KETAHANAN TANAMAN KOPI (
COFFEA SPP.)
TERHADAP HAMA PENGGEREK BUAH KOPI
(HYPOTHENEMUS HAMPEI
FERR.)
Coffee Resistance to Coffee Berry Borer
(Hypothenemus hampei Ferr.)
Agung Wahyu Susilo Peneliti Pemuliaan Tanaman Pusat Penelitian Kopi dan Kakao Indonesia
Jl. PB. Sudirman No. 90 Jember
Ringkasan
Salah satu permasalahan pada pertanaman kopi (Coffea spp.) adalah serangan hama penggerek buah kopi (Hypothenemus hampei Ferr., PBKo). Informasi mengenai ketahanan tanaman kopi terhadap hama PBKo diperlukan untuk strategi pengendalian dan perakitan varietas tahan. Tulisan ini mengulas ketahanan tanaman kopi terhadap hama PBKo bersumber dari berbagai informasi hasil-hasil penelitian yang menyangkut aspek fenologi tanaman kopi, biologi kumbang H. hampei dan interaksi antara inang tanaman kopi dengan kumbang H. Hampei. Hama PBKo menyebabkan kerusakan jaringan endosperma biji sehingga terjadi penurunan kualitas biji. Permasalahan hama PBKo lebih serius dijumpai pada spesies kopi Robusta (Coffea canephora) dibandingkan pada spesies kopi Arabika (Coffea arabica) terkait dengan perbedaan tipe pembungaan dan kesesuaian lingkungan tumbuh. Mekanisme ketahanan antixenosis dipengaruhi oleh perbedaan fenologi buah seperti ukuran, bentuk diskus, warna, dan aroma. Tingkat kekerasan kulit tanduk (parchment) diduga yang berperan dalam mekanisme antibiosis, sedangkan keserempakan waktu pemasakan buah dan ketinggian tempat dapat berpengaruh terhadap ekspresi ketahanan semu. Telah diinformasikan penemuan beberapa klon harapan tahan hasil seleksi yang dapat dimanfaatkan untuk perakitan varietas tahan dan studi mekanisme ketahanan PBKo.
Summary
Coffee berry borer, Hypothenemus hampei Ferr. (CBB) is the main pest on coffee. Information concerning the resistance would be so important for breeding and pest control. This paper will review the resistant mechanism which has been reported in several papers related with plant phenology, biology of H. hampei and host-pest interaction. CBB can affect yield losses due to endosperm was infected by the pest. The pest incidence was found higher in Robusta coffee (Coffea canephora) than in Arabica coffee (Coffea arabica) related to the differences on flowering type and agroclimatic adaptability. Antixenosis mechanism was influenced by berry
phenology such as size, discus, color and flavour. Parchment hardness was identi-fied as the antibiosis factor on suppressing larva movement and the ecological resistance was performed by the plants in which relatively bearing flower in the same time and higher altitude lowering infestation. It has been selected some promising resistant clones for the purpose of breeding and study on resistant mechanism.
Key words: Coffee berry borer, Coffea spp., resistance.
LATAR BELAKANG
Kopi (Coffea spp.) merupakan salah satu komoditas ekspor andalan penyumbang devisa negara senilai USD 521,3 juta per tahun. Nilai tersebut memberi kontribusi terhadap perolehan total devisa sebesar 0,57% atau 24,18% dari perolehan devisa sub sektor pertanian. Produksi kopi Indone-sia mencapai 674.800 ton (tahun 2005) yang diperoleh dari areal seluas 1.302.042 ha dengan komposisi 91,05% jenis kopi Robusta dan 8,95% jenis kopi Arabika. Luasan areal pertanaman kopi tersebut merupakan yang terluas kedua di dunia setelah Brazil, namun berdasarkan produksi dan volume ekspor, Indonesia menempati urutan ke empat setelah Brazil, Vietnam, dan Columbia. Selain sebagai sumber devisa kopi juga berperan penting dalam mendorong agri-bisnis, sebagai sumber pendapatan, dan penciptaan lapangan pekerjaan yang berbasis masyarakat pedesaan. Hal ini terlihat dari komposisi luas areal pertanaman kopi per-kebunan rakyat yang mencapai 1.249.554 ha (95,96%) dibandingkan areal pertanaman perkebunan besar yang hanya 26.597 ha (2,04%). Secara keseluruhan ada sekitar 2 juta tenaga kerja yang terlibat dalam pengusahaan kopi dengan jumlah jiwa yang ditanggung mencapai 5 juta orang (Ditjenbun, 2006).
Komoditas kopi yang dikomersialkan umumnya adalah jenis kopi Arabika dan kopi Robusta. Di pasaran dunia, kopi Arabika dibedakan atas 3 kelompok berdasarkan kualitas citarasanya, yaitu kopi Arabika biasa, kopi spesialti, dan kopi organik. Kopi spesialti merupakan produk yang memiliki kualitas citarasa tertinggi karena kekhasannya. Indonesia memiliki peluang akan produk-produk kopi spesialti, seperti Mandailing Coffee dari Sumatera Utara, Gayo Mountain Coffee dari Aceh Tengah, Java Coffee dari Jawa Timur, Toraja Coffee dari Sulawesi Selatan (Ditjenbun, 2006). Saat ini tuntutan konsumen akan produk organik telah membuka peluang pasar bagi produk-produk kopi yang dikelola secara organik dengan tetap mempertimbangkan aspek kekhasan citarasa tersebut. Meskipun secara ekonomi pengusahaan kopi Arabika lebih menguntungkan namun komposisi produksi kopi Arabika relatif rendah disebabkan ar-eal pengembangan kopi Arabika terbatas pada daerah-daerah dataran tinggi. Kopi merupakan produk pertanian yang mengandalkan aspek kualitas citarasa sehingga sasaran akhir budidaya kopi adalah produk biji yang berkualitas tinggi.
