PEMANFAATAN MIKROBA DALAM BIOREMEDIASI:
SUATU TEKNOLOGI ALTERNATIF UNTUK
PELESTARIAN LINGKUNGAN
Pidato Pengukuhan
Jabatan Guru Besar Tetap
dalam Bidang Mikrobiologi
pada Fakultas Matematika dan Ilmu Pengetahuan Alam,
Diucapkan di Hadapan Rapat Terbuka Universitas Sumatera Utara
Gelanggang Mahasiswa, Kampus USU, 1 Mei 2006
Oleh:
ERMAN MUNIR
UNIVERSITAS SUMATERA UTARA
M E D A N
Yang terhormat,
Bapak Menteri Pendidikan Nasional Republik Indonesia,
Bapak Ketua dan Anggota Majelis Wali Amanat Universitas Sumatera Utara, Bapak Ketua dan Anggota Senat Akademik Universitas Sumatera Utara, Bapak Ketua dan Anggota Dewan Guru Besar Universitas Sumatera Utara, Bapak Rektor Universitas Sumatera Utara,
Bapak/Ibu para Pembantu Rektor Universitas Sumatera Utara, para Dekan, Ketua Lembaga dan Unit Kerja, para Dosen dan Karyawan di lingkungan Universitas Sumatera Utara,
Bapak dan Ibu para undangan, keluarga, teman sejawat, mahasiswa, dan hadirin yang saya muliakan.
Assalamu’alaikum Warahmatullahi Wabarakatuh
Pada hari yang berbahagia ini, pertama kali saya memanjatkan puji dan syukur ke hadirat Allah SWT yang telah dan senantiasa memberikan karunia dan nikmat-Nya kepada kita semua sehingga kita dapat hadir pada kesempatan ini. Serta selawat dan salam kepada Nabi Muhammad SAW beserta keluarga dan sahabat-sahabatnya yang kita harapkan syafaatnya di kemudian hari.
Atas izin dan rida-Nya perkenankanlah saya menyampaikan pidato pengukuhan sebagai Guru Besar Tetap pada Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Sumatera Utara ini dengan judul:
PEMANFAATAN MIKROBA DALAM BIOREMEDIASI:
SUATU TEKNOLOGI ALTERNATIF UNTUK PELESTARIAN LINGKUNGAN
PENDAHULUAN
Dapatkah kita hidup tanpa mikroba? Banyak orang tidak suka dengan mikroba. Mendengar kata mikroba, yang terbayang bagi kita adalah kerugian yang ditimbulkannya seperti penyakit, pencemaran, dan juga kerusakan makanan. Mungkin sedikit orang yang memahami manfaatnya. Mikroba ada di mana-mana: di air, dalam tanah, di kulit, dalam mulut, dalam saluran pencernaan, di lantai rumah, di pakaian, dan lain-lain. Bahkan udara yang kita hirup sehari-hari dihuni oleh mikroba, yang kehadirannya memberikan kecemasan kepada manusia. Apakah memang begitu mengerikannya?
Walaupun banyak mikroba yang menyebabkan kerugian kepada makhluk hidup seperti hewan, tumbuhan, dan manusia, kontribusi mikroba terhadap kelangsungan kehidupan di permukaan bumi ini tidak ternilai besarnya. Contoh sederhana, tanpa mikroba maka antibiotika dan vaksin tidak akan pernah ada. Tanpa peran mikroba yang ada dalam lambung hewan memamah biak, kita tidak akan pernah makan daging sapi dan beberapa jenis makanan hasil fermentasi mikroba seperti tempe, kecap, terasi,
yoghurt, minuman beralkohol, dan masih banyak lagi yang mungkin tidak
akan pernah kita konsumsi tanpa adanya mikroba.
Di lingkungan, mikroba memiliki fungsi atau peranan yang cukup beragam. Dari penghasil oksigen di ekosistem perairan, berperan dalam siklus biogeokimia, membantu tanaman dalam penyerapan unsur hara, sebagai pengurai, sampai pada degradasi atau remediasi polutan. Buku ini membahas peranan mikroba dalam degradasi beberapa polutan.
