Klasifikasi, Morfologi dan Nomenklatur Virus Influenza
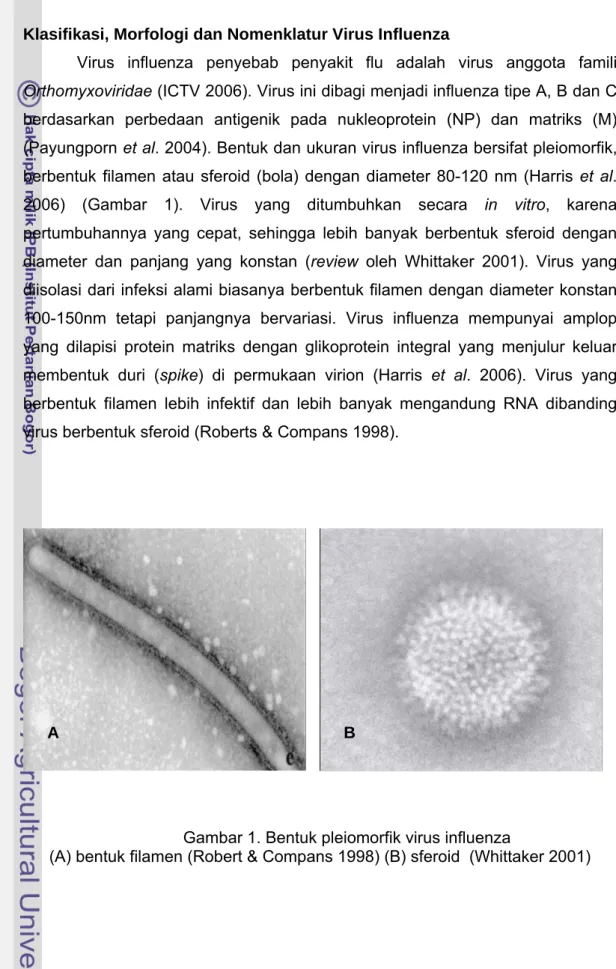
Virus influenza penyebab penyakit flu adalah virus anggota famili Orthomyxoviridae (ICTV 2006). Virus ini dibagi menjadi influenza tipe A, B dan C berdasarkan perbedaan antigenik pada nukleoprotein (NP) dan matriks (M) (Payungporn et al. 2004). Bentuk dan ukuran virus influenza bersifat pleiomorfik, berbentuk filamen atau sferoid (bola) dengan diameter 80-120 nm (Harris et al. 2006) (Gambar 1). Virus yang ditumbuhkan secara in vitro, karena pertumbuhannya yang cepat, sehingga lebih banyak berbentuk sferoid dengan diameter dan panjang yang konstan (review oleh Whittaker 2001). Virus yang diisolasi dari infeksi alami biasanya berbentuk filamen dengan diameter konstan 100-150nm tetapi panjangnya bervariasi. Virus influenza mempunyai amplop yang dilapisi protein matriks dengan glikoprotein integral yang menjulur keluar membentuk duri (spike) di permukaan virion (Harris et al. 2006). Virus yang berbentuk filamen lebih infektif dan lebih banyak mengandung RNA dibanding virus berbentuk sferoid (Roberts & Compans 1998).
B
A B
Gambar 1. Bentuk pleiomorfik virus influenza
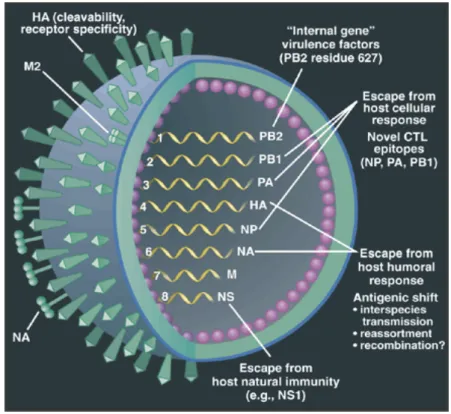
Di dalam virion influenza tipe A dan B terdapat 8 segmen genom RNA serat tunggal (single-stranded RNA) berpolaritas negatif yang menyandi 11 protein (Tabel 1). Kedelapan segmen RNA bersama-sama dengan nukleoprotein (NP) membentuk ribonukleoprotein (RNP) (Bui et al. 2000; Elton et al. 2001; Munch et al. 2001). RNP dikelilingi oleh protein matriks M1. Pada permukaan amplop virus terdapat glikoprotein HA dan NA serta kanal ion (ion channel) M2 (Elton et al. 2001). Struktur dan segmen-segmen genom virus influenza A terlihat pada Gambar 2. Virus influenza tipe C mempunyai 7 segmen genom RNA, karena hanya mempunyai satu jenis glikoprotein permukaan yaitu hemagglutinin esterase fusion (HEF). HEF berfungsi sebagai pengikat reseptor (H), fusi membran (F) dan esterase (E) (review oleh Whittaker 2001).
Virus influenza tipe A secara natural dapat menginfeksi unggas dan manusia (Khawaja et al. 2005). Virus ini dibagi ke dalam berbagai subtipe berdasarkan analisis serologis dan genetis glikoprotein hemaglutinin (HA) dan neuraminidase (NA) (Lee et al. 2001). Sampai saat ini ada 16 subtipe HA (H1-H16) dan 9 subtipe NA (N1-N9) (Russell and Webster 2005). Subtipe H16 baru ditemukan tahun 2004, pertama kali diisolasi dan diidentifikasi pada burung camar laut kepala hitam (Fouchier et al. 2005). Semua subtipe HA dan NA ditemukan pada unggas air, dan hanya 3 subtipe HA (H1-H3) dan 2 subtipe NA (N1-N2) ditemukan pada manusia (Hoffman et al. 2001). Subtipe H5 dan H7 yang sangat virulen pada unggas (Lee et al. 2001; Khawaja et al. 2005) dilaporkan berpotensi sebagai penyebab pandemi (Russell & Webster 2005).
Semua strain virus influenza diberi nama sesuai nomenklatur standar, berturut-turut tersusun dari tipe virus influenza/spesies hewan (jika bukan manusia)/wilayah isolasi/urutan nomor isolasi laboratorium/tahun isolasi (subtipe) (WHO 2002). Misalnya Influenza A/goose/Guangdong/1/1996 (H5N1).
Tabel 1. Segmen genom virus influenza A serta fungsi protein yang disandinya Segmen/ Gen Protein yang disandi Fungsi 1/PB2 Polimerase Basa 2
Perampasan tudung (cap snatching) Faktor virulensi
Polimerase Basa 1
Perampasan tudung Polimerisasi sintesis mRNA
Perakitan kompleks RdRp (RNA dependent RNA polymerase)
Menghambat respon imun seluler 2/PB1
PB1-frame 2 (PB1-F2)
Menginduksi apoptosis makrofag
Menurunkan penghilangan (clearance) virus Meningkatkan terjadinya infeksi sekunder oleh bakteri 3/PA Polimerase Asam Perakitan kompleks RdRp Endonuklease Replikasi
Menghambat respon imun seluler
4/HA Hemaglutinin Attchment (penempelan virus pada sel hospes) Fusi dengan membran endosom
Target netralisasi antibodi
5/NP Nukleoprotein Tanda isyarat (signal) impor vRNP Menghambat respon imun seluler
6/NA Neuraminidase Memotong ujung asam sialat dari reseptor sel hospes sehingga progeni virion lepas dari sel Target netralisasi antibodi
Matriks 1 (M1) Perakitan (assembly) progeni virus 7/M
Matriks 2 (M2) Tanda isyarat transpor ke permukaan sel Kanal ion
Nonstruktural 1 (NS1)
Ekspor mRNA virus dari nukleus
Menghambat pemotongan dan penyambungan (splicing) pre mRNA seluler
Menghambat ekspor mRNA seluler
Menghambat respon anti virus interferon (IFN) Menginduksi badai sitokin (sitokines storm) 8/NS
Nonstruktural 2 (NS2) atau Nuclear export protein (NEP)
Bersama-sama dengan M1 sebagai tanda isyarat ekspor vRNP dari nukleus
Gambar 2. Struktur dan segmen-segmen genom virus influenza A (Webster 2001)
Siklus Replikasi Virus Influenza
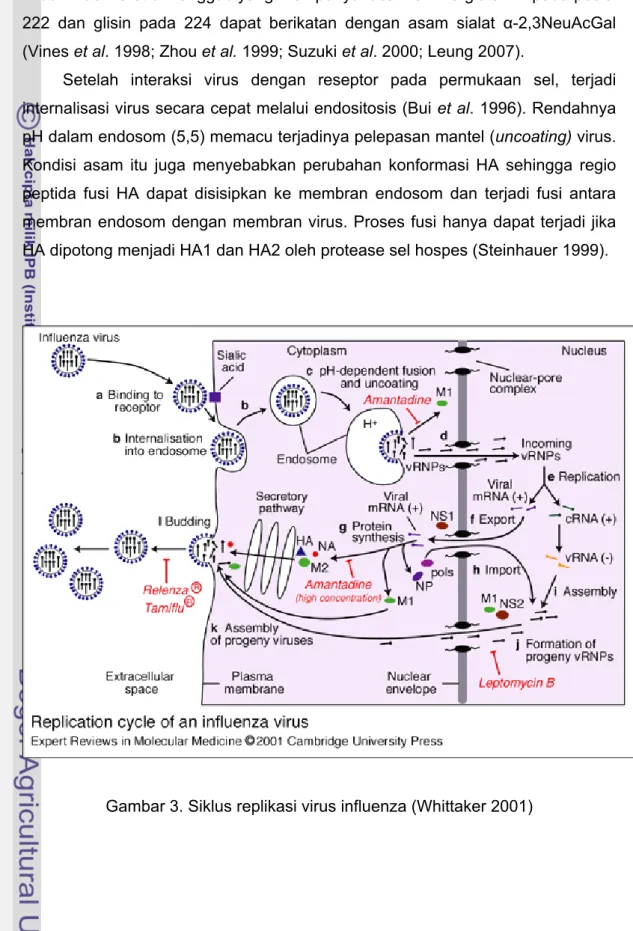
Siklus replikasi virus influenza A mempunyai keunikan karena semua sintesis mRNA dan replikasi genom terjadi di dalam nukleus sel hospes yang terinfeksi. Proses replikasi virus sangat cepat, sekitar 10 jam/siklus (Coleman 2007). Infeksi virus influenza diawali dengan masuknya virus ke dalam sel hospes (entry), diikuti transkripsi, translasi, perakitan dan budding virion-virion baru keluar sel hospes (Gambar 3).
