIC 100.
Pertumbuhan lilit batang akan mempengaruhi produksi tanaman karet,
dimana semakin besar lilit batang maka jumlah pembuluh, diameter dan kerapatan
kulitnya banyak, sehingga menghasilkan produksi yang tinggi
(Danimihardja, 1988).
Tabel 9. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Lilit Batang (cm) Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
Tebal Kulit
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 10. Dapat diketahui bahwa pada parameter tebal kulit menunjukkan
perbedaan yang nyata ke arah yang lebih tinggi dengan klon pembanding yaitu
pada IRR 307, sedangkan IRR 317, IRR 305 dan IRR 321 menunjukkan
perbedaan ke arah yang lebih kecil jika dibandingkan dengan klon PB 260 sebagai
klon pembanding. Jika dibandingkan dengan klon RRIC 100 terdapat beberapa
klon yang memiliki perbedaan yang nyata pada parameter tebal kulit, dan
perbedaan tersebut terjadi ke arah yang kebih kecil, yaitu pada IRR 304, IRR 317,
IRR 306, IRR 309 IRR 305, IRR 321 dan IRR 302. Dan jika dibandingkan dengan
klon BPM 24 terdapat beberapa klon yang memiliki perbedaan yang nyata pada
parameter tebal kulit, dan perbedaan tersebut juga terjadi ke arah yang lebih kecil,
yaitu pada klon IRR 304, IRR 317, IRR 305, IRR 321 dan IRR 302. Selain klon
tersebut seluruhnya menunjukkan perbedaan yang tidak nyata dengan klon
pembanding. Tebal kulit terbesar terdapat pada IRR 316 (8,67) dan yang terendah
terdapat pada IRR 317 (5,90). Nilai rataan dari klon pembanding pada parameter
indeks penyumbatan dapat dilihat pada Lampiran 28.
Hal ini menjelaskan bahwa adanya variasi yang dimiliki dari klon-klon
IRR seri 300 dan klon pembanding RRIC 100, BPM 24, dan PB 260.
Pertumbuhan tebal kulit merupakan karakteristik spesifik pada klon tertentu.
Tebalnya kulit dapat dipengaruhi oleh lingkungan, tetapi pada umumnya kulit
yang tipis karena kemungkinan terjadinya luka-luka ketika penyadapan dilakukan.
Tujuan melakukan evaluasi ketebalan kulit adalah untuk mendapatkan tanaman
lateksnya juga banyak. Tanaman yang mempunyai kulit yang terlalu tipis tidak
diinginkan karena ketika terjadi penyadapan dapat melukai kambium.
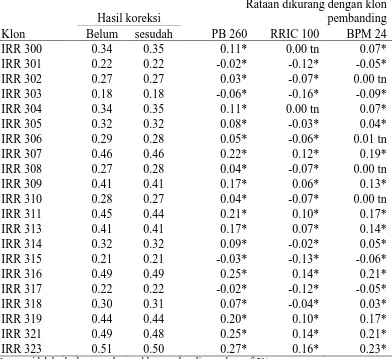
Tabel 10. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Tebal Kulit (mm)
Klon
Hasil koreksi Rataan dikurang dengan klon pembanding belum sesudah PB 260 RRIC 100 BPM 24 Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
* = berbeda nyata dengan klon pembanding pada taraf 5%
Anatomi Kulit
Jumlah Baris Pembuluh Lateks (buah)
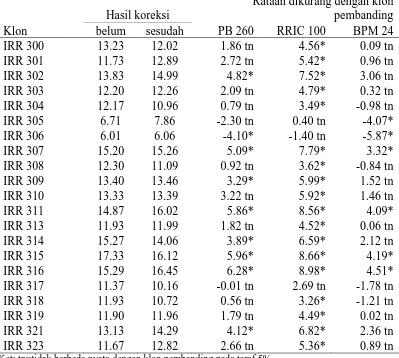
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 11. Dapat diketahui bahwa klon dengan jumlah baris pembuluh terbanyak
dijumpai pada klon IRR 315, dan nilai ini berbeda nyata dengan klon pembanding
PB 260, RRIC 100 dan BPM 24. Rataan nilai klon pembanding dapat dilihat pada
terdapat pada klon IRR 303 yang nilainya lebih rendah dengan ketiga klon
pembanding namun belum menunjukkan perbedaan yang signifikan dengan ketiga
klon pembanding.
Menurut Webster dan Baulkwill (1989), menyatakan bahwa baik
ketebalan asli maupun jumlah baris pembuluh lateks yang ada didalam semakin
meningkat dengan bertambahnya usia tanaman. Jumlah baris pembuluh lateks
pada prinsipnya merupakan ciri khas suatu klon tetapi perkembangannya
tergantung pada tingkat pertumbuhan tanaman yang dipengaruhi oleh
faktor-faktor seperti kepadatan tanaman dan status hara dan juga oleh klon.
Tabel 11. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Jumlah Baris Pembuluh Lateks (buah)
Klon
Hasil koreksi Rataan dikurang dengan klon pembanding Belum sesudah PB 260 RRIC 100 BPM 24 Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
Diameter Pembuluh Lateks (mikron)
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 12. Dapat diketahui bahwa pada parameter diameter pembuluh secara
umum menunjukkan nilai yang lebih rendah dengan klon pembanding PB 260
kecuali klon IRR 308, IRR 317 dan IRR 314, walaupun nilai dari ketiga klon ini
juga menunjukkan perbedaan yang tidak nyata dengan klon pembanding. Nilai
klon pembanding dapat dilihat pada Lampiran 34. Terdapat perbedaan yang nyata
antara klon yang diuji dengan klon RRIC 100 sebagai klon pembanding yaitu
pada klon IRR 318, IRR 304, IRR 317 dan IRR 314. Sedangkan jika
dibandingkan dengan klon BPM 24 terdapat perbedaan yang nyata ke arah yang
lebih kecil (nilai negatif) yaitu pada klon IRR 305 dan IRR 321.
Hal ini disebabkan karena pembuluh semakin dekat dengan kambium
maka aliran pembuluh semakin kecil. Diketahui bahwa pembuluh lateks
membentuk struktur, dimana pembuluh yang muda berada dekat dengan kambium
Tabel 12. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Diameter Pembuluh Lateks (mikron)
Klon
Hasil koreksi Rataan dikurang dengan klon pembanding belum sesudah PB 260 RRIC 100 BPM 24 Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
* = berbeda nyata dengan klon pembanding pada taraf 5%
4. Produksi (gr/p/s)
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 13. Dapat diketahui bahwa pada parameter produksi menunjukkan
perbedaan yang nyata ke arah yang lebih tinggi dengan klon pembanding pada
beberapa klon yang diuji, seperti pada klon IRR 300, IRR 317, IRR 315, IRR 308,
IRR 314, IRR 310, IRR 307, IRR 316, jika dibandingkan dengan klon PB 260
sebagai klon pembanding. Jika dibandingkan dengan klon RRIC 100 terdapat
beberapa klon yang memiliki perbedaan yang nyata pada parameter produksi, dan
perbedaan tersebut terjadi ke arah yang lebih tinggi, yaitu pada klon IRR 300,
dibandingkan dengan klon BPM 24 terdapat beberapa klon yang memiliki
perbedaan yang nyata pada parameter produksi, dan perbedaan tersebut juga
terjadi ke arah yang lebih tinggi, yaitu pada klon klon IRR 300, IRR 315,
IRR 308, IRR 310 dan IRR 316. Produksi tertinggi terdapat pada klon IRR 300
(56,04) dan yang terendah terdapat pada klon IRR 313 (43,19). Nilai rataan dari
klon pembanding pada parameter produksi dapat dilihat pada Lampiran 37.
Tabel 13. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Produksi (gr/p/s)
Klon
Hasil koreksi Rataan dikurang dengan klon pembanding belum sesudah PB 260 RRIC 100 BPM 24 Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
Kadar Karet Kering (%)
Berdasarkan hasil pengamatan yang telah diuji secara statistik pada
Tabel 14. Dapat diketahui bahwa pada parameter kadar karet kering secara
umum menunjukkan perbedaan yang tidak nyata dengan klon pembanding pada
beberapa klon yang diuji, kecuali pada klon IRR 318 jika dibandingkan dengan
klon PB 260. Jika dibandingkan dengan klon RRIC 100 terdapat beberapa klon
yang memiliki perbedaan yang nyata pada parameter kadar karet kering , dan
perbedaan tersebut terjadi kearah yang lebih tinggi, yaitu pada klon IRR 300,
IRR, 318, IRR 304, IRR 315, IRR 310, IRR 309, IRR 313, IRR 307dan IRR 311.