Di antara permasalahan dalam budi-daya kopi adalah serangan hama penggerek
buah kopi, Hypothenemus hampei Ferr. (PBKo). Hama PBKo ini selain menyerang biji kopi di pertanaman juga dapat me-nyerang biji kopi sewaktu di penyimpanan. Serangan hama PBKo menyebabkan penurunan produktivitas dan kualitas hasil secara nyata. Serangan pada stadia buah muda dapat menyebabkan keguguran buah sebelum buah masak, sedangkan serangan pada stadia buah masak (tua) menyebabkan biji berlubang sehingga terjadi penurunan berat dan kualitas biji (Sulistyowati, 1992). Kehilangan hasil akibat serangan PBKo bervariasi tergantung kondisi pengelolaan tanaman. Pada pertanaman yang tidak dilakukan tindakan pengendalian serang-an hama PBKo dapat mencapai 100% (Baker, 1999 cit. Prakasan et al., 2001). Kisaran kehilangan hasil akibat PBKo yang dilaporkan mencapai 5—80% (Idowu cit. Prakasan et al., 2001), 30—80% (Sulistyowati, 1992). Situasi harga biji kopi secara tidak langsung juga dapat berpengaruh terhadap intensitas serangan hama PBKo di pertanaman. Pada saat harga biji kopi baik, maka semua biji-biji kopi dapat terpanen secara maksimal sehingga akan memutus siklus hidup H. hampei yang dapat menurun-kan intensitas serangan PBKo pada periode panen berikutnya (Damon, 2000). Durham (2004) menyebut-kan total kerugian akibat serangan hama PBKo di dunia mencapai USD 500 juta. Kasus serangan hama PBKo lebih banyak dijumpai pada pertanaman kopi Robusta dibandingkan pada pertanaman kopi Arabika disebabkan fenologi pembuahan kopi Robusta yang terus menerus dan kondisi lingkungan tumbuh kopi Robusta
yang lebih sesuai untuk mendukung perkembangan H. hampei (Damon, 2000). Strategi pengendalian hama PBKo di-arahkan melalui sistem pengendalian hama terpadu (PHT) yang memadukan antara komponen bahan tanam tahan, agens hayati, dan manajemen lingkungan. Sulistyowati (1992) telah menjabarkan beberapa kom-ponen pengendalian PBKo secara terpadu. Secara kultur teknis pengendalian hama PBKo dapat dilakukan untuk memutus daur hidup kumbang PBKo dengan cara melaku-kan petik bubuk (petik buah merah 15—30 hari menjelang panen besar), lelesan (tindak-an pemungut(tindak-an buah-buah y(tindak-ang terjatuh di tanah), dan racutan (pemetikan buah-buah yang masih tersisa pada akhir masa panen). Pengelolaan tanaman penaung yang tepat, yaitu menjaga kondisi penaungan tidak terlalu gelap juga dapat menekan per-kembangan kumbang H. hampei. Pengen-dalian secara biologis dapat menggunakan parasitoid Cephalonomia stepanoderis Betr., Prorops nasuta, dan Heterospilus coffeicola Schm. Jamur entomopatogen Beuveria bassiana juga telah direkomendasikan sebagai agens hayati hama PBKo. Namun demikian hingga kini belum tersedia bahan tanam tahan anjuran untuk pengendalian hama PBKo. Anjuran penanaman adalah penggunaan varietas/klon yang memiliki sifat pembungaan serempak.
Dalam aplikasi teknologi pengendalian diperlukan pemahaman yang baik mengenai mekanisme hubungan antara tanaman inang dengan serangga hama. Ketahanan tanaman merupakan salah satu bentuk ekspresi hubungan antara inang dengan serangga
hama yang proses interaksinya dipengaruhi oleh faktor lingkungan tumbuh. Terkait dengan masalah hama PBKo tersebut, tulisan ini akan menguraikan aspek-aspek ketahanan tanaman terhadap serangan hama PBKo sebagai informasi dasar yang di-perlukan dalam tindakan pengendalian dan perakitan bahan tanam tahan.
FENOLOGI TANAMAN KOPI
(COFFEA SPP.)
