Oseana, Volume XXI, Nomor 4, 1996 : 43 - 59 ISSN 0216-1877 MAKANAN DAN CARA MAKAN BERBAGAI JENIS TERIPANG
oleh
Aznam Aziz *)
ABSTRACT
FOOD AND FEEDING HABITS OF VARIOUS SPECIES OF HOLOTHU-RIANS. Holothurians usually utilize three sources of food : detritus, the organic contents of mud, and plankton. The deposit-feeders collect sediment and particles with their buccal tentacles on soft or hard bottom, on soft bottoms the tentacles dig into the sediment, or take only on the surface layer. On hard bottoms the tentacles sweep and pick up particles from the surface. The food of suspension-feeding holothuroids is composed of plankton and suspended organic matter. Most of aspidochirote holothuroids are deposit feeders, and some of dendrochirote holothuroids are suspension-feeders. The digestive tract is prominent within the body cavity, extending from the mouth to the anus. Commonly their consist of pharynx, oesophagus, stomach, large intestine, small intestine, rectum, cloaca, and anus. Digestion efficiency as well as feeding effects on their habitat will be discussed in this article.
PENDAHULUAN
Teripang adalah salah satu kelompok ekhinodermata yang mempunyai arti ekonomi penting. Biota ini merupakan salah satu produk perikanan penghasil devisa negara. Untuk menunjang usaha pembudidayaannya, maka pengetahuan mengenai macam makanan dan cara makan dari biota ini perlu diketahui. Pada kelompok teripang dikenal dua macam cara makan, yaitu deposit feeder atau pemakan endapan pada hampir sebagian besar anggota
kelompok, suspension feeder atau pemakan materi tersuspensi (termasuk plankton) hanya p a d a s a t u b a n g s a ( o r d o ) , y a i t u Dendrochirotida.
Teripang pemakan endapan sangat tergantung kepada kandungan zat organik dalam lumpur, detritus, organik debris, dan plankton. Yang disebutkan belakangan ini adalah dalam jumlah yang terbatas. Sedangkan teripang pemakan materi tersuspensi, sangat tergantung kepada kehadiran plankton dan partikel-partikel melayang (seston).
Tipe dari tentakel bukal sangat menentukan macam makanan dari teripang. Pada kelompok teripang dikenal lima macam tipe tentakel bukal. Tentakel bukal ini terdapat di seputar mulut dan merupakan alat yang penting untuk menangkap makanan. Dari hasil penelitian para pakar diketahui 4 macam tipe tentakel bukal (peltate, digitate, pinnatae, dan semidendrit), adalah merupakan ciri dari teripang pemakan endapan. Sedangkan tentakel bukal tipe dendritik, termodifikasi untuk menangkap plankton langsung dari air laut di sekitarnya. Tentakel bukal tipe dendritik ini hanya dimiliki oleh teripang bangsa Dendrochirotida. Dari sudut pandang sistematik, tipe tentakel bukal dapat pula dipakai sebagai pemisah pada tingkat ordo (bangsa).
Teripang komersial pada umumnya termasuk kedalam bangsa Aspidochirotida, yang telah dibuktikan sebagai pemakan endapan. Banyak kepustakaan di Indonesia yang menyangkut penulisan tentang teripang komersial, dengan latah mencantumkan sederetan plankton yang hadir di lokasi penelitian yang dianggap mempunyai relevansi dengan makanan teripang tersebut. Upaya pemberian pupuk kandang, dedak, pupuk organik, pada budidaya teripang bertujuan untuk memperkaya kandungan zat organik pada substrat pemeliharaan teripang dewasa. Hal ini jelas memperlihatkan secara tidak langsung bahwa, teripang komersial adalah termasuk kedalam teripang pemakan endapan. Tetapi hal ini bertentangan dengan kebanyakan pakar kita yang menghubungkan teripang dengan kehadiran plankton di sekitarnya. Pada tulisan ini lebih lanjut akan diuraikan secara garis besar mengenai sistematika, tentakel bukal, macam makanan, cara makan, sistem pencernaan, sistem enzim, dan pengaruh cara makan terhadap lingkungan.
Semoga tulisan yang pendek ini dapat menambah pengetahuan kita.
SISTEMATIKA DAN PENYEBARAN
Teripang ditemukan pada semua laut dan lautan, dengan batas kedalaman antara 0 sampai dengan 10.000 m. Mereka dapat hidup bebas sebagai epifauna baik menyendiri ataupun hidup dalam kelompok. Beberapa anggota dari kelompok teripang i n i mempunyai kebiasaan membenamkan diri dalam lumpur dan pasir, atau bersembunyi di bawah batu dan karang. Kelas Holothuroidea yang hidup saat ini diperkirakan sekitar 900 jenis. Mereka dapat beradaptasi dengan bermacam-macam habitat yang meliputi lumpur, pasir, batu, koral, padang lamun, dan daerah pertumbuhan algae (HYMAN 1955, PAWSON 1966).
Di perairan Indonesia diperkirakan terdapat sekitar 200 jenis teripang. Untuk kedalaman 0 m sampai dengan 20 m di Indonesia dilaporkan terdapat sekitar 155 jenis teripang (CLARK & ROWE 1971). Faktor penting yang menghalangi penyebaran teripang adalah salinitas dan suhu. Teripang hidup pada batasan pergoyangan salinitas air laut normal (30 - 34 permil), tetapi beberapa jenis di antaranya bisa bertahan sampai dengan salinitas sekitar 21 permil (BAKUS 1973). Teripang yang hidup di laut jeluk dan di daerah ugahari lebih toleran terhadap suhu rendah. Penyebaran teripang muda (juvenile) diduga dibatasi oleh kehadiran senyawa methan dan hidrogen sulfida yang dihasilkan dari proses pembusukan (YAMANOUTI dalam BAKUS 1973). Diduga teripang muda ini tidak toleran terhadap dua zat pencemar tersebut.
