FORDA - Jurnal
Teks penuh
Gambar
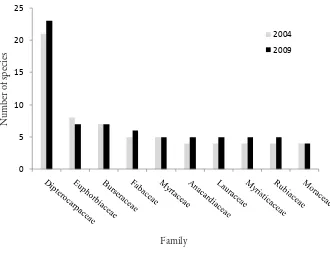
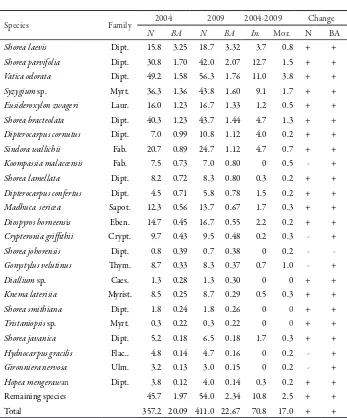
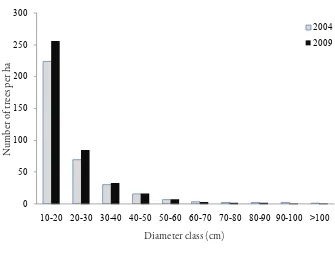
Garis besar
Dokumen terkait
• Pembuatan sambungan dengan menggunakan alat pertukangan baik secara manual maupun masinal ( Portabel atau Stationer ) secara benar dan aman berbagai jenis sambungan
28 Table 3.1 Categories for Student Questionnaire in Needs Analysis 51 Table 3.2 Categories for Interview with English Teacher in Needs Analysis 52 Table 3.4 Categories for
6.1.3 For the operation into the intermediate and/or beyond point(s) each Participating Party shall agree to designate fifth freedom traffic rights for passenger and
Altruism Ratio – atau yang akhir-akhir ini sering disebut Local Wisdom - tersebut kini terancam oleh berdirinya korporasi yang akan membeli lahan mereka, karena Pegunungan
KENCANA MULIA SEMESTA 15 Juli 2011 AKADEMI TEKNIK INDUSTRI MAKASSAR PANITIA PENGADAAN
Struktur yang berbasis fungsi yang diterapkan kepada KPP dengan sistem administrasi yang modern untuk dapat merealisasikan debirokratisasi pelayanan sekaligus
Keenam peristiwa dalam kehidupan manusia itu yakni (1) selamatan dalam siklus hidup seseorang, seperti hamil tujuh bulan, kelahiran, upacara potong rambut, upacara turun
Berdasarkan hasil diskusi yang digambarkan menggunakan fishbone seperti diatasselama sesi brainstorming yang dilakukan dengan mahasiswa Pegunungan Bintang, maka dapat