Spesies kopi yang dikomersialkan umumnya adalah Coffea arabica (kopi arabika) dan Coffea canephora (kopi Robusta) dari kelompok subseksi Erythro-coffea marga Coffea. Secara sitologi kedua spesies kopi tersebut berbeda, yaitu kopi arabika termasuk tanaman allotetraploid (2n= 4x=44) dan kopi Robusta termasuk tanaman diploid (2n = 22). Berdasarkan sifat penyerbukannya, kopi Arabika bersifat menyerbuk sendiri (self compatible) dan tanaman kopi Robusta bersifat menyerbuk silang (cross compatible) (Charrier & Berthaud, 1985). Adanya perbedaan sifat-sifat ini membawa konsekuensi secara mendasar dalam strategi pemuliaan dan metode perbanyakan kedua spesies tersebut. Habitat tumbuh asli tanaman kopi berada di kawasan hutan hujan tropis di wilayah Afrika (Charrier & Berthaud, 1985). Dalam pengembangannya budidaya kopi memerlukan tanaman penaung sebagai pelindung terhadap pencahayaan matahari langsung guna mengurangi proses evapo-transpirasi. Curah hujan dan suhu merupakan faktor iklim yang berpengaruh terhadap produksi kopi. Dalam hal ini volume dan
distribusi curah hujan sepanjang tahun, dan ketinggian tempat menentukan kesesuaian tumbuh tanaman kopi (Rothfos, 1980). Curah hujan tahunan yang diperlukan untuk pertumbuhan kopi Arabika berkisar antara 1.100—1.300 mm dan untuk kopi Robusta 1.550—2.000 mm. Distribusi curah hujan sangat berpengaruh dalam menentukan pola pembuahan tanaman karena terkait dengan proses pematahan dormansi bunga. Sedangkan ketinggian tempat yang sesuai untuk pertumbuhan kopi Arabika adalah 1000—2000 m dpl. yang kondisi suhunya berkisar antara 15 OC hingga 24 OC, dan
untuk tanaman kopi Robusta <700 m dpl. yang kondisi suhunya berkisar antara 24 OC—30 OC (Wilson, 1985).
Pembentukan buah kopi terpengaruh oleh kondisi musim yang ekstrim, dimana kondisi musim kering berkepanjangan atau pun hujan yang berlebihan akan menganggu proses pembungaan dan pembuahan. Kondisi curah hujan tinggi akan menganggu proses penyerbukan bunga kopi yang dibantu oleh angin dan kumbang penyerbuk. Sebelum mekar, bunga kopi secara fisiologi akan mengalami masa dormansi. Pada saat itu kuncup bunga terhenti perkembangannya selama beberapa bulan (1—4 bulan). Faktor pemacu perkembangan bunga setelah masa dormansi tersebut adalah ketersediaan air tanah. Meskipun masa dormansi telah terlewati namun apabila air tanah belum mencukupi kebutuhan proses pemekaran bunga maka bunga belum akan mekar. Proses pemekaran bunga biasanya dipicu oleh tibanya musim hujan atau melalui proses penyiraman (Cannell, 1985). Karena itu pada daerah-daerah yang bertipe iklim
basah dengan sebaran curah hujan merata sepanjang tahun pembuahan kopi dapat berlangsung sepanjang tahun.
Buah kopi memiliki dua biji yang posisinya berhadapan satu sama lain disatu-kan oleh kulit yang berwarna merah ketika masak, mengandung pulp yang rasanya manis. Setiap biji tersebut endospermanya diselubungi oleh kulit tanduk (parchment) yang keras (Rothfos, 1980). Ukuran biji tersebut juga dipengaruhi oleh kondisi curah hujan saat pembentukan biji. Pada daerah-daerah yang memiliki tipe curah hujan tinggi ukuran bijinya lebih besar dibanding daerah-daerah kering.
HAMA PENGGEREK BUAH KOPI (HYPOTHENEMUS
HAMPEI)
Hama penggerek buah kopi (PBKo), Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae) pertama kali ditemukan di Uganda tahun 1867, kemudian hama ini menyebar ke berbagai areal pertanaman kopi di dunia. Negara produsen kopi yang di-laporkan pertanaman kopinya belum ter-serang hama PBKo adalah Costa Rica, Papua New Guinea, dan Hawai (Giorda-nengo et al., 1993). Di Indonesia, hama PBKo pertama kali ditemukan tahun 1909 di perkebunan Lampegan, Jawa Barat yaitu menyerang tanaman kopi jenis Liberika (Cramer, 1957), dan penyebarannya diduga melalui pemasukan kopi dari Uganda (Sulistyowati, 1992).
Prakasan et al. (2001) menguraikan proses kumbang H. hampei menggerek buah kopi. Kumbang H. hampei dapat menyerang
buah stadia muda hingga stadia tua. Kumbang H. hampei menggerek di bagian diskus buah, kemudian masuk ke dalam jaringan endosperma yang menyebabkan kerusakan biji. Umumnya dalam satu buah ditemukan ada satu kumbang betina H. hampei, namun kadang-kadang dapat ditemukan lebih dari satu kumbang. Meskipun serangan ditemukan pada buah muda namun H. hampei lebih menyukai buah-buah yang endospermanya sudah mengeras. Biasanya apabila kumbang H. hampei menyerang buah muda yang endospermanya belum mengeras, kumbang akan tetap tinggal di lubang gerekan menanti sampai endosperma biji mengeras atau keluar mencari alternatif buah yang endosperma bijinya sudah mengeras. Buah muda yang terserang H. hampei akan terganggu perkembangannya, biasanya buah menjadi busuk lalu gugur sebelum masak.