Virus influenza masuk sel hospes melalui endositosis yang diperantarai reseptor (receptor–mediated endositosis) (Elton et al. 2001). Ikatan pada reseptor merupakan determinan awal patogenesitas, dan spesifisitas ikatan reseptor menentukan tropisme suatu virus pada spesies hospes tertentu. Residu asam amino bagian dari HA yang berikatan dengan reseptor adalah asam amino nomor 222 dan 224 (penomoran menurut H5). Glikoprotein HA virus influenza strain manusia yang mempunyai asam amino leusin pada posisi 222 dan serin pada 224 dapat berikatan dengan asam sialat α-2,6NeuAcGal. Sementara HA
virus influenza strain unggas yang mempunyai asam amino glutamin pada posisi 222 dan glisin pada 224 dapat berikatan dengan asam sialat α-2,3NeuAcGal (Vines et al. 1998; Zhou et al. 1999; Suzuki et al. 2000; Leung 2007).
Setelah interaksi virus dengan reseptor pada permukaan sel, terjadi internalisasi virus secara cepat melalui endositosis (Bui et al. 1996). Rendahnya pH dalam endosom (5,5) memacu terjadinya pelepasan mantel (uncoating) virus. Kondisi asam itu juga menyebabkan perubahan konformasi HA sehingga regio peptida fusi HA dapat disisipkan ke membran endosom dan terjadi fusi antara membran endosom dengan membran virus. Proses fusi hanya dapat terjadi jika HA dipotong menjadi HA1 dan HA2 oleh protease sel hospes (Steinhauer 1999).
Gambar 3. Siklus replikasi virus influenza (Whittaker 2001)
Rendahnya pH endosom juga menyebabkan aliran ion ke bagian interior virus melalui protein M2 dan memutus interaksi M1-vRNP (Pinto et al. 1992; Bui et al. 1996). Pelepasan mantel virus menyebabkan vRNP dan M1 masing-masing lepas ke sitoplasma dan menuju nukleus melewati nuclear core protein (NCP) (Bui et al. 1996). Impor vRNP melalui NCP diperantarai oleh nuclear localization signal (NLS) 1 dan 2 pada protein NP (Cros et al. 2005; Ozawa et al. 2007; Wu et al. 2007). Sementara, impor protein M1 ke dalam nukleus terjadi secara difusi pasif (Bui et al. 1996). Amantadin dan rimantadin yang banyak dipakai sebagai antivirus bekerja dengan menghambat aktivitas M2 untuk memutus interaksi M1-vRNP sehingga materi genetika virus tidak dapat masuk nukleus (Hayden 2006) Transkripsi dan replikasi genom RNA virus (vRNA) influenza dilakukan di dalam nukleus sel hospes, dikatalisis oleh enzim RdRp terdiri dari enzim PB1, PB2 dan PA (Honda et al. 2002; Crow et al. 2004; Hara et al. 2006). Genom vRNA membentuk kompleks dengan RdRp dan NP membentuk vRNP sebagai cetakan transkripsi (membentuk mRNA) dan cetakan replikasi (membentuk genom vRNA dari cRNA) (Vreede et al. 2004).
Sintesis mRNA virus diawali dengan penambahan fragmen tudung pada ujung ‘5 mRNA sebagai primer inisiasi transkripsi. Fragmen tudung (7mGppp dan 10-13 nukleotida setelah tudung) dari pre-mRNA sel hospes dipotong oleh enzim PB1 kemudian dikenal dan diikat oleh enzim PB2. Proses perampasan tudung dari pre-mRNA seluler tersebut disebut dengan cap snatching (Rao et al. 2003; Crow et al. 2004; Hara et al. 2006). Setelah penambahan tudung, pemanjangan (elongasi) rantai mRNA berjalan sampai pada sekuen kaya uridin yang terletak 15-22 nukleotida sebelum ujung 3’ mRNA. Pemanjangan mRNA virus ini dikatalisis oleh enzim PB1. Seperti juga mRNA eukariot, mRNA virus yang baru disintesis juga mengalami poliadenilasi pada ujung 3’, dikatalisis oleh RdRp (Honda et al. 2002).
mRNA virus tetap terlindung dari degradasi selama kompleks RdRp terikat pada sekuen spesifik 5’AGCAAAAGCAGG’3 yang ditemukan pada semua mRNA virus. Sekuen ini komplementer dengan 12 nukleotida ujung 3’ dari genom vRNA. Semua segmen genom virus influenza mempunyai 12 nukleotida pada ujung ‘3 dan 13 nukleotida pada ujung ‘5 yang bersifat stabil (Bae et al. 2001; Crow et al. 2004). Primer untuk mengamplifikasi secara lengkap semua genom virus
didisain berdasarkan regio genom yang identik dan stabil ini (Hoffman et al. 2001).
Replikasi genom vRNA tidak memerlukan primer dan dibentuk dari cetakan cRNA. Pada tahap I replikasi, vRNA dikopi menjadi cRNA berpolaritas positif. Inisiasi pembentukan cRNA tidak memerlukan tudung 7mGppp (Crow et al. 2004; Vreede et al. 2004; Hara et al. 2006). Tahap II replikasi adalah sintesis vRNA berpolaritas negatif dengan cRNA sebagai cetakannya (Hara et al. 2006). Seluruh serat cRNA disebut anti genom karena merupakan cetakan untuk sintesis vRNA. RdRp yang mengkatalisis replikasi genom, tidak mempunyai mekanisme untuk memperbaiki kesalahan (proofreading) sehingga tingkat kesalahan mencapai 1 dari 104 nukleotida per siklus replikasi (review oleh
Webster et al. 1992).
Pembentukan cRNA tidak memerlukan tudung 7mGppp. Perubahan fungsi katalitik polimerase dari transkripsi mRNA ke replikasi cRNA diperantarai oleh protein NP yang berikatan langsung dengan PB1 dan PB2 (Portela & Digard 2002). Perubahan fungsi katalitik polimerase PB1 memerlukan perubahan strukter sekunder polimerase, karena regio ikatan cRNA berbeda dengan regio ikatan mRNA (Gonzales & Ortin 1999). Berbeda dengan mRNA, 2 bentuk RNA lainnya (yaitu cRNA dan vRNA) dibungkus (encapsidated) oleh protein NP membentuk vRNP (Portela & Digard 2002).
Translasi (sintesis protein) dari mRNA virus influenza seluruhnya menggunakan mekanisme translasi dalam sitoplasma sel hospes. Protein PA, PB1, PB2 dan NP hasil translasi selanjutnya masuk ke nukleus untuk mengkatalisis transkripsi dan replikasi, kemudian dirakit dengan vRNA yang baru dan disintesis membentuk vRNP (Klumpp et al. 1997). vRNP diekspor ke sitoplasma melalui pembentukan kompleks NEP-M1-RNP, dan berinteraksi dengan reseptor ekspor nuklear sel hospes (superfamili importin-β) yang bersifat stabil (Cullen 2000; Neumann et al. 2000; Sandri-Goldin 2004). Ekspor vRNP virus influenza dihambat oleh antibiotik leptomisin B yang berikatan dengan CRM-1 (Elton et al. 2001).
Fragmen gen virus influenza A ada yang menyandi satu protein (PB1, PB2, PA, NA, HA, NP) ada yang lebih dari satu protein (gen NS dan M). Splicing mRNA dari gen NS menjadi mRNA NS1 dan mRNA NS2 (berturut-turut
menyandi protein NS1 dan NS2) dilakukan di nukleus sel hospes menggunakan mekanisme splicing pre-mRNA sel hospes. Splicing yang sama juga dilakukan terhadap mRNA gen M menjadi mRNA M1 dan mRNA M2 (Whittaker 2001).
