Cara Genetis untuk Menentukan
Kepekaan Bakteri terhadap Antibiotik
Pendahuluan
Dalam beberapa dasawarsa terakhir, jumlah antibiotik yang beredar di pasaran bertam-bah banyak. Selaras dengan pemakaiannya, banyak bakteri patogen beradaptasi dalam lingkungannya dan menjadi resisten terha-dap antibiotik tersebut. Selain mempersukar pengobatan penyakit, akibat lainnya adalah perubahan pola penyebab infeksi, khusus-nya infeksi nosokomial. Enterokokus yang dahulu bukan penyebab infeksi nosokomial menonjol, saat ini telah menjadi salah satu penyebab infeksi nosokomial yang perlu
1,2
diperhatikan. Contoh lainnya adalah resistensi terhadap vankomisin, yang pada periode sebelum tahun 80-an tidak pernah ditemukan, tetapi sekarang sudah banyak
3
ditemukan. Kecenderungan peningkatan resistensi bakteri yang diisolasi dari penderita terhadap berbagai antibiotik juga
4-6
ditemukan di Indonesia.
Da s a r G e n e t i k R e s i s te n s i B a k te ri terhadap Antibiotik
Berbagai penelitian menunjukkan bahwa mekanisme terjadinya resistensi bakteri terhadap antibiotik beraneka ragam, baik melalui pembentukan enzim penghancur antibiotik, penurunan aktivitas protein pengikat antibiotik, dan sebagainya. Fenotip yang tampil semuanya mempunyai dasar genetik.
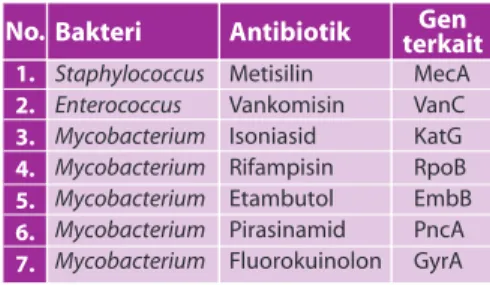
Beberapa contoh gen yang dikaitkan dengan resistensi bakteri terhadap antibiotik dapat dilihat pada tabel 1.
8
pada kodon nomor 43 gen rsplt. Pada M. leprae, M. tuberculosis, N. meningitidis, dan E. coli, mutasi pada gen rpo yang menyandi RNA polimerase terbukti menghasilkan
8,9
fenotip yang resisten terhadap rifampisin. Pada bakteri Gram negatif, bakteri Gram positif, dan mikobakteria, resistensi terhadap fluorokuinolon juga telah banyak
di-8,10,11
amati. Dalam hal ini, resistensi dikaitkan dengan kejadian mutasi pada gen penyandi DNA girase, khususnya gen gyrA yang menyandi subunit A DNA girase.
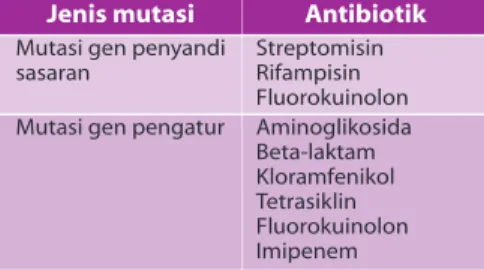
Dalam menjalankan fungsinya, gen yang menyandi protein sasaran antibiotik dipe-ngaruhi/dikendalikan oleh gen pengatur. Karena itu, fenotip yang resisten juga dapat timbul akibat mutasi pada gen pengatur tersebut. Telah diketahui, misalnya, bahwa semua bakteri Gram negatif membawa gen beta-laktamase pada kromosomnya dan bakteri akan menjadi resisten terhadap sefalosporin serta penisilin jika bakteri memproduksi enzim dalam jumlah cukup
12
besar. Khususnya pada bakteri Citrobacter, Centerobacter, Pseudomonas, dan Serratia, produksi enzim beta-laktamase di atas dapat dirangsang oleh antibiotik sefoksitin dan
13
imipenem. Enzim beta-laktamase di atas disandi oleh gen ampC dan fungsi gen ampC dikendalikan oleh aktivitas gen ampR, ampD, ampE, serta ampG. Mutasi pada ampD, misalnya, dapat menimbulkan peningkatan aktivitas ampC yang mengakibatkan peningkatan resistensi bakteri terhadap antibiotik golongan beta-laktam, kecuali imipenem yang fenotip resistensinya Pembicaraan mengenai resistensi bakteri
terhadap antibiotik akan menyangkut dua jenis bakteri:
1) Bakteri yang secara alamiah resisten terhadap antibiotik tertentu (resistensi intrinsik). Faktor genetik yang melandasinya bersifat kromosomal.
2) Bakteri yang berubah sifatnya dari peka menjadi resisten. Perubahan fenotip ini dapat terjadi karena mutasi kromosomal dan/atau didapatnya materi genetik dari luar.
Telah lama diketahui bahwa galur bakteri resisten dapat timbul lewat pemaparan bakteri dengan antibiotik dalam konsentrasi
7
tinggi untuk waktu yang lama. Pada percobaan di laboratorium, fenomena ini relatif mudah teramati pada streptomisin,
8
rifampisin, dan fluorokuinolon.
Tabel 1. Gen yang terkait dengan resistensi bakteri terhadap antibiotik
Salah satu mekanisme timbulnya fenotip resisten ini adalah mutasi gen yang menyandi protein sasaran antibiotik. Pada M. tuberculosis, resistensi terhadap streptomisin terjadi akibat subtitusi adenin oleh guanin
Agus Sjahrurachman
Bagian Mikrobiologi, Fakultas Kedokteran Universitas Indonesia, Jakarta, Indonesia
Sejalan dengan makin lamanya penggunaan antibiotik, banyak bakteri yang semula sensitif terhadap antibiotik tersebut kini menjadi resisten. Mekanisme perubahan tersebut mempunyai landasan genetis, baik melalui mutasi gen atau melalui masuknya gen asing penyandi resistensi dari luar bakteri. Kecenderungan peningkatan bakteri yang resisten ini mampu mengurangi ketepatgunaan pemakaian antibiotik untuk pengobatan penyakit infeksi. Karena itu, pengujian resistensi bakteri penyebab infeksi terhadap macam-macam antibiotik sangat diperlukan. Selama ini, pengujian tersebut dilakukan dengan memeriksa fenotip bakteri.
Dengan berkembangnya teknologi genetika molekuler, terbuka kemungkinan memakai teknologi tersebut untuk menentukan adanya gen penyandi resistensi pada bakteri. Hasil berbagai penelitian menunjukkan bahwa saat ini penggunaan teknologi tersebut secara rutin masih menjadi perdebatan, khususnya ditinjau dari sisi manfaat klinisnya. Namun, teknologi tersebut tidak diragukan lagi unggul untuk penetapan resistensi bakteri yang belum dapat dibiak in vitro atau bakteri yang lambat tumbuhnya. Selain itu, teknologi tersebut lebih aman bagi petugas laboratorium.
ABSTRAK
1. 2. 3. 4. 5. 6. 7. Bakteri Staphylococcus Enterococcus Mycobacterium Mycobacterium Mycobacterium Mycobacterium Mycobacterium No. Antibiotik Metisilin Vankomisin Isoniasid Rifampisin Etambutol Pirasinamid Fluorokuinolon Gen terkait MecA VanC KatG RpoB EmbB PncA GyrATabel 4. Dasar genetik mutasi gen yang didapat
Deteksi Gen Penyandi Resistensi
Sampai saat ini, penentuan resistensi bakteri terhadap antibiotik hampir semuanya menggunakan cara difusi cakram, pengen-ceran dalam agar kaldu, dan epsilometer. Untuk dapat melakukan pengujian di atas, terlebih dahulu bakteri harus diisolasi, dimumikan, dan selanjutnya dipaparkan pada antibiotik. Untuk antibiotik tertentu, cerminan resistensi terhadapnya juga dapat ditentukan dengan cara lain, termasuk kit komersial yang mampu mendeteksi pro-duksi enzim penghancur antibiotik dari bakteri tersebut. Tes Cefinase atau iodometri, misalnya, dapat dipakai untuk mendeteksi produksi enzim beta-laktamase yang banyak dihasilkan oleh Staphylococcus spp., Haemo-philus influenzae, dan Bacterioides spp. Cara-cara konvensional yang mencari fenotip bakteri seperti di atas mempunyai kele-mahan, khususnya dalam hal lamanya waktu sampai mendapatkan hasil. Karena itu, dicoba dikembangkan cara lain, khususnya untuk mencari faktor genotipnya.