Biologi H. Hampei
Hama PBKo (H. hampei) berbentuk kumbang kecil berwarna hitam, ukuran kumbang betina panjang 1,5—2,5 mm (Venkatesha et al., 1998, Prakasan et al., 2001) yang lebih panjang dibandingkan kumbang jantan 1,0 mm (Prakasan et al., 2001). H. hampei meletakkan telur di dalam buah antara 20—60 butir, dan telur akan menetas setelah 4—9 hari (Prakasan et al., 2001). Periode larva berlangsung selama 10—26 hari, periode pra pupa selama 2 hari dan pupa selama 4 hari. Satu siklus hidup H. hampei berlangsung selama 25—35 hari (Le Pelley, 1968 cit. Prakasan et al., 2001). Kumbang-kumbang yang baru terbentuk di
dalam buah kemudian akan melakukan perkawinan sesamanya (sibling). Rasio antara kumbang betina dan kumbang jantan sekitar 10:1. Setelah proses kawin kumbang betina akan meninggalkan buah untuk mencari buah-buah yang masih segar untuk proses infestasi berikutnya, dan kumbang jantan tetap tinggal di dalam buah. Aktivitas infestasi tersebut dilakukan pada waktu sore hari. Kumbang H. hampei mampu bertahan lebih dari 5 bulan di dalam buah, baik pada buah yang masih melekat di pohon maupun buah yang sudah gugur (Venkatesha et al., 1998).
Kumbang betina H. hampei daya jangkaunya dapat mencapai jarak sejauh 345 m, menggunakan sayap dan dapat juga dibantu oleh angin. Kemampuan hidup kumbang betina rata-rata selama 156 hari sedangkan kemampuan hidup kumbang jantan lebih pendek, yaitu selama 103 hari (Bergamin, 1943 cit. Prakasan et al., 2001). Daya hidup yang lama bagi kumbang H. hampei tersebut memungkinkan infestasi dapat dilakukan pada periode panen berikutnya. Buah-buah yang tidak terpanen atau pun buah yang gugur merupakan media tumbuh bagi H. hampei selama
Kumbang H. hampei
(Foto : Venkatesha et al., 1998)
Lubang gerekan H. hampei
(Foto : Venkatesha et al., 1998)
Larva H. hampei
(Foto : Venkatesha et al., 1998) Sumber foto : ICCRI
Gambar 1. Fenologi buah kopi dan gejala serangan hama PBKo (foto dari berbagai sumber; ICCRI, Venkatesha et al., 1998).
Figure 1. Phenology of coffee berry and visual symptoms affected by coffee berry borer (Photo by ICCRI and Venkatesha et al., 1998).
menunggu musim panen berikutnya (Prakasan et al., 2001).
Lingkungan tumbuh H. Hampei
Suhu optimum untuk perkembangan PBKo berkisar antara 25—26 OC (Barrera
& Baker, 1985 cit. Prakasan et al., 2001), sedangkan kelembaban optimum yang dibutuhkan berkisar antara 90—95% (Baker et al., 1994 cit. Prakasan et al., 2001). Selain faktor suhu dan kelembaban, ketinggian tempat (elevasi) juga ber-pengaruh terhadap perkembangan PBKo. Serangan hama PBKo dijumpai lebih tinggi pada kisaran ketinggian tempat 500—1000 m dpl. daripada pada ketinggian tempat <500 m dpl. atau >1000 m dpl. (Venkatesha et al., 1998; Prakasan et al., 2001). Pertanaman kopi dengan penaungan yang berlebihan (gelap) mendukung perkembangan PBKo lebih baik di-bandingkan pertanaman yang kondisi penaungannya kurang (terbuka) (Le Pelley, 1968 cit. Prakasan et al., 2001). Pada per-tanaman dengan penaungan rapat, dilaporkan bahwa buah yang terinfestasi PBKo 5 kali lebih banyak dan per-kembangan H. hampei lebih cepat dibandingkan pada pertanaman dengan kondisi penaungan kurang (terbuka). Demikian juga pertanaman yang pem-buahannya sepanjang tahun akan men-dukung keberlanjutan pembiakan PBKo. Tanaman inang H. Hampei
H. hampei termasuk kumbang monofag (manophagous insect) yang hanya mampu bertahan hidup pada biji-bijian tanaman kopi
(Coffea spp.) (Swarupa et al., 2000, Prakasan et al., 2001). Namun demikian dijumpai ada sebagian tahapan daur hidup kumbang H. hampei yang ditemukan pada tanaman Caesalpinia, Centrosemo, Crota-laria (Swarupa et al., 2000), keberhasilan pembiakan H. hampei pada media biji-bijian Melicocca bijuga dan Cajanus cajan (Baker, 1999 cit. Prakasan et al., 2001).
Musuh alami H. Hampei
H. hampei memiliki musuh alami dari kelompok predator; Dindymus rubiginous, kelompok parasitoid; Cephalonomia stepha-noderis, Prorops nasuta (Hymenoptera: Bethylidae), Phymastichus coffea (Hy-menoptera: Eulophidae), dan Heterospilus coffeicola (Hymenoptera: Braconidae), kelompok nematoda; Heterorhabditis sp., Penagrolaimus sp., kelompok bakteri; Bacillus thuringiensis, dan kelompok jamur; Beuveria bassiana, Paecilomyces fumosoroseus (Prakasan et al., 2001).