Sebagai target protein transmembran, protein HA, NA dan M2 mengalami modifikasi pascatranslasi, berupa glikosilasi dan pelipatan, selama melintasi retikulum endoplasma dan aparatus Golgi (Gomez-Puertas et al. 2000). Sebagian molekul M1 berikatan dengan vRNP dan sebagian lagi membentuk selubung di bawah amplop virus. Glikoprotein HA pada transmembran menstimulasi M1 untuk berikatan/menempel pada membran. Interaksi antara ekor transmembran HA dengan M1 merupakan target perakitan virion (Ali et al. 2000; Gomez-Puertas et al. 2000; Ruigrok et al. 2000). Pada perakitan virion, semua komponen virus (HA, NA, M2, M1 dan vRNP) dibawa ke regio plasma membran sel yang kaya dengan detergent-insoluble glikolipid (DIG) atau disebut lipid raft (Zhang et al. 2000).
Perakitan virion diikuti dengan budding, yaitu pembentukan dan penutupan kuncup vRNP yang dikelilingi amplop pada membran sel hospes sehingga virion terlepas ke ekstrasel tanpa merusak membran sel (review oleh Garoff et al. 1998; Chazal & Gerlier 2003). Budding dapat terjadi melalui permukaan apikal atau basolateral sel epitel. Jika budding terjadi pada permukaan membran basolateral epitel, virus akan menyebar secara sistemik (Whittaker 2001).
Progeni virus dilepaskan ke ekstrasel jika NA memotong asam sialat dari reseptor sel hospes, sehingga progeni virus yang baru dilepaskan tidak berikatan kembali dengan reseptornya (Stray et al. 2000; Mishin et al. 2005). Peningkatan afinitas HA pada reseptor asam sialat dapat meningkatkan patogenesitas infeksi, namun di sisi lain dapat menghambat aktivitas NA pada proses budding. Pelepasan virus dan penyebarannya memerlukan keseimbangan fungsi antara kedua glikoprotein (HA dan NA) tersebut (Kobasa et al. 2001). Inhibitor neuraminidase sebagai antivirus yaitu zanamivir (Relenza) dan oseltamivir (Tamiflu) menghambat aktivitas NA sehingga budding tidak dapat terjadi (Hayden 2006).
Patogenesitas Virus Avian Influenza
Sebagai patogen intraseluler, virus avian influenza (VAI) mempunyai mekanisme untuk menghindari respon imun hospes sehingga virus dapat bertahan hidup dan bereplikasi dalam tubuh hospes. Peningkatan kemampuan virus untuk menghindari sistem imun hospes, secara langsung berkorelasi dengan peningkatan patogenesitas virus. VAI mempunyai berbagai mekanisme untuk menghindar dari sistem imun bawaan dan respon imun perolehan (adaptif) hospes (Coleman 2007).
Protein NS1
Protein NS1 merupakan faktor virulensi VAI, berperan sebagai antagonis IFN α/β. NS1 menghambat pengaktifan dsRNA-dependent protein kinase R (PKR) dari signal IFN α/β sehingga produksi IFN α/β terhambat (Talon et al. 2000; Fernandez-Sesma et al. 2006; Garcia-Sastre 2006; Hale et al. 2006; Coleman 2007). Hambatan sintesis IFN α/β oleh sel terinfeksi VAI juga terjadi melalui penghambatan NS1 pada pengaktifan 2’-5’ oligoadenilat sintetase (2’-5’-OAS) (Hale et al. 2006; Min & Krug 2006). Patogenesitas NS1 juga terjadi melalui ikatan langsung NS1 dengan p85β, yaitu subunit regulator fosfatidilinositol-3-kinase (PI3K), sehingga replikasi virus dalam sel terinfeksi tidak terhambat (Hale et al. 2006).
Patogenesitas infeksi VAI H5N1 pada manusia berbeda dengan influenza biasa (H1N1, H3N2), yaitu adanya hiperinduksi sitokin proinflamasi sehingga menimbulkan hipersitokinemia, yang secara populer disebut ‘badai sitokin’ (Guan et al. 2004; Lee et al. 2005). Sintesis berlebihan (overekspresi) protein HA, NP atau M di dalam sel terinfeksi VAI akan memicu NF-кB signaling pathway melalui pengaktifan IкB kinase (IKK) (Flory et al. 2000). Kedua virus influenza A subtipe H5N1 dan H1N1 ini menginduksi produksi sitokin melalui pengaktifan NF-кB atau degradasi IкB-α. Namun, VAI subtipe H5N1 mengaktifkan mitogen activated protein kinase (MAPK) secara dominan, termasuk p38 MAPK dan extracellular signal regulated kinase 1 dan 2 (ERK1/2), sehingga meningkatkan ekspresi TNFα oleh makrofag (Lee et al. 2005). Mutasi NS1 menyebabkan penurunan kemampuan virus menghambat produksi IFN α/β, sehingga NS1 berpotensi
besar sebagai target pembuatan antivirus dan vaksin (Solorzano et al. 2005; Fernandez-Sesma et al. 2006; Garcia-Sastre 2006).
Imunitas adaptif juga dihambat NS1 melalui induksi transkripsi faktor-faktor yang terlibat dalam maturasi sel dendritik serta migrasi dan stimulasi sel T. Hal ini menyebabkan maturasi dan kapasitas sel dendritik untuk menstimulasi respon imun sel T terhambat. Imunitas seluler melalui sel T merupakan mekanisme pertahanan tubuh untuk penghilangan virus dari tubuh (Fernandez-Sesma et al. 2006). Sel T sitotoksik (cytotoxic T lymphocyte; CTL) berperan penting mengontrol infeksi virus (Wherry & Ahmed 2004; Thomas et al. 2006). Untuk menghindari respon CTL, VAI mengakumulasi substitusi asam amino pada epitop atau dekat epitop CTL, seperti asam amino 380-388 dan 383-391 pada NP. Substitusi R384G pada NP menyebabkan hilangnya epitop CTL sehingga virus terhindar dari respon CTL (Berkoff et al. 2005).
Di dalam nukleus, NS1 menghambat ekspresi gen sel hospes dengan menghambat ekspor mRNA seluler ke sitoplasma. Mekanisme tersebut dilakukan NS1 melalui penghambatan poliadenilasi mRNA seluler dan interaksi NS1 dengan kompleks protein ekspor nuklear sel hospes (Satterly et al. 2007). Terhambatnya ekspor mRNA seluler dari nukleus menyebabkan translasi hanya dilakukan pada mRNA lama yang diekspor ke sitoplasma sebelum sel terinfeksi VAI.
Protein PB1-F2
Protein PB1-F2 adalah protein yang diekspresikan dari bingkai pembacaan terbuka (open reading frame:ORF) alternatif pada segmen gen polimerase PB1. Protein ini merupakan faktor patogenesitas virus influenza secara in vivo (Garcia-Sastre 2006; Coleman 2007). Protein PB1-F2 mampu menginduksi apoptosis makrofag sehingga menurunkan kemampuan sel hospes untuk menghilangkan virus dan meningkatkan infeksi sekunder bakteri oportunistik (Coleman 2007). ORF alternatif terletak pada basa ke (+)120 setelah ORF gen PB1. Ekspresi ORF alternatif ini melalui mekanisme ribosomal scanning, yaitu pembacaan ribosom pada kodon inisiasi translasi (AUG). Jika ribosom mengenali AUG pada ORF alternatif, polipeptida baru akan terbentuk. PB1-F2 hanya bertahan dalam sel selama 5 jam pascainfeksi (Coleman 2007). Protein ini
terlokalisasi di membran mitokondria dan secara dramatik menyebabkan degradasi morfologi mitokondria, menurunkan potensial membran dan menginduksi apoptosis (Garcia-Sastre 2006; Coleman 2007).
Virus AI yang dihilangkan ORF alternatif PB1-F2 tetap hidup secara in vitro, namun secara in vivo menurunkan kemampuan induksi apoptosis makrofag sampai 50%. Hal ini menunjukkan bahwa protein PB1-F2 hanya meningkatkan patogenesitas virus secara in vivo (Zamarin et al. 2006). Karena PB1-F2 dapat menyebabkan apoptosis makrofag sebagai antigen presenting cell (APC) profesional, kematian makrofag juga mempengaruhi presentasi antigen pada cabang respon imun adaptif (Coleman 2007).
Glikoprotein hemaglutinin
Glikoprotein HA, merupakan faktor patogenesitas virus influenza. HA berperan sebagai pengikat reseptor sel, fusi membran serta target utama netralisasi oleh antibodi sel hospes (Cross et al. 2001; Hulse et al. 2004; Hoffman et al. 2005; Gambaryan et al. 2006). Daerah pemotongan HA menjadi subunit HA1 dan HA2 menentukan patogenesitas VAI. Proteolisis HA pada daerah pemotongan bersifat spesifik dan spesifisitas jenis protease membatasi distribusi jaringan yang dapat diinfeksi virus ini. Kebanyakan VAI nonvirulen atau patogenik rendah mempunyai daerah pemotongan satu asam amino basa, namun strain patogenik tinggi mempunyai daerah pemotongan lebih dari satu asam amino basa (Munch et al. 2001).
