IDENTIFIKASI KERAGAMAN GEN β-LAKTOGLOBULIN
PADA SAPI
FRIESIAN HOLSTEIN
DI BALAI
PENGEMBANGAN PERBIBITAN TERNAK
SAPI PERAH CIKOLE, LEMBANG
SKRIPSI
HENTI SYLVIA NURY
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR 2010
RINGKASAN
HENTI SYLVIA NURY. D14063554. 2010. Identifikasi Keragaman Gen β-laktoglobulin pada Sapi Friesian Holstein di Balai Pengembangan Perbibitan Ternak - Sapi Perah Cikole, Lembang. Skripsi. Departemen Ilmu Produksi dan Teknologi Peternakan. Fakultas Peternakan. Institut Pertanian Bogor
Pembimbing utama : Prof. Dr. Ir. Cece Sumantri M. Agr. Sc Pembimbing Anggota : Ir. Anneke Anggraeni M.Si., Ph.D
Kualitas susu sangat dipengaruhi oleh komposisi dan kadar protein yang dikandungnya. Protein dalam susu sapi terdiri dari dua jenis yaitu kasein dan whey. Protein utama dalam whey adalah β-laktoglobulin. Upaya perbaikan kualitas protein dapat dilakukan melalui pendekatan seleksi menggunakan penciri genetik pada tingkat DNA. Penelitian ini bertujuan untuk mengidentifikasi keragaman gen β-laktoglobulin terhadap materi berupa DNA ektsraksi menerapkan metode Polymerase Chain Reaction - Restriction Fragmen Length Polymorphism (PCR-RFLP). DNA ekstraksi berasal dari sampel darah diambil dari secara vena jugularis bersumber dari 88 ekor sapi perah induk laktasi Friesian Holstein (FH) di BPPT-SP Cikole, Lembang. Penelitian genotyping gen β-laktoglobulin ini berlangsung selama enam bulan di Laboratorium Genetika Ternak, bagian Genetika dan Molekuler Ternak, Fakultas Peternakan, Institut Pertanian Bogor. Frekuensi genotipe dan alel serta nilai heterozigositas dihitung menggunakan metode Nei (1987). Kesetimbangan alel dan genotipe berdasarkan hukum Hardy-Weinberg dihitung menerapkan uji chi-square.
Genotyping gen β-laktoglobulin pada sapi FH bersifat polimorfik atau beragam. Ada dua tipe alel yang dideteksi, yaitu alel A dan B yang menghasilkan tiga tipe genotipe, yaitu AA, AB dan BB. Frekuensi alel A dan B adalah 0,40 dan 0,60. Nilai heterozigositas yang diperoleh sebesar 0,483. Hasil uji chi-square menunjukkan bahwa χ2 lebih kecil dari χ2 tabel yang artinya gen β-laktoglobulin dari sapi FH pengamatan berada dalam keseimbangan Hardy-Weinberg, dengan varian genotipe untuk AA, AB dan BB berurutan adalah 0,10, 0,60, dan 0,30.
ABSTRACT
Identification on the Polymorphism of the β-lactoglobuline Gene in Holstein-Friesian Cows in BPPT-SP Cikole. Lembang
Nury, H.S., C. Sumantri, and A. Anggraeni
Milk quality in dairy cattle is strongly influenced by the composition and the content of its protein. Proteins in dairy cow mainly consist of two types, namely casein and whey. The main protein in whey is β-lactoglobuline. Improving protein quality can be afforded through selection approaches using genetic markers at the DNA level. This study was aimed to identify polymorphims of the β-lactoglobuline gene from the collected DNA extractions by using Polymerase Chain Reaction - Restriction Fragment Length Polymorphism (PCR-RFLP) methode. Those of DNA extractions were from blood samples taken via vena jugularis from a total of Holstein Friesian (HF) lactating cows of 88 heads at BPPT - SP Cikole, Lembang, West Java. Genotyping process of the β-lactoglobuline was carried out for six months at the Laboratory of Animal Genetics, the Molecular Genetics and Animal Husbandry, Faculty of Animal Science, Bogor Agricultural University. Genotype and allele frequencies and heterozygosity values were calculated by Nei Methode (1987). The Hardy - Weinberg equilibrium of the allele and genotype frequencies was calculated by the Chi-square test. Genotyping the β-lactoglobuline gene in HF cows was polymorphic. Two types of alleles were detected, namely the A and B resulting three types of genotypes, namely AA, AB and BB. The frequencies of the A and B alleles were 0.40 and 0.60 respectively. The heterozygosity value was obtained at 0.483. Chi-square test resulted that χ2 calculation was less than χ2 table meaning the β-lactoglobuline gene in HF observations in the equilibrium, with the variant genotypes of the AA, AB and BB genotypes were respectively 0.10, 0.60, and 0.30.
IDENTIFIKASI KERAGAMAN GEN β-LAKTOGLOBULIN
PADA SAPI
FRIESIAN HOLSTEIN
DI BALAI
PENGEMBANGAN PERBIBITAN TERNAK
SAPI PERAH CIKOLE, LEMBANG
HENTI SYLVIA NURY D14063554
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR 2010
Judul : Identifikasi Keragaman Gen β-laktoglobulin pada Sapi Friesian- Holstein di Balai Pengembangan Perbibitan Ternak Sapi Perah Cikole, Lembang
Nama : Henti Sylvia Nury NIM : D14063554
Menyetujui,
Pembimbing Utama Pembimbing Anggota
(Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc.) (Ir. Anneke Anggraeni, M.Si., Ph.D) NIP. 19591212 198603 1 004 NIP. 19630924 199803 2 001
Mengetahui Ketua Departemen,
Ilmu Produksi dan Teknologi Peternakan
(Prof. Dr. Ir. Cece Sumantri, M. Agr. Sc.) NIP. 19591212 198603 1 004
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 6 November 1987 di Medan, Sumatera Utara. Penulis adalah anak ketiga dari 5 bersaudara dari pasangan bapak Drs. H. Maraenda Hrp SH,MH dan Ibu Dra. Hj. Pitta Hara Siregar.
Pendidikan dasar diselesaikan pada tahun 2000 di SD Negeri 163099 Tebing Tinggi. Pendidikan lanjutan menengah pertama diselesaikan pada tahun 2003 di SLTP Negeri 27 Medan, dan pendidikan lanjutan menengah atas diselesaikan pada tahun 2006 di SMA Negeri 8 Medan, Sumatera Utara.
Penulis diterima sebagai mahasiswa departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan Institut Pertanian Bogor melalui jalur Ujian Seleksi Masuk IPB (USMI) pada tahun 2006. Selama mengikuti pendidikan, penulis aktif di Lembaga Dakwah Fakultas FAMM Al-An’aam, Fakultas Peternakan IPB periode 2007/2008 DAN 2008/2009, HIMAPROTER (English Club) Fakultas Peternakan IPB periode 2007-2008. Penulis pernah menjadi asisten mata kuliah Pendidikan Agama Islam pada tahun ajaran 2008/2009, dan 2009/2010. Penulis juga pernah terlibat dalam Kepanitiaan Open House Asrama 44 periode 2007/2008, SALAM ISC tahun 2008 divisi Muslim Generation Inside (MER-C) dan Masa Perkenalan Fakultas (MPF) sebagai staf divisi MEDIS pada tahun 2008.
KATA PENGANTAR
Puji dan syukur senantiasa dipanjatkan kehadirat Allah SWT, karena atas Taufik dan RahmatNya, penelitian dan penulisan skripsi ini dapat terselesaikan. Skripsi yang berjudul Identifikasi Keragaman Gen β-laktoglobulin pada Sapi Perah Friesian Holstein di Balai Pengembangan Perbibitan Ternak Sapi Perah Cikole, Lembang merupakan salah satu syarat untuk memperoleh gelar sarjana pada Fakultas Peternakan Institut Pertanian Bogor.
Upaya dalam peningkatan kualitas susu dapat dilakukan dengan perbaikan karakteristik genetik pada kualitas susu. Keragaman genetik pada protein susu telah diketahui terdapat hubungan yang signifikan antara variasi genotipe gen protein susu dengan kadar protein susu dan secara ekonomi penting dalam suatu peternakan sapi perah. Gen-gen tersebut dapat digunakan sebagai penanda molekuler tingkat DNA untuk seleksi terhadap karakter kualitas susu sapi secara cepat dan tepat.
Penelitian ini bertujuan untuk memberikan informasi mengenai keragaman gen β-laktoglobulin pada sapi FH (Friesian Holstein) di BPPT-SP Cikole, Lembang. Penulis berharap semoga skripsi menjadi langkah awal untuk berkarya di dunia pendidikan dan dapat memberikan kontribusi bagi kemajuan dunia peternakan khususnya di Indonesia. Amin.
Bogor, Juli 2010
DAFTAR ISI Halaman RINGKASAN ... ii ABSTRACT ... iii LEMBAR PERNYATAAN ... iv LEMBAR PENGESAHAN ... v RIWAYAT HIDUP ... vi
KATA PENGANTAR ... vii
DAFTAR ISI ... viii
DAFTAR TABEL ... x
DAFTAR GAMBAR ... xi
DAFTAR LAMPIRAN ... xii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan ... 1
TINJAUAN PUSTAKA ... 2
Sapi FH dan Produksi Susunya ... 2
Seleksi dengan Marked Assisted Selection (MAS) ... 3
Protein Susu ... 3
Struktur gen β-laktoglobulin ... 4
Keragaman gen β-laktoglobulin ... 5
Metode PCR-RFLP ... 5
Elektroforesis ... 6
Keragaman Genetik ... 7
METODE ... 8
Lokasi dan Waktu ... 8
Materi ... 8
Sampel ... 8
Primer ... 8
Polymerase Chain Reaction- Restriction Fragment Length Polymorphism ... 8
Elektroforesis ... 9
Prosedur ... 9
Pengambilan Sampel Darah ... 9
Polymerase Chain Reaction- Restriction Fragment Length Polymorphism... 10
Elektroforesis ... 11
HASIL DAN PEMBAHASAN ... 13
Amplifikasi Gen β-laktoglobulin ... 13
Pendeteksian Keragaman Gen β-laktoglobulin ... 14
Frekuensi Gen β-laktoglobulin pada sapi FH ... 16
Distribusi Genotipe Gen β-laktoglobulin ... 17
KESIMPULAN DAN SARAN ... 19
Kesimpulan ... 19
Saran ... 19
UCAPAN TERIMA KASIH ... 20
DAFTAR PUSTAKA ... 21
DAFTAR TABEL
Nomor Halaman
1. Komposisi Susu beberapa Bangsa Sapi Perah ... 2 2. Komposisi Protein Susu Ternak Ruminansia ... 4 3. Keragaman Gen β-laktoglobulin dari Beberapa Penelitian
pada Ternak Ruminansia ... 5 4. Sekuens Primer ... 8 5. Nilai Frekuensi Genotipe, Frekuensi Alel dan Nilai
Heterozigositas Lokus Hae III pada sapi Friesian Holstein ... 16 6. Distribusi Genotipe Gen β-laktoglobulin pada Sapi FH ... 17
DAFTAR GAMBAR
Nomor Halaman
1. Struktur Gen β-laktoglobulin ... 4
2. Hasil Amplifikasi Gen β-laktoglobulin pada Sapi FH ... 13
3. Sekuens Gen β-laktoglobulin Bos taurus pada Ekson 4 ... 14
4. Pola Pita Gen β-laktoglobulin Sapi FH ... 15
5. Perbedaan sekuen Gen β-laktoglobulin nomor akses GenBank X14710 yang disebabkan mutasi titik C – T ... 16
DAFTAR LAMPIRAN
Nomor Halaman
1. Sekuens Gen β-laktoglobulin berdasarkan Akses
PENDAHULUAN Latar Belakang
Konsumsi susu nasional yang semakin tinggi belum dapat diimbangi dengan produksi susu dalam negeri. Konsumsi susu di masyarakat telah mengalami peningkatan sebesar 98.9% selama kurun waktu 6 tahun dan diperkirakan akan terus meningkat pada tahun-tahun selanjutnya (Ditjenak, 2008). Hal ini dapat diatasi dengan perbaikan produksi terhadap sapi perah. Namun, peningkatan dan perbaikan terhadap aspek kualitas susu sapi perah juga perlu diperhatikan mengingat manfaat yang terkandung dalam susu terutama dalam mencerdaskan anak bangsa.
Sapi perah Friesian Holstein (FH) merupakan ternak penghasil susu yang sangat dominan dibandingkan ternak perah lainnya. Sapi perah sangat efisien dalam mengubah makanan ternak berupa hijauan dan konsentrat menjadi susu dengan substansi gizi yang lengkap, salah satunya adalah protein. Protein susu dapat diklasifikasikan ke dalam dua kelompok utama yaitu kasein dan whey. Kedua protein ini memiliki komponen penyusun yang berbeda-beda. Protein susu merupakan salah satu komponen penentu kualitas susu yang dapat ditingkatkan. Hasil pengamatan dari beberapa penelitian menunjukkan bahwa gen pengontrol protein susu telah diketahui berasosiasi signifikan terhadap kadar protein susu dan komposisi protein. Gen-gen tersebut diharapkan dapat digunakan sebagai penanda molekuler tingkat DNA untuk program seleksi ternak terhadap karakter kualitas susu sapi.
Fokus utama pada pembahasan ini adalah protein whey dengan komponen utama β-laktoglobulin. β-laktoglobulin menduduki proporsi terbesar sehingga dikatakan sebagai protein major dari whey. Lunden et al, (1997) menyatakan bahwa β-laktoglobulin yang merupakan komponen utama protein whey yang dapat memberikan dampak positif terhadap komposisi susu.
Tujuan
Penelitian ini bertujuan untuk mengidentifikasi keragaman genetik gen β-laktoglobulin pada sapi perah Friesian Holstein (FH) dengan menggunakan metode Polymerase Chain Reaction- Restriction Fragment Length Polymorphism (PCR-RFLP).
TINJAUAN PUSTAKA
Sapi Friesian Holstein dan Produksi Susunya
Sapi perah dipelihara untuk menghasilkan air susu, ini berarti produktivitas sapi perah ditentukan oleh jumlah air susu yang dihasilkan. Air susu merupakan suatu bahan makanan alami yang mendekati sempurna dengan kandungan protein, mineral dan vitamin yang tinggi, menjadikan susu sebagai sumber bahan makanan yang esensial (Blakely dan Bade, 1994). Bangsa sapi Friesian Holstein (FH) merupakan bangsa sapi perah yang memiliki tingkat produksi tertinggi dibandingkan dengan bangsa sapi perah lainnya. Dengan tingkat produksi rata-rata setiap satu masa laktasi (10 bulan ) adalah sekitar 3.050 liter atau sekitar 10 liter per ekor per hari, di tempat asalnya produksi susu per masa laktasi rata-rata sebanyak 7.245 liter atau sekitar 20 liter per hari (Putranto, 2006). Rendahnya tingkat produksi ini menyebabkan peternak memerlukan input produksi yang tinggi untuk mempertahankan usaha ternak dan pencapaian produksi optimal. Di daerah tropis seperti Indonesia sifat tersebut tidak terekspresi secara maksimal karena kondisi lingkungan di Indonesia yang tidak sesuai seperti daerah asalnya, meskipun daya adaptasi ternak ini relatif tinggi (Anwar, 2008).
Tabel 1. Komposisi Susu Berbagai Bangsa Sapi Perah
Bangsa
Air Protein Lemak Laktosa Abu Bahan Kering ---(%)--- Jersey Guernsey Ayrshire Friesian Holstein Shorthorn 85,27 85,45 87,10 88,01 87,43 3,80 3,84 3,34 3,15 3,32 5,14 4,98 3,85 3,45 3,63 5,04 4,98 5,03 4,65 4,89 0,75 0,75 0,69 0,68 0,73 14,73 14,55 12,90 11,93 12,57 Sumber : Sudono (1999)
Berdasarkan data di atas, dapat diketahui sapi FH merupakan salah satu bangsa sapi perah yang memiliki kemampuan produksi yang cukup tinggi, namun di sisi lain ada kekurangan yang menjadi perhatian dalam peningkatan kualitas komposisi susu yaitu kadar protein yang masih rendah dari sapi FH jika dibandingkan dengan bangsa sapi perah lainnya.
Seleksi dengan Marked Assisted Selection (MAS)
Seleksi dapat didefinisikan sebagai suatu proses dalam memilih individu dari suatu populasi untuk dijadikan tetua bagi generasi berikutnya. Salah satu metode seleksi adalah menggunakan penciri genetik. Banyak gen teridentifikasi mempunyai potensi sebagai penciri genetik atau Marked Assisted Selection (MAS) seperti gen-gen pengontrol protein susu akan bermanfaat dalam mempercepat kegiatan seleksi dari sifat produksi dan bernilai ekonomis (Sumantri et al., 2007). Seleksi keunggulan genetik melalui identifikasi gen yang diprediksi berasosiasi kuat dengan sifat produksi dan kualitas susu akan sangat mendukung bagi program perbaikan genetik sapi FH (Bovenhuis et al., 1992).
Perkembangan aplikasi teknologi pemuliaan telah memberikan banyak informasi mengenai pemetaan gen pada genom sapi sampai pada tingkat keragaman sekuens DNA (Deoxyribose Nucleic Acid ) yang terjadi secara intensif selama dekade terakhir. Sifat produksi susu termasuk sifat kuantitatif yang dikendalikan oleh banyak gen dan diwariskan serta dipengaruhi oleh faktor lingkungan (Noor, 2000).
Protein Susu
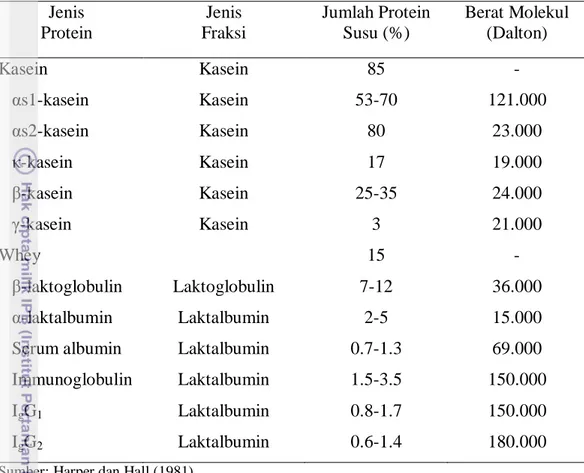
Protein susu bervariasi secara kuantitatif dan kualitatif pada masing individu dalam satu spesies. Secara kuantitatif dapat dilihat dari komposisi elemen-elemen penyusun protein susu yaitu proporsi dari kasein dan whey yang secara umum menurut Harper dan Hall (1981) disajikan pada Tabel 2 :
Protein susu pada dasarnya dapat diklasifikasikan menjadi dua kelompok yaitu kasein dan whey. Kasein merupakan komponen protein yang terbesar dengan proporsi sekitar 80% dalam susu dan 20% berupa whey. (Yahyaoui et al., 2003). Protein whey merupakan protein butiran (globular). Kadar whey dalam susu mencapai 20%. Komponen penyusun whey protein terdiri dari β-laktoglobulin, α-laktalbumin, Bovine Serum Albumin (BSA), Immunoglobulin dan Laktoferrin (Kontopidis, 2000). Protein whey pada susu sapi yang utama adalah β-laktoglobulin yang terdiri dari 7-12 % dari total protein pada susu (Meza et al., 2007).
Tabel 2. Komposisi Protein Susu Ternak Ruminansia. Jenis Protein Jenis Fraksi Jumlah Protein Susu (%) Berat Molekul (Dalton) Kasein αs1-kasein αs2-kasein κ-kasein β-kasein γ-kasein Whey β-laktoglobulin α-laktalbumin Serum albumin Immunoglobulin IgG1 IgG2 Kasein Kasein Kasein Kasein Kasein Kasein Laktoglobulin Laktalbumin Laktalbumin Laktalbumin Laktalbumin Laktalbumin 85 53-70 80 17 25-35 3 15 7-12 2-5 0.7-1.3 1.5-3.5 0.8-1.7 0.6-1.4 - 121.000 23.000 19.000 24.000 21.000 - 36.000 15.000 69.000 150.000 150.000 180.000 Sumber: Harper dan Hall (1981)
Struktur Gen β-laktoglobulin
Pengkodean DNA untuk gen β-laktoglobulin telah dilakukan untuk ternak ruminansia yaitu sapi, domba, dan kambing. Posisi gen β-laktoglobulin pada sapi dan kambing terdapat pada kromosom 11 (Hayes dan Petit, 1993). Struktur gen β-laktoglobulin terdiri dari 7 ekson yang ditunjukkan pada Gambar 1.
E1 E2 E3 E4 E5E6 E7 Keterangan: Intron = Exon =
Exon 1 = 2171-2305 (135 bp) Exon 4 = 5167-5277 (111 bp) Exon 2 = 2975-3114 (135 bp) Exon 5 = 5953-6057 (105 bp) Exon 3 = 3976-4049 (74 bp) Exon 6 = 6279-6321 (43bp) Exon 7 = 6711-6893 (183 bp)
Sumber : Alexander et al (1989).
Keragaman Gen β-laktoglobulin
β-laktoglobulin merupakan salah satu mayor protein susu yang terdapat pada susu ruminansia. β-laktoglobulin termasuk kelompok protein lipocalin yang dapat mengikat molekul-molekul yang bersifat hidrofobik dan berperan penting dalam metabolisme lemak (Kontopidis, 2000). Manfaat dari β-laktoglobulin adalah sebagai pengangkut retinol, asam palmitat, asam lemak, vitamin D dan kolesterol. Selain itu berfungsi untuk membantu regulasi metabolisme phospor pada kelenjar susu. (Madureira et al., 2007)
Keragaman gen β-laktoglobulin dapat meningkatkan persentase protein susu (Kumar et al., 2006). Gen β-laktoglobulin pada ruminansia diteliti untuk melihat hubungan dengan produksi susu dan komposisi susu. Pada ternak sapi dilaporkan terdapat dua varian untuk gen β-laktoglobulin yaitu A dan B (Yahyaoui et al., 2003). Beberapa penelitian yang mengambil topik utama mengenai keragaman gen β-laktoglobulin dan berhasil mengidentifikasi alel-alelnya pada ternak ruminansia yang disajikan pada Tabel 3.
Tabel 3. Keragaman Gen β-laktoglobulin dari Beberapa Penelitian pada Ternak Ruminansia
Ternak Tipe Jumlah Alel Metode Enzim Restriksi Sumber Sapi Sapi Sapi Kambing Sapi Perah Perah Perah Perah Perah 2 2 2 2 2 PCR-RFLP PCR-RFLP PCR-RFLP PCR-RFLP - HaeIII HaeIII HaeIII SaeII - Karimi et al., 2009 Rachagani et al., 2006 Maskur et al., 2005 Kumar et al., 2006 Ojala et al., 1997
Metode PCR-RFLP (Polymerase Chain Reaction- Restriction Fragment Length Polymorphism)
Polymerase chain reaction (PCR) merupakan suatu teknik untuk menggandakan jumlah molekul DNA secara in vitro. Proses ini berjalan dengan bantuan enzim polymerase dan primer. Primer merupakan oligonukleotida spesifik yang menempel pada bagian sampel DNA yang akan diperbanyak (Williams, 2005). Enzim polymerase merupakan enzim yang dapat mencetak urutan DNA baru. Hasil
dari proses PCR dapat langsung divisualisasikan dengan elektroforesis atau dapat digunakan untuk analisis lebih lanjut (Williams, 2005).
Reaksi yang terjadi dalam mesin PCR secara umum dapat dibagi menjadi tiga tahap yaitu tahap denaturasi (pemisahan untai ganda DNA), tahap annealing (penempelan primer) dan tahap ekstensi (pemanjangan primer). Reaksi ini umumnya terjadi dalam 25-30 siklus. Pada tahap denaturasi, DNA dipanaskan hingga 94oC sehingga DNA untai ganda berpisah menjadi DNA untai tunggal. Tahapan yang paling menentukan dalam proses PCR adalah tahap penempelan primer, karena tiap pasangan primer memiliki suhu penempelan yang spesifik. Tahap ekstensi/elongasi/pemanjangan primer terjadi pada suhu 72oC. Pada tahapan ini enzim taq polymerase, buffer PCR, dNTP, dan Mg2+ memulai aktifitasnya memperpanjang primer (Viljoen et al., 2005).
Restriction Fragment Length Polymorphism (RFLP) adalah profil DNA berupa fragmen-fragmen DNA hasil pemotongan enzim endonuklease untuk berbagai individu. Enzim endonuklease atau enzim restriksi (RE) mengenali situs pemotongan empat dan enam basa. RFLP memiliki kemampuan untuk mendeteksi keragaman di tingkat alel didasarkan pada polimorfisme yang muncul karena adanya basa yang mengalami substitusi, penambahan, pengurangan dan perpindahan (translokasi) pada genom DNA. Kelebihan RFLP adalah dapat mendeteksi sifat kodominan, artinya dapat membedakan antara yang homozigot dan heterozigot. Selain itu kelebihan yang lain adalah diharapkan didapatkan homologi polimorfik (Gupta et al., 2002).
Yahyaoui et al. (2001) menyatakan bahwa PCR-RFLP merupakan suatu metode yang sederhana dan bisa digunakan untuk mencari keragaman genotip. Analisis RFLP biasa digunakan untuk mendeteksi adanya keragaman pada gen yang berhubungan dengan sifat ekonomis, seperti produksi dan kualitas susu (Sumantri et al., 2005).
Elektroforesis
Elektroforesis adalah suatu teknik pemisahan molekul seluler berdasarkan ukurannya. Elektroforesis menggunakan medan listrik yang dialirkan pada suatu medium yang mengandung sampel yang akan dipisahkan. Teknik ini dapat digunakan dengan memanfaatkan muatan listrik yang ada pada makromolekul,
misalnya DNA yang bermuatan negatif (Triwibowo, 2008). Winarno dan Agustinah (2007) menyatakan elektroforesis adalah suatu cara pemisahan campuran dan beberapa senyawa dengan melakukan suspense ke dalam air kemudian diberikan aliran listrik. Gel yang ditempatkan ke dalam sumur elektroforesis yang mengandung larutan buffer dan dialiri listrik, molekul DNA yang bermuatan negatif pada pH netral akan bergerak ke arah positif. DNA bergerak melalui gel pada kecepatan yang berbeda tergantung ukurannya.
Muladno (2002) menyatakan hasil analisis DNA dapat dilihat melalui proses elektroforesis. Komponen bahan kimia terpenting yang digunakan dalam proses tersebut adalah gel. Kecepatan migrasi DNA ditentukan oleh beberapa faktor yaitu ukuran molekul DNA, konsentrasi agarose, konformasi DNA, voltase yang digunakan, adanya ethidium bromide di dalam gel dan komposisi larutan buffer. Gel yang biasa digunakan adalah yaitu gel agarose dan gel poliakrilamida.
Keragaman Genetik
Keragaman genetik dalam suatu populasi digunakan untuk mengetahui dan melestarikan bangsa-bangsa dalam populasi terkait dengan penciri suatu sifat khusus. Pengetahuan akan keragaman genetik suatu bangsa akan sangat bermanfaat bagi keamanan dan ketersediaan bahan pangan yang berkesinambungan (Blot et al., 1998).
Estimasi perhitungan keragaman genetik dalam populasi secara kuantitatif dapat diperoleh melalui dua ukuran keragaman variasi populasi yaitu proporsi lokus polimorfisme dalam populasi dan rata-rata proporsi individu heterozigot dalam setiap lokus (Nei, 1987). Keragaman genetik pada protein susu telah menjadi perhatian besar dalam industri sapi perah, terutama karena kemampuan dalam berasosiasi dengan komposisi, koagulasi rennet, dan pembuatan keju dan secara ekonomi penting dalam suatu industri keju (Celik, 2003).
Keseimbangan keragaman genetik dapat ditentukan dengan menggunakan hukum Hardy-Weinberg yang menyatakan bahwa jumlah frekuensi gen dominan dan resesif adalah 1 dan jumlah proporsi dari ketiga macam genotip (p2, 2pq dan q2) adalah 1. Adapun faktor-faktor yang mempengaruhi frekuensi gen antara lain adalah seleksi, mutasi, pencampuran populasi, dan genetic drift (Noor, 2000).
MATERI METODE Lokasi dan Waktu
Penelitian ini akan dilaksanakan pada bulan Desember 2009 sampai dengan bulan Mei 2010. Penelitian ini dilaksanakan di Laboratorium Genetika Molekuler Ternak, Bagian Pemuliaan dan Genetika Ternak, Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Materi Sampel
Sebanyak 88 sampel DNA sapi betina laktasi diperoleh dari koleksi sampel DNA Laboratorium Genetika Molekuler Ternak, Bagian Pemuliaan dan Genetika Ternak Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor. Sapi Friesian-Holstein berasal dari BPPT-SP Cikole, Lembang yang telah mengalami proses ekstraksi.
Primer
Primer adalah DNA utas tunggal dengan ukuran pendek, biasanya 18 sampai 25 pb (pasang basa)yang akan menempel pada DNA cetakan ditempat yang spesifik. Primer yang digunakan adalah EA3 dan EA4. Amplifikasi gen β-laktoglobulin pada ekson 4 mengikuti Karimi et al., (2009) pada Tabel 4.
Tabel 4. Sekuens Primer
.
Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP)
Amplifikasi gen (DNA Target) menggunakan bahan-bahan yang terdiri dari air destilata, 10×buffer, MgCl2, dan dNTP, pasangan primer (forward dan reverse), enzyme taq DNA polymerase, enzim restriksi HaeIII dengan buffernya. Alat–alat yang digunakan adalah tabung eppendorf, pipet mikro, alat sentrifugasi, vortex, mesin PCR (DNAThermal Cycler), deep freezer dan inkubator.
Gen Sekuen Primer Produk
PCR Enzim Restriksi Β-LG 5´TGTGCTGGACACCGACTACAAAAA3´ (forward) 5´GCTCCCGGTATATGACCACCCTCT-3´ (reverse). 247 pb HaeIII
Elektroforesis
Elektroforesis DNA produk PCR menggunakan gel agarose berukuran besar (Agarose 1,5 %) yang terbuat dari 0,45 g agarose, 30 ml 0,5 x TBE , 2,5 µl EtBr (Ethidium bromide). Proses elektroforesis menggunakan bahan seperti 10 x buffer, loading dye, dan marker. Alat-alat yang digunakan adalah timbangan digital (Sartorius basic), gelas ukur, gelas kimia, microwave, stirrer, plate cetakan gel, power supply, pipet mikro, tip, plastik mika, sarung tangan dan UV-transilluminator.
Prosedur Pengambilan Sampel Darah
Sampel darah yang digunakan dalam penelitian ini adalah darah yang berasal dari sapi FH laktasi yang telah diketahui data produksi dan kualitas susunya dari BPPT-SP Cikole sebanyak 88 ekor, sampel darah diambil dari bagian vena juguralis dengan menggunakan jarum vaccutainer no. 21 G dan disimpan dalam tabung vaccutainer 10 ml dengan alkohol.
Isolasi DNA dilakukan dari sampel darah dengan menggunakan metode ekstraksi phenol-chloroform (Sambrook et al. 1989) yang telah dimodifikasi untuk penggunaan sampel darah yang disimpan dalam alkohol, dengan prosedur sebagai berikut:
Preparasi sampel. Sampel darah dalam EtOH sebanyak 200 ul dipindahkan ke tabung 1,5 ml. Kemudian dihilangkan alkoholnya dengan menambahkan air destilasi 1000 µl. Sampel disentrifugasi pada kecepatan 8000 rpm selama 5 menit dan bagian supernatannya dibuang. Pencucian tersebut diulangi untuk menghilangkan alkohol dalam darah sebanyak mungkin.
Degradasi protein. Sampel yang telah bersih dari alkohol ditambahkan 1x STE (sodium tris EDTA) sampai volume 340 µl (± 200 µl), 40 µl SDS (sodium dosesil sulfat) 10% dan 20 µl proteinase K 5 mg/ml. Campuran diinkubasi dan digoyang pada suhu 50 ºC selama semalam.
Degradasi bahan organik. Sampel yang telah diinkubasi ditambahkan 400 µl larutan phenol, 400 µl CIAA (choloform : isoamyl alkohol (24 :1) dan 40 µl 5M NaCl. Campuran digoyang pada suhu ruang selama satu jam.
Presipitasi DNA. Sampel selanjutnya disentrifugasi pada kecepatan 5000 rpm selama 10 menit hingga fase DNA terpisah dengan fase phenol. Fase DNA dipindahkan dalam tabung baru. Ditambahkan 800 µl alkohol absolut dan 40 µl 5M NaCl, kemudian diinkubasi pada suhu -20 ºC selama semalam. Larutan disentrifugasi pada kecepatan 12000 rpm selama 10 menit, bagian alkohol dibuang sebanyak mungkin. Bagian endapan (DNA) yang tersisa ditambahkan 800 µl alkohol 70% dan disentrifugasi seperti sebelumnya, dan bagian alkohol dibuang sebanyak mungkin. Bagian DNA yang tersisa dibiarkan dalam keadaan terbuka pada suhu ruang sampai kering. Tambahkan 100 µl larutan pengelusi yang telah dipanaskan sebelumnya. DNA disimpan pada suhu -20ºC dan siap untuk digunakan.
Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP)
PCR dikondisikan pada volume reaksi 25 µl tiap tabung yang terdiri dari sampel sebanyak 2 µl, air destilata 16,7 µl, 0,5 µl masing-masing primer, 10×buffer sebanyak 2,5 µl, 2,5 mM MgCl2 2 µl, dan 2 mM dNTP 0,3 µl, BSA 1 µl dan enzim
taq 0,1 µl. Tabung PCR dihomogenkan dengan menggunakan vortex, kemudian diendapkan dengan menggunakan centrifuge pada kecepatan 5.000 rpm selama setengah menit.
Amplifikasi dilakukan selama 35 siklus dengan tahapan pertama adalah denaturasi awal pada suhu 95 ºC selama lima menit, tahap kedua yaitu denaturasi pada suhu 95 ºC selama 30 detik, annealing (penempelan primer) pada suhu 60 ºC selama 45 detik, pemanjangan molekul DNA pada suhu 72 ºC selama satu menit, dan tahap ketiga yaitu pemanjangan akhir molekul DNA pada suhu 72 ºC selama lima menit.
Produk PCR sebanyak 5µl dipindahkan ke tabung baru dan ditambahkan 0,3 µl enzim restriksi HaeIII 10 unit, 0,7 buffer R dan 1 µl air destilasa kemudian diinkubasi selama 1 malam (overnight) pada suhu 37 ºC.
Elektroforesis
Elektorforesis DNA produk PCR menggunakan gel agarose 2,5 % dengan mencampurkan 0,75 g agarose dan 30 ml 0,5 x TBE yang dipanaskan selama empat menit di dalam microwave, didinginkan di atas stirrer yang sudah dicelupkan magnet stir dan ditambahkan sebanyak 2,5 µl EtBr. Gel dipindahkan ke dalam cetakan agar
dan didinginkan kurang lebih 30 menit. Produk PCR sebanyak 7 µl yang dihomogenkan dengan loading dye dimasukkan ke dalam sumur-sumur gel agarose yang sudah digenangi larutan 0,5 x TBE buffer kemudian elektroforesis dilakukan selama 45 menit. Hasil elektroforesis divisualisasikan dengan mengamatinya di atas UV-transilluminator.
Analisis data
Keragaman genotipe tiap-tiap individu dapat ditentukan dari pita-pita DNA gen yang ditemukan. Masing-masing sampel dibandingkan berdasarkan ukuran (marker) yang sama dan dihitung frekuensi alelnya.
Frekuensi alel pada lokus dihitung berdasarkan rumus Nei (1987): Xi = 2nii + ∑nij
2N Keterangan :
Xi = frekuensi alel ke- i
nij = jumlah individu yang bergenotipe ij
nii = jumlah individu yang bergenotipe ii
N = jumlah individu sampel
Derajat heterozigositas (ĥ) dihitung berdasarkan frekuensi alel pada tiap lokus DNA dengan rumus Nei (1987) :
ĥ = 2n (1 - ΣXi)
(2n – 1) Keterangan :
Xi = frekuensi alel
ĥ = nilai heterozigositas lokus n = Jumlah individu sampel
Uji chisquare dihitung untuk mengetahui kesetimbangan alel/genotipe berdasarkan hukum Hardy-Weinberg oleh (Hartl dan Clark, 1997) yaitu :
χ2
= ∑(obs – exp)2 exp
Keterangan : χ2
= uji Khi-kuadrat
obs = jumlah nilai pengamatan exp = jumlah nilai harapan Derajat Bebas (db) = k - 1
HASIL DAN PEMBAHASAN Amplifikasi Gen β-laktoglobulin
Gen β-laktoglobulin pada ternak sapi perah Friesian Holstein (FH) betina laktasi yang berasal dari BPPT-SP Cikole telah berhasil diidentifikasi melalui proses amplifikasi DNA. Proses amplifikasi dilakukan dengan metode PCR-RFLP. Penelitian ini menggunakan pasangan primer berdasarkan literatur Karimi et al. (2009). Perkiraan Panjang fragmen yang diamplifikasi dapat diketahui dengan menyesuaikan sekuens gen β-laktoglobulin pada ekson 4 yang diperoleh dari GeneBank no.akses X14710 (Lampiran 1). Hasil amplifikasi gen β-laktoglobulin tersebut diperlihatkan pada Gambar 2.
Keterangan : M = Marker
1-12 = sampel sapi FH
Gambar 2. Pita DNA Amplifikasi Gen β-laktoglobulin Sapi FH Metode PCR pada Gel Agarose 2,5 % .
Amplifikasi DNA dengan menggunakan primer tertentu menghasilkan produk PCR sepanjang 247 pasang basa. Keberhasilan dalam mengamplifikasi DNA bergantung pada interaksi komponen PCR dalam konsentrasi yang tepat. Beberapa hal yang umum dilakukan untuk optimasi PCR diantaranya adalah suhu annealing (penempelan primer), konsentrasi Mg 2+, konsentrasi primer dan konsentrasi DNA target (Viljoen et al., 2005).
Suhu annealing merupakan kisaran suhu yang membuat pasangan primer menempel dengan komplemennya pada fragmen DNA target saat proses PCR dilakukan. Suhu annealing sangat menentukan keberhasilan amplifikasi karena
M 1 2 3 4 5 6 7 8 9 10 11 12
500 pb 247 pb 100 pb
proses perpanjangan DNA dimulai dari primer. Biasanya suhu yang digunakan agar terjadinya penempelan adalah 50-65 o C dan pada penelitian ini suhu yang digunakan adalah 60oC sesuai dengan Karimi et al. (2009).
Pendeteksian Keragaman Gen β-laktoglobulin
Amplifikasi gen β-laktoglobulin menghasilkan panjang fragmen 247 pb. Proses pemotongan gen dilakukan dengan menggunakan enzim restriksi HaeIII yang mengenali situs pemotongan empat basa pada posisi 5’....GGCC...3’. Literatur yang menjadi acuan yaitu Karimi et al. (2009) menginformasikan bahwa genotipe AA memiliki potongan dua fragmen yaitu 148 pb dan 99 pb, genotipe BB memiliki tiga fragmen 74 pb, 74 pb dan 99 pb, sedangkan genotipe AB terdiri dari 3 fragmen yaitu 148 pb, 99 pb, dan 74 pb. Hasil penelitian sesuai dengan acuan yang dipakai. Enzim HaeIII berhasil memotong di dua titik yaitu pada posisi basa ke 74 dan 148 dari produk PCR. Pemotongan tersebut menghasilkan tiga fragmen yang panjangnya berturut-turut adalah 74, 74 dan 99 pb. Ketiga fragmen tersebut menunjukkan genotipe dengan homozigot BB yang berarti alel B.
Alel A akan terbentuk jika terdapat mutasi pada salah satu basa dari ke empat basa sebagai situs pengenal enzim restriksi HaeIII. Situs tersebut tidak dikenali oleh enzim sehingga tidak terjadi pemotongan. Alel yang muncul pada populasi sapi FH di Cikole adalah A dan B sedangkan genotipe yang ditemukan AA, AB dan BB. Genotipe AA terdiri dari 2 fragmen pada basa ke 148 dan 99, genotipe AB menunjukkan adanya tiga fragmen yaitu pada posisi basa ke 148, 99, dan 74 sedangkan genotipe BB terdiri dari tiga fragmen yaitu pada posisi 74, 74, dan 99 .
Keragaman alel pada sapi Friesian Hosltein ditunjukkan dengan sekuens gen β-laktoglobulin ekson 4 pada Gambar 3.
Primer forward
1 ttccagcctt gaatgagaac aaagtccttg tgctggacac cgactacaaa aagtacctgc 61 tcttctgcat ggagaacagt gctgagcccg agcaaagcct GG*CCtgccag tgcctgggtg 121 ggtgccaacc ctggctgccc agggagacca gctgtgtggt cctcgctgca acggGG*CCgg 181 gggggacggt gggagcaggg agcttgattc ccaggaggag gagggatggg gggtccccga 251 gtcccgccag gagagggtgg tcatataccg ggagccggtg tcctgggggc ctgtgggtga
Gambar 3. Posisi Penempelan Primer (cetak tebal) pada Sekuen Gen β-laktoglobulin nomor akses GenBank X14710. Terjadi Mutasi pada Situs Pemotongan HaeIII (GG|CC) pada Posisi 102.
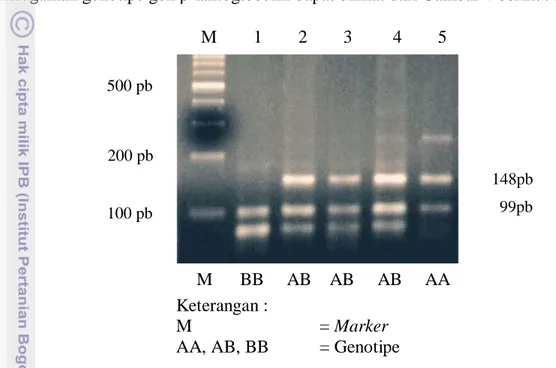
Asumsi yang mendukung dalam penentuan tipe genotipe yaitu semua pita yang memiliki laju migrasi yang sama merupakan alel yang homolog (Nei, 1987). Keragaman genotipe gen β-laktoglobulin dapat dilihat dari Gambar 4 berikut :
Keterangan :
M = Marker
AA, AB, BB = Genotipe
Gambar 4. Pola Pita Gen β-laktoglobulin Sapi FH dalam Gel Agarose 2,5 % dengan Enzim Restriksi HaeIII.
Keragaman terjadi akibat adanya mutasi. Mutasi adalah suatu perubahan struktur kimia gen yang berakibat berubahnya fungsi gen. Keragaman DNA adalah salah satu akibat dari mutasi (Toland, 2008). Point mutations atau mutasi titik adalah mutasi yang terjadi yang terjadi hanya pada satu nukleotida atau bagian kecil dari gen. Mutasi titik dapat dibedakan berdasarkan tipe perubahan runutan nukleotida yaitu delesi, insersi, subtitusi (transisi dan transversi), kesalahan pembacaan (proofreading errors), dan perubahan struktur kimia pada basa (Paolella, 1998). Mutasi yang terjadi pada penelitian ini adalah mutasi substitusi tipe transisi yaitu perubahan basa antar basa purin (A-G) atau basa pirimidin (T-C). Basa sitosin menjadi timin menyebabkan perubahan ekspresi asam amino pada proses translasi. Asam amino yang diterjemahkan yaitu dari valine menjadi alanin. Mutasi yang terjadi dapat dilihat pada Gambar 5.
M 1 2 3 4 5 148pb 100 pb 500 pb 200 pb 99pb M BB AB AB AB AA
Basa Timin Mutasi Transisi T- C
Alel A : 5’--GCCCG AGCAAAGCCT GG *TCTGCCAG TGCCTG… --- 3’ Alel B : 5’--GCCCG AGCAAAGCCT GG*CCTGCCAG TGCCTG… --- 3’
Basa Sitosin
Gambar 5. Perbedaan sekuen Gen β-laktoglobulin nomor akses GenBank X14710 yang disebabkan mutasi titik C – T (Alel B mempunyai basa C pada posisi basa ke 102 pada gambar 3 sedangkan alel A mempunyai basa T). Frekuensi Gen β-laktoglobulin pada Sapi Friesian Holstein
Sapi FH menduduki populasi terbesar, bahkan hampir di seluruh dunia, baik di negara-negara sub-tropis maupun tropis seperti di Indonesia. Sampel sapi yang diamati sebanyak 88 ekor dengan genotipe AA, AB dan BB berturut-turut berjumlah 9, 53 dan 26 ekor dari total sampel.
Penelitian ini menghasilkan frekuensi alel yang berbeda. Alel A sebesar 0,40 dan alel B 0,60 sedangkan frekuensi genotipe diperoleh 0,10, 0,60, dan 0,30 berturut-turut untuk AA, AB, dan BB. Hal ini menunjukkan bahwa gen β-laktoglobulin bersifat polimorfik (beragam) sesuai dengan Nei (1987) yang menyatakan bahwa suatu alel dikatakan polimorfik jika memiliki frekuensi alel sama dengan atau kurang dari 0,99. Nilai frekuensi dapat dilihat pada Tabel 5 berikut.
Tabel 5. Nilai Frekuensi Genotipe, Frekuensi Alel dan Nilai Heterozigositas Lokus Hae III pada sapi Friesian Holstein
N Frekuensi Genotipe Frekuensi Alel Heterozigositas (ĥ) AA AB BB A B 88 0,10 0,60 0,30 0.40 0,60 0.483 88 9 53 26 62 79
Keterangan : N = Jumlah sampel
Keragaman genotipe gen β-laktoglobulin sapi perah Friesian Holstein dapat dibandingkan dengan beberapa penelitian sebelumnya. Penelitian ini sesuai dengan Bobe et al.(1999) yang melaporkan bahwa sapi FH di Amerika terdiri dari tiga variasi genotip yaitu AA, AB, dan BB dan dua alel yaitu A dan B. Frekuensi genotipe AA, AB, dan BB adalah 0,125, 0,476, dan 0,399. Lunden et al. (1997) juga melaporkan hasil penelitiannya, ditemukan 2 alel pada sapi FH Swedia yaitu alel A
dan B dengan frekuensi masing-masing 0,498 dan 0,502. Frekuensi genotipe yaitu 0,235, 0,525, dan 0,240 untuk AA, AB, dan BB. Namun, penelitian ini berbeda dengan Heck et al. (2009) yang menemukan bahwa sapi FH Belanda memiliki frekuensi alel A yang lebih tinggi yaitu 0,583 dibandingkan alel B sebesar 0,471.
Karimi et al. (2009) melaporkan terdapat dua alel gen β-laktoglobulin pada sapi Nadji India yaitu alel A sebesar 0.0875 dan alel B sebesar 0,9125 sedangkan variasi genotipe terdiri dari AA (0), BB (0,175), dan AB (0,825). Rachagani et al. (2006) juga menemukan dua alel (A dan B) untuk gen β-laktoglobulin pada Sapi Sahiwal dengan frekuensi alel masing-masing adalah 0,17 dan 0,83. Sapi Tharparkar menghasilkan frekuensi alel A sebesar 0,39 dan B sebesar 0,61. Maskur et al. (2005) dalam penelitiannya melaporkan terdapat dua alel pada sapi Hissar dengan frekuensi alel A dan B berturut-turut sebesar 0,19 dan 0,81. Hasil penelitian ini juga tidak berbeda dengan hasil penelitian Curi et al. (2005) yang menyatakan bahwa keragaman β-laktoglobulin pada sapi Angus, dan Nelore dengan enzim HaeIII memiliki frekuensi alel B yang lebih tinggi dibandingkan dengan alel A.
Distribusi Genotipe Gen β-laktoglobulin
Keseimbangan variasi genotipe yang muncul pada populasi sapi FH dapat diukur dengan menggunakan uji signifikansi chi-square test ( 2). Hasil uji 2 dapat dilihat pada Tabel 6.
Tabel 6. Distribusi Genotipe Gen β-laktoglobulin pada Sapi FH
Genotipe Frekuensi Nilai Nilai χ2 χ2 Genotipe Observasi Ekspektasi Hitung Tabel AA 0,10 9 14,08 1,83
AB 0,60 53 42,24 2,74 5,99 BB 0,30 26 31,68 1,02
Total 1 88 88 5,59
Berdasarkan uji signifikansi tersebut diperoleh 2 Hitung < 2 Tabel, artinya bahwa distribusi genotipe β-laktoglobulin pada populasi sapi FH berada dalam keseimbangan Hardy-Weinberg. Keseimbangan genotipe pada sapi FH Cikole dapat disebabkan tidak terjadinya seleksi pada ternak. Populasi tersebut tidak mengalami
pertukaran atau pergantian ternak, artinya pihak BPPT-SP Cikole masih mempertahankan ternak yang ada.
Heterozigositas disebut juga sebagai keragaman genetik. Nilai heterozigositas merupakan cara yang paling akurat untuk mengukur keragaman suatu populasi (Nei,1987). Nilai heterozigositas dapat dipengaruhi oleh jumlah sampel, jumlah alel dan frekuensi alel. Populasi sapi perah di Cikole memiliki nilai heterozigositas sebesar 0,483, ini berarti bahwa berbanding lurus dengan keragaman genetik yang tinggi pula.
Hubungan antara keragaman gen β-laktoglobulin dengan susu dan komposisi susu dapat ditemukan pada beberapa penelitian. Sapi FH dengan genotip BB dapat memproduksi susu 30% lebih rendah dari genotip AA. Genotipe BB dapat meningkatkan persentase lemak susu lebih tinggi dibandingkan genotype AA. (Berry et al., 2010). β-laktoglobulin berasosiasi dengan perubahan jumlah dan komposisi susu. Sapi perah dengan alel B berpengaruh rendah terhadap produksi akan tetapi dapat meningkatkan kandungan kasein pada susu (Bobe et al., 2009). Bobe et al. (1999) menemukan keragaman genotip gen β-laktoglobulin dan κ-casein yaitu AA, AB, dan BB pada populasi sapi FH di Amerika sebagai protein major mampu memberikan pengaruh terhadap total protein susu, akan tetapi tidak memberikan hasil yang signifikan terhadap konsentrasi protein. Ojala et al. (1997) juga meneliti efek keragaman genotipe protein susu sapi FH dan Jersey terhadap komposisi susu, ditemukan bahwa genotipe β-laktoglobulin lebih sedikit menyumbangkan kontribusi terhadap variasi fenotipik persentase lemak, susu dan produksi protein.
KESIMPULAN DAN SARAN Kesimpulan
Identifikasi keragaman gen β-laktoglobulin pada ekson 4 menggunakan metode PCR-RFLP menghasilkan produk amplikon sepanjang 247 pb. Proses pemotongan gen β-laktoglobulin oleh enzim restriksi HaeIII memperlihatkan adanya keragaman atau bersifat polimorfik. Genotyping gen β-laktoglobulin menghasilkan dua tipe alel, yaitu A dan B; sehingga diperoleh tiga variasi genotipe, yaitu AA, AB dan BB. Keragaman gen tersebut didukung oleh nilai heterozigositas yang cukup tinggi sebesar 0,483. Genotipe BB yang memiliki pengaruh positif terhadap kualitas susu mempunyai nilai frekuensi lebih tinggi dibandingkan genotipe AA (0,30 vs 0,10). Meskipun demikian, varian alel A dan B (0,40 vs 0,60) sapi FH pengamatan masih berada dalam keseimbangan Hardy –Weinberg.
Saran
Memperbanyak sampel dengan mempertimbangkan beberapa kondisi manajemen pemeliharaan untuk mengetahui lebih jauh keragaman gen β-laktoglobulin serta pengaruhnya terhadap kualitas dan kadar protein susu sapi FH di dalam negeri perlu dilakukan.
UCAPAN TERIMA KASIH
Puji syukur penulis panjatkan kehadirat Allah SWT yang telah memberikan taufik dan karunia-Nya yang tak terhingga sehingga penulis dapat menyelesaikan skripsi ini dengan baik. Shalawat serta salam semoga tercurahkan kepada teladan ummat manusia, Muhammad SAW, beserta para keluarga, sahabat serta para pengikutnya hingga akhir zaman.
Penulis mengucapkan terima kasih yang sebesar-besarnya kepada Bapak Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc dan Ibu Ir. Anneke Anggraeni, M.Si., Ph.D yang telah membimbing, mengarahkan, meluangkan waktu bagi penulis, mulai saat penyusunan proposal, tahap penulisan skripsi dan ujian akhir sarjana. Penulis juga mengucapkan terima kasih kepada Bapak Dr.Jakaria, S.Pt.,M.Si dan Dr. Ir. Kartiarso, M.Si atas masukan dan saran dalam perbaikan skripsi pada saat ujian akhir penulis.
Penulis mengucapkan terima kasih kepada Ayahanda Drs. H. Maraenda Hrp, SH. MH dan Ibunda Dra. Pitta Hara Siregar yang tiada henti menyayangi dengan tulus, mengajarkan, mendidik dan mendo’akan yang terbaik untuk keberhasilan penulis. Terima kasih kepada kakak dan adikku tercinta, Mawaddah Tsaniyah, SKM, Mufidah Ulfah, SH, Lukman Hakim dan si kecil Farhan Baleo Alamsyah yang senantiasa menyemangati penulis dan memberikan do’a sehingga penulis dapat menyelesaikan studi selama di IPB, serta seluruh keluarga besar yang ada di Medan.
Ucapan terima kasih juga penulis sampaikan kepada kakak-kakak dan teman-teman seperjuangan dalam penelitian ini yang telah banyak membantu penulis dan memberikan banyak pelajaran terutama Mbak Restu, Kak Erik, Kak Kia, Kak Zul, Kak Tikno, Yadin, Adit dan Ratna serta seluruh anggota Laboratorium Genetika dan Molekuler, tak lupa pula sahabat-sahabat tercinta atas ukhuwah yang telah diberikan, teman-teman di IPTP 43, Famm Al-An’aam, dan keluarga besar Wismo Ayu Depan (Macik, Mba ncun, Rani, Puspa, Endang, Anti, Eka, Trisna, Alim dan Mba eka), teman-teman ROIA-D dan ID yang banyak memberikan pelajaran hidup yang sangat berharga bagi penulis. Terakhir, penulis ucapkan terima kasih kepada semua pihak yang tidak dapat disebutkan satu-persatu.
Bogor, Juli 2010
Penulis
DAFTAR PUSTAKA
Alexander,L. J., C.W. Beattie, G. Hayes, M. J. Pearse, A. F. Stewart, & A.G. Mackinlay. 1989. Isolation and characterization of the bovine beta-lactoglobulin gene. University of New South Wales, School of Biochemistry, P.O.Box 1, Kensington, N.S.W. 2033,Australia
Anwar. 2008. Tentang Ternak Perah. file:///journal/item/2/tentang_ternak_perah. [30 Juni 2010]
Berry S. D., N. Lopez-Villalobos , E. M. Beattie , S. R. Davis , L. F. Adams , N. L. Thomas , A. E. Ankersmit-Udy , A. M. Stanfield, K. Lehnert , H. E. Ward, J. A. Arias, R. J. Spelman, & R. G. Snell. 2010. Mapping a quantitative trait locus for the concentration of beta-lactoglobulin in milk, and the effect of beta-lactoglobulin genetic variants on the composition of milk from Holstein-Friesian x Jersey crossbred cows. J. Vet. NZ. 58:1-5.
Blakely, J. & D. H. Bade. 1994. Ilmu Peternakan. Edisi keempat. Gajah Mada University Press. Yogyakarta.
Blott, S. C., J. L. Wiliams & C. S. Haley. 1998. Genetic relationship among European cattle breeds. Anim. Genetics. 29: 273-282.
Bobe, G., D. C. Beitz, A. E. Freeman, & G. L. Lindberg. 1999. Effect of milk protein genotypes on milk protein composition and its genetic parameter estimates. J. Dairy Sci. 82:2797–2804.
Bobe, G., G. L. Lindberg, L. F. Reutzel & M. D. Hanigan. 2009. Effects of lipid supplementation on the yield and composition of milk from cows with different β-lactoglobulin phenotypes. J. Dairy Sci. 92: 197–203.
Bovenhuis, H., J.A.M. Van Arendonk & S. Kerver. 1992. Associations between milk protein polymorphism and milk production traits. J. Dairy Sci. 75: 2549– 2559.
Celik, S. 2003. β-lactoglobulin genetic variants in Brown Swiss breed and its association with composional properties and rennet cloting time of milk. Int. Dairy J. 13: 727-731.
Curi, R. A., H. N. Oliveira, M. A. Gimenes, A. C. Silveira, & C. R. Lopes. 2005.
Effects of CSN3 and LGB gene polymorphisms on production traits in beef cattle. Gene and Molec Bio. 28: 262-266.
[Ditjennak] Direktorat Jenderal Peternakan. 2008. Statistik Peternakan 2008. Jakarta: Departemen Pertanian.
Gupta, P. K., R. K. Varshney & M. Prasad, 2002. Molecular Markers: Principles and Methodology. Dalam : Jain, S. M., D. S. Brar, and B. S. Ahloowalia (Eds.). Molec. Tech. in Crop Improv. 9: 54.
Harper, W.J., & E.W. Hall. 1981. Dairy Technology and Engineering. AVI Publ. co. Inc., Westport.
Hartl, D. L., & A. G. Clark. 1997. Principle of Population Genetic. Sinauer Associates, Sunderland. MA.
Hayes, H.C., & E.J. Petit. 1993. Mapping of the β-lactoglobulin gene and of immuno-globulin M heavy chain-like sequence to homologous cattle, sheep and goat chromosomes. Mamm. Gen. 4: 207–210.
Heck, J. M. L., A. Schennink, H. J. F. van Valenberg, H. Bovenhuis, M. H. P. W. Visker, J. A. M. van Arendonk, & A. C. M. van Hooijdonk. 2009. Effects of milk protein variants on the protein composition of bovine milk. J. Dairy Sci. 92: 1192–1202.
Karimi. K., M. T. B. Nassiry, K. Mirzadeh, A. Ashayerizadeh., H. Roushanfekr, & F. Fayyazi. 2009. Polymorphism of the β-laktoglobulin gene and its association with milk production traits in Iranian Nadji cattle. Iranian Journal of Biotechnology.7: 2
Kontopidis, G. C. Hold & L. Sawyer. 2004. Inveted review: β-laktoglobulin: binding properties, structure, and function. J. Dairy Sci. 87: 785-796.
Kumar, A., P. K. Rout & R. Roy. 2006. Polymorphism of β-lactoglobulin gene in Indian goats and its effect on milk yield. J. Appl. Genet 47: 1
Lunden. A., M. Nilson, & L. Janson. 1997. Marked effect of β-lactoglobulin polymorphism on the ratio of casein to total protein in milk. J. Dairy Sci. 80: 2996-3005.
Madureira, A. R., C. I. Pereira, A. M. P. Gomes, M. E. Pintado, & F. X. Malcata. 2007. Bovine whey proteins : Overview on their main biological properties. Food Res. Int. 40: 1197-1211.
Maskur, C. Sumantri & Muladno. 2005. Karakterisasi gen β-laktoglobulin dan hubungannya dengan sifat produksi susu pada sapi Hissar. Zuriat.16: 2
Meza, N. M. A., V. B. Cordoba, A. F. G. Cordova, F. Felix, & F. M Goycoolea. 2007. Effect of β-lactoglobulin A and B whey protein variants on the rennet-induced gelation of skim milk gels in a model reconstitude skim milk system. J. Dairy Sci. 90: 582-593.
Muladno. 2002. Seputar Teknologi Rekayasa Genetika. Pustaka Wirausaha Muda dan USESE Foundation. Bogor.
Nei, M. 1987. Molecular Evalutionery Genetics. Columbia University Press. New York.
Noor, R. R. 2008. Genetika Ternak. Penebar Swadaya, Jakarta.
Ojala, M., T.R. Famula & J.F. Medrano. 1997. Effects of milk protein genotypes on the variation for milk production traits of Holstein and Jersey Cows in California. J. Dairy Sci. 80: 1776–1785
Paolella, P. 1998. Introduction to Molecular Biology. McGraw Hill Companies, Inc.. Massachusetts.121-143.
Putranto, E.H. 2006. Analisis keuntungan usaha peternakan sapi perah rakyat di Jawa Tengah. Tesis. Program Pasca Sarjana, Universitas Diponegoro, Semarang. Rachagani, S., I. D. Gupta, N. Gupta, & S. C. Gupta. 2006. Genotyping of
β-Lactoglobulin gene by PCR-RFLP in Sahiwal and Tharparkar cattle breeds. BMC Genet. 7: 31-34.
Sambrok J, F. Fritsch, & T. Miniatis. 1989. Molecular Clooning Laboratory Manual. 3rd Edition. New York: Cold Spring Harbor Laboratory Press.
Sumantri, C., A. Anggraeni, R. R. A. Maheswari, K. Dwiyanto. & A. Fajarallah. 2005. Pengaruh genotype kappa-kasein terhadap kualitas susu pada sapi perah FH di BPTU Baturaden. Prosiding. Seminar Nasional Teknologi Peternakan dan Veteriner 2005.
Sumantri, C., A. Anggraeni., A. Farajallah., & D. Perwitasari. 2007. Keragaman mikrosatelit DNA sapi perah FH di balai pembibitan ternak unggul Baturraden. JITV 12: 124-133.
Sudono, A. 1999. Ilmu Produksi Ternak Perah. Fakultas Peternakan, Institut Pertanian Bogor, Bogor.
Toland, A.E. 2001. DNA Mutations. http://genetichealth.com/G101_ Changes_in_ DNA.shtml. Juni 2010].
Viljoen, G.J., L.H. Nel & J.R. Crowther. 2005. Molecular Diagnostic PCR Handbook. Springer, Dordrecht, Netherland.
Williams, J. L. 2005. The Use of Marker-Assisted Selection in Animal Breeding and Biotechnology. Rev. sci. tech. Off. int. Epiz., 24: 379-391.
Winarno & W. Agustinah. 2007. Pengantar Bioteknologi. M-Brio Biotekindo Press, Bogor.
Yahyaoui, M. H., A. Angoilillo, F. Pila, A. Sanchez, & J. M. Folch. 2003. Charaterization and genotyping of the caprine κ-Casein variants. J. Dairy Sci. 86: s2715-2720.
Lampiran 1. Sekuens Gen β-laktoglobulin yang diakses di GenBank (No. X14710)
LOCUS X14710 7877 bp DNA linear MAM 20-MAY-1992
DEFINITION B.taurus beta-lactoglobulin gene. ACCESSION X14710
VERSION X14710.1 GI:127
KEYWORDS Alu repetitive sequence; beta-lactoglobulin; transport protein.
SOURCE Bos taurus (cattle) ORGANISM Bos taurus
Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi;
Mammalia; Eutheria; Laurasiatheria; Cetartiodactyla; Ruminantia;
Pecora; Bovidae; Bovinae; Bos. REFERENCE 1
AUTHORS Alexander,L.J., Beattie,C.W., Hayes,G., Pearse,M.J., Stewart,A.F.
and Mackinlay,A.G.
TITLE Isolation and Characterization of the Bovine Beta-lactoglobulin
gene
JOURNAL Unpublished
REFERENCE 2 (bases 1 to 7877) AUTHORS Mackinlay,A.G.
TITLE Direct Submission
JOURNAL Submitted (16-MAR-1989) Mackinlay A.G., University of New South
Wales, School of Biochemistry, P.O.Box 1, Kensington, N.S.W. 2033, Australia COMMENT F100. FEATURES Location/Qualifiers source 1..7877 /organism="Bos taurus" /mol_type="genomic DNA" /db_xref="taxon:9913" /clone="lambda F100" /tissue_type="calf thymus" /clone_lib="lambda EMBL3A" TATA_signal 2143..2148 exon 2171..2305 /product="beta-lactoglobulin" /note="exon 1" exon 2975..3114 /product="beta-lactoglobulin" /note="exon 2" exon 3976..4049 /product="beta-lactoglobulin" /note="exon 3" exon 5167..5277 /product="beta-lactoglobulin" /note="exon 4" exon 5953..6057 /product="beta-lactoglobulin" /note="exon 5" exon 6279..6321
/product="beta-lactoglobulin" /note="exon 6" exon 6711..6893 /product="beta-lactoglobulin" /note="exon 7" polyA_signal 6869..6874 polyA_site 6893 /note="polyA site" repeat_region 7372..7877
/note="artiodactyl Alu-like repetitive sequence"
ORIGIN
1 gcgcttggaa aagattgtcc agcctccctc ccatagtggt cagtgccagc tgccccaggc 61 cagaggtgct ttatttccgt ctctctctct ggatggtatt ctctggaagc tgaaggttcc 121 tggaagttat gaatagcttt gccatgaagg gcatggtttg tggtcatggt tcacaggaac 181 ttgggagacc ctgcagctcg gacgttcctg aggttggtgg caccctgatt tcctaagctc 241 gctggggaac ggggtgctac ttctccctgg ctgacctccc tctgctctcg atcacccagt 301 tctgagagca gagtggtgct gggggcacag cctctcgcat ctgacacttg tgttcaaacc 361 acccatgctg gtgttcgggg ggccacctat ggggaaggct cctcactgca ggggtgcccc 421 tgtcccctga gagatcagaa gtcccagtct ggatgtcgaa tggccgagct ccctccagag 481 gctccaggga gggatccttg ccccctccgc cgccgcctcc agctcctggt gccgcaccct 541 tgggcccgat ctcgtagacg cctcagtcca gtctctgcct ccgtgttcac tggcattctc 601 cccatgtccc ctctgtgtcc ccgttttctc tcacaaggac accggacata agattagccc 661 ccgttccagc atcacctgaa cagctcacat ctgtaaagac ctagattcca aacaagattc 721 catcctgaag ttcctggtgg acgtgagttc tggagcgacg cccttcaacc ccatcacagc 781 ttgcggttca tcgcaaaaca cggaacctgg gatttatcgt aaaacccagg ttcttcgtga 841 aacactgagc ttcgaggctt gttgcaagaa ttaaaggtgc taatacagat cagggcaagg 901 accgaagctg gccaagcctc ctctttccat cacaggaaag ggaggtctgg gggcggccgg 961 gggtctgctc ccgtgggtgg gctctttctg gtacagtcac caacagtctc tccgggaagg 1021 aaaccagagg ccagagagca agccagagct agtctaggag atccctgagc ctccacccaa 1081 gatgccgacc agccagcggg ccccctggaa agaccctaca gtctaggggg ggaacaggag 1141 ccgacccgcc aggcccccgc tatcaggaga caccccaacc ttgctcctgt tcccctaccc 1201 cagtacgccc acccgacccc tgagatgagt ggtttacttg cttagaatgt caattgaagg 1261 cttttgtacc ccctttgcca gtggcacagg gcacccacag cccgctgggt actgatgccc 1321 atgtggactc agccaggagg actgtcctgc gccctccctg ctcgggcccc ctccatactc 1381 agcgacacac ccagcaccag cattcccacc actcctgagg tctgaaggca gctcgctgtg 1441 gtctgagcgg tgcggaggga agtgccctgg gagatttaaa atgtgagagg tgggaggtgg 1501 gaggttgggt cctgtaggcc ttcccatccc acgtgcctgc acggagccct agtgctactc 1561 agtcatgccc ccgcagcacc cctcaggtca ctttcccatc ctgggggtta ttatgactgt 1621 tgtcattgtt gttgcctttt tgctacccta actgggcagc gggtgcttgc agagccctcg 1681 atactgacca ggttcccccc tcggagctcg acctgaaccc catgtcaccc tcgccccagc 1741 ctgcagaggg tgaggtgact gcagagatac cctttaccca aggccacagt cacatggttt 1801 ggaggagatg gtgcccaagg cagaagccac cctccaggac acacctgccc ccagtgctgg 1861 ctctgacctg tccttgtcta agaggctgac cccagaagtg ttcctggcgc tggcagccag 1921 cctggaccca gagcctggac accccctgcg cccccacttc tggggcgtac caggaaccgt 1981 ccaggcccag agggggcctt cctgcttggc ctcgaatgga agaaggcctc ctattgtcct 2041 cgtagaggaa gcaaccccag ggcccaagga taggccaggg gggattcggg gaaccgcgtg 2101 gctgggggcc cggcccgggc tggctggctg gccctcctcc tgtataaggc cccgagcccg 2161 ctgtctcagc cctccactcc ctgcagagct cagaagcgtg accccagctg cagccatgaa 2221 gtgcctcctg cttgccctgg ccctcacttg tggcgcccag gccctcattg tcacccagac 2281 catgaagggc ctggatatcc agaaggttcg agggtgcccg ggtgggtggt gagttcgagg 2341 gctggctggg gagctgggcc tcagagacca agggaggctg tgacgtctgg gattcccatc 2401 agtcagctag agccgcctga caaatcgccc gccacaggct tcaaccaggc ctttagtgtc 2461 ttgcattctg gaggctggaa gctgcaatcc gggcatcggc ccagctggct tctcctgcgg 2521 ccactctccg gggagcagac agccatcttc tccctgtgtc ctttgcgtgc cctggtttcc 2581 tcttcctgtg aggtcaccag gcctgctgga tccacgcccg cccacacagc ctcacgtaac 2641 ctttgtcatc tctttaaagg ccgtgtctcc agtcctgtgt tgaggttctg ggggttaatg 2701 ggacacagtt cagcccctaa aagagtccgc tctgcccctc aaattttccc cacctccagc 2761 tatggtctcc ccaagatcca aatgttgcca cgtgtgcggg ggctcatctg ggtccctctt
2821 tgggctcaga gtgagtctgg ggagagcatt cctcagggtg ccgagttggg gggaggcatc 2881 tcagggctgc ccaggccagg gtgggacaga gagcccactg tggggctggg ggccccttcc 2941 cgcccctgga gtgcagctca aggtccctcc ccaggtggcg gggacttggt actccttggc 3001 catggcggcc agcgacatct ccctgctgga cgcccagagt gcccccctga gagtgtatgt 3061 ggaggagctg aagcccaccc ctgagggcga cctggagatc ctgctgcaga aatggtgggc 3121 gtccccccca aaaaaagcat ggaaccccca ctccccaggg atatggaccc cccggggtgg 3181 ggtgcaggag ggaccagggc cccagggctg gggaacgggg cttggagttt cctggtaccc 3241 ctggaggtcc acccaaggct gcttatccag ggctttctct ttcttttttt cccccaactt 3301 ttattaattt gatgcttcag aacatcatca aacaaatgaa cacaaaacat cattttcgtt 3361 aacttggaag gggagataaa atccactgaa gtggaaatgc ataggaaaga tacatacagt 3421 aaggcaggta ttctgaattc gctgttagtt tgaggattac aaatgcactt gagcaacaga 3481 gagacgtttt cattatttct cggtctgaac agctcagtat ctaaaatgaa caagatgtca 3541 tggagacaaa gccggcgggg gagaggcccg tgtgaaggcc gctgggcgtg cagacctggg 3601 tcctcggggc ccaggcagtt cccactacca gccctgtcca ccctcagacg ggggtcagag 3661 tgcaggagag agctgggtgg gtgtggggca gagatgggga cctgaacccc aggactgcct 3721 tttggggtgc ctgtggtcaa ggctctcccc aaccttttct ccctggctcc atctgacttc 3781 tcctggccca tccacccggt cacctgtggc cccagaggtg acagtgagtg cagccaaggc 3841 cggttggcca gccggccccc tatgcccacg ccacccgcct ccagcccctc ctggggccgc 3901 cttctgcccc tggccctcag ttcatcctga tgaaaatggt ccatgcccgt ggctcagaaa 3961 gcagctgtct ttcagggaga acggtgagtg tgctcagaag aagatcattg cagaaaaaac 4021 caagatccct gcggtgttca agatcgatgg tgagtgctgg gtccccaggg gacgcccacc 4081 accccccagg gactgtgggc aggtgcaggg ggctggcgtc aggccccgag atgctaaggg 4141 gctggtggtg atgaagacac tgccgtgcca cctgcttccc tggcctccct gccacctgcc 4201 cggggccttg ggccggtggc cgtgggcagg tcccggctgg gcaggtctga caccccaggg 4261 tgacacccga gctctctttg ctgagggtgg ggtggtgctc ggggccctca ggctgagctc 4321 aggaggtccc tgtgcccacc caggggtaac cgagagccgc tgcccgctcc aggggtccag 4381 gtgccccacg atcccagccc accccacggc tccttcatct cctgaagacg aactctgtcc 4441 gccctcgctc attcacttgt ttgtcctaaa tccaagatga gaaagcttcg aggtggggtt 4501 ggggttccat cagggcctgc ccttccgccg ggcagcctgg gccacatctg cccttggcct 4561 cttcaggact cactctgact ggaggccctg cactgactga tgccagggtg cccagcccag 4621 ggtctcctgt gccatccggc tgcacggggt ttggatgctg gtcctgcccc caagctgccc 4681 agacactgca gggcagctgg ggccacccgc aggcctcggt cagggagagc cccagctgcc 4741 cccgctcagc gctgcccccc aacaattccc cagtcctcag gacgcatccc tcttcccttg 4801 ctgggcagtg ttcagcccca cccgagatcg gggaagccct atttcttgac cactccggtc 4861 cctggggagg gcggcctcag actcagtggt gagtgttccc aagtccagga ggtggtggag 4921 ggtccctggc ggatccagag ttgggcttcc agagtgaggg cttcctgggc cccatgtgcc 4981 tggcagtggc agcagggaag gggccacacc attttggggc tgggggatgc cagagggcgc 5041 tccccacccc gtcctcacca agtggtgacc ccgggggagc cccgctggtt gtggggggtg 5101 ctgggggctg accagaaacc cccctcctgc tggaactcac tttcctcccg tcttgatctc 5161 ttccagcctt gaatgagaac aaagtccttg tgctggacac cgactacaaa aagtacctgc 5221 tcttctgcat ggagaacagt gctgagcccg agcaaagcct ggcctgccag tgcctgggtg 5281 ggtgccaacc ctggctgccc agggagacca gctgtgtggt cctcgctgca acggggccgg 5341 gggggacggt gggagcaggg agcttgattc ccaggaggag gagggatggg gggtccccga 5401 gtcccgccag gagagggtgg tcatataccg ggagccggtg tcctgggggc ctgtgggtga 5461 ctggggacgg gggccagaca cacaggctgg gagacggggg gctgcagcgc tctggtgtga 5521 ccatcacgat ggagccggcg gtcactatga atctaacagc ctttgttacc ggggagtttc 5581 aattatttca tcaaataaga actcaggcac aaagctgtct ttcaactgtc acgtcctgaa 5641 aacaaatggc aggtgacatt ttccatgcca tagcagtgcc actgggcatt ttcagggccc 5701 atgtgccagg agggcgtggg catcggcgag tggaggctcc tggccgtgtc agctggccca 5761 gggggaggag gggacccaga cagccagagg tggggagcag gctttccccc tgtgacgctg 5821 cagacccacc gcactgccct gggaggaagg ggagggaact gggccaaggg ggaagggcag 5881 gtgtgctgga ggccaaggca gacctgcaca ccaccctgga gagcaggggt tgaccccgtc 5941 ccggccccac agtcaggacc ccggaggtgg acgacgaggc cctggagaaa ttcgacaaag 6001 ccctcaaggc cctgcccatg cacatccggc tgtccttcaa cccaacccag ctggagggtg 6061 agcacccagg ccccaccctg ctcctggggc aggaagccac ccggcccagg accacctcct 6121 cccatggtga cccccagctc cccaggcctc ccgggaggat ggagacgggg tgcagggccc 6181 cgaggtggcc ccctccccac cccctcccca gctccctctg tcctggggtg tccagtccca 6241 tcctgacgct cccccgccac ggctctccct cccccacaga gcagtgccac atctaggtga 6301 gcccctgccg gcgcctctgg ggtaagctgc ctgccctgcc ccacgtcctg ggcacacaca
6361 tggggtaggg gttcttggtt gggcccggga gcccccatta ggccctgggg tcccccgtag 6421 gaatggctgg aagctggggt ccttcctgga gactacagag ccggctggcc acatgctcgc 6481 tcttgtgggg tgacctgtgt cctggcctca ctcacacgct gatctcctcc acctccttcc 6541 tggcagacct aagggccaag gtggagggct caggaagtga cacctaaggg ggaggctagg 6601 ggggtccttc tcccaaagga ggggccgtcc tgaatcccca gccacggaca ggctggcaag 6661 ggtctggcag gtaccccagg aatcacaggg gagccccatg tccatttcag agcccgggag 6721 ccttggcccc tctggggaca gacgatgtca tccccgcctg ccccatcagg ggaccaggag 6781 gaaccgggac cacattcacc cctcctggga cccaggcccc tccaggcccc tcctggggcc 6841 tcctgcttgg ggccgctcct ccttcagcaa taaaggcata aacctgtgct ctcccttctg 6901 agtctttcct ggatgatggg ccgggggtgg agaaggcccg ggacagggtg gggagtggtc 6961 tggctcagag gatgatggta gggctgggat ccagggcgtc tgcattacag tcttgtgaca 7021 tctgggggcc cacacacatc tacggctctt tgaaactttc aggaaccagg tagggagtcg 7081 gcagagacat ctgccagtta acttggagtg ttcagtcaac acccaaactc gacaaaggac 7141 aggaagtgga aaatggctgc tcttagtcta ataaatattg atatgaaaac tcaagttgct 7201 catggatcaa attatgcctt tttatgaatc cagccactac agtcggtatc aaacttcatg 7261 tactcaaaac gcactgatct tttctgtgct aaaatgaaat aaagagattt ccccaagata 7321 caggagctgg gcaaaagagg tcacggttgg aaggggactt gttctgcaca cacagcaagg 7381 agatccagcc agtccatcct aaaggagatc agtcctgggt gttcattgga gggactgatg 7441 ttgaagctga aactccaata ctttggccac atgatgcgga gagctgactc atttgaaaag 7501 accctgatac tgggaaagat tgagggcagg aggagaaggg gatgacagag gatgagatgg 7561 ttggatggca tcaccgacaa aatggacatg ggtttgggtg gactccagga gttggtgatg 7621 gacagaaagg cctggcatgc tgcggtccat ggggtcacaa agagtcggac acgactgagc 7681 gactgaactg aactgaactg aatggaaaag aggtatacag caacgtgggg attttttaga 7741 taataagaat aatttacgca taacatagtg tatactcata tttatatata tacgtgaatg 7801 ctcagtcaca ctcagtcata tctgactctg tgacccatgg actgtagccc tccaggctcc 7861 ttctgtccac agaattc