KETAHANAN TANAMAN TERHADAP HAMA PBKO Hubungan H. hampei dengan tanaman
inang
Kumbang H. hampei betina setelah dewasa akan keluar dari dalam biji kopi yang diinfestasi oleh induknya, terbang mencari biji-biji kopi baru sebagai sumber pakan dan tempat berkembang biaknya. Dalam hal ini kumbang jantan tetap tinggal di dalam biji. Kumbang tersebut keluar dari biji umumnya setelah kawin. Proporsi
kumbang betina yang telah kawin keluar dari dalam biji mencapai 62% (Lopez & Frérot, 1993 cit., Damon, 2000; Baker, 1999 cit. Prakasan et al., 2001). Proses keluarnya kumbang H. hampei dari dalam biji dipicu oleh kondisi kelembaban udara yang tinggi, berkisar antara 90—100%. Waktu efektif bagi kumbang H. hampei terbang mencari inangnya adalah sore hari antara pukul 16.00—18.00 (Cramer, 1957). Renwick & Chew (1994) menyebutkan bahwa proses penemuan inang oleh kumbang dipengaruhi oleh faktor visual. Presepsi kumbang H. hampei terhadap inangnya ditentukan berdasarkan sensor warna (vission) dan aroma (olfaction) biji kopi. Hasil uji tingkat kesukaan (preferency) bahwa kumbang H. hampei lebih menyukai biji-biji kopi yang berwarna merah (masak) dibanding biji kopi yang hijau (muda) meskipun pada buah muda juga ditemukan adanya infestasi kumbang H. hampei (Giordanengo et al., 1993). Dalam uji warna perangkap disebutkan bahwa kumbang H. hampei lebih tertarik pada perangkap berwarna merah (Mathieu et al., 1997; Wiryadiputra, 2006). Hasil uji olfaktori membuktikan adanya senyawa volatil dikeluarkan oleh biji-biji yang masak, menyebabkan kumbang H. hampei tertarik untuk mendekat (Giordanengo et al. (1993). Karena itu dapat diduga bahwa kandungan senyawa volatil tersebut terkait dengan tingkat kemasakan buah sehingga H. hampei lebih tertarik pada buah-buah masak yang memiliki kandungan volatil lebih tinggi. Hasil uji senyawa perangkap dilaporkan bahwa campuran metanol dan etanol (1:1) terbukti paling efektif menarik kumbang
H. hampei, dan dalam hal ini metanol merupakan senyawa kunci penarik kumbang tersebut (Mathieu et al., 1997; Prakasan et al., 2001). Metanol dan etanol termasuk metabolit sekunder kelompok fenolik yang diproduksi oleh lebih dari 70 keluarga tanaman dikotiledon (Panda & Khush, 1995). Karena itu diketahui bahwa proses penemuan inang oleh kumbang H. hampei diawali oleh respons positif terhadap signal warna, kemudian diikuti oleh respons terhadap senyawa volatil yang dikeluarkan oleh biji kopi.
Kumbang H. hampei terbang secara horisontal menyukai daerah di sekitar kanopi tanaman. Dalam pencarian inang tersebut terdapat perbedaan tingkat kesukaan jenis inang berdasarkan spesies dalam marga Coffea. Hasil penelitian di India pada skala lapangan dan laboratorium menunjukkan bahwa serangan oleh H. hampei lebih tinggi dijumpai pada spesies C. arabica dibanding-kan pada spesies C. canephora, dan spesies kopi lain seperti C. liberica, C. eugenoides (Prakasan et al., 2001). Sebaliknya Mathieu et al. (1997) melaporkan bahwa H. hampei lebih tertarik pada spesies C. canephora daripada spesies C. arabica. Di Indonesia disebutkan bahwa spesies C. arabica lebih peka terhadap serangan hama PBKo di-bandingkan spesies C. canephora. Meskipun masalah hama PBKo lebih serius dijumpai pada pertanaman kopi Robusta hal ini terkait dengan lingkungan tumbuh kopi Arabika yang kurang mendukung perkembangan H. hampei (Cramer, 1957) dan fenologi pembuahan kopi Robusta yang relatif berkesinambungan (Damon, 2000). Belum
ada laporan yang secara jelas menyebutkan hubungan tingkat kesukaan tersebut dengan karakteristik biji antar spesies marga Coffea tersebut. Friederichs (1924) cit. Damon (2000) menyebutkan bahwa H. hampei lebih tertarik pada buah-buah kopi yang berukuran kecil, sedangkan Ticheler (1961) cit. Damon (2000) menyebukan buah kopi yang memiliki diskus kecil kurang disukai H. hampei. Kelimpahan jumlah buah di pertanaman juga berpengaruh terhadap mobilitas kumbang H. hampei. Pada saat lepas panen, kumbang H. hampei banyak berkoloni pada buah-buah sisa panen sebelum akhirnya menemukan buah-buah segar untuk proses perkembangbiakan selanjutnya. Buah-buah lepas panen merupakan tempat yang baik bagi kumbang H. hampei untuk berkoloni sambil menunggu periode pembuahan berikutnya. Hasil penelitian di India ditemukan >20% dalam satu buah dikoloni oleh lebih dari 50 kumbang H. hampei (Prakasan et al., 2001). Setelah kumbang betina H. hampei berhasil menemukan inangnya, aktivitas yang kemudian dilakukan adalah menggerek di bagian diskus buah dengan membentuk lubang gerekan kecil (ø1 mm) sepanjang kulit buah hingga tembus di bagian endo-sperma (Sulistyowati, 1992). Kumbang H. hampei lebih menyukai biji-biji kopi yang endo-spermanya sudah menggeras (Cramer, 1857; Venkatesha et al., 1998; Prakasan et al., 2001). Hal ini diduga terkait dengan aktivitas peletakan telur sebab kumbang H. hampei juga dapat memanfaatkan buah-buah muda yang endospermanya masih lunak sebagai sumber pakannya (Cramer, 1957; Prakasan et al., 2001). Apabila
infestasi dilakukan pada buah yang endospermanya masih lunak, kumbang H. hampei tidak akan melanjutkan proses berbiak di dalam buah tersebut melainkan akan pindah mencari buah-buah lain yang endospermanya sudah menggeras (Prakasan et al., 2001). Buah muda yang telah ter-serang oleh H. hampei akhirnya akan gugur (Cramer, 1957). Hal ini jika dikaitkan dengan proses pencarian inang dapat diketahui bahwa senyawa volatil yang dikeluarkan biji-biji kopi juga terkait dengan ketertarikan kumbang untuk peletakan telur. Seluruh daur hidup kumbang H. hampei akan dilalui di dalam buah yang di-infestasinya tersebut (Durham, 2004). Faktor yang berpengaruh terhadap lama waktu daur hidup kumbang H. hampei di dalam buah adalah suhu. Semakin tinggi suhu akan semakin cepat waktu daur hidupnya dan sebaliknya pada kondisi suhu lingkungan rendah daur hidup kumbang akan semakin lama. Kondisi suhu ini juga terkait dengan faktor ketinggian tempat, dimana semakin tinggi ketinggian tempat suhu lingkungan semakin rendah maka daur hidup kumbang H. hampei juga akan semakin lama. Di dataran rendah lama waktu daur hidup kumbang H. hampei rata-rata selama 25 hari (Friederichs, 1924 cit. Cramer, 1957). Suhu optimum untuk per-kembangbiakan H. hampei adalah 25 OC
(Prakasan et al., 2001).
Mekanisme ketahanan tanaman Belum banyak informasi hasil penelitian yang menjelaskan mengenai mekanisme ketahanan hama PBKo.
Hasil-hasil penelitian umumnya baru menyebut-kan perbedaan intensitas serangan PBKo pada berbagai spesies atau klon yang menggambarkan respons ketahanan tanaman hasil infestasi alami. Fenomena ketahanan kebal (immune) pernah di-laporkan oleh Schweizer (1925) cit. Cramer (1957) pada spesies C. schumanniana. Mekanisme ketahanan kebal merupakan ekspresi non-host resistance (Chahal & Gosal, 2002) sehingga dalam hal ini dapat diketahui bahwa tidak semua spesies dalam marga Coffea dapat digunakan oleh H. hampei sebagai inang. Dalam laporan Cramer (1957) disebutkan bahwa intensitas serangan PBKo pada spesies-spesies komersial, terutama C. arabica dan C. canephora lebih tinggi dibandingkan pada spesies lain yang kurang dibudi-dayakan seperti C. dyowskii, C. stenophylla. Chahal & Gosal (2002) menyebutkan bahwa fenomena ketahanan kebal umumnya dijumpai pada spesies-spesies tanaman
liar atau pun spesies tanaman yang tidak dibudidayakan.
Perbedaan intensitas serangan hama PBKo antarspesies tanaman atau antarklon yang dilaporkan (Cramer, 1957; Mathieu et al., 1997; Laporan PHT, 2000; Prakasan et al., 2001) menunjukkan adanya per-bedaan respons ketahanan antarspesies ataupun antarklon dalam spesies terhadap infestasi H. hampei. Mekanisme ketahanan tersebut diduga bersifat antixenosis dan antibiosis. Perbedaan fenologi buah dalam hal bentuk, intensitas warna biji, dan aroma diduga berpengaruh dalam mengendalikan mekanisme antixenosis ini. Di antara karak-ter buah yang menunjukkan keragaman tinggi antar spesies kopi adalah diskus buah. Keragaman diskus buah tersebut diduga ikut berperan dalam mempengaruhi presepsi kumbang H. hampei dalam proses penemuan inangnya. Selain bentuk buah, hasil penelitian yang menunjukkan adanya
Gambar 2. Fenologi diskus buah kopi yang diduga berpengaruh terhadap presepsi H. hampei dalam penemuan tanaman inang
perbedaan tingkat kesukaan H. hampei terhadap warna (Mathieu et al., 1997; Wiryadiputra, 2006) dan senyawa volatil biji (Giordanengo et al., 1993) merupakan faktor yang berperan dalam pengaturan mekanisme anixenosis ini. Preferensi H. hampei ini terkait dengan aktivitas makan dan peletakan telur. Laporan Cramer (1957) dan Bagpro PHT (2000) yang menyebutkan bahwa klon-klon spesies C. canephora yang dilaporkan tahan PBKo memiliki kulit tanduk (parchment) yang keras menunjukkan bahwa mekanisme antibiosis juga ikut berperan dalam pengaturan ketahanan hama PBKo.
Mekanisme ketahanan semu yang di-pengaruhi oleh faktor ekologi dan sifat pembungaan tanaman dapat menghindarkan tanaman dari infestasi kumbang H. hampei. Mekanisme ini terkait dengan keserempak-an waktu pemasakkeserempak-an buah, tinggi tempat penanaman, dan manajemen lingkungan. Waktu pemasakan buah yang serempak dapat menyebabkan terputusnya siklus hidup H. hampei karena ketersediaan buah untuk infestasi H. hampei di pertanaman terbatas sehingga akan mengurangi populasi kumbang H. hampei. Dalam rekomendasi penanaman dianjurkan menanam varietas/ klon yang memiliki sifat pemasakan buah serempak. Klon/varietas yang dianjurkan adalah USDA 230762 untuk kopi Arabika, dan klon BP 42, BP 288, dan BP 234 untuk kopi Robusta (Sulistyowati, 1992). Faktor tinggi tempat penanaman juga berperan mengatur mekanisme ketahanan semu ini. Dalam hal ini pada lokasi-lokasi dengan ketinggian tempat yang kurang mendukung perkembangan H. hampei akan
meng-hindarkan tanaman dari sebaran H. hampei, seperti daerah-daerah berketinggian tempat >1.600 m dpl. yang cocok untuk per-tanaman kopi arabika (Cramer, 1957). Sedangkan faktor lingkungan yang ber-pengaruh adalah kondisi penaung. Dalam hal ini pertanaman yang penaungannya kurang (terbuka) dapat menghambat perkembangan H. hampei.
STRATEGI PEMULIAAN KETAHANAN HAMA PBKO
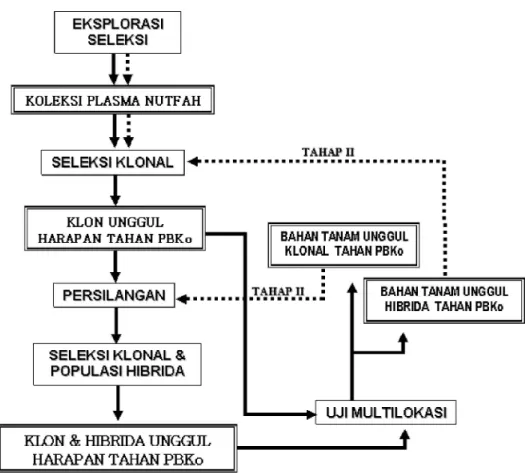
Masalah hama PBKo di Indonesia lebih banyak ditemukan pada pertanaman kopi Robusta (C. canephora) daripada kopi Arabika (C. arabica) sehingga program pemuliaan diarahkan untuk mendapatkan bahan tanam kopi Robusta tahan hama PBKo. Sasaran kegiatan adalah mendapat-kan bahan tanam dalam bentuk klonal atau pun hibrida F1 (biklonal/poliklonal) unggul dalam hal produktivitas, kualitas hasil dan ketahanan hama PBKo. Terkait dengan sifat penyer-bukan kopi robusta yang menyerbuk silang maka metode seleksi berulang (recur-rent selection) dapat diterapkan untuk program pemuliaan ketahanan hama PBKo. Dalam hal ini penerapan metode seleksi berulang dimodifikasi sesuai kondisi daur hidup tanaman kopi yang panjang. Sasaran mendapatkan klon dan hibrida tahan PBKo dapat dicapai sekaligus dalam setiap daur seleksi. Pada Gambar 3 terlihat tahapan-tahapan kegiatan yang akan dilakukan dalam daur seleksi berulang pemuliaan ketahanan hama PBKo.
Sumber genetik ketahanan hama PBKo diperoleh dari hasil eksplorasi dan seleksi.
Klon-klon tahan terseleksi selanjutnya akan digunakan sebagai induk persilangan untuk membentuk bahan tanam baru ataupun dapat dikategorikan sebagai genotipe unggul harapan bila telah memenuhi kriteria keunggulan bahan tanam unggul kopi. Persilangan bertujuan membentuk rekom-binasi baru untuk seleksi populasi atau pun seleksi individual. Dalam hal ini per-silangan juga dirancang untuk mendeteksi dayagabung antartetua klonal. Rancangan persilangan diallel umumnya digunakan
untuk tujuan tersebut. Bahan tanam unggul, baik hibrida maupun klonal, sebelum direkomendasikan kepada pengguna (petani/pekebun) terlebih dahulu dilakukan uji multilokasi untuk mengetahui adap-tabilitas bahan tanam unggul di berbagai lokasi. Setiap daur pemuliaan dengan cara ini membutuhkan waktu sekitar 15—20 tahun.
Hasil seleksi generasi awal pada kopi Robusta diperoleh klon SA13 dan SA34 yang menunjukkan tahan hama PBKo
Gambar 3. Skema metode seleksi berulang (recurrent selection) pemuliaan ketahanan hama PBKo pada tanaman kopi Robusta (C. canephora).
(Cramer, 1957). Namun demikian hingga kini klon-klon tersebut belum dimanfaatkan dalam program pemuliaan untuk menghasilkan bahan tanam tahan PBKo. Klon-klon tersebut juga dapat digunakan sebagai pembanding sifat ketahanan hama PBKo dalam setiap tahapan seleksi klon-klon koleksi plasma nutfah. Bagpro PHT (2000) melaporkan hasil uji saring pada klon-klon kopi Robusta tahan PBKo pada koleksi plasma nutfah di Kebun Bangelan dan KP Sumber Asin, Jawa Timur. Hasil seleksi tersebut mendapatkan beberapa klon yang menunjukkan respons tahan seperti klon SA13 dan SA34, adalah SA 13-02, SA 34-01, SA 53, SA94, SA 129-05, SA 129-OP, SA 142, SA 245, SA 254, Wringin Anom 2, Wringin Anom 3, Wringin Anom 8, dan BGN 371. Seleksi ini baru mendasarkan pada sifat ketahanan PBKo sehingga perlu ditindaklanjuti dengan evaluasi sifat dayahasil dan mutuhasil dalam proses seleksi tahap lanjut.