Pada subunit HA1 terdapat domain-domain penting, seperti daerah antigenik, posisi glikosilasi dan kantong pengikat reseptor. Daerah antigenik fungsinya berkaitan erat dengan pertahanan terhadap respon imun hospes, sementara kantong pengikat reseptor fungsinya berkaitan dengan daya adaptasi pada hospes dan patogenesitas strain. Posisi glikosilasi turut menentukan afinitas ikatan reseptor serta pengenalan daerah antigenik oleh antibodi (Hulse et al. 2004; Gambaryan et al. 2006; Smith et al. 2006a; Stevens et al. 2006)
Mutasi dan Perubahan Antigen
Enzim RdRp tidak mempunyai mekanisme enzimatik perbaikan kesalahan replikasi, sehingga perubahan nukleotida terjadi terus menerus. Berbeda dengan
polimerase DNA yang hanya mempunyai kesalahan 1 dari 109 basa, kesalahan replikasi oleh RdRp adalah 1 dari 104 nukleotida per siklus replikasi (review oleh
Webster et al. 1992). Substitusi titik/poin dapat dibedakan atas substitusi sinonim dan substitusi nonsinonim.
Substitusi sinonim adalah perubahan nukleotida tidak diikuti perubahan ekspresi asam amino. Hal ini terjadi pada semua asam amino, kecuali metionin dan triptofan yang hanya disandi oleh 1 kodon. Substitusi sinonim ini menyebabkan kodon bias, yaitu ketidakseimbangan penggunaan kodon sinonim yang menyandi asam amino. Kodon bias ini terlihat pada semua spesies di semua bagian genom, baik daerah intron maupun ekson. Karena kodon bias tidak mengubah fenotip produk ekspresi, sehingga kodon bias selalu ada dalam genom. Penggunaan kodon (codon usage) pada gen berkorelasi dengan akurasi
dan tingkat translasi. Kodon pilihan (codon preference) biasanya adalah kodon yang tRNA untuk kodon tersebut melimpah sehingga dapat ditranslasi lebih cepat (Lavler & Kotlar 2005; Wu & Freeland 2005).
Seperti organisme lainnya, substitusi sinonim pada VAI juga berkaitan dengan kelimpahan tRNA (Plotkin & Dushoff 2003). Namun, mengingat translasi mRNA pada VAI menggunakan mekanisme translasi sel hospes, substitusi sinonim tersebut lebih dikarenakan seleksi penyesuaian terhadap penggunaan kodon sel hospes. Hal ini terjadi karena perbedaan penggunaan kodon antara virus dengan sel hospes dapat mempengaruhi kecepatan translasi protein (Garmory et al. 2003).
Substitusi nonsinonim adalah perubahan nukleotida diikuti dengan perubahan ekspresi asam amino. Substitusi nonsinonim hanya terjadi pada bagian tertentu dari gen yang mengalami tekanan. Semakin sering mengalami tekanan, semakin tinggi tingkat substitusinya (Plotkin & Dushoff 2003). Adanya tekanan seleksi akan menyebabkan munculnya varian dengan tingkat efektivitas replikasi yang tinggi (Jong et al. 2000). Tingkat perubahan asam amino virus di dalam tubuh hospes (in vivo) lebih tinggi dibandingkan virus yang ditumbuhkan secara in vitro. Hal ini menunjukkan bahwa tingginya tekanan imun berkorelasi dengan perubahan asam amino (Nakajima et al. 2003).
Kecepatan substitusi nonsinonim virus influenza mencapai 2-3x substitusi per posisi per tahun (Tumpey et al. 2002; Swayne & Suarez 2003). Rasio
kecepatan mutasi nonsinonim dan sinonim sangat penting untuk mempelajari mekanisme evolusi molekuler sekuen gen tertentu. Rasio kecepatan mutasi nonsinonim/sinonim (ω = dN/dS) juga merupakan indikator tekanan seleksi pada level protein. Jika ω=1 berarti seleksi netral, ω<1 berarti seleksi pemurnian (purifying selection) dan ω=>1 berarti seleksi positif (Yang et al. 2000).
Analisis genom VAI subtipe H5N1 yang menginfeksi unggas dan manusia dari tahun 1997-2004 menunjukkan bahwa gen PB2, HA dan NS1 mengalami tekanan seleksi positif, sementara gen lainnya (PA, PB1, M, NA, NS2, NP) mengalami tekanan seleksi pemurnian (Campitelli et al. 2006). Namun, isolat VAI H5N1 penyebab wabah di Indonesia dan Vietnam pada dekade terakhir menunjukkan bahwa hanya gen M2 (ω=1,23) dan PB1-F2 (ω=3,01) yang mengalami seleksi positif. Hal ini menunjukkan keterlibatan gen ini dalam adaptasi VAI pada hospes baru dan transmisi interspesies. Seleksi positif pada gen M2 terjadi akibat tekanan seleksi untuk adaptasi VAI dari unggas air ke unggas darat. Perbedaan pH dan lingkungan seluler antara unggas air dan unggas darat merupakan tekanan seleksi pada M2 sebagai kanal ion hidrogen. Tekanan seleksi pada PB1-F2 dikarenakan peran protein ini dalam menginduksi apoptosis makrofag (Smith et al. 2006a).
Adaptasi selalu dilakukan VAI, baik adaptasi terhadap tekanan imun maupun adaptasi pada spesies hospes baru (Voeten et al. 2000; Taubenberger et al. 2005). Adaptasi merupakan kekuatan utama dari evolusi. Perbedaan spesies hospes dan perbedaan tekanan menyebabkan pebedaan kecepatan evolusi VAI (Brown et al. 2001). Lama infeksi dan frekuensi reinfeksi virus influenza pada manusia, menyebabkan tingginya tekanan seleksi oleh sistem imun (Bush et al. 1999; Suzuki & Nei 2002).
Kecepatan mutasi glikoprotein HA kira-kira 2 x 10-3 nukleotida per posisi per
replikasi (Webster et al. 1992). Kecepatan mutasi HA tersebut lebih tinggi dibanding NA karena NA bukan merupakan determinan antigenik utama dan jumlah NA pada permukaan virion hanya 1/5 jumlah HA (Plotkin & Dushoff 2003).
Protein internal tidak berperan dalam pengikatan dengan reseptor sel hospes dan tersembunyi dari antibodi, sehingga protein ini lebih stabil dibanding glikoprotein permukaan (Plotkin & Dushoff 2003; Berkhoff et al. 2005). Struktur dan fungsi protein internal juga sangat mendasar sehingga tidak menguntungkan
VAI jika mutasi terjadi secara cepat. Hal ini menyebabkan VAI menghadapi konflik intragenom tentang kecepatan mutasi. Gen atau bagian spesifik gen tertentu dalam genom tersebut mengalami seleksi positif untuk berubah, sementara gen lain mengalami seleksi pemurnian untuk tidak berubah (Plotkin & Dushoff 2003).
Protein/regio protein yang fungsinya berkaitan erat dengan pertahanan terhadap respon imun hospes, daya adaptasi dan patogenesitas mempunyai tingkat substitusi nonsinonim lebih tinggi dibanding substitusi sinonim (Plotkin & Dushoff 2003). Kecepatan substitusi nonsinonim gen subunit HA1 dari VAI subtipe H3 sebesar 5,7 x10-3 per posisi per tahun. Hal ini disebabkan karena
pada HA1 terdapat daerah antigenik, kantong pengikat reseptor dan posisi glikosilasi (Bush et al. 1999).
Hanyutan antigenik
Adaptasi terhadap tekanan imun hospes dilakukan VAI untuk menghindar dari pengenalan dan netralisasi antibodi dan sel T sitotoksik. Antibodi netralisasi terhadap protein HA bersifat protektif melawan infeksi, sehingga protein ini paling tinggi mengalami tekanan imun dibandingkan protein internal (Berkhoff et al. 2005). Mekanisme VAI untuk menghindar dari sistem imun hospes merupakan tekanan untuk mutasi secara gradual sehingga muncul strain-strain virus baru yang secara imunologik berbeda (hanyutan antigenik) (Horimoto & Kawaoka 2001; Munch et al. 2001; Smith et al. 2004).
Hanyutan antigenik adalah perubahan secara periodik akibat mutasi genetik struktur glikoprotein permukaan VAI sehingga antibodi yang telah terbentuk oleh tubuh akibat infeksi atau vaksinasi sebelumnya tidak dapat mengenali keberadaan virus tersebut (Munch et al. 2001). Hanyutan antigenik berjalan lambat namun progresif dan cenderung menimbulkan penyakit yang terbatas pada suatu kawasan tertentu (Tumpey et al. 2002; Swayne & Suarez 2003). Hanyutan antigenik menuntut pembuatan vaksin selalu diperbarui mengikuti munculnya strain virus baru (Plotkin et al. 2002; Smith et al. 2004).
Reasorsi dan transmisi Virus Avian Influenza
Pandemi dapat terjadi jika subtipe virus influenza baru dapat melintasi barier hospes antara unggas dan mamalia, termasuk manusia. Adaptasi VAI strain unggas ke manusia antara lain melalui reasorsi (reassortment), yaitu pertukaran atau pencampuran gen. Genom RNA yang tersusun bersegmen-segmen memudahkan terjadinya reasorsi, yaitu bersegmen-segmen gen pada strain tertentu digantikan segmen gen sealel dari strain lainnya. Reasorsi menyebabkan perubahan struktur antigen secara dominan, sehingga disebut lompatan antigenik (antigenic shift). Reasorsi hanya dapat terjadi jika suatu sel secara simultan terinfeksi oleh 2 atau lebih strain VAI yang berbeda, sehingga terjadi penyusunan kembali suatu strain virus baru yang bermanifestasi sebagai genotipe virus baru. Hospes yang dapat diinfeksi oleh 2 jenis strain VAI yaitu strain avian dan manusia dikenal dengan “mixing vessel”. Hospes ini memungkinkan sebagai hospes perantara transmisi VAI dari unggas ke manusia (Ito et al. 1998; Hoffman et al. 2001; Li et al. 2004).