Klasifikasi jenis-jenis teripang, seperti yang dianjurkan oleh FRIZZEL et al (1966),
adalah sebagai berikut : Kelas : Holothuroidea Anak kelas : Dendrochirotacea Bangsa : Dendrochirotida
Dactylochirotida Anak kelas : Aspidochirotacea Bangsa : Aspidochirotida
Elasipoda Anak kelas : Apodacea Bangsa : Apodida
Molpadiida
Teripang yang mempunyai nilai ekonomi penting termasuk kedalam bangsa Aspidochirotida, dengan dua suku utamanya, yaitu Holothuriidae dan Stichopodidae. Pada umumnya teripang komersial menempati perairan dangkal kurang dari 30 meter.
TENTAKEL BUKAL
Pada pangkal faring dari teripang, terdapar semacam cincin kapur yang diduga homolog dengan organ lentera Aristoteles pada kelompok bulu babi (HYMAN 1955). Cincin kapur ini berperan dalam memperkuat kedudukan faring. Cincin tersebut pada umumnya dibangun oleh 5 keping radial dan 5 keping interradial. Kadang-kadang jumlahnya lebih dari 10 keping, dan biasanya sesuai dengan jumlah tentakel bukal. Tentakel bukal bertumpu pada cincin kapur ini. Dengan koordinasi kerja otot, cincin kapur dan tentakel bukal dapat ditarik kedalam mulut atau diulurkan mendekati bibir mulut. Selain itu setiap tentakel bukal dapat memendek dan memanjang, berkat kerja otot yang berada dalam struktur dindingnya. Hal ini terlihat jelas sewaktu aktifitas makan sedang berlangsung.
Menurut HYMAN (1955), tentakel bukal dan kaki tabung mempunyai struktur umum yang sama dengan struktur dinding tubuh teripang tersebut. Tetapi dalam hal
struktur rinci, kaki tabung, tentakel bukal dan dinding tubuh memperlihatkan susunan yang berbeda. Baik kaki tabung, ataupun tentakel bukal berhubungan dengan sistem pembuluh air. Dalam hal ini pembuluh air dan sistem otot secara sinergik mengontrol pergerakan kaki tabung dan tentakel bukal.
Selanjutnya dilaporkan bahwa kaki tabung dan tentakel bukal juga berperan sebagai organ sensor yang peka terhadap rangsang kimia dan rangsang mekanik. Menurut BOULAND et al. (1982), tentakel bukal merupakan modifikasi dari kaki tabung dengan bagian ujung (distal)nya yang sangat terspesialisasi sebagai komponen sensor. Fungsi utama dari tentakel bukal adalah untuk mengumpulkan partikel-partikel makanan yang kaya akan kandungan zat organik.
ROBERTS (1982), menyebutkan bahwa pada kelompok teripang dikenal lima macam tipe tentakel bukal (Gambar 1), yaitu: 1. Digitate atau tentakel berbentuk jari 2. Pinnate atau tentakel berbentuk sikat 3. Peltate atau tentakel berbentuk perisai 4. Semidendrit atau peltato-dendrit, i.e.
modifikasi antara bentuk perisai dan bentuk pohon
5. Dendritik atau berbentuk pohon
Selanjutnya dikatakan, bahwa tentakel bukal tipe peltate adalah merupakan bentuk yang paling primitif. Tentakel bukal tipe peltate ini dimiliki oleh teripang bangsa Aspidochirotida dan bangsa Elasipoda dan merupakan teripang pemakan endapan.
Perubahan bentuk tentakel tipe peltate ke arah tipe pinnate, dianggap sebagai arah kemajuan dimana terjadi peningkatan efisiensi dalam pengumpulan partikel makanan, sehubungan dengan bertambahnya luas permukaan tentakel tersebut. Tentakel bukal tipe pinnate dimiliki oleh teripang bangsa Apodida.
Gambar 1. Tipe tentakel bukal pada berbagai jenis teripang Keterangan : 1. tipe dendritik
2. tipe pinnate 3. tipe digitate 4. tipe digitate 5. tipe peltate 6. tipe digitate
Tentakel bukal berbentuk jari (digitate) dimiliki oleh teripang bangsa Dactylochirotida dan sebagian teripang bangsa Apodida. Tentakel tipe digitate ini dianggap sebagai penyederhanaan secara sekunder dari tipe dendritik dan secara simultan juga bisa dipandang sebagai modifikasi dari tentakel tipe pinnate. Terjadinya perubahan bentuk tentakel ini diduga erat hubungannya dengan perubahan habitat dan perubahan macam makanan. Sebagai contoh beberapa anggota bangsa Aspidochirotida dengan tipe tentakel peltate termodifikasi menjadi peltato-dendrit. Dalam hal ini pertambahan jumlah percabangan dari ujung tentakel bukal dipandang sebagai peningkatan efisiensi untuk menangkap partikel-partikel melayang dalam medium air laut di sekitarnya.
Hipotesa selanjutnya terjadi perubahan dari semidendrit ke arah tentakel dendritik. Dalam kondisi ini teripang tidak lagi bersifat pemakan endapan, tetapi lebih bersifat pemakan partikel melayang. Tentakel bukal dendritik (bentuk pohon), dimiliki oleh teripang bangsa Dendrochirotida. Sebagian dari anggota bangsa Dendrochirotida juga beradaptasi untuk hidup melayang sebagai megalo plankton.