Gen yang dicari tersebut berupa segmen DNA atau RNA. Karena jumlah salinan asam nukleat sasaran tersebut biasanya sedikit, biasanya terlebih dahulu dilakuk an amplifikasi DNA bakteri langsung dari bahan pemeriksaan dengan reaksi rantai
poli-20
merase (RRP). Selain dengan cara RRP, amplifikasi asam nukleat kadang-kadang juga dilakukan dengan cara reaksi rantai ligase, amplifikasi secara OBReplikase. Amplifikasi cara lain dilakukan dengan terle-bih dahulu membiakkan bakteri tersebut secara konvensional, lalu asam nukleatnya diekstraksi. Untuk menentukan apakah proses amplifikasi menghasilkan amplikon atau amplimer yang benar, dapat dilakukan berbagai pemeriksaan lanjutan, di antaranya adalah penentuan besar amplikon dengan cara elektroforesis, hibridisasi amplikon dengan pelacaknya, analisis restriction fragment length polymorphism (RFLP), atau
21-34
sequencing asam nukleat. puan ini secara alami hanya dimiliki oleh
beberapa jenis bakteri, seperti Streptococcus pneumoniae, Strepto co ccus viridans, Haemophilus influenzae, dan Neisseria
8,18
meningitidis. Timbulnya resistensi akibat transformasi, misalnya, teramati pada resistensi bakteri-bakteri tersebut di atas terhadap penisilin akibat pembentukan gen mosaik penyandi penicillin-binding protein
19
(PBP).
Konjugasi adalah pertukaran materi genetik antara dua bakteri akibat kontak fisik di antara keduanya. Proses konjugasi ini biasanya diinduksi oleh plasmid atau mobile genetic element yang berintegrasi pada kromosom. Plasmid sendiri, sebagai materi genetik ekstra-kromosomal, dapat memba-wa satu atau lebih gen penyandi resistensi terhadap antibiotik. Sampai saat ini, kon-jugasi dianggap sebagai salah satu meka-nisme utama penyebaran gen penyandi
8
resistensi pada bakteri.
Akhir-akhir ini, juga banyak dikupas tentang mobile DNA yang mampu mengalami trans-lokasi dari satu replikon ke replikon lain, independen dari perangkat rekombinasi sel hospes. Mobile DNA element dapat memba-wa gen-gen penyandi resistensi terhadap
8
antibiotik atau tranposon. Contohnya adalah Tn 552 (yang membawa penyandi resistensi terhadap antibiotik golongan beta-laktam), Tn 1546 (yang membawa penyandi resistensi terhadap vankomisin), Tn 5 (yang membawa penyandi resistensi terhadap kanamisin, bleomisin, strepto-misin), dan Tn 10 (yang membawa penyandi resistensi terhadap tetrasiklin).
Tabel 3. Dasar genetik resistensi akibat masuknya gen asing
Dari uraian di atas, dapat disimpulkan bahwa perubahan bakteri sensitif menjadi bakteri resisten terhadap antibiotik dapat terjadi karena mutasi preexisting genes, masuknya gen baru (yaitu gen penyandi resistensi), dan mutasi pada gen yang baru masuk (tabel 3 dan 4).
memerlukan penurunan ekspresi membran
8,14
luar yang disandi oleh gen ompD. Contoh lain resistensi yang timbul akibat mutasi pada gen pengendali adalah resistensi E. coli terhadap berbagai antibiotik sekaligus, seperti terhadap kloramfenikol, tetrasiklin, dan fluorokuinolon. Pada contoh ini, mutasi pada gen pengendali marR atau marO dapat menyebabkan peningkatan aktivitas gen marA yang menyebabkan penurunan kadar
15,16
antibiotik intrabakteri. Beberapa contoh dasar genetik resistensi yang disandi oleh gen kromosomal dapat dilihat pada tabel 2. Selain mekanisme mutasi gen kromosomal, mekanisme resistensi yang dalam tahun-tahun terakhir dianggap lebih penting adalah masuknya DNA asing ke dalam bakteri. Misalnya, yang berkaitan dengan fenomena peningkatan bakteri resisten terhadap antibiotik golongan beta-laktam, kloramfenikol, tetrasiklin, aminoglikosida, eritromisin, trimetoprim, dan vankomisin. Dalam hal ini, DNA asing dapat masuk ke dalam bakteri melalui tiga cara, yaitu transduksi, transformasi, dan konjugasi. Tabel 2. Dasar genetik resistensi bakteri akibat mutasi kromosomal
Transduksi adalah pemindahan DNA dari satu bakteri ke bakteri lainnya melalui infeksi bakteri oleh bakteriofaga. Pada saat infeksi, DNA bakteriofaga berintegrasi dengan DNA bakteri dan saat replikasi sebagian dari DNA bakteri terakit pada DNA bakteriofaga. Saat bakteriofaga menginfeksi bakteri lain, terjadilah pemindahan DNA tersebut. Jika dalam DNA yang dipindahkan terdapat gen penyandi resistensi, bakteri yang terinfeksi oleh bakteriofaga tadi akan menjadi resisten terhadap antibiotik tertentu. Fenomena tim-bulnya resistensi bakteri melalui transduksi, misalnya, teramati pada penyebaran resis-tensi Staphylococcus aureus terhadap anti-biotik golongan beta-laktam dan logam
17
berat.