Virus influenza A subtipe H1N1 penyebab pandemi influenza tahun 1918 mengalami lompatan antigenik sehingga tahun 1958 muncul subtipe H2N2 dan tahun 1968 muncul subtipe H3N2 (Belshe 2005). Transmisi langsung VAI Vdari unggas ke manusia biasanya mengakibatkan kematian, seperti terjadi di Hongkong tahun 1997-1998. Virus HPAI H5N1 yang menyerang dan mematikan manusia dan ayam di Hongkong tersebut (Lee et al. 2001), merupakan produk reasorsi dengan VAI H9N2 yang bertindak sebagai donor gen internal (Guan et al. 1999). Virus tersebut kemudian berkembang cepat di pasar unggas Hongkong, dan mempunyai kemampuan untuk transmisi langsung ke manusia (Zhou et al. 1999; Cauthen et al. 2000).
Kejadian tersebut merupakan kasus pertama, dimana infeksi VAI H5N1 langsung pada manusia tanpa terlebih dulu beradaptasi pada hospes mamalia perantara (Tumpey et al. 2002; Rowe et al. 2003; Sturm-Ramirez et al. 2004). Virus HPAI H5N1 penyebab wabah di Danau Qianghai Cina tahun 2005 yang mematikan ribuan unggas air migratori dilaporkan juga merupakan virus hasil reasorsi (Zhou et al. 2006). Burung puyuh menyediakan lingkungan yang memungkinkan VAI H3N2 babi mengalami reasorsi dan menghasilkan virus influenza yang berpotensi menyebabkan pandemi (Perez et al. 2003).
Transmisi VAI H5N1 dari manusia ke manusia belum pernah dilaporkan (Buxton et al. 2000; The Writing Committee WHO 2005; Kandun et al. 2006). Namun, VAI subtipe H5N1 berpotensi sebagai penyebab pandemi influenza pada manusia melalui 2 mekanisme. Manusia yang terinfeksi VAI H5N1 dan strain influenza manusia (misalnya H1N1) akan memicu reasorsi, sehingga memunculkan VAI subtipe H5 yang mampu ditransmisikan dari manusia ke manusia. Alternatif lain adalah mutasi langsung VAI H5N1 yang berkemampuan untuk transmisi dari manusia ke manusia (
Russell & Webster 2005).
Profil Glikoprotein Hemaglutinin
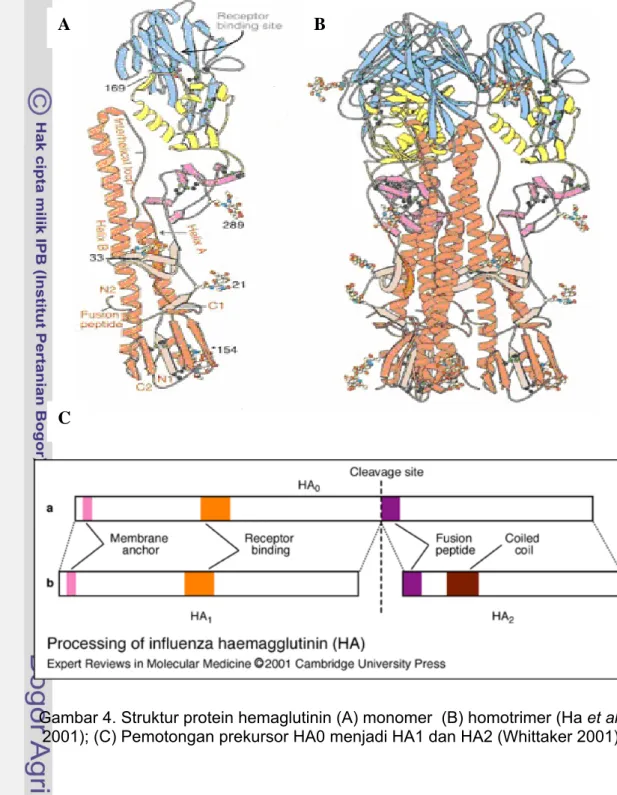
Protein HA disintesis sebagai polipeptida 76 kDa. Setelah translasi di retikulum endoplasma, HA mengalami maturisasi di aparatus Golgi menjadi homotrimer HA masing-masing 220 kDa (Gambar 4A, 4B). Setiap monomer awalnya merupakan prekursor polipeptida tunggal (HA0) kemudian dipotong
menjadi 2 subunit yaitu HA1 dan HA2 (Gambar 4C). Kedua subunit ini
dihubungkan oleh ikatan disulfida antara residu asam amino 14 dari HA1 dengan
residu asam amino 137 dari HA2. Tanpa proteolisis HA menjadi HA1 dan HA2,
proses fusi virus dengan membran endosom tidak terjadi sehingga virus bersifat noninfeksius (Steinhauer 1999).
Pada HA1 terdapat daerah antigenik, kantong pengikat reseptor, residu pengikat reseptor dan posisi glikosilasi. Daerah antigenik fungsinya berkaitan erat dengan pertahanan terhadap respon imun hospes, sementara kantong pengikat reseptor fungsinya berkaitan dengan daya adaptasi pada hospes dan patogenesitas strain. Posisi glikosilasi turut menentukan afinitas ikatan reseptor serta pengenalan daerah antigenik oleh antibodi (Hulse et al. 2004; Gambaryan et al. 2006; Smith et al. 2006a; Stevens et al. 2006). Peptida fusi pada ujung N subunit HA2 berperan pada fusi membran saat infeksi virus ke dalam sel hospes (Cross et al. 2001). Meskipun tingkat mutasi HA paling tinggi dibanding protein lain, namun pada bagian-bagian tertentu dari HA bersifat stabil pada semua influenza A (Wagner et al. 2005)
A
B
C
Gambar 4. Struktur protein hemaglutinin (A) monomer (B) homotrimer (Ha et al. 2001); (C) Pemotongan prekursor HA0 menjadi HA1 dan HA2 (Whittaker 2001)
Sekuen yang bersifat stabil
Pada ujung C glikoprotein HA mempunyai 3 residu sistein pada posisi 551, 559, 562 yang bersifat stabil (Wagner et al. 2005). Bagian transmembran HA terdiri dari 10-11 asam amino, dan 5 diantaranya bersifat stabil pada semua
influenza A. Meskipun sekuen ini tidak esensial untuk perakitan dan daya infeksi virus (Jin et al. 1994), namun interaksi bagian transmembran HA dengan protein internal sangat menentukan bentuk virion (Jin et al. 1997). Asam amino pertama (D:aspartat) dan terakhir (P:prolin) dari subunit HA1 juga bersifat stabil pada semua influenza A (Stevens et al. 2006).
Daerah pemotongan (cleavage site) hemaglutinin
Daerah pemotongan HA adalah sekuen asam amino pada HA sebagai tempat pemotongan prekursor HA (HA0) menjadi HA1 dan HA2 oleh protease sel
hospes, sehingga proses fusi dengan membran endosom pada saat infeksi VAI ke dalam sel hospes dapat terjadi. Daerah pemotongan HA0 tergantung pada
keberadaan asam amino basa arginin (R) atau lisin (K). Daerah pemotongan bersifat spesifik dan spesifisitas jenis protease membatasi distribusi jaringan yang dapat diinfeksi virus ini. Kebanyakan VAI non-virulen atau low pathogenic mempunyai satu asam amino basa pada daerah pemotongan, namun strain highly pathogenic mempunyai lebih dari satu asam amino basa (polybasic) pada posisi tersebut (Munch et al. 2001).
Sekuen HA dengan daerah pemotongan monobasic (contoh: HA1
-PSIQVR-GL-HA2) dapat dipotong oleh tryptase yang dihasilkan sel epitel traktus respirasi
dan pencernaan (Whittaker 2001; Chen et al. 2004). Secara in vitro, daerah pemotongan HA monobasic juga dapat dipotong oleh trypsin-like enzyme, seperti faktor pembeku darah “Xa”, mini plasmin dan protease bakteri (Murakami et al. 2001). Protease dari Staphylococcus aureus dan Pseudomonas aeruginosa juga dilaporkan dapat memotong daerah pemotongan monobasic (Zhirnov et al. 2002).
Sekuen HA dengan daerah pemotongan polybasic (contoh: HA1
-KKREKR-GL-HA2), memungkinkan proses proteolitik dapat dilakukan oleh protease lain
seperti furin dan proprotein konvertase 6 (PC6) yang terdapat di aparatus Golgi semua sel (Horimoto et al. 1994). Enzim proteolitik furin mengenal sekuen asam amino motif B-X-B-R (B=asam amino basa, X=asam amino nonbasa) (Walker et al. 1994). VAI dengan daerah pemotongan polybasic mempunyai jaringan distribusi yang tidak terbatas dan menyebabkan infeksi sistemik yang fatal (Whittaker 2001; Chen et al. 2004). Daerah pemotongan polybasic pada VAI
H5N1 bertanggung jawab terhadap infeksi sistemik sehingga virus dapat diisolasi dari darah, cairan serebrospinal dan feses (WHO et al. 2005).