Sebagian besar teripang yang hidup saat ini terutama bersifat pemakan endapan (deposit feeders), dengan tipe tentakel bervariasi antara peltate, digitate, pinnate, dan peltato-digitate. Dari sudut pandang taksonomi, tentakel bukal ini dapat dipakai sebagai pembeda di tingkat subkelas dan bangsa (ordo).
Pada saat ini studi khusus mengenai struktur dan fungsi dari tentakel bukal masih sangat terbatas. FANKBONER (1978, 1981), telah melaporkan tentang peranan dan fungsi
dari tentakel bukal dari teripang kelompok Dend rochir oti da. ROBE RTS ( 197 9 ), ROBERTS & BRYCE (1982), BOULAND et al. (1982), dan CAMERON & FANKBONER (1984), telah melaporkan tentang fungsi dan struktur dari tentakel bukal dari kelompok Aspidochirotida. Selanjutnya peran dan fungsi dari tentakel bukal tipe pinnate dari suku Synaptidae telah dilaporkan oleh ROBERTS (1979).
Tentakel bukal tipe peltate secara umum dikatakan berbentuk perisai, tetapi pada pengamatan dengan mikroskop elektron ternyata bahwa bentuk umum dari tentakel tipe peltate ini lebih mirip dengan miniatur kembang kol (BOULAND et al. 1982). Bagian paling ujung dari tentakel bukal berbentuk tonjolan-tonjolan kecil yang disebut sebagai nodul atau nodular (Gambar 2). Tonjolan-tonjolan ini baik secara mekanis ataupun dengan bantuan mukosa berperan penting dalam menangkap partikel-partikel makanan dari substrat di sekitarnya. Ada kecenderungan nodular ini berkemampuan untuk menyeleksi ukuran partikel ataupun menyeleksi kandungan zat organik dalam partikel tersebut. Hal ini tampaknya tergantung kepada jenis dari teripang dan habitat tempat hidupnya (ROBERTS 1979, ROBERTS & BRYCE 1982, dan BOULAND et al. 1982).
BOULAND et al. (1982), melaporkan tentang struktur dan fungsi tentakel bukal dari teripang jenis Holothuria forskali yang hidup di Laut Tengah. Tentakel bukal tipe peltate terdiri dari tangkai dan perisai (Gambar 2). Bagian tangkai sebelah ujungnya terbagi kedalam 10 sampai 15 percabangan. kemudian setiap cabang ini terbagai lagi menjadi cabang-cabang yang lebih kecil. Setiap ujung percabangan akan berakhir pada sekumpulan papila yang disebut sebagai nodular.
Gambar 2. Diagram tentakel bukal tipe peltate Keterangan 1. 2. P. T. bagian perisai bagian tangkai papilla tunas
LA. lumen ambulakral
Di bawah mikroskop elektron kumpulan papila akan berbentuk seperti miniatur kembang kol. Setiap papila mempunyai tangkai yang relatif pendek dan berakhir pada ujung yang berbentuk tunas (bud), biasanya 2 sampai 5 tunas. Secara rinci bentuk permukaan tunas (bud) adalah seperti sikat (brushlike). Bagian dari tunas (bud) tersebut dibangun oleh tiga macam sel, yaitu
sel mukus, sel epitel bersilia, dan sel "GV". Sel "GV" adalah sel yang bersifat kemosensitif. Ketiga macam sel ini secara bersama berperan dalam mendeteksi partikel- partikel makanan (BOULAND et al. 1982). Berbeda dengan bangsa Aspidochirotida lainnya, teripang jenis Holothuria cinerascens mempunyai tentakel bukal dengan tipe semidendrit atau peltato-
dendrit. Dalam hal ini tentakel tersebut juga termodifikasi untuk menangkap partikel-partikel makanan dari air laut di sekitarnya (ROBERTS & BRYCE 1982). Anggota suku Synaptidae (Ophiodesoma grisea) sering terlihat berada di antara daun lamun atau algae. Tentakel bukal tipe pinnate dari hewan ini berperan aktif mengumpulkan partikel makanan yang menempel pada daun lamun ataupun algae (ROBERTS 1979). Teripang jenis Bohadschia graffei aktif menyapukan tentakel bukalnya pada permukaan koloni spons untuk mengumpulkan partikel-partikel makanan (BIRKELAND 1989). Sebelumnya tingkah laku mengumpulkan partikel makanan dari tumbuhan bentik, juga telah dilaporkan oleh BERRILL (1966). Teripang jenis Ophiodesoma spectabilis yang hidup di perairan sekitar Hawaii aktif mengumpulkan partikel makanan yang menempel di daun algae marga Sargassum.
Tentakel bukal tidak saja peka terhadap rangsang kimia dan mekanik, tapi juga peka terhadap rangsang cahaya. Teripang jenis
Ophiodesoma spectabilis peka terhadap intensitas cahaya yang kuat. Hewan ini akan bergerak menghindar dan mencari tempat yang terlindung dari cahaya matahari (BERRILL 1966).