Transformasi adalah proses pemasukan DNA bebas dari lingkungan ke bakteri.
Kemam-Jenis mutasi
Mutasi gen penyandi sasaran
Mutasi gen pengatur
Antibiotik Streptomisin Rifampisin Fluorokuinolon Aminoglikosida Beta-laktam Kloramfenikol Tetrasiklin Fluorokuinolon Imipenem
Gen yang didapat dari luar Antibiotik
Gen-gen penyandi aminoglikosidase Gen-gen penyandi beta-laktamase Gen-gen penyandi chloramphenicol acetyl transferase (CAT)
Gen MLS Gen mecA
Gen-gen penyandi tetrasiklin Gen penyandi ligase abnormal dan gen-gen penyerta Aminoglikosida Beta-laktam Kloramfenikol Eritromisin Metisilin Tetrasiklin Vankomisin Jenis mutasi
Mutasi gen penyandi sasaran
Mutasi gen pengatur
Gen yang dihasilkan
Extended-spectrum beta-lactamase Extended-spectrum beta-lactamase Gen penyandi resisten metisilin (mec)
Konfirmasi amplikon dengan elektroforesis saat ini merupakan cara tersering untuk
21
mengidentifikasi gen penyandi resistensi. Uji ini spesifik jika bahan pemeriksaan tidak mengandung mikroba lain yang mengan-dung rantai asam nukleat serupa dan RRP menghasilkan amplikon dalam jumlah yang cukup. Jika menggunakan dua jenis atau lebih primer yang sasarannya berbeda, teknik RRP multipleks ini sekaligus dapat dipakai untuk menentukan adanya gen penyandi resistensi dan identifikasi bakteri. RRP multipleks ini, misalnya, telah dicobakan untuk mendeteksi gen mecA pada kuman Staphylococcus yang terlebih dahulu
diper-21,29,35,36
banyak dengan cara konvensional. Hasilnya menunjukkan bahwa cara RRP multipleks ini lebih sensitif dibandingkan dengan cara konvensional dalam hal mendeteksi resistensi yang disandi oleh gen mecA untuk selain Staphylo-coccus aureus. Selain itu, teknik ini juga telah dicobakan pada biakan darah, hanya saja perlu dipertimbangkan kemungkinan adanya penghambat RRP dalam bahan, seperti antikoagulan natrium polianetol
37,38
sulfonat.
Untuk konfirmasi dengan cara RFLP, sebelum elektroforesis amplikon, terlebih dahulu dilakukan fragmentasi dengan enzim endonuklease restriksi, suatu enzim yang hanya memecah DNA di tempat spesifik. Untuk memilih jenis enzim yang dipakai, diperlukan data rangkaian basa penyusun gen sasaran amplifikasi. Pita-pita potongan DNA yang terlihat pada hasil elektroforesis selanjutnya dibandingkan dengan bakunya. Teknik ini, misalnya, telah dipakai untuk mendiferensiasi gen vanA, vanB, dan vanC yang menyandi resistensi enterococcus
24
terhadap vankomisin. Selain itu, teknik ini telah pula dipakai untuk mendeteksi mutasi gen M. tuberculosis penyebab resistensi terhadap isoniazid dan streptomisin serta untuk mengklasifikasikan gen penyandi extended beta-lactamase pada bakteri Gram
20, 26, 27, 39-45
negatif.
Konfirmasi adanya gen penyandi resistensi pada bakteri juga dilakukan dengan teknik hibridisasi. Teknik ini sangat spesifik meskipun mempunyai kelemahan, yaitu keharusan tersedianya pelacak. Teknik ini telah dicobakan untuk mendeteksi gen
35,36
mecA pada .
staphylococcus
staphylococcus
Mutasi gen yang menyebabkan resistensi juga telah dicoba dengan teknik analisis universal heteroduplex generator-RRP untuk
30
gen rpoB pada M. tuberculosis. Dalam hal ini, DNA bakteri yang diuji dan bakteri kontrol didenaturasi dan dibiarkan berhibridisasi ulang secara bersamaan di tempat yang sama. Mutasi yang terjadi menyebabkan ketidaksesuaian pasangan basa saat hibri-disasi heterodupleks, membentuk hibrid berupa DNA dengan bubble. DNA untai ganda dengan bubble tersebut akan mem-bentuk pita yang berbeda dengan DNA bakunya jika dielektroforesis dengan gel yang beresolusi tinggi.