Sekuen daerah pemotongan QRERRRKKR khas pada strain patogenik VAI H5N1 penyebab kematian unggas di Indonesia dan Vietnam (Smith et al. 2006a; Stevens et al. 2006). Titik pemotongan QRESRRKKR adalah khas pada VAI H5N1 penyebab kematian manusia di Indonesia tahun 2005-2007 (CDC 2007).
Daerah antigenik (antigenic sites)
Pada glikoprotein HA dikenal 5 epitop daerah antigenik sebagai target netralisasi antibodi (Smith et al. 2004). Penentuan 5 epitop (A sampai E) daerah antigenik tersebut didasarkan pada struktur HA virus influenza A subtipe H3N2 penyebab pandemi flu di Hongkong 1968. Daerah antigenik tersebut tampaknya tidak bersifat linier, melainkan konformasional. Masing-masing epitop terbentuk pada struktur tersier molekul HA, sehingga asam-asam amino yang saling berjauhan bersama-sama membentuk satu epitop. Kelima epitop tersebut terdapat pada 25 residu asam amino subunit HA1. Epitop A dibentuk oleh residu asam amino 135, 124, 133, 145, 144, 142, dan 131. Epitop B dibentuk oleh asam amino 156, 197, 189, 190, 157, 196, 193 dan 158. Epitop C dibentuk oleh asam amino 276, 278, 275 dan 299. Epitop D dibentuk oleh asam amino 226, 121 dan 172. Epitop E dibentuk oleh residu asam amino 262, 62 dan 83 (penomoran menurut H3) (Plotkin & Dushoff 2003).
Substitusi asam amino pada daerah antigenik merupakan salah satu pendorong evolusi gen hemaglutinin (Shih et al. 2007). Dari 25 asam amino pada 5 epitop tersebut, 14 residu asam amino diantaranya (121, 124, 133, 135, 142, 145, 156, 158, 190, 193, 197, 226, 262, 275) menunjukkan seleksi positif (Bush et al. 1999). Seleksi positif pada daerah antigenik disebabkan tekanan seleksi untuk menghindar dari respon imun hospes, termasuk penghindaran pengenalan antibodi yang terbentuk akibat vaksinasi (Plotkin & Dushoff 2003; Smith et al. 2004; Campitelli et al. 2006). Perubahan epitop terjadi terus menerus, dan perubahan epitop pada jangka 2-5 tahun biasanya hanya didominasi oleh satu epitop saja. Dengan demikian setiap 2-5 tahun terjadi pergeseran dominasi perubahan epitop (Plotkin et al. 2002).
Kantong pengikat reseptor (receptor binding pocket)
Kantong pengikat reseptor adalah residu asam amino yang terlibat mempertahankan integritas struktur residu pengikat reseptor serta mempengaruhi afinitas ikatan pada reseptor sel hospes. Secara struktural, terdapat 3 elemen dasar kantong pengikat reseptor VAI H5N1 yaitu α-helix (asam amino HA1 188-190), loop-130 (asam amino HA1 134-138) dan loop-220 (asam amino HA1 221-228) (Stevens et al. 2006). Residu asam amino kantong pengikar reseptor yang bersifat stabil adalah Y94, W149, H179. Sekuen stabil ini (Y94, W149 dan H179) (penomoran menurut H5) terlibat kontak secara langsung dengan residu asam sialat reseptor hospes (Stevens et al. 2006). Substitusi asam amino kantong pengikat reseptor menyebabkan perubahan pelipatan protein sehingga mempengaruhi afinitas ikatan virus pada reseptor (Harvey et al. 2004; Gambaryan et al. 2006; Auewarakul et al. 2007).
Virus influenza strain avian yang mengalami perubahan afinitas pada reseptor sel hospes berpeluang memunculkan strain virus yang mempunyai afinitas tinggi pada sel mamalia (Campitelli et al. 2006). Substitusi asam amino 129 dan 134 pada subunit HA1 dari VAI H5N1 isolat manusia menyebabkan pergeseran spesifisitas ikatan reseptor dari 2,3NeuAcGal menjadi α-2,3NeuAcGal dan α-2,6NeuAcGal (Auewarakul et al. 2007).
Residu pengikat reseptor (receptor binding sites)
Residu pengikat reseptor menentukan spesifisitas ikatan virus dengan reseptor sel hospes. Teori yang menyatakan bahwa ayam hanya mempunyai reseptor 2,3NeuAcGal dan manusia hanya mempunyai reseptor α-2,6NeuAcGal tampaknya tidak berlaku lagi. Pada sel epitel tak bersilia dan sel goblet saluran respirasi manusia predominan mempunyai asam sialat α-2,6NeuAcGal (Ibrecevich et al. 2006; Thompson et al. 2006), namun pada jaringan trakheobronkhial bersilia mengandung 2,6NeuAcGal dan α-2,3NeuAcGal meskipun dalam proporsi terbatas (Thompson et al. 2006). Pada manusia, reseptor α-2,3NeuAcGal juga dapat ditemukan pada pneumosit tipe II (Ibrecevich et al. 2006). Pada saluran respirasi dan pencernaan ayam dominan mempunyai reseptor α-2,3NeuAcGal, namun reseptor α-2,6NeuAcGal juga dapat ditemukan pada paru-paru (minor) maupun sel epitel kolon (mayor) (Kim et al.
2005). Seperti juga ayam, sel epitel kolon burung puyuh juga banyak terdapat reseptor α-2,6NeuAcGal. Hal ini menunjukkan bahwa ayam dan puyuh berpotensi sebagai hospes intermediate untuk transmisi VAI ke manusia (Guo et al. 2007).
Substitusi asam amino pada residu pengikat reseptor menyebabkan perubahan spesifisitas ikatan reseptor. Substitusi S224G dan L222Q (penomoran menurut H5) pada virus influenza strain manusia menyebabkan virus ini dapat bereplikasi pada intestinum itik (Vines et al. 1998). Bahkan disebutkan bahwa hanya dengan mutasi 1 asam amino pada residu pengikat reseptor menyebabkan perubahan spesifisitas ikatan dengan reseptor (Glasser et al. 2005; Gambaryan et al. 2006). Substitusi asam amino S223N virus HPAI H5N1 garis Asia, menurunkan afinitas pada reseptor α-2,3NeuAcGal dan memberi kemampuan virus berikatan pada α-2,6NeuAcGal secara moderat (Gambaryan et al. 2006). Hal ini menunjukkan bahwa untuk transmisi lintas spesies tidak selalu melalui reasorsi, tetapi dapat juga melalui mutasi pada residu pengikat reseptor (Harvey et al. 2004).
Virus AI yang menginfeksi manusia dapat mengalami mutasi pada residu pengikat reseptor, sehingga afinitas HA terhadap α-2,3NeuAcGal menurun dan afinitas terhadap α-2,6NeuAcGal meningkat (Harvey et al. 2004). Pergeseran spesifisitas reseptor ini terjadi pada awal setelah transmisi virus pada hospes (Matrosovich et al. 2000). Perubahan spesifisitas reseptor akibat tekanan seleksi juga terjadi pada kultur virus secara in ovo. Virus influenza strain manusia yang ditumbuhkan pada sel amnion (mempunyai reseptor 2,6NeuAcGal dan α-2,3NeuAcGal), sampai pasase ke-2 masih mempertahankan spesifisitas reseptor pada α-2,6NeuAcGal. Sementara jika dikultur pada sel alantois yang hanya mempunyai reseptor α-2,3NeuAcGal menyebabkan substitusi L222Q sehingga spesifisitas reseptor bergeser dari α-2,6NeuAcGal menjadi α-2,3NeuAcGal (Ito et al. 1997). Hal ini menunjukkan bahwa defisiensi suatu reseptor spesifik terhadap residu pengikat reseptor virus influenza merupakan tekanan seleksi yang memacu terjadinya substitusi sehingga kompatibel dengan reseptor yang dimiliki sel hospes. Hal ini dilakukan virus influenza sebagai prasyarat untuk efisiensi replikasi.
Posisi glikosilasi (N-link glycosylation sites)
Posisi glikosilasi asam amino terkait asparagin (N) dengan pola sekuen NXS dan NXT yang berpotensi sebagai tempat penempelan oligosakarida (Hoffman et al. 2005; Smith et al. 2006a; Stevens et al. 2006). Penempelan oligosakarida pada asam amino tertentu sangat penting untuk orientasi pelipatan HA untuk berikatan dengan reseptor sel atau antibodi. Posisi glikosilasi biasanya tidak stabil karena berhubungan dengan patogenesitas dan imunogenesitas. Posisi glikosilasi pada asam amino 154-156 dan 193-195 berdekatan dengan daerah antigenik dan residu pengikat reseptor, sehingga mempengaruhi afinitas ikatan pada reseptor dan mekanisme virus VAI menghindar respon imun hospes (Matrosovich et al. 1999; WHO 2005b; Campitelli et al. 2006; Gambaryan et al. 2006; Smith et al. 2006a). Penambahan posisi glikosilasi pada asam amino 84-86 merupakan mekanisme virus untuk menghindar pengenalan daerah antigenik oleh antibodi (WHO 2005b; Smith et al. 2006a). Hal ini merupakan strategi virus VAI untuk menutup atau membuka daerah antigenik dari sistem imun (Hoffmann et al 2005; Campitelli et al. 2006; Stevens et al. 2006).
Penambahan glikan pada asam amino residu pengikat reseptor menurunkan efisiensi ikatan HA pada reseptor sel (Mishin et al. 2005). Substitusi S223N dapat meningkatkan sensitivitas uji HI melalui perubahan spesifisitas reseptor dan/atau ikatan antigen-antibodi (Hoffmann et al. 2005). Penambahan glikosilasi pada protein NA juga terlibat dalam peningkatan virulensi VAI (Hulse et al. 2004).
Peptida fusi (fusion peptide)
Peptida fusi adalah peptida yang berperan pada fusi membran virus influenza saat infeksi virus ke dalam sel hospes, terdapat pada ujung N dari HA2. Peptida ini bersifat stabil pada semua influenza A, terdiri dari 23 asam amino hidrofobik kaya glisin (G). Sebelas asam amino pertama ujung N dari HA2 (GLFGAIAGFIE) lebih stabil dibandingkan 12 asam amino berikutnya. Hidrofobisitas asam amino pada peptida fusi sangat diperlukan untuk destabilisasi membran, sehingga fusi membran virus influenza dapat dilakukan dengan mudah. Substitusi glisin (G) menjadi asam amino serin (S), leusin (L) atau fenilalanin (F) secara signifikan menurunkan fusi virus influenza. Namun
substitusi asam amino pada peptida fusi menjadi glisin juga menurunkan kemampuan fusi. Hal ini menunjukkan bahwa stabilitas glisin diperlukan bukan hanya karena hidrofobisitasnya tetapi juga strukturnya (Cross et al. 2001).
Sekuen asam amino peptida fusi VAI H5N1 penyebab wabah di Asia adalah GLFGAIAGFIEGGWQGMVDGWYG. Jika dibandingkan dengan peptida fusi virus influenza subtipe H3N2 penyebab pandemi influenza di Hongkong tahun 1968 (GLFGAIAGFIENGWEGMIDGWYG), peptida fusi ini hanya mengalami substitusi 3 asam amino selama hampir 40 tahun (Cross et al. 2001; Smith et al. 2006a).
Peran Unggas Air pada Penyebaran Virus Avian Influenza
Unggas air, termasuk ordo Anseriformes (itik, entok, angsa) dan Charadriiformes (burung camar dan burung dara laut) adalah inang alami semua subtipe virus influenza A, sehingga sangat memungkinkan sebagai reservoir (penampung) virus influenza A (Olsen et al. 2006; Fouchier et al. 2007; Webster et al. 2007). Sementara, semua unggas termasuk unggas domestik (ayam, kalkun, puyuh) termasuk rentan terinfeksi. Pada inang alami, virus berada dalam keadaan seimbang dan tidak menunjukkan gejala klinis. Dalam tubuh hospes alami ini, secara evolusioner virus dalam keadaan statis, yang secara molekuler ditandai dengan rendahnya rasio substitusi N/S (Taubenberger et al. 2005). Antara hospes dengan virus terjadi toleransi yang seimbang, dimana replikasi virus secara efisien dan tidak menyebabkan penyakit. Virus bereplikasi di saluran pencernaan unggas air, sehingga ekskresi virus bersama feses dapat ditransmisikan ke unggas atau mamalia lain melalui fecal-oral (Sturm-Ramirez et al. 2004).
Penelitian di Pakistan menunjukkan bahwa 15% itik dan angsa merupakan reservoir VAI. Selain unggas air, burung liar juga dilaporkan sebagai reservoir VAI (Khawaja et al. 2005). Prevalensi VAI subtipe H5N1 di pasar unggas di Hongkong tahun 1997 paling tinggi ditemukan pada ayam (19,5%), diikuti angsa (2,5%) dan itik (2,4%) (Sortridge 1997). Sementara di pasar unggas Nanchang, Cina tahun 2000, prevalensi VAI paling tinggi dijumpai pada itik (1,3%), diikuti ayam (1,2%), puyuh (0,8%) dan merpati (0,5%) (Liu et al. 2003). Prevalensi virus HPAI subtipe H5, H7, H9 pada unggas air di Minnesota mencapai 21,5%,
sementara prevalensi subtipe H3, H4 dan H6 mencapai 63,8%. Prevalensi subtipe H5 dan H9 masing-masing sebesar 0,4% dan prevalensi subtipe H7 mencapai 0,7% (Hanson et al. 2003).
Sistem penggembalaan itik secara bebas, terutama pada saat panen padi dilaporkan juga merupakan faktor yang berperan pada penyebaran virus HPAI H5N1 (Gilbert et al. 2006). Sebanyak 27% flock itik backyard di Thailand positif terinfeksi VAI H5N1. Dan pada saat wabah VAI H5N1 pada unggas dan manusia akhir tahun 2004, 47% flock itik backyard positif terinfeksi VAI H5N1. Infeksi VAI H5N1 pada itik tersebut di atas bersifat subklinis, namun virus tetap diekskresikan bersama feses dalam waktu cukup lama (Songserm et al. 2006).
Seroprevalensi VAI pada unggas air (itik, entok dan angsa) secara signifikan lebih tinggi dibandingkan seroprevalensi pada ayam kampung. Hal ini semakin nyata terlihat pada sampling pemeriksaan terhadap itik di daerah sangat tercemar seperti daerah Jawa Barat (FKH IPB 2006).
Salah satu unggas air, yaitu Itik, dianggap sebagai sumber VAI H5N1 pada wabah di Cina tahun 2000-2004 (Chen et al. 2004; Li et al. 2004). Wabah VAI H5N1 di Hongkong tahun 2001 juga berasal dari reservoir itik dan angsa yang mengalami reasorsi dengan VAI lainnya sehingga muncul virus yang bersifat patogenik pada unggas darat (Sturm-Ramirez et al. 2004). Inokulasi 23 isolat VAI H5N1 pada itik jantan menunjukkan bahwa semua isolat VAI H5N1 dapat bereplikasi secara efisien dan 22 diantaranya ditransmisikan pada hewan peka melalui kontak (Sturm-Ramirez et al. 2005). Strain patogenik VAI H5N1 hanya menyebabkan gejala klinis ringan pada itik, tetapi tetap mengekskresikan virus bersama kotorannya yang berpotensi untuk menular ke unggas lain dan (bahkan) juga ke manusia (Kishida et al. 2005; Strurm-Ramirez et al. 2005).
Penelitian yang dilakukan Chen et al. (2004) dan Li et al. (2005) juga menunjukkan bahwa isolat VAI H5N1 dari itik sehat secara progresif dapat bereplikasi dan menyebabkan berbagai penyakit pada mencit. VAI H5N1 yang bersifat patogenik tinggi pada unggas darat, menjadi patogenik rendah jika diinokulasikan pada itik. Pada itik tidak menyebabkan gejala klinis tetapi ekskresi virus dari itik terjadi terus menerus sehingga berpotensi menyebarkan virus yang bersifat patogenik bagi unggas lain dan (bahkan) juga pada manusia (Hulse-Post et al. 2005).
Infeksi VAI sangat jarang terjadi bersifat letal pada unggas air. Wabah VAI H5N1 di Hongkong akhir tahun 2002 menyebabkan kematian pada burung migratori dan unggas air domestik termasuk itik, merupakan laporan pertama setelah tahun 1961, dimana infeksi VAI bersifat letal pada unggas air (Sturm-Ramirez et al. 2004). Virus HPAI H5N1 juga menyebabkan wabah di Danau Qinghai Cina tahun 2005 yang mematikan ribuan unggas air migratori (Zhou et al. 2006).
Transmisi VAI dari unggas air ke unggas lain kemungkinan melalui pasar unggas, dimana kontak antara unggas air dan unggas lainnya seperti ayam, puyuh, dan burung-burung lainnya tidak terhindarkan lagi (Weaver 2005; Gilbert et al. 2006). Puyuh disebutkan merupakan hospes perantara transmisi VAI dari unggas air ke unggas darat. Puyuh menyediakan lingkungan untuk adaptasi VAI subtipe H9 dari itik sehingga membentuk varian baru yang dapat menginfeksi unggas lainnya (Perez et al. 2003).
Hasil penelitian wabah VAI di Hongkong menunjukkan bahwa VAI H5N1 isolat ayam berbeda dengan isolat unggas air. VAI H5N1 isolat ayam mengalami delesi 19 asam amino pada tangkai (stalk) NA dan ada tambahan posisi glikosilasi pada HA. Delesi asam amino pada NA menyebabkan penurunan kemampuan pelepasan virus dari sel, sementara penambahan glikosilasi pada HA menyebabkan penurunan afinitas ikatan HA pada reseptor sel hospes. Perubahan NA dan HA pada ayam ini merupakan bentuk adaptasi virus dari unggas air ke unggas darat dan memperbesar peluang unggas domestik sebagai hospes intermedier pada transmisi zoonotik (Matrosovich et al. 1999)
Transmisi virus dari unggas air ke unggas darat dapat terjadi dua arah. VAI H9N2 yang awalnya berasal dari unggas air ditransmisikan ke unggas darat dan mengalami reasorsi. Virus dari unggas darat ini ditransmisikan kembali ke unggas air dan mengalami reasorsi kembali membentuk varian baru yang berpotensi untuk menginfeksi manusia secara langsung (Li et al. 2003). Hal ini menunjukkan bahwa unggas air juga berpotensi sebagai “mixing vessel” yang memunculkan varian-varian virus baru yang berpotensi sebagai penyebab pandemi influenza pada manusia (Xing et al. 2007).