Sedangkan jika dibandingkan dengan klon BPM 24 secara umum memiliki
perbedaan yang tidak nyata pada parameter kadar karet kering, kecuali pada klon
IRR 316 yang memiliki kadar karet kering yang lebih rendah dengan klon
pembanding. Kadar karet kering tertinggi terdapat pada klon IRR 318 (36,02) dan
yang terendah terdapat pada klon IRR 316 (31,72). Nilai rataan dari klon
pembanding pada parameter kadar karet kering dapat dilihat pada Lampiran 40.
Subronto dan Harris (1977), menyatakan kadar karet kering yang tinggi
terutama disebabkan oleh viskositas lateks yang tinggi yang menyebabkan proses
Tabel 14. Hasil Rataan Yang Telah Dikoreksi dan Yang Belum Pada Parameter Ket: tn=tidak berbeda nyata dengan klon pembanding pada taraf 5%
* = berbeda nyata dengan klon pembanding pada taraf 5%
Korelasi Antar Parameter
Untuk melihat korelasi antar parameter berikut dibawah ini disajikan Tabel korelasi antar parameter sifat fisiologis dan sifat aliran lateks terhadap produksi.
Tabel 15. Korelasi Antar Parameter
Pr - r Tabel N = 21 λ 0,05 = 0, 433
Keterangan : PAS = Panjang Alur Sadap, IP = Indeks Penyumbatan,
KA = Kecepatan Aliran Lateks, IPr = Indeks Produksi, S = Sukrosa, T = Tiol,
F = Fosfat, Pr = Produksi.
Dari hasil analisa korelasi pada Tabel 15, memperlihatkan bahwa
sifat fisiologis (sukrosa, tiol, dan fosfat) dan aliran lateks (panjang alur sadap,
indeks penyumbatan, kecepatan aliran lateks dan indeks produksi) menunjukkan
pengaruh yang tidak nyata terhadap produksi. Tetapi menunjukkan nilai positif
kecuali pada parameter sukrosa dan tiol yang memiliki nilai negatif terhadap
produksi. Hal ini merupakan bahwa kadar sukrosa rendah (< 5 mM) memberikan
indikasi penggunaan bahan asimilat menjadi lateks sangat intensif atau dapat
dikatakan tanaman mengalami lelah fisiologi dengan kemampuan produksi
semakin menurun. Demikian juga ditunjukkan dengan kadar tiol yang umumnya
KESIMPULAN DAN SARAN
Kesimpulan
1. Dari hasil penelitian karakteristik sifat fisiologis lateks berdasarkan
pengamatan kadar sukrosa, tiol, dan fosfat anorganik secara umum
klon IRR seri 300 menunjukkan perbedaan yang nyata dengan ketiga klon
pembanding.
2. Pada karakteristik sifat aliran lateks berdasarkan parameter indeks produksi
dan kecepatan aliran lateks secara umum menunjukkan adanya perbedaan
yang nyata dengan ketiga klon pembanding.
3. Pada karakteristik pertumbuhan pada parameter jumlah pembuluh lateks klon
IRR 315 adalah klon terbaik yang menunjukkan perbedaan yang nyata dengan
ketiga klon pembanding.
4. Pada pengamatan produksi menunjukkan bahwa adanya variasi diantara klon
IRR seri 300 seperti IRR 300, IRR 303, IRR 305, IRR 306, IRR 309, IRR 310,
IRR 311, IRR 313, IRR 315, IRR 316, IRR 321 yang menunjukkan perbedaan
yang nyata dengan ketiga klon pembanding.
5. Adapun pada korelasi antar parameter yaitu produksi dengan karakteristik
sifat fisiologis (sukrosa, tiol, dan fosfat anorganik) dan sifat aliran lateks
(indeks produksi, indeks penyumbatan, panjang alur sadap, dan kecepatan
Saran
Klon IRR 315, IRR 318, IRR 319, IRR 302, dan IRR 309 merupakan klon
harapan untuk dapat dibudidayakan dalam skala terbatas untuk melihat potensi
DAFTAR PUSTAKA
Azwar, R., 1993. Analisis Konstribusi Klon Unggul Terhadap Peningkatan Produktifitas Perkebunan Karet. Perhimpunan Ilmu Pemuliaan Tanaman (PERIPI), Jakarta.
Azwar, R dan A. Daslin, 1993. Performance of 1974. Multilateral Exchange Klons Advarious Location in Indonesia, Jakarta.