Pencemaran atau polusi bukanlah merupakan hal baru, bahkan tidak sedikit dari kita yang sudah memahami pengaruh yang ditimbulkan oleh pencemaran atau polusi lingkungan terhadap kelangsungan dan keseimbangan ekosistem. Polusi dapat didefinisikan sebagai kontaminasi lingkungan oleh bahan-bahan yang dapat mengganggu kesehatan manusia, kualitas kehidupan, dan juga fungsi alami dari ekosistem. Walaupun pencemaran lingkungan dapat disebabkan oleh proses alami, aktivitas manusia yang notabenenya sebagai pengguna lingkungan adalah sangat dominan sebagai penyebabnya, baik yang dilakukan secara sengaja ataupun tidak.
Berdasarkan kemampuan terdegradasinya di lingkungan, polutan digolongkan atas dua golongan:
1. Polutan yang mudah terdegradasi (biodegradable pollutant), yaitu bahan seperti sampah yang mudah terdegradasi di lingkungan. Jenis polutan ini akan menimbulkan masalah lingkungan bila kecepatan produksinya lebih cepat dari kecepatan degradasinya. 2. Polutan yang sukar terdegradasi atau lambat sekali terdegradasi
(nondegradable pollutant), dapat menimbulkan masalah lingkungan
yang cukup serius.
Bahan polutan yang banyak dibuang ke lingkungan terdiri dari bahan pelarut (kloroform, karbontetraklorida), pestisida (DDT, lindane), herbisida (aroklor, antrazin, 2,4-D), fungisida (pentaklorofenol), insektisida (organofosfat), petrokimia (polycyclic aromatic hydrocarbon [PAH], benzena,
toluena, xilena), polychlorinated biphenyls (PCBs), logam berat, bahan-bahan radioaktif, dan masih banyak lagi bahan-bahan berbahaya yang dibuang ke lingkungan, seperti yang tertera dalam lampiran Peraturan Pemerintah RI Nomor 18 Tahun 1999 tentang Pengelolaan Bahan Berbahaya dan Beracun.
APAKAH YANG DIMAKSUD DENGAN BIOREMEDIASI?
Bioremediasi merupakan pengembangan dari bidang bioteknologi lingkungan dengan memanfaatkan proses biologi dalam mengendalikan pencemaran. Bioremediasi bukanlah konsep baru dalam mikrobiologi terapan, karena mikroba telah banyak digunakan selama bertahun-tahun dalam mengurangi senyawa organik dan bahan beracun baik yang berasal dari limbah rumah tangga maupun dari industri. Hal yang baru adalah bahwa teknik bioremediasi terbukti sangat efektif dan murah dari sisi ekonomi untuk membersihkan tanah dan air yang terkontaminasi oleh senyawa-senyawa kimia toksik atau beracun.
Keberhasilan proses bioremediasi harus didukung oleh disiplin ilmu lain seperti fisiologi mikroba, ekologi, kimia organik, biokimia, genetika molekuler, kimia air, kimia tanah, dan juga teknik. Mikroba yang sering digunakan dalam proses bioremediasi adalah bakteri, jamur, yis, dan alga. Degradasi senyawa kimia oleh mikroba di lingkungan merupakan proses yang sangat penting untuk mengurangi kadar bahan-bahan berbahaya di lingkungan, yang berlangsung melalui suatu seri reaksi kimia yang cukup kompleks. Dalam proses degradasinya, mikroba menggunakan senyawa kimia tersebut untuk pertumbuhan dan reproduksinya melalui berbagai proses oksidasi.
DAPATKAH DIBAYANGKAN BILA TIDAK ADA MIKROBA PENGURAI DI MUKA BUMI INI?
Tanpa adanya mikroba, proses penguraian di lingkungan tidak akan berlangsung. Kotoran, sampah, hewan, dan tumbuhan yang mati akan menutupi permukaan bumi, suatu kondisi yang tidak akan pernah kita harapkan. Sebagai akibatnya, siklus nutrisi atau rantai makanan akan terputus.
Lintasan biodegradasi berbagai senyawa kimia yang berbahaya dapat dimengerti berdasarkan lintasan mekanisme dari beberapa senyawa kimia alami seperti hidrokarbon, lignin, selulosa, dan hemiselulosa. Sebagian
besar dari prosesnya, terutama tahap akhir metabolisme, umumnya berlangsung melalui proses yang sama. Polimer alami yang mendapat perhatian karena sukar terdegradasi di lingkungan adalah lignoselulosa (kayu) terutama bagian ligninnya.
DEGRADASI MATERIAL LIGNIN
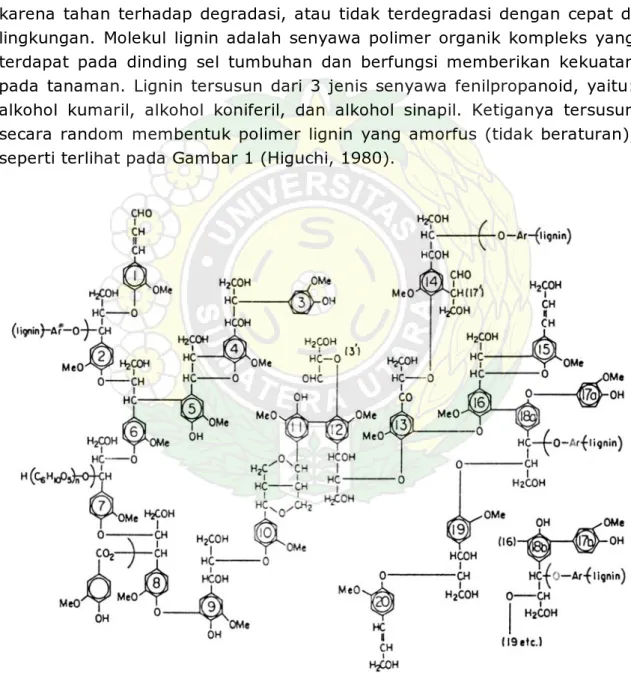
Lignin adalah polimer alami dan tergolong ke dalam senyawa rekalsitran karena tahan terhadap degradasi, atau tidak terdegradasi dengan cepat di lingkungan. Molekul lignin adalah senyawa polimer organik kompleks yang terdapat pada dinding sel tumbuhan dan berfungsi memberikan kekuatan pada tanaman. Lignin tersusun dari 3 jenis senyawa fenilpropanoid, yaitu: alkohol kumaril, alkohol koniferil, dan alkohol sinapil. Ketiganya tersusun secara random membentuk polimer lignin yang amorfus (tidak beraturan), seperti terlihat pada Gambar 1 (Higuchi, 1980).
Jamur basidiomisetes merupakan kelompok utama pendegradasi lignoselulosa. Walaupun beberapa bakteri diketahui dapat mendegradasi lignin, tetapi bakteri yang mampu mendegradasi lignin secara kompleks belum pernah dilaporkan. Jamur pembusuk kayu menghasilkan enzim-enzim pendegradasi lignoselulosa seperti golongan selulase, ligninase, dan hemiselulase.
Berdasarkan mekanisme degradasi, jamur pembusuk kayu digolongkan ke dalam jamur pembusuk putih dan jamur pembusuk cokelat, yang masing-masing memiliki metabolisme degradatif yang berbeda. Jamur busuk putih mampu mendegradasi seluruh komponen material lignoselulosa termasuk lignin, sedang jamur busuk cokelat lebih cenderung mendegradasi bagian selulosa dan hemiselulosa tetapi tidak lignin (Green and Highley, 1997).
Penggunaan kultur campuran antara jamur pembusuk putih dan jamur pembusuk cokelat memiliki prospek yang cukup tinggi untuk mendapatkan glukosa alternatif dari material lignoselulosa (Munir dan Goenadi, 1999). Cooke and Rayner (1984), jamur basidiomisetes dan askomisetes memiliki peran yang utama dalam degradasi lignoselulosa yang setiap tahunnya diperkirakan terbentuk sebanyak 100 gigaton, di mana 20 gigatonnya adalah lignin.
BAGAIMANA MIKROBA DAPAT MENDEGRADASI LIGNIN YANG BEGITU KOMPLEKS?
Pada Gambar 1 terlihat monomer-monomer pembentuk lignin tersusun secara tidak beraturan sehingga sukar untuk didegradasi oleh mikroba, seperti halnya pada degradasi molekul selulosa dan kitin. Suatu pendapat menyatakan bahwa jamur busuk putih mendegradasi lignin adalah untuk mendapatkan selulosa dari material lignoselulosa. Ketidakteraturan struktur lignin ini menyebabkan proses degradasi menjadi sangat kompleks, dan enzim-enzim yang berperan dalam degradasi lignin bekerja secara nonspesifik. Proses ini berlangsung melalui pembentukan radikal-radikal bebas yang dapat menyerang sejumlah besar molekul organik. Hal ini menyebabkan jamur pendegradasi lignin mendapat perhatian yang sangat besar dalam biodegradasi berbagai jenis polutan organik.
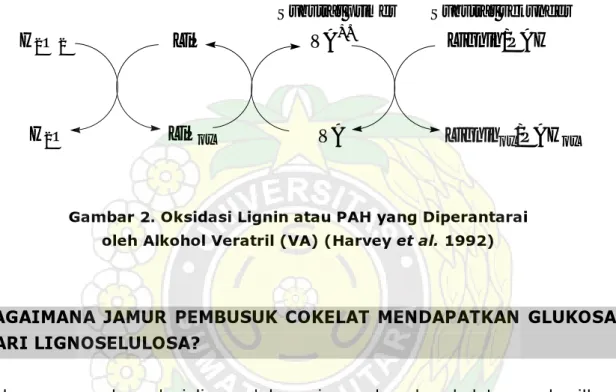
Filed et al. (1993); Evans et al. (1994) menyatakan bahwa kelompok peroksidase (lignin peroksidase [LiP] dan manganese peroksidase [MnP]) yang menggunakan H2O2 dan lakase (polifenol oksidase) yang
menggunakan molekul oksigen berperan dalam degradasi lignin. Gambar 2 berikut menunjukkan seri oksidasi lignin atau hidrokarbon poliaromatik (PAH). Radikal alkohol veratril (VA+.) yang dihasilkan adalah sebagai produk
utama oksidasi H2O2 yang dikatalisis oleh LiP.
LiP
VA
+.Lignin/PAH
H
2O
2H
2O
LiP
oxVA
Lignin
ox/PAH
oxSubstrat primer Substrat sekunder
Gambar 2. Oksidasi Lignin atau PAH yang Diperantarai oleh Alkohol Veratril (VA) (Harvey et al. 1992)
BAGAIMANA JAMUR PEMBUSUK COKELAT MENDAPATKAN GLUKOSA DARI LIGNOSELULOSA?
Dalam proses degradasi lignoselulosa, jamur busuk cokelat menghasilkan sejumlah besar asam oksalat (COOH)2. Hal ini menyebabkan turunnya pH
lingkungan yang cukup drastis, yang selanjutnya menyebabkan hidrolisis selulosa secara nonenzimatik (Shimada et al. 1991). Proses ini sangat penting karena aktivitas enzim selulase belum dapat berlangsung sempurna karena enzim ini tidak dapat menembus pori-pori dinding sel yang ukurannya lebih kecil dari ukuran enzim. Peranan jamur penghasil asam oksalat dalam bioremediasi lingkungan dibicarakan pada bagian lain dalam buku ini.
BAGAIMANA ASAM OKSALAT DISINTESIS OLEH JAMUR?
Baru-baru ini telah ditemukan proses biosintesis asam oksalat oleh jamur pembusuk cokelat. Biosintesis asam oksalat merupakan proses fisiologis yang sangat penting bagi jamur, di mana jamur memperoleh energi dengan mengoksidasi karbohidrat menjadi asam oksalat (Munir et al. 2001a), seperti yang terlihat dalam persamaan reaksi berikut:
C6H12O6 + 5O2 2(COOH)2 + 2CO2 + 4H2O
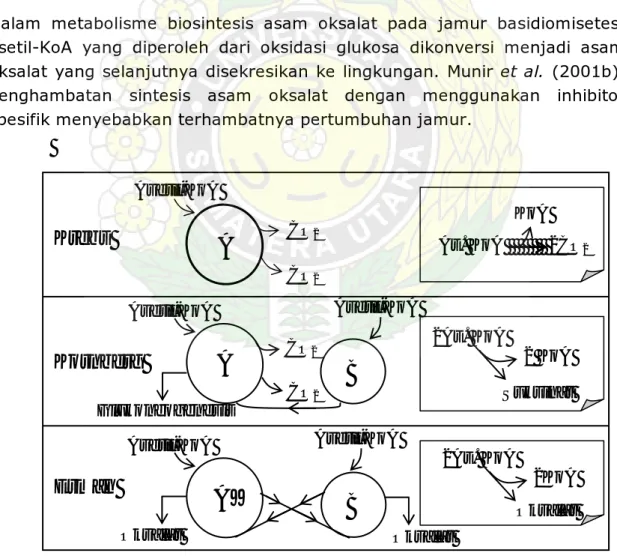
Dalam tulisan tersebut juga dikemukakan tentang lintasan metabolisme asam oksalat pada jamur pembusuk kayu. Lintasan metabolisme ini mendapat perhatian yang cukup besar bagi kalangan biokemis. Gambar 3 berikut menunjukkan perbandingan lintasan metabolisme asam oksalat pada jamur dengan siklus Krebs dan siklus Glioksilat (Kornberg).
Dalam metabolisme biosintesis asam oksalat pada jamur basidiomisetes, asetil-KoA yang diperoleh dari oksidasi glukosa dikonversi menjadi asam oksalat yang selanjutnya disekresikan ke lingkungan. Munir et al. (2001b), penghambatan sintesis asam oksalat dengan menggunakan inhibitor spesifik menyebabkan terhambatnya pertumbuhan jamur.
Asetil-KoA CO2 CO2 Asetil-KoA Glukoneogenesis Asetil-KoA CO2 CO2 Asetil-KoA Oksalat Oksalat Asetil-KoA
Krebs
Kornberg
Erman
2As. KoA KoA 2 KoA As. KoA 2CO2Suksinat 2As. KoA 2KoA Oksalat
A
A
A’
B
B
Gambar 3. Metabolisme Asetil-KoA pada Siklus Krebs dan Analognya: A, Siklus Krebs; A’, Modifikasi Siklus Krebs; B, Siklus Glioksilat
MENGAPA JAMUR BUSUK PUTIH SANGAT BERPOTENSI DALAM DEGRADASI POLUTAN?
Karena kemampuannya dalam mendegradasi berbagai senyawa aromatik, jamur pendegradasi lignin telah mendapat perhatian besar dalam bidang bioremediasi. Sistem degradasi enzimatis ekstraseluler menyebabkan jamur busuk putih lebih toleran terhadap konsentrasi polutan toksik yang lebih tinggi. Selanjutnya, mekanisme degradasi nonspesifik yang dimiliki oleh jamur pembusuk putih menyebabkan mereka mampu mendegradasi sejumlah besar polutan. Keunggulan lain dari jamur pembusuk putih dalam degradasi polutan adalah mereka tidak memerlukan pengkondisian untuk polutan tertentu, karena kekurangan nutrien dapat menginduksi proses degradasi. Di samping itu, induksi sintesis enzim-enzim pendegradasi polutan biasanya tidak terpengaruh oleh banyak sedikitnya polutan (Barr and Aust, 1994).
BIOREMEDIASI POLUTAN YANG SUKAR TERDEGRADASI
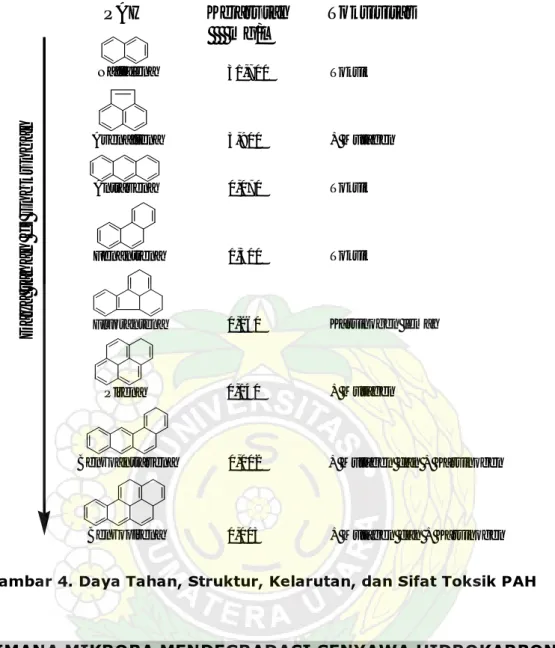
Toksisitas senyawa hidrokarbon seperti hidrokarbon minyak (bensin) terhadap mikroba, tumbuhan, hewan, dan manusia telah banyak dipelajari. Senyawa hidrokarbon aromatis polisiklis (PAH) dalam minyak memiliki toksisitas yang cukup tinggi. Efek toksik dari hidrokarbon yang terdapat dalam minyak berlangsung melalui larutnya lapisan lemak yang menyusun membran sel, sehingga menyebabkan hilangnya cairan sel atau kematian terhadap sel (Rosenberg and Ron, 1998). Ketahanan PAH di lingkungan dan toksisitasnya meningkat sejalan dengan peningkatan jumlah cincin benzenanya (Mueller et al. 1998), seperti terlihat pada Gambar 4.
Di samping itu, PAH terikat kuat pada material organik tanah dan kelarutannya juga rendah. Hal ini menyebabkan ketersediaannya untuk degradasi oleh mikroba menjadi terbatas. Gambar ini selanjutnya menujukkan bahwa benzopirena dengan lima cincin benzena lebih sukar terdegradasi bila dibandingkan dengan naftalena yang memiliki dua cincin benzena.
Naftalena Asenaftena Antrasena Fenantrena Benzopirena Benzoantrasena Pirena Fluorantena PAH Da y a t ah an d i li n g k u n g an Kelarutan mg/l Toksisitas 31,700 3,900 0,070 1,300 0,140 0,260 0,002 0,003 Toksik Toksik Toksik Karsinogen lemah
+ Mutagen dan + Karsinogen + Mutagen dan + Karsinogen
+ Mutagen + Mutagen
Gambar 4. Daya Tahan, Struktur, Kelarutan, dan Sifat Toksik PAH
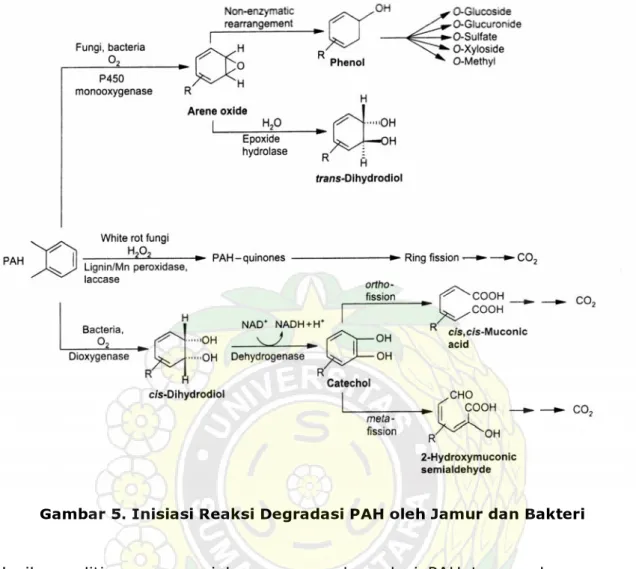
BAGAIMANA MIKROBA MENDEGRADASI SENYAWA HIDROKARBON?
Beberapa golongan mikroorganisme telah diketahui memiliki kemampuan dalam memetabolisme PAH. Bakteri dan beberapa alga menggunakan dua molekul oksigen untuk memulai pemecahan cincin benzena PAH, yang dikatalis oleh enzim dioksigenase untuk membentuk molekul cis-dihidrodiol. Kebanyakan jamur mengoksidasi PAH melalui pemberian satu molekul oksigen untuk membentuk senyawa oksida aren yang dikatalisis oleh sitokrom P-450 monooksigenase. Pada jamur busuk putih, bila terdapat H2O2, enzim lignin peroksidase yang dihasilkan akan menarik satu elektron
dari PAH yang selanjutnya membentuk senyawa kuinon (Cerniglia and Sutherland, 2001). Cincin benzena yang sudah terlepas dari PAH selanjutnya dioksidasi menjadi molekul-molekul lain dan digunakan oleh sel mikroba sebagai sumber energi. Gambar 5 berikut menunjukkan lintasan metabolisme PAH oleh mikroorganisme.
Gambar 5. Inisiasi Reaksi Degradasi PAH oleh Jamur dan Bakteri
Hasil penelitian mengenai kemampuan degradasi PAH termasuk senyawa aromatik klor, nitroaromatik, zat warna, pestisida, dan pencemar lingkungan lainnya oleh jamur dan bakteri cukup banyak dilaporkan. Untuk keperluan lebih lanjut dapat dirujuk tulisan Cerniglia and Sutherland (2001) dan Mueller et al. (1998). Selanjutnya, Mueller et al. menyatakan bahwa bioremediasi senyawa PAH dapat ditempuh melalui tiga metode berikut, yaitu: fase padat (solid-phase), dengan menggunakan bioreaktor, dan proses in situ. Masing-masing metode memiliki kelebihan dan batasan.
APAKAH MIKROBA RIZOSFER JUGA BERPERAN DALAM DEGRADASI POLUTAN?
Mikroorganisme tanah seperti jamur, bakteri, aktinomisetes, dan protozoa merupakan komponen yang sangat penting dalam ekosistem tanah karena mereka memiliki peranan utama dalam siklus nutrisi, mempertahankan
struktur tanah, dan juga mengatur pertumbuhan tanaman melalui berbagai mekanisme.
Aktivitas dan populasi mikroorganisme sekitar perakaran tanaman (rizosfer) biasanya lebih dinamis dari daerah nonrizosfer. Hal ini disebabkan oleh adanya molekul organik seperti gula dan asam organik yang dikeluarkan oleh akar atau produk regenerasi dari akar yang dapat dimanfaatkan oleh mikroorganisme tanah. Tanpa adanya sekresi dari akar, mikroba di sekitar rizosfer akan sukar bertahan dalam ekosistem tanah.
Kelompok mikroba yang memiliki fungsi penting di daerah rizosfer adalah jamur, bakteri, dan protozoa yang membantu pertumbuhan tanaman melalui berbagai mekanisme seperti peningkatan penyerapan nutrisi, sebagai kontrol biologi terhadap serangan patogen, dan juga menghasilkan hormon pertumbuhan bagi tanaman (Chanway, 1997). Kajian mengenai peranan bakteri tanah yang hidup bebas seperti Pesudomonas, Bacillus,
Agrobacterium, dan Erwinia dalam mengurangi serangan patogen telah
banyak dilaporkan. Reddy et al. (1994) melaporkan bahwa serangan jamur
Fusarium oxysporum terhadap pertumbuhan bibit Douglas-fir menurun
tajam setelah diinokulasi beberapa strain Pseudomonas.
Bakteri simbiotik dari genus Rhizobium dan Barahyrhizobium, di samping telah dikenal luas sebagai bakteri penambat nitrogen bebas, juga memiliki kemampuan dalam mