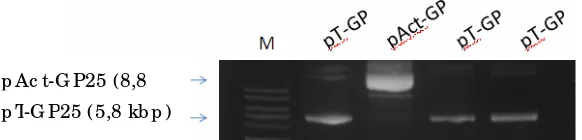
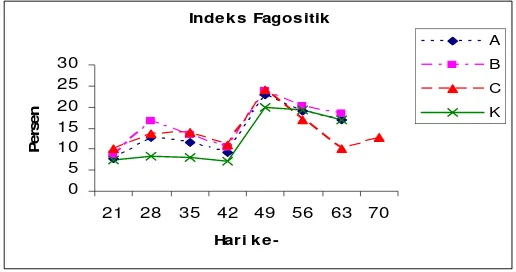
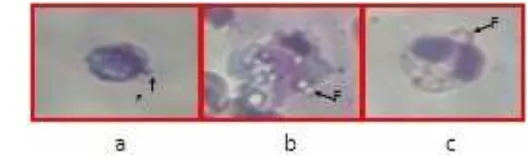
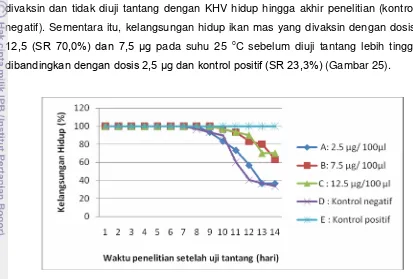
Pengembangan Vaksin DNA Penyandi Glikoprotein Virus KHV (Koi Herpesvirus) Menggunakan Isolat Lokal
Teks penuh
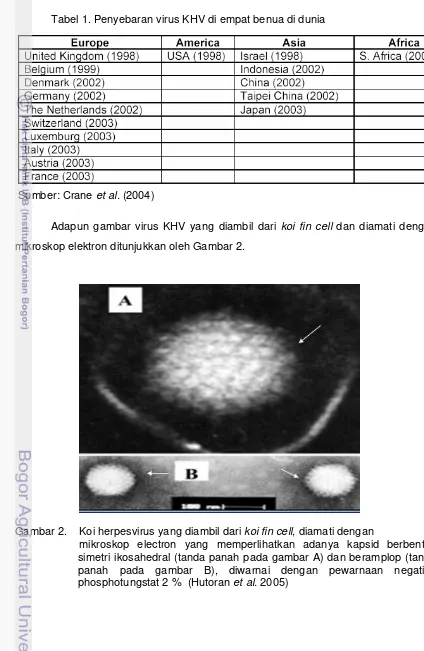
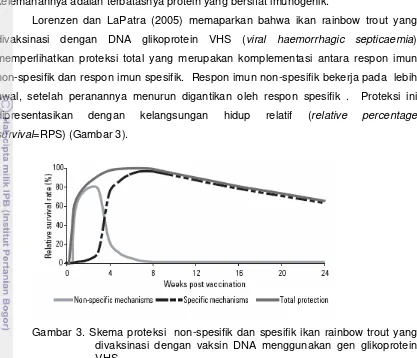
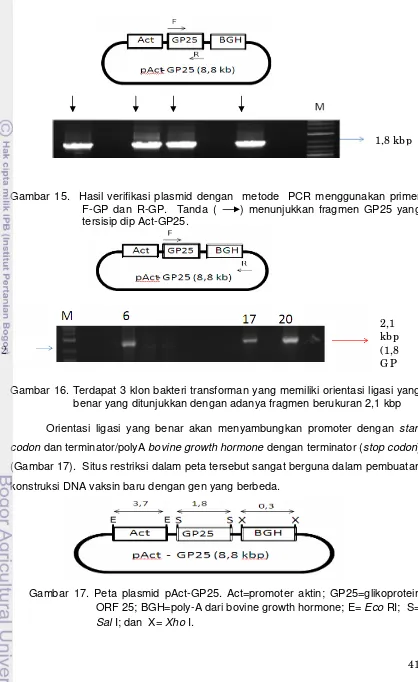
Gambar


Dokumen terkait
Boru (bagi isteri) dengan sisipan nama cucunya yang tertua (laki-laki ataupun perempuan) dari anaknya laki-laki ataupun perempuan yang sudah mempunyai keturunan, sampai
Film mempunyai media komunikasi sosial yang terbentuk dari penggabungan dua indra, penglihatan dan pendengaran, yang mempunyai inti atau tema sebuah cerita banyak
4.6 Cara Tindak Tutur yang Diucapkan pada Upacara Perkawinan Batak Toba Pemakaian bahasa merupakan suatu hal yang penting dalam pembahasan penelitian ini dalam acara
Untuk membicarakan beban apa saja yang harus dipikul, biasanya makan waktu bertele-tele, sebab akan terjadi pembicaraan yang sambung- menyambung, sehingga kalau
Oleh karena itu, dapat dikatakan bahwa tradisi adalah kebiasaan yang dilakukan oleh suatu masyarakat etnik dan diturunkan secara turun-temurun kepada generasi setelahnya dengan
Dengan konfigurasi ini nampaknya sangat mudah untuk mencapai puncak yang rata pada pass bandnya, dan sekaligus mempunyai bentuk lereng yang mendekati symetris, sehingga mudah
Jika suatu graph diberi label pada setiap simpul dan sisi dengan bilangan sebanyak simpul dan sisi, maka graph tersebut mempunyai sifat total sisi ajaib jika label pada setiap sisi
Analisa data kandungan logam berat timbal (Pb) diuji secara statistik menggunakan SPSS uji T, masing-masing terhadap : 1) Kandungan logam berat timbal (Pb) air