Azwar, R., S. Woelan, A. Daslin, dan I. Suhendry,1998. Klon Harapan Seri IRR. Pros. Lok. Nas. Pemuliaan Karet 1998 dan Diskusi Nasional Prospek Karet Alam Abad 21. Pusat Penelitian Karet.
Boerhendy, I., 1988. Efek Okulasi Tajuk Terhadap Beberapa Sifat Anatomis dan Fisiologi Tanaman Karet. Balai Perkebunan Rakyat. BPP Sembawa.
Bricard, P., dan D. Nicolas, 1989. Possibility of The Use of Phisiological Parameters of Lateks in Early Selection. In d’ Auzac, J., J.L. Jacob and H. Chrestin, Florida.
Danimihardja, S., 1988. Hasil Pengujian Pendahuluan Klon Seri BPPB. Balai Perkebunan Rakyat. BPP Sembawa.
Ditjenbun (Direktorat Jenderal Perkebunan), 2007. Indonesia Miliki Perkebunan Karet Terluas di Duni
Djikman, M. J., 1951. Hevea. Thirthy Years of Research. University of Miammi Press, Coral Gobles Florida.
Edgar, A. T., 1958. Manual of Rubber Planting. The Incorporated Society of Planters, Kuala Lumpur.
Gills, G. E. Van dan H. Suharto, 1976. Aliran Lateks Komposisi dan Sifat Lateks. Menara Perkebunan.
Http:// www.binaukm.com, 2010. Persyaratan Tumbuh Tanaman Karet Dalam Budidaya Tanaman Karet
Jacob, J. L., J. C. Prevot and R.G.O. Kekwick, 1989. General Metabolish of Hevea brasiliensis. In d’ Auzac, J., J.L. Jacob and H. Chrestin. Physiologi of Rubber Tree Lateks. CRC Press, Florida.
. Diakses Pada Tanggal 18 Oktober 2010.
Lukman, 1980. Pembukaan Sadapan dan Stimulasi Sehubungan dengan Besarnya Lilit Batang. BPP Medan.
, 1984. Pembentukan Lateks dan Hubungannya dengan Penyadapan. Warta Perkaretan. BPP Sungai Putih.
Narayanan, R., Ho, C. Y., and Chen, K. T., 1973. Clonal Nursery Studies in Hevea III. Correlation Betwen Yield, Structural Characteristic, Latecs Constituent and Plugging Indecs. J. Rubb. Res. Ins., Malaysia.
Nazaruddin dan Paimin, 1992. Budidaya dan Pengolahan Karet. Strategi Tahun 2000. Penebar Swadaya, Jakarta.
Perries,O. S., and D. M. Fernando., 1983. A Hand Book of Rubber Culture and Processing. Rubber Research Institute of Srilanka.
Rasjidin, 1989. Bercocok Tanaman Karet (Hevea brasiliensis Muell. Arg) FP-UISU, Medan.
Setiawan, D. H., dan A. Andoko, 2005. Petunjuk Lengkap Budaya Karet. Agromedia Pustaka, Jakarta.
Southorn, W. A., 1961. Micropy of Hevea Lateks. Proc. Nat. rub. Res. Conf., Malaysia.
Southorn, W. A., and E. Yip, 1968. Lateks Flow Studies III. Elektrostatis Consideration in The Colloidals Stability of Fresh Hevea Lateks. Rubb. Res. Inst., Malaysia.
Subronto dan A. Harris, 1977. Indeks Aliran Sebagai Parameter Fisiologi Penduga Produksi Lateks. BPP Medan.
Subronto dan L. A. Napitupulu, 1978. Pengujian Klon Dengan Menggunakan Parameter Fisiologis Untuk Menkasir Kemampuan Produksi. BPP Medan.
Sumarmadji, Karyudi, dan THS Siregar, 2006. Rekomendasi System Eksploitasi Pada Klon Quick Dan Slow Stater Serta Penggunaan Irisan Ganda Untuk Meningkatkan Produktifitas Tanaman Karet. Pros. Lok. Nas. Bududaya Tanaman Karet 2006. Balai Penelitian Sungei Putih, Pusat Penelitian Karet.
Sutjihno, 2002. Rancangan Augmented RCB. Balitbio, Bogor.
Tan, A. 1987. Strategies in Ruber Tree Breeding. In Cambel, A. I. Abbots, A. J. Attan, R. R. Improvement of Vegetative Propagated Plants, Academic Press London.