Epidemiologi Virus Avian Influenza
Sejak 1959 sampai akhir tahun 2003, dilaporkan hanya terjadi 24 wabah virus influenza pada ternak unggas di seluruh dunia. Kebanyakan wabah tersebut terbatas secara geografis pada daerah tertentu, dan tidak satupun dari wabah-wabah tersebut yang ukurannya mendekati wabah H5N1 di Asia tahun 2004. Sampai saat ini, semua wabah virus HPAI disebabkan oleh virus influenza A dari subtipe H5 dan H7. Faktor utama penyebaran virus HPAI adalah perdagangan unggas hidup dan produknya serta melalui mobilitas manusia (wisatawan dan pengungsi) (WHO 2004).
Dimensi baru wabah virus HPAI mencuat di akhir tahun 2003. Dari pertengahan Desember 2003 sampai awal Februari 2004, wabah yang disebabkan oleh virus HPAI H5N1 garis Asia dilaporkan telah menyerang unggas di Korea Selatan, Vietnam, Jepang, Thailand, Kamboja, Republik Demokratik Rakyat Laos, Indonesia dan Cina (Maines et al. 2005; OIE 2005). Kejadian wabah yang serentak di banyak negara oleh virus HPAI H5N1 belum pernah terjadi sebelumnya. Virus HPAI H5N1 dijumpai pertama kali tahun 1997, merupakan hasil reasorsi dari VAI H5N1 isolat angsa domestik (A/goose/Guangdong/1/96) yang menyumbangkan gen HA dan VAI H9N2 yang menyumbangkan segmen-segmen gen penyandi protein internal (Guan et al. 1999). Meskipun beberapa genotipe pernah dilaporkan (Cauthen et al. 2000), namun genotipe “Z” telah mendominasi wabah yang terjadi sejak Desember 2003 (Li et al. 2004).
Pada bulan April 2005, untuk pertama kalinya, VAI H5N1 dapat mematikan ungas liar dalam skala besar (Zhou et al. 2006). Di danau Qinghai Barat Laut Cina, beberapa ribu unggas air migratori sakit dan mati terkena infeksi VAI H5N1 (Feare & Yasua 2006). Virus “Qinghai-like” tersebut selanjutnya menyebar ke Rusia, Eropa dan Afrika oleh burung migratori (Webster & Govorkova 2006). Kematian ternak unggas banyak dilaporkan di daerah yang berdekatan dengan danau dan rawa-rawa yang menjadi tempat singgah unggas air liar. Hal ini memperkuat dugaan bahwa unggas migratori menjadi penyebar virus, termasuk virus HPAI H5N1 garis Asia. Unggas air liar diperkirakan dapat membawa virus hanya selama masa inkubasi, atau beberapa spesies yang masih bertahan meskipun sudah terinfeksi H5N1 (Sturm-Ramirez et al. 2004).
Virus AI subtipe H5N1 dari berbagai negara, secara filogenetik terpisah menjadi 2 clade. Clade 1 adalah virus yang diisolasi pada unggas dan manusia di Kamboja, Thailand, Vietnam, Laos, Korea Selatan dan Jepang tahun 2003-2004. Clade 2 terbagi menjadi 3 subclade. Subclade 1 adalah virus dari Indonesia tahun 2004-2006 dan isolat Hongkong tahun 2003. Subclade 2 adalah isolat virus dari Rusia, Turki dan Timur Tengah tahun 2005-2006. Subclade 3 adalah isolat dari Laos, Thailand, Kamboja dan Vietnam tahun 2005-2006 (WHO 2005b; Webster & Govorkova 2006).
Virus-virus AI dalam clade dan subclade terpisah mempunyai perbedaan struktur antigenik, sehingga setiap clade atau subclade memerlukan vaksin yang berbeda. Studi pada feret menunjukkan bahwa vaksin terhadap satu clade tidak protektif terhadap clade lainnya (Webster & Govorkova 2006). Disamping struktur antigenik, antar clade dan subclade yang berbeda juga menunjukkan perbedaan sensitifitas terhadap obat antivirus influenza (Webster & Govorkova 2006). Mayoritas virus clade 1 resisten terhadap amantadin dan rimantadin, namun mayoritas virus clade 2 sensitif terhadap kedua jenis antivirus tersebut. Meskipun demikian, semua VAI H5N1 sensitif terhadap inhibitor neuraminidase (Webster & Govorkova 2006; Kandun et al. 2006).
Penyebaran VAI H5N1 secara global disebabkan oleh perdagangan unggas dan/atau produk unggas serta pergerakan unggas migratori (Capua & Marangon 2006; Chen et al. 2006). Analisis penyebaran global VAI H5N1 di Asia menunjukkan bahwa 9 dari 21 introduksi virus ke negara-negara Asia melalui perdagangan unggas atau produk unggas. Burung migratori juga berperan pada penyebaran dan introduksi VAI H5N1 ke 3 dari 21 negara-negara di Asia. Sementara introduksi VAI H5N1 pada 20 dari 23 negara di Eropa terjadi melalui unggas migratori. Di Afrika, 2 dari 8 negara mengalami introduksi VAI H5N1 melalui perdagangan unggas dan 3 dari 8 negara melalui unggas migratori (Kilpatrick et al. 2006).
Telaah Virus Avian Influenza di Indonesia
Kasus VAI H5N1 pada unggas di Indonesia muncul pertama kali pada bulan Agustus 2003 di beberapa peternakan ayam ras komersial di Jawa Barat dan Jawa Tengah. Kasus ini kemudian meluas ke berbagai daerah di Jawa
Tengah, Jawa Barat, Jawa Timur, DIY, Lampung, Bali serta beberapa daerah di Sumatera dan Kalimantan. Sampai akhir tahun 2003, kasus VAI dinyatakan endemik di 9 propinsi, terdiri dari 51 kabupaten/kota dengan jumlah kematian unggas mencapai 4,13 juta ekor (Data Dirjen Peternakan RI 2004). Jumlah kematian unggas sampai bulan November 2005 diperkirakan mencapai 10,45 juta ekor. Jumlah kematian unggas pada tahun 2005 cenderung menurun drastis dibanding tahun 2003 dan 2004, meskipun daerah yang terserang cenderung lebih luas. Hingga saat ini, virus HPAI H5N1 dinyatakan endemik di 31 dari 33 propinsi di Indonesia (Depkes 2007).
Virus AI subtipe H5N1 di Indonesia termasuk genotipe Z. Genotipe ini pertama kali ditemukan pada unggas di Cina Selatan tahun 2002 (Smith et al. 2006a). Analisis filogenetik gen HA dari VAI H5N1 dari berbagai wilayah geografi dan berbagai spesies hewan di Indonesia menunjukkan bahwa VAI H5N1 di Indonesia membentuk satu klaster (cluster), terpisah dengan isolat dari negara lain. Analisis serupa juga menunjukkan bahwa virus Indonesia mengelompok berdasarkan wilayah geografis, sehingga terbentuk 3 grup (A, B dan C). Grup A adalah kelompok virus dari kawasan tengah dan timur Indonesia, yaitu Jawa, Sulawesi Selatan dan Timor Barat. Grup B juga terdiri dari isolat virus di kawasan tengah dan timur Indonesia yaitu Jawa, Bali, Flores dan Timor Barat. Termasuk grup C adalah isolat virus dari pulau Jawa, Sumatera dan Bangka (Smith et al. 2006a).
Infeksi VAI H5N1 pada manusia mulai terjadi pada Juli 2005. Infeksi VAI H5N1 pada manusia terjadi secara sporadis dan menyerang beberapa klaster famili (Kandun et al. 2006; Sedyaningsih et al. 2007). Sejak awal infeksi sampai bulan Juni 2006, tercatat 54 kasus infeksi dan 21 infeksi diantaranya terjadi pada 7 klaster famili. Tingkat fatalitas kasus VAI H5N1 mencapai 76%, terutama menginfeksi manusia usia kurang dari 40 tahun dan 57,4% menyerang laki-laki (Sedyaningsih et al. 2007). Kasus infeksi VAI H5N1 pada klaster famili kemungkinan dipengaruhi oleh faktor genetik, tingkah laku, imunologik, dan lingkungan (Kandun et al. 2006). Semua kasus infeksi VAI H5N1 di Indonesia merupakan VAI H5N1 clade 2 subclade 1 (Kandun et al. 2006; Sedyaningsih et al. 2007).
Sampai saat ini (24 Januari 2008), dikonfirmasi sebanyak 120 kasus penularan VAI H5N1 pada manusia dan 97 diantaranya meninggal dunia (Depkes 2008; WHO 2008). Jumlah kematian manusia akibat VAI H5N1 di 12 propinsi di Indonesia ini tercatat paling tinggi di dunia (WHO 2008). Dari tahun 2003-2008 dilaporkan WHO 352 kasus penularan, dan 219 diantaranya meninggal dunia (WHO 2008). Transmisi VAI dari unggas ke manusia masih dianggap terjadi secara langsung dari unggas (ayam) atau lingkungan yang terkontaminasi virus AI (Smith et al. 2006a).