SISTEM PENCERNAAN
Secara umum sistem perncernaan pada semua kelompok teripang dibangun oleh unit yang sama, yaitu terdiri dari mulut, faring, esofagus, lambung, usus, rektum, kloaka, dan anus (HYMAN 1955, FERAL & MASSIN 1982). Mulut berbentuk bulat atau sedikit oval, terletak di tengah selaput bukal dan dikelilingi oleh sederetan tentakel bukal. Mulut dilingkari oleh semacam otot polos yang
diduga berfungsi sebagai bibir. Faring dan esofagus relatif pendek, dan esofagus kadang-kadang tereduski. Faring melekat dengan erat ke dinding tubuh dengan bantuan suspensor yang terdiri dari otot dan jaringan ikat. Lambung ditandai oleh kehadiran otot polos yang membesar, tetapi pada beberapa anggota Holothuroidea pembesaran otot polos ini tidak begitu jelas. Selain dari pembesaran otot polos, lambung pada awal dan akhirnya juga ditandai oleh semacam penyempitan (contriction). Usus merupakan bagian saluran pencernaan yang paling panjang. Kurang lebih 2 sampai 3 kali panjang total tubuh. Usus tersusun berbelok dalam tubuh, mula-mula usus berjalan memanjang sepanjang sisi middorsal ke arah belakang, kemudian membelok kembali ke arah depan melalui sisi tubuh sebelah kiri. Setelah sedikit melewati faring, usus ini berbelok kembali ke arah belakang sepanjang sisi midventral. Bagian usus paling belakang sarat dengan makanan yang ditelan, bagian ini kadang-kadang disebut sebagai usus besar, dan bagian sebelumnya disebut sebagai usus halus. Usus akan berlanjut ke bagian rektum dan kloaka. Rektum dan kloaka mempunyai struktur dinding yang tebal (Gambar 3). Kloaka melekat ke bagian posterior tubuh dengan bantuan suspensor (kloakal-suspensor). Ukuran dan bentuk dari komponen saluran makanan ini bervariasi dari jenis ke jenis lainnya.
Secara umum saluran pencernaan dibangun oleh 5 lapisan, yaitu : lapisan epitel, lapisan jaringan penyokong dalam, lapisan otot polos dan otot bergaris, lapisan jaringan penyokong luar, dan lapisan peritoneum bersilia. Lapisan peritoneum ini kadang-kadang absen pada sebagian anggota kelompok teripang. Sel-sel kelenjar dan sel-sel mukosa yang berperan penting dalam pencernaan terdapat di antara sel-sel epitel.
Posisi anus pada teripang juga bervariasi, biasanya terletak di ujung posterior dari tubuh. Pada bangsa Elasipoda anus terletak pada posisi posterio-ventral dari ujung tubuh. Sedangkan pada bangsa Dactylochirotida, sebagian anggota Aspidochirotida dan Dendrochirotida, mempunyai anus yang terletak di ujung poterio-dorsal.
Sebagaimana lazimnya pada biota lainnya, gerak partikel makanan dalam saluran pencernaan dimungkinkan oleh daya peristaltik. Daya peristaltik timbul akibat gerakan dinding saluran pencernaan yang fungsinya dikoordinasikan oleh kerja otot polos yang dibantu oleh otot bergaris (MASSIN 1980, FERAL & MASSIN 1982).
MACAM MAKANAN
PAWSON (1966), menyimpulkan ada tiga macam sumber makanan bagi teripang, yaitu : kandungan zat organik dalam lumpur, detritus, dan plankton. Teripang primitif dan teripang yang hidup di laut jeluk, lebih tergantung kepada detritus dan kandungan zat organik dalam lumpur. Dalam hal ini tipe tentakel bukal sangat erat kaitannya dengan macam makanan. Tentakel bukal dengan tipe sederhana, dan berukuran relatif pendek dengan luas sapuan yang sangat terbatas (tipe peltate, digitate, dan pinnate) merupakan ciri dari teripang pemakan endapan (deposit feeder).
Mengingat pergerakan teripang yang relatif sangat terbatas, dan dengan kemampuan gerak yang sangat lamban, menyebabkan biota ini tidak mungkin bertindak sebagai hewan predator atau karnivora. Tidak adanya alat kunyah dan pemotong seperti organ Aristoteles pada kelompok bulu babi, juga mengakibatkan tidak mungkin menjadi hewan herbivora dan menyebabkan biota ini sangat
tergantung kepada substrat di sekitarnya.
Teripang bangsa Aspidochirotida, Elasipoda, Apoda, Molpadiida, dan Dactylochirotida mempunyai 10 sampai 20 tentakel bukal yang berukuran relatif kecil dan mempunyai bentuk yang sederhana. Biota ini pada umumnya bersifat deposit feeder, dengan sumber makanan utamanya berupa kandungan zat organik dalam lumpur, detritus dan plankton (PAWSON 1966, YINGST 1976, 1982, LAMBERSON 1978, HAUKSSON 1979, KHRIPOUNOFF & SIBUET 1980, MASSIN 1982, COULON & JANGOUX 1993 dan KUNGER et al. 1994). Selanjutnya MORIARTY ( 1 9 8 2 ) , menambahkan bahwa selain kandungan zat organik dalam lumpur, teripang juga tergantung kepada massa bakteri yang terdapat dalam pasir. Selain lumpur, detritus, bakteri, beberapa biota berukuran kecil juga dimanfaatkan sebagai makanan, seperti fora-minifera, plankton, dan potongan-potongan kecil dari hewan ataupun tumbuhan laut lainnya (organik debris). Tetapi makanan yang disebutkan belakangan ini bukanlah menjadi sumber makanan utama.
Teripang bangsa Dendrochirotida, dianggap lebih maju dalam sudut pandang evolusi. Teripang ini mempunyai tentakel bukal tipe dendritik. Tentakel bukal tipe dendritik mempunyai percabangan berbentuk pohon dan berukuran relatif lebih panjang, sehingga mempunyai daya sapuan yang lebih luas. Berbeda dengan teripang yang diceritakan terdahulu. kelompok Dendrochirotida ini bersifat suspension feeder, dan hidup dari memakani plankton dan partikel tersuspensi (seston). Dalam hal ini plankton merupakan makanan utamanya (PAWSON 1966, FANKBONER 1978, MASSIN 1982, dan BIRKELAND 1989). Dari hasil penelitian FANKBONER ( 1 9 8 1 ) , ternyata bahwa tidaklah semua anggota kelompok
Dendrochirotida bersifat suspension feeder. Teripang jenis Leptopenctata elongata yang hidup di perairan Inggris ternyata bersifat sebagai deposit feeder atau pemakan endapan. Teripang ini selalu berada dalam lumpur dengan bagian anusnya menghadap ke arah permukaan substrat. Pada lubang di atasnya terlihat adanya tumpukan fecal pellet yang menunjukkan hewan ini melakukan aktifitas makan dengan normal. Sumber makanan teripang ini adalah kandungan zat organik yang terdapat dalam lumpur.
B e b e r a p a a n g g o t a b a n g s a Aspidochirotida mempunyai tentakel bukal yang merupakan modifikasi antara tipe peltate dan tipe dendritik. Tentakel bukal seperti ini disebut sebagai semidendrit atau peltatodendreit. Di samping memakani lumpur di sekitarnya. teripang ini juga aktif menangkap plankton langsung dari air laut di sekitarnya. Dua jenis teripang yang termasuk ke dalam kelompok ini, yaitu Holothuria
cinerascen dan Actinopyga mauritiana.
Teripang ini memilih hidup di kaloran terumbu karang yang relatif berarus kuat (ROBERTS & BRYCE 1982 dan BIRKELAND 1989).
Para pakar sependapat bahwa teripang pemakan endapan tidak selektif memilih partikel yang kaya akan kandungan zat organik. Ukuran partikel dalam saluran pencernaan relatif sama dengan ukuran partikel medium di sekitarnya. Kandungan zat organik dalam saluran pencernaan dan dalam fecal pellet, jauh lebih tinggi dari kandungan zat organik dalam lumpur di sekitarnya (BONHAM & HELD 1963, PAWSON 1966, YINGST 1976, 1982, KHRIPOUNOFF & SIBUET 1980, HAUKSSON 1979, MASSIN 1982, HAMMOND 1982, 1983, MORIARTY 1982, BIRKELAND 1989 dan KLINGER et al. 1994).
CARA MAKAN
Selain mengandalkan kemampuan tentakel bukal untuk mengumpulkan partikel makanan, teripang pemakan endapan juga mempunyai kemampuan untuk menelan lumpur dan pasir di sekitarnya secara langsung (PAWSON 1966). Cara makan dengan jalan menelan lumpur, jelas tidak bersifat selektif, baik terhadap ukuran granula ataupun terhadap kandungan zat organik.
Kemampuan selektif dari tentakel bukal disebabkan karena pada bagian ujung dari tentakel bukal terdapat komponen kecil yang disebut nodular. Pada nodular terdapat sel-sel yang bersifat kemosensitif. Dengan kehadiran sel-sel tersebut teripang dapat menjadi lebih selektif dalam memilih partikel makanan (ROBERTS 1979, ROBERTS & BRYCE 1982, dan BOULAND et al. 1982).
Baik pada kelompok teripang pemakan endapan ataupun pada kelompok teripang pemakan materi tersuspensi, terlihat bahwa tentakel bukal berperan aktif untuk pengumpulan makanan. Selain sel-sel kemosensitif, pada bagian ujung tentakel bukal juga terdapat sel-sel penghasil mukosa. Dengan bantuan mukus ini kerja tentakel bukal menjadi lebih efisien (PAWSON 1966, FANKBONER 1978, 1981, dan BOULAND et al. 1982).
Kelompok teripang pemakan plankton dan materi tersuspensi memanfaatkan tentakel bukalnya yang relatif lebih panjang dan mempunyai percabangan seperti pohon. Tentakel bukal ini secara aktif mengumpulkan plankton dan seston langsung dari medium air laut di sekitarnya. Beberapa anggota teripang bangsa Dendrochirotida beradaptasi untuk hidup sebagai megaloplankton.
Teripang suku Synaptidae dengan tentakel bukal berbentuk sikat (pinnate) juga tergolong sebagai teripang pemakan endapan. Tetapi berbeda dengan anggota kelompok Aspidochirotida lainnya, partikel makanan tidak langsung diambil dari substrat di sekitarnya, tetapi diperoleh dari sedimen yang menempel di daun algae, lamun ataupun pada koloni sessil bentik (spons, karang lunak, tunikata dan seterusnya). Hal ini telah dilaporkan oleh BERRILL (1966), yang meneliti teripang jenis Opheidesoma spectabilis yang hidup di perairan sekitar Hawaii. Lebih lanjut pakar ini melaporkan bahwa, dalam batasan tertentu teripang suku Synaptidae juga mempunyai kemampuan untuk menangkap partikel melayang langsung dari medium air laut di sekitarnya. Selanjutnya cara makan berbagai jenis teripang dapat dilihat pada Tabel 1.
SISTEM ENZIM
Dari penelitian mengenai ekstrak dinding usus dan cairan saluran pencernaan, diketahui adanya aktifitas enzimatik pada teripang. Enzim bersama mukosa diduga dihasilkan oleh lapisan epitelium. Mukus dan enzim secara bersama menguraikan senyawa-senyawa kimia dari partikel makanan di sepanjang saluran pencernaan (FILINOVA & TOKIN 1980). Selanjutnya FERAL (1989), meneliti teripang jenis, Leptosynapta gallinnei, Leptosynapta inhaerens, dan Holothuria forskali melaporkan adanya 25 macam enzim pada teripang tersebut. Lima macam enzim mampu menguraikan rantai senyawa ester, 12 macam enzim mampu menguraikan rantai glikosida, dan 8 macam enzim lainnya mampu menguraikan rantai senyawa peptida. Berbeda dengan pendapat LAWRENCE (1982), selanjutnya FERAL (1989), juga mendapatkan adanya enzim yang mampu memecah ikatan cellulosa dan chitin. Sistem enzim pada teripang juga mampu mengurai senyawa
kompleks, seperti protein dan gliko-lipida. Keduapuluh lima macam enzim tersebut dapat dikelompokkan kedalam Disacchari-dase, PolysacchariDisacchari-dase, amylase, maltase, pro-tease, esterease, dan chitinase. Aktifitas enzimatik dapat berlangsung sepanjang saluran pencernaan, tetapi aktifitas yang paling menonjol adalah pada usus depan (anterior intestine).
Selanjutnya FERAL (1989), melaporkan bahwa baik pada teripang kelompok pemakan endapan ataupun pemakan mated tersuspensi, diduga mempunyai komposisi enzim yang sama. Hal ini berkaitan dengan samanya sumber makanan, yaitu de-tritus dan biota kecil, perbedaan hanyalah pada cara memperolehnya (substrat pasir dan medium air laut). Dari sudut pandang enzimatik, teripang bisa dianggap sebagai biota omnivora, karena sumber makanannya bisa berasal dari sisa tanaman ataupun sisa he wan.
LAW RENCE (1982), dalam ikhtisarnya mengenai pencernaan fauna ekhinodermata, sebelumnya telah melaporkan adanya enzim amylase, cellulase, pectmase, maltase, protease, dan peptidase dengan kemampuan reaksi enzimatik sedang sampai kuat. Juga dilaporkan adanya enzim-enzim lainnya dengan reaksi enzimatik yang relatif lemah atau tidak nyata.
Tingkat asam-basa atau pH level dalam cairan lambung selama proses reaksi enzimatik bervariasi antara 5,0 sampai dengan 7,9 tetapi biasanya suasana pH adalah sedikit asam pada kebanyak teripang (LAWRENCE 1982).
EFISIENSI ASIMILASI DAN PERIODA ASIMILASI
Asimilasi adalah proses penguraian dan penyerapan material makanan, yang kemudian akan dirubah menjadi molekul kompleks yang berlangsung dalam tubuh suatu organisme. Sedangkan efisiensi asimilasi
adalah persentase material makanan yang dapat dimanfaatkan oleh suatu organisme dalam proses asimilasi tersebut.
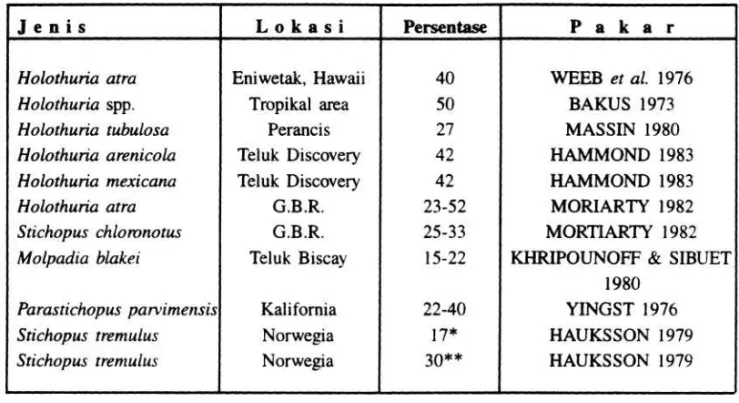
BAKUS (1973), melaporkan bahwa efisiensi asimilasi teripang yang hidup di daerah tropis adalah sekitar 50 %. Efisiensi asimilasi ini bervariasi untuk berbagai jenis teripang, dimana letak geografis dan macam habitat juga ikut mempengaruhi. Selain jenis dan tempat, menurut HAUKSSON (1979), kondisi periodisitas dari organ reproduksi juga ikut berpengaruh. Pada waktu musim memijah efisiensi asimilasi akan menurun dari waktu-waktu lainnya. Lebih lanjut MASSIN (1980), melaporkan bahwa efisiensi asimilasi pada kelompok teripang adalah lebih rendah bila dibandingkan dengan cacing laut ataupun sipunkulida. Efisiensi asimilasi berbagai jenis teripang selanjutnya dapat dilihat pada Tabel 2.
Perioda asimilasi adalah waktu yang dibutuhkan oleh sedimen untuk melewati saluran pencernaan mulai saat ditelan sampai dikeluarkan lewat anus. Waktu asimilasi ini juga sangat bervariasi menurut jenis teripang.
Faktor yang menentukan waktu asimilasi adalah ukuran panjang tubuh teripang dan panjang saluran pencernaan dari teripang tersebut (BAKUS 1973, MASSIN & JANGOUX 1976). Salah satu metode yang sederhana untuk menentukan waktu asimilasi adalah dengan jalan menggunakan zat pewarna. Dalam hal ini digunakan zat pewarna yang tidak beracun dan tidak mempunyai dampak yang merugikan terhadap kelangsungan hidup teripang tersebut (MASSIN & JANGOUX 1976, HAMMOND 1982). Pengujian waktu asimilasi ini dapat dilakukan di laboratorium ataupun di lapangan.
Hasil penelitian terhadap perioda asimilasi ini masih jauh dari memuaskan, mengingat hasilnya masih menunjukkan perbedaan waktu yang sangat bervariasi. Waktu asimilasi berbagai jenis teripang lebih
lanjut dapat dilihat pada Tabel 3.
Kebanyakan teripang aktif makan di malam hari terutama dari kelompok Apodida (BIRKELAND 1989). YAMANOUCHI (1956), mendapatkan berbagai jenis teripang kelompok Aspidochirotida bervariasi dalam aktifitas makan ini. Kelompok pertama adalah teripang yang aktif mencari makan siang dan malam, seperti teripang jenis Holothuria atra,
Holothuria edulis, dan Holothuria
flavomaculata. Sedangkan kelompok lainnya adalah teripang yang terutama aktif mencari makan di malam hari, seperti teripang jenis
Holothuria scabra, Holothuria vitiens, Actinopyga lecanora, Stichopus variegatus,
dan Stichopus chloronotus. Teripang kelompok kedua ini pada siang hari bersembunyi pada tempat yang terlindung.
PENGARUH AKTIFITAS MAKAN TERHADAP LINGKUNGAN
Berbeda dengan bintang laut dan kelompok bulu babi, pengaruh aktifitas makan pada teripang tidaklah terlalu mempengaruhi lingkungan tempat hidupnya (BIRKELAND 1989). Salah satu pengaruhnya adalah memindahkan volum tertentu dari sedimen di sekitarnya (BAKUS 1973, BIRKELAND 1989).
MASSIN (1982), menyimpulkan adanya dua macam pengaruh dari aktifitas makan teripang terhadap lingkungannya. Pertama adalah merubah komposisi ukuran partikel pasir (sedimen) melalui proses pencernaan. Hal ini terutama dilakukan oleh kelompok teripang pemakan endapan. Efek kedua adalah pengadukan partikel sedimen (bioturbation). Tetapi dari hasil penelitian diketahui bahwa, baik daya "giling" ataupun reaksi kimiawi pada teripang adalah sangat lemah sehingga tidak begitu jelas dampaknya dalam merubah (memperkecil) ukuran partikel pasir yang ditelannya.
Pemindahan partikel pasir, terbatas kepada perubahan batasan tertentu dari stratifikasi sedimen dan mempengaruhi stabilitas dari sedimen. Aksi bioturbasi ini pada kelompok teripang tidaklah begitu menonjol. Dengan tetap tingginya kandungan
zat organik dalam fecal pellet teripang, secara biologis teripang bisa dipandang menyediakan semacam fasilitas untuk kelompok biota pemakan endapan lainnya. Semoga tulisan yang ringkas ini bisa menambah pengetahuan kita terhadap kelompok teripang ini.
Tabel 1. Cara makan berbagai jenis teripang (Holothuroidea)
Catatan : Terlihat dari tabel ini hampir semua teripang komersial (Aspidochirotida) adalah termasuk kedalam kelompok teripang pemakan endapan
Tabel 2. Effisiensi asimilasi berbagai jenis teripang (Holothuroidea)
Keterangan : GBR = Karang Penghalang Besar, Australia * = Saat musim memijah
** = Di luar musim memijah
Tabel 3. Waktu asimilasi berbagai jenis teripang (Holothuroidea)
J e n i s L o k a s i Perioda (jam) P a k a r Holothuria atra Holothuria difficilis Holothuria tubulosa Stichopus japonicus Eniwetak, Hawaii 10-36 Eniwetak, Hawaii 15 Mediterania 2,5-24 Hokkaido 30 BAKUS 1973 BAKUS 1968 MASSIN & JANGOUX 1976
DAFTAR PUSTAKA
BAKUS, G.J. 1973. The Biology and the Ecology of tropical holothurians. In : JONES, O.A. and R. ENDEAN (eds.), Biology and Geology of Coral Reefs. 2. Biologi. 1. Academic Press, New York : 325-367.
BERRILL, M. 1966. Synaptid holothurian, Opheidesoma spectabilis. Can. J. Zool. 44 (3) : 457-482.
BIRKELAND, C. 1989. The influence of echinoderms on coral reef communi-ties. In: JANGOUX, M. and J. M. LAWRENCE (eds.), Echinoderms studies 3. Balkema, Rotterdam : 79 pp. BONHAM, K. and E.E. HELD 1963. Eco-logical observations on the sea cucum-ber Holothuria atra and Holothuria leucospilota at Rongelap Atoll, Marshall Islands. Pac. Sci. 17 (1) : 305-314.
BOULAND, C, C. MASSIN, and M. JANGOUX 1982. The fine structure of the bucal tentacles of Holothuria forskali (Echinodermata, Holothuroidea) Zoomorphology 101 : 133-149.
CAMERON, J.L. and P.V. FANKBONER 1984. Tentacle structure and feeding processes in life stages of the commer-cial sea cucumber Parastichopus californicus (Stimpson). J. Exp. Mar. Biol. Ecol 81 : 193-209
CLARK, A.M. and F.W.E. ROWE 1971. Monograph of shallow water Indo-west Pacific echinoderms. Trustees of the British Museum (Nat. Hist.) Lon-don : pp.
COULON, P.and M. JANGOUX 1993. Feed-ing rate and sediment reworkFeed-ing by the holothuroid Holothuria tubulosa (Echinodermata) in a Mediterranean seagraas bed off Ischia Island, Italy. Mar. Ecol. Progr. Ser. 92 : 201-204. FANKBONER, P.V. 1978.
Suspension-feed-ing mechanisms of the armoured sea cucumber Psolus chitinoides Clark. J. Exp. Mar. Biol Ecol 31 : 11-25. FANKBONER, P.V. 1981. A reexamination
of mocus feeding by the sea cucumber Leptopentacta elongata. J. Mar. Biol Ass. U.K. 61 : 679-683.
FERAL, J.P. 1989. Activity of the principal digestive enzymes in the detritivorous apodous holothuroid Leptosynapta galliennei and two other shallow-wa-ter holothuroids. Mar. Biol. 101 : 367-379.
FERAL, J.P. and C. MASSIN 1982. Diges-tive systems : Holothuroidea. In : JANGOUX, M. and J.M. LAWRENCE (eds.), Echinoderm Nutrition. Balkema, Rotterdam : 191-212.
FILIMONOVA, G.F. and I.B. TOKIN 1980. (Structural and functional peculiarities of the Digestive system of Cucumaria frondosa (Echinodermata : Holothhuroidea). Mar. Biol. 60 : 9-16. FRIZZELL, D.L., EXLINE, and D.L.
PAWSON 1966. Phylogeny and evo-lution Holothurians. In : Tratise on invertebrate paleontology. 3. Univ. Kansas Press, New York : U641-U645.
HAMMOND, L.S. 1981. An analysis of grain size modification in biogenic carbonate sediments by deposit-
feeding holothurians and echinoids (Echinodermata). Limnol. Oceanogr. 26 (5) : 898-906.
HAMMOND, L.S. 1982. Patterns of feeding and activity in deposit feeding ho lothurians and echinoids (Echinodermata) from a shallow back- reef lagoon, Discovery Bay, Jamaica. Bull. Mar. Sci. 31(2) : 549-574. HAMMOND, L.S. 1983. Nutrition of
de-posit-feeding holothuroids and echi-noids (Echinodermata) from a shallow reef lagoon, Discovery Bay, Jamaica. Mar.Ecol. Progr. Ser. 64 (3) : 155-160.
HAUKSON, E. 1979. Feeding biology of Stichopus tremulus, a deposit-feeding holothurian. Sarsia 64 (3) : 155-160. KLINGER, T.S., C.R. JOHNSON, and J.JELL
1994. Sediment utilization, feeding niche breadth, and feeding-niche over-lap of Aspidochirotida (Echinodermata : Holothuroidea) at Heron Island, Great Barrier Reef. In : DAVID, GUILLE, FERAL and ROUX (eds.), Echino-derms through time. Balkema, Rotterdam : 523-528.
KHRIPOUNOFF, A. and M. SIBUET 1980. La nutrition d'echinodermes abyssaux I. Alimentation des holothuries. Mar. Biol. 60 : 17-26.
HYMAN, L. 1955. The Invertebrates. IV. Echinodermata the coelomata bilateria. Me Graw-Hill Co., New York : 156-162
LAMBERSON, J.O. 1978. Notes on the Morphology, ecoogy and distribution of Thelonota anax (Holothuroidea : Stichopodidae.) Micronesica 14 (1) : 115-122.
LAWRENCE, J.M. 1982. Digestion of Echinodermata. In : JANGOUX, M. and J.M. LAWRENCE (eds.), Echino-derm Nutrition. Balkema, Rotterdam : 283-316.
MASSIN, C. 1980. The sediment ingested by Holothuria tubulosa (Holothuroidea : Echinodermata). In : JANGOUX, M. (ed.), Echinoderms : Present and Past. Balkema, Rotterdam : 206-208 MASSIN, C. 1982. Food and feeding
mechanisme : Holothuroidea; Effects of feeding on the environment : Holothuroidea. In : JANGOUX, M. and J.M. LAWRENCE (eds.), Echino-derm Nutrition. Balkema, Rotterdam : 43-55, 493-497.
MASSIN, C. and C. DOUMEN 1986. Distri-bution and feeding of epibenthic holothuroids on the reef flat of Laing Island (Papua New Guinea). Mar. Ecol.Progr. Ser. 31 : 185-195. MASSIN, C. et JANGOUX 1976.
Observa-tion ecologiques sur Holothuria tubulosa, H. poll et H. forskali (Echinodermata-Holothuroidea) et comportement alimentaire de H.
tubulosa. Cah. Biol. Mar. 17 : 45-59. MORIARTY, D.J.W. 1982. Feeding of
Holothuria atra and Stichopus
chloronotus on bacteria, organic car-bon and organic nitrogen in sediment of the Great Barrier Reef. Austr. J. Mar. Freshw. Res. 33 : 255-263. PAWSON, D.L. 1966. Ecology of
Holothuri-ans. In : BOOLOTIAN, R.A. (ed.), Physiology of Echinodermata. Intersci, Publ. New York : 63-71.
ROBERTS, D. 1979. Deposit-feeding mecha-nisms and resource partitioning in
tropical Holothurians. J. Exp. Mar. Biol. Ecol. 37 : 43-56.
ROBERTS, D. 1982. Classification and ho-lothurian tentacle. In : LAWRENCE. J.M. (ed.), Echinoderms : Proceedings of the International Conference, Tampa Bay. Balkema, Rotterdam : 1 1 7 - 1 2 1 .
ROBERTS, D. and C. BRYCE 1982. Further observations on tentacular feeding mechanisms in Holothurians. J. Exp. Mar. Biol. Ecol. 59 : 151-163.
TANAKA, Y. 1958. Feeding and digestive processes of Stichopus japonicus. Bull. Fac. Fish. Hokkaido Univ. 9 ( 1 ) : 14-25.
WEBB, K.L., W.D. DUPAUL, and C.F. D'ELIA 1977. Biomass and nutrient flux measurements on Holothuria atra
populations on windward reef flats at Eniwetak, Marshall Islands. Proc. Third International Coral Reef Symposium : 409-415.
YAMANOUCHI, T. 1956. The daily activity rhythms of the holothurians in the coral reef of Palao Islands. Publ. Seto Mar. Biol. Lab. 5 : 347-362.
YINGST, J.Y. 1976. The utilization of organic matter in shallow marins sediments by an
epibenthic deposit-feeding Holothurian. J. Exp. Mar. Biol Ecol.
23 : 55-69.
YINGST, J.Y. 1982. Factors influencing rates of sediment ingestion by Parastichopus parvimensis, an epibenthic deposit-feeding Holothurian. Estuarine, Coastal, and Shelf Science 14 : 1 1 9 -134.