Klon-klon tersebut merupakan materi genetik dasar untuk proses perakitan bahan tanam tahan PBKo. Hingga kini belum diketahui informasi pewarisan ketahanan hama PBKo dan gen-gen pengatur ke-tahanan PBKo pada klon-klon tersebut. Pemilihan tetua tahan sebagai sumber gen ketahanan ditentukan berdasarkan derajat ketahanan. Melalui persilangan akan ter-bentuk rekombinasi genetik yang merupakan gabungan antara sifat tahan dan sifat-sifat unggul lainnya. Apabila sifat ketahanan PBKo diatur oleh gen dominan maka per-silangan dengan tetua tahan akan meng-hasilkan populasi F1 tahan PBKo. Sedangkan apabila sifat ketahanan PBKo
diatur oleh banyak gen (polygenic) maka perlu pengabungan gen-gen ketahanan ke dalam genotipe berbasis populasi maupun individu. Karena itu dalam pemilihan tetua persilangan perlu mempertimbangkan sumber gen ketahanan berasal lebih dari satu genotipe tahan.
P E NU T U P
Hama PBKo hingga saat ini masih menjadi masalah penting pada pertanaman kopi di Indonesia. Perakitan teknologi pengendalian hama PBKo belum mengarah pada upaya pencarian bahan tanam tahan sebagai komponen teknologi pengendalian yang efektif dan ramah lingkungan. Informasi mekanisme ketahanan hama PBKo sebagai landasan dalam penentuan strategi pengen-dalian maupun perakitan bahan tanam tahan belum banyak terungkap dalam hasil-hasil penelitian. Demikian pula klon-klon hasil seleksi sebagai sumber gen ketahanan belum termanfaatkan secara optimal dalam pe-rakitan bahan tanam tahan dan studi mekanisme ketahanan. Perlu langkah-langkah strategis dalam upaya pemanfaatan klon-klon tahan sehingga tercapai sasaran mendapatkan bahan tanam tahan PBKo dan informasi mekanisme ketahanan PBKo.
DAFTAR PUSTAKA
Bagian Proyek Penelitian PHT Tanaman Perkebunan (2000). Pengendalian
Hama Penggerek Buah Kopi (PBKo) Menggunakan Bahan Tanam Tahan.
Pusat Penelitian Kopi dan Kakao Indonesia, p. 28—38.
Cannell, M.G.R. (1985). Physiology of the coffee crop. p. 108—134. In: M.N.
Clifford & K.C. Wilson (Eds.).
Botany, Biochemistry of Beans and Beverage. AVI Publisihing,
Cobbec-ticut, USA.
Chahal, G.S. & S.S. Gosal (2002). Principle and Procedure of Plant Breeding: Bio-technological and Conventional Ap-proach. Alpha Science Int., Harrow U . K .
Charrier, A. & J. Berthaud (1985). Botani-cal classification of Coffea. p. 13— 47. In: M.N. Clifford & K.C.
Wil-son (Eds.). Botany, Biochemistry of
Beans and Beverage. AVI
Publish-ing, Cobbec-ticut, USA.
Damon, A. (2000). A review of the biology and control of the coffee berry borer,
Hypothenemus hampei (Coleoptera:
Scolytidae). Bulletin of
Entomologi-cal Research, 90, 453—465.
Durham, S. (2004). Stopping the Coffee Berry Borer from Boring into Profits. Agric.
Research, 52, 10—11.
Giordanengo, P.; Luc O. Brun; B. Frerot (1993). Evidence for allelchemical at-traction of the coffee berry borer, Hypothenemus hampei, by coffee ber-ries. J. of Chemical Eco., 19, 763— 769.
Mathieu, F.; L.O. Burn; C. Marcillaud & B. Frérot (1997). Trapping of the coffee berry borer within a mesh-enclosed environment: interaction of olfactory and visual stimuli. J. Appl. Ent. 121, 181—186.
Panda, N. & G.S. Khush (1995). Host Plant Resistance to Insects. 1st Edt. CAB International, International Rice Re-search Institute, Manila.
Renwick, J.A.A. & F.S. Chew (1994). Ovi-position Behavior in Lepidoptera.
Annu. Rev. Entomol., 39, 377—400.
Rothfos, B. (1980). Coffee Production. Niedersächsische buchdruckerei, Ger-many. 366 p.
Sulistyowati, E. (1992). Hama Utama
Tanam-an Kopi dTanam-an Cara PengendaliTanam-annya. In: Buku III : Bahan pelatihan teknik
budidaya dan pengolahan kopi. Pusat Penelitian Perkebunan, Jember. Swarupa, S.G.; A.G.S. Reddy & P. C. Shekar
(2000). A compendium of coffee
re-search in India. Central Coffee
Re-search Institute, India.
Wilson, K.C. (1985). Climate and soil. p. 97— 107. In: M.N. Clifford & K.C. Wilson
(Eds.). Botany, biochemistry of beans
and beverage. AVI Publishing,
Cobbecticut, USA.
Wiryadiputra, S. (2006). Penggunaan perangkap dalam penggendalian hama penggerek buah kopi (Hypothenemus hampei).
Pelita Perkebunan, 22, 101—118.
Venkatesha, M.G.; H.G. Seetharama & K. Sreedharan (1998). Coffee Pests and
Their Management. p. 1—28. In: C.S.
Srinivasan (Ed.). A Compendium on
Pests and Diseases of Coffee and Their Management in India. Central
Coffee Research Institute. Karnataka. * * * * * * * *