Teknik lain yang lebih mudah pembacaan-nya adalah kombinasi RRP dan immunoassay, atau dikenal dengan nama RRPLiPA (RRP-line probe assay). Pada teknik ini, DNA hasil amplifikasi RRP yang telah diberi penanda biotin, dihibridisasikan dengan DNA pelacak yang telah diimobilisasi. Adanya hibridisasi selanjutnya dideteksi dengan substrat kromogen. Teknik ini telah dicobakan untuk mendeteksi mutasi gen M. tuberculosis yang
29
resisten terhadap rifampisin. Teknik yang menyerupai RRP-LiPA adalah teknik analisis
(20)
rangkaian molecular beacon-RRP . Dalam teknik terakhir, DNA pelacak yang telah ter-tandai oleh fluorofor akan mengalami peru-bahan konformasional jika berhibridisasi dengan DNA sasaran dan mengaktifkan fluorofor berfluorosensi.
Perbandingan Uji Resistensi Cara Konvensional dan Genetis
Pemanfaatan teknik deteksi gen penyandi resistensi saat ini belum luas. Selain lebih rumit dan masih lebih mahal dibandingkan teknik konvensional, juga karena tidak semua fenotip resistensi hanya bergantung pada (satu) gen saja. Misalnya, dalam hal resistensi terhadap penisilin, bakteri dapat menjadi resisten akibat ekspresi gen mecA yang menyandi protein pengikat penisilin atau oleh gen lain yang menyandi protein beta-laktamase. Dengan demikian, jelas bahwa bakteri yang tidak membawa gen mecA masih mungkin resisten terhadap penisilin.
Dibandingkan dengan teknik konvensional, teknik genetis mempunyai potensi
keung-20
gulan berupa:
·Dapat dilakukan langsung pada bahan pemeriksaan tanpa terlebih dahulu Konfirmasi amplikon yang terbaik adalah
dengan DNA sequencing. Dibandingkan dengan cara lainnya, teknik ini memerlukan perangkat yang mahal dan lebih kompleks pengerjaannya. Dengan dikembangkannya automatisasi, teknik ini menjadi lebih murah
29
dan lebih cepat memberikan hasil. Teknik ini telah dilakukan untuk mendeteksi mutasi gen yang menimbulkan fenotip resistensi M. tuberculosis terhadap rifampisin, isoniazid, etambutol, streptomisin, dan
pirazina-27,28,31-33,46
mid.
Cara deteksi lain yang relatif baru adalah cara assay bDNA. Teknik ini berbeda dengan teknik RRP karena yang diamplifikasi bukan DNA sasaran dan amplifikasinya terjadi melalui proses hibridisasi bertingkat. Ringkasnya, DNA penangkap yang terfiksasi pada fase padat akan berhibridisasi dengan DNA sasaran. Di tempat lainnya, DNA sasaran akan berhibridisasi dengan fragmen DNA pelacak, kemudian fragmen lain DNA pelacak berhibridisasi dengan DNA penguat. Akhirnya, keberadaan DNA penguat dide-teksi dengan cara chemiluminescence atau reaksi enzimatik dengan substrat kromo-genik. Teknik ini telah dicobakan untuk deteksi gen mecA pada
dengan hasil yang sebanding dengan cara
22,23
amplifikasi RRP.
Deteksi mutasi gen yang menimbulkan fenotip resisten juga dapat dilakukan dengan menguraikan amplikon dari DNA untai ganda menjadi DNA untai tunggal sebelum dilakukan elektroforesis. Pita-pita DNA yang tampak dibandingkan dengan bakunya. Teknik ini dikenal sebagai teknik gabungan RRP dan SSCP (single-strand conformation polymorphism) dan telah digunakan untuk mengidentifikasi mutasi gen penyandi extended beta-lactamase pada beberapa bakteri famili Enterobacteriaceae, gen yang menimbulkan resistensi M. tuberculosis terhadap isoniazid, rifampisin,
31,34,42-44,47-52
etambutol, dan fluorokuinolon.
Salah satu pengembangan teknik ini dikenal dengan nama cleavage fragment length polymorphism (CFLP). Dalam teknik ini, pasca-denaturasi amplikon, fragmen DNA diuraikan oleh enzim endonuclease cleavage yang mengurai DNA pada pertemuan untai tunggal dan dupleksnya. Setelah dielektro-foresis pada gel pendenaturasi, polimor-fisme yang unik terjadi selaras dengan
20
mutasinya.
melalui proses isolasi bakteri sehingga dapat mempercepat hasil pemeriksaan. Hal ini akan sangat bermanfaat jika diterapkan untuk kasus infeksi gawat yang memerlukan intervensi antibiotik segera atau pada bakteri yang belum berhasil dibiak in vitro, seperti M. leprae.
·Penilaian langsung pada potensi bakteri menjadi resisten dan tidak pada fenotip-nya. Ini terutama berkaitan dengan kuman yang lambat tumbuhnya sehingga pe-nampilan fenotip lebih sukar diamati. Selain itu dengan teknik genetis dapat dihindari adanya kemungkinan false resistance akibat pengaruh artifisial media pertumbuhan bakteri pada fenotip resis-tensi.
·Karena tidak memerlukan pembiakan bakteri, teknik ini jelas lebih aman bagi petugas dari kemungkinan terjadinya infeksi laboratorik.
Di samping berbagai keuntungan di atas, teknik genetis juga mempunyai dampak
20
negatif:
·Untuk tiap antimikroba, diperlukan uji yang spesifik. Hal ini berkaitan dengan perbedaan gen yang akan dideteksi. ·Dibandingkan dengan cara konvensional,
deteksi secara genetis pada dasarnya hanya mendeteksi gen dan kurang komprehensif. Hal ini berkaitan dengan fakta bahwa fenotip resistensi dapat disebabkan oleh banyak gen berbeda atau mutasi yang berbeda. Selain itu, belum semua mekanisme resistensi diketahui latar belakang genetisnya. Lebih lanjut, cara genetis tidak tepat jika dikerjakan untuk mencari fenomena resistensi yang baru.
·Pada saat ini, teknik genetis masih memerlukan perbaikan agar deteksi dapat dilakukan dengan jumlah gen yang makin kecil; sensitivitas cara genetis belum optimal jika dalam sediaan hanya terdapat beberapa bakteri.
·Teknik genetis mampu mendeteksi gen penyandi resistensi yang secara klinis belum tentu relevan, misalnya untuk gen yang tingkat ekspresinya rendah.
·Pengerjaannya harus hati-hati karena kontaminasi yang kecil sekalipun akan teramplifikasi dan dapat menyebabkan timbulnya hasil yang salah.
·Pada saat ini, pembakuan teknik genetis masih sangat minim.
DAFTAR PUSTAKA
1. Murray BE. The life and times of enterococcus. Clin Microb Rev. 1990; 3:46-65.
2. Schaberg DR, Culver DH, Gaynes RP. Major trends in microbial etiology of nosocomial infection. Am J Med. 1991; 92:72S-5S.
3. Center for Diseases Control and Prevention. Nosocomial enterococci resistant to vancomycin -United States, 1989-1993. MMWR. 1993; 42:597-9.
4. Sjahrurachman A. Resistensi kuman saluran pernapasan dan pus terhadap berbagai antibiotik di Bagian Mikrobiologi Fakultas Kedokteran Universitas Indonesia. Maj Kes Masy Indon. 1996; 8:521-5.
5. Sjahrurachman A. Resistensi bakteri terhadap aminoglikosida. Cermin Dunia Kedokt. 1996; 108:49-53. 6. Usman CW, Suharno J, Santoso AUS, et al. Aktivitas antibakteri sefuroksim dan antibiotik oral lain terhadap
kuman penghasil beta laktamase. Mikrobiol. Min. Indo. 1992; 6: 206-8. 7. Bryson V, Szibalsky W. Microbial selection. Science 1952; 11:45-51.
8. Rice LB, Bonomo RA. Genetic and biochemical mechanisms of bacterial resistance to antimicrobial agents In: Lorian, V (ed). Antibiotics in Laboratory Medicine. 4th ed. Maryland: Williams & Wilkins Co. 1996; pp.453-501. 9. Carter PE, Abadi JFR, Yakubu DE, Pennington TH. Molecular characterization of rifampicin resistant Neisseria
meningitidis. Antimicrob. Agents Chemother. 1994; 38:1256-61.
10. Yoshida H, Bogaki M, Nakamura M, Nakamura S. Quinolone resistance determining region in the DNA gyrase gyr A gene of Escherichia coli. Antimicrob. Agents Chemother. 1990; 34:1271-72.
11. Zhanel GG, Karlowsky JA, Sauders MH, et al. Development of multiple antibiotic resistant (mar) mutants of
Pseudomonas aeruginosa after serial exposure to fluoroquinolones. Antimicrob. Agents Chemother. 1995;
39:489-95.
12. Bergstrom S, Olsson O, Normark S. Common evolutionary origin of chromosomal beta lactamase genes in enterobacteria. J. Bacteriol. 1982; 150:528-34.
13. Bennett PM, Chopra I. Molecular basis of beta lactamase induction in bacteria. Antimicrob. Agents Chernother. 1993; 37:153-8.
14. Yoneyama H, Nakae T. Mechanism of efficient elimination of protein D2 in outer membrane of imipenem resistant Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1993; 37:2385-90.
15. Ariza RR, Cohen SB, Bachahawat N, et al. Repressor mutations in the marRAB operon that activate oxidative stress gene and multiple antibiotic resistance. J. Bacteriol. 1994; 176:143-8.
16. Cohen SP, Hachler H, Levy SB. Genetic and functional analysis of the multiple antibiotic resistance (mar) locus in Escherichia coli. J. Bacteriol. 1993 ; 175:1484-92.
17. Lyon BR, Skurray R. Antimicrobial resistance in Staphylococcus aureus Genetic basis. Microbiol. Rev. 1987; 51: 88-134.
18. Davis BD, Gene variation and transfer. In: Davis BD, Dulbecco R, Eisen HN, Ginsberg HS (eds). Microbiology. Philadelphia, USA. 1990; pp.123-42.
19. Grebe T, Hakenbeck R, Penicillin-binding protein 2b and 2x of Streptococcus pneumoniae are primary resistance determinants for different classes of beta lactam antibiotics. Antimicrob. Agents Chemother. 1996; 40:829-34.
20. Cockerill FR II. Genetic methods for assessing antimicrobial resistance. Antimicrob. Agents. Chemother. 1999; 43:199-212.
21. Geha DJ, Uhl JR, Gustafero CA, Persing DH. Multiplex PCR for identification of methicillin-resistant staphylococci in the clinical laboratory. J Clin Microb. 1994;1768-72.
22. Kolbert CP, Varga-Demore P, Arruda J, et al. A novel branched DNA assay for detection of mecA gene in oxacillin resistant and oxacillin susceptible staphylococci. J. Clin. Microb. 1998; 36:2640-4.
23. Zheng X, Kolbert CP, Delmore P, et al. Direct mecA detection from blood cultures bottles by branched DNA (bDNA) signal amplification. Abstract of the 37 Meeting of International Conference on Antimicrobial Agents and Chemotherapy. Toronto, Canada. 1997;p.87.
24. Patel R, Uhl J, Kohner P, et al. Multiplex PCR detection of vanA, vanB, vanC-1 and vanC2/3 in enterococci. J. Clin. Microbiol. 1997; 35:703-7.
25. Patel R, Uhl J, Kohner P, et al. DNA sequence variation within vanA, vanB, vanC-1 and vanC2/3 genes of clinical Enterococcus isolates. Antimicrob. Agents. Chemother. 1998; 42:202-5.
26. Martha HJ, Soini P, Huovinen V and Vijanen MK. katG mutations in isonazid resistant Mycobacterium
tuberculosis isolates recovered from Finnish patients. Antimircob. Agents. Chemother. 1996; 40:2187-9.
27. Musser JM, Kapur V, Williams DL, et al. Characterization of the catalaseperoxidase gene (katG) and inhA locus in isoniazid-resistant and susceptible strains of Mycobacterium tuberculosis by automated DNA sequencing : restricted array of mutations associated with drug resistance. J. Infect. Dis. 1996; 713:196-202.
28. Sreevatsan S, Pan X, Zhang Y, et al. Analysis of the oxyR-aphC region in isoniazid-resistant and susceptible
Mycobacterium tuberculosis complex organisms recovered from diseased humans and animals in diverse
localities. Antimicrob. Agents. Chemother. 1997; 41: 600-6.
29. Rossau, Traore H, de Beenower H, et al. Evaluation of INNO-LIPA Rif TB assay, a reverse hybridization assay simultaneus detection of Mycobacterium tuberculosis complex and its resistance to rifampicin. Antimicrob. Agents. Chemother. 1997 ; 41:2093-8.
30. Williams DL, Waguespack C, Gillis TP, et al. Characterization of rifampin resistance in pathogenic mycobacteria. Antimicrob. Agents. Chemother. 1994; 38:2380-6.
31. Sreevatsan S, Pan X, Stockbauer KE, et al. Ethambutol resistance in Mycobacterium tuberculosis: Critical role of embB mutations. Antimicrob. Agents. Chemother. 1997; 41:1677-81.
32. Sreevatsan S, Pan X, Stockbauer KE, et al. Characterization of rspL and rrs mutations in streptomycin-resistant
Mycobacterium tuberculosis isolates from diverse geographic localities. Antimicrob. Agents. Chemother. 1996;
33. Sreevatsan S, Pan X, Kreiswirth BN and Musser JM. Mutations associated by pyrazinamide resistance in pncA of Mycobactecium tuberculosis complex organisms. Antimicrob. Agents. Chemother. 1997; 41:636-40.
34. Takiff HE, Salazar L, Guerrero C, et al. Cloning and nucleotide sequence of Mycobacterium tuberculosis gyrA and gyrB genes and detection of quinolone resistance mutations. Antimicrob. Agents. Chemother. 1994; 38:773-80.
35. Kolbert CP, Connolly JE, Lee MJ, Pershing DH. Detection of the Staphylococcal mecA gene by chemiluminescent DNA hybridization. J. Clin. Microb. 1995; 33:2179-82. 36. Mulde GJ. Comparison of disk diffussion, E test and detection of mecA for determination of methicillin resistance in coagulase-negative Staphylococci. Eur J. Clin. Microb.
Infect. Dis. 1996; 15:567-73.
37. Ubakata K, Nakagami S, Nitta A, et al. Rapid detection of niecA gen in methicillin resistant Staphylococci by enzymatic detection of polymerase chain reaction product. J. Clin. Microbiol. 1992; 30:1728-33.
38. Fredrick DN, Relman DA. Improved amplification of microbial DNA from blood cultures by removal of the PCR inhibitor sodium polyanetholesulfonate. J. Clin. Microbiol. 1998: 36:2810-6.
39. Brow MA, Olderbur MC, Lyamichev V. et. al. Differentiation of bacterial 16SRNA genes and intergenic regions in Mycobacterium tuberculosis katG genes by structure-specific endonuclease cleavage. J. Clin. Microb. 1996; 34:2139-42.
40. Cockerill FR III, Uhl JR, Temezgen Z. et. al. Rapid identification of a point mutation of the Mycobacterium tuberculosis catalase-peroxidase (katG) gene assoiciated with isoniazid resistance. J. Infect. Dis. 1995; 171:240-5.
41. Haas WH, Schilke K, Brand J. et al. Molecular analysis of katG gene mutation in strains of Mycobacterium tuberculosis complex from Africa. Antimicrob. Agents. Chemother. 1997; 41:1601-3.
42. Heym B, Alzari PM, Honore N, Cole ST. Missense mutation in the catalase-peroxidase gene, katG, are associated with isoniazid resistance in Mycobacterium tuberculosis. Mol. Microbiol. 1995; 15:235-45.
43. Rouse DA, Zhongming L, Bai GH, Morris SL. Characterization of katG and inhA genes of isoniazid resistant clinical isolates of Mycobacterium tuberculosis. Antimicrob. Agents. Chemother. 1995; 39:2472-7.
44. Victor TC, Pretorius GS, Felix JV, et al. KatG mutation in isoniazid resistant strains of Mycobacterium tuberculosis are not infrequent. Antimicrob. Agents. Chemother. 1996; 40:1572 (Letter).
45. Nuesch-Inderbinen MT, Hacher H and Ksyser FH. Detection of genes coding for extended spectrum beta SHV-lactamase in clinical isolates by molecular genetic method and comparison with E test. Eut J. Clin. Microb. Infect. Dis. 1996; 15:398-402.
46. Kapur V, Li LL, Iordanescu S. et al. Characterization by automated DNA sequencing of mutations in the gene (rpoB) encoding RNA polymerase beta sub unit in rifampin-resistant Mycobacterium tuberculosis strains from New York City and Texas. J. Clin Microbiol. 1994; 32:1095-8.
47. M'Zali FH, Heritage J, Gasoyne-Binzi DM. et. al. PCR single-strand conformational polymorphism can be used to detect the gene encoding SHV-7 extended spectrum beta lactamase and to identify different SHV genes within the same strain. Antimicrob. Agents Chemother. 1998; 41:123-5.
48. Telenti A, Honore N, Bernasconi C, et al. Genotypic assessment of isoniazid and rifampicin resistance in Mycobacterium tuberculosis: a blind study at reference laboratory level. J. Clin. Microbiol. 1997; 35:719-23.
49. Felmlee TA, Liu Q, Whelen AC. et. al. Genotype detection of Mycobacterium rifampicin resistance : Comparison of single strand poly-morphism and dideoxy fingerprinting. J. Clin. Microb. 1995; 33:1617-23.
50. Telenti A, Imboden P, Marchesi F, et al. Detection of rifampicin resistance mutations in Mycobacterium tuberculosis. Lancet 1993; 341:647-50.
51. Telenti A, Imboden P, Marchesi F, et al. Direct automated detection of rifampicin-resistant Mycobacterium tuberculosis by polymerase chain reaction and single strand conformation polymorphism analysis. Antimicrob. Agents Chemother. 1993; 37:2054-8.
52. Whelen AC, Femlee TA, Hunt JM. et. al. Direct genotypic detection of Mycobacterium tuberculosis rifampicin resistance in clinical specimens by using single tube heminested PCR. J. Clin. Microbiol. 1995; 33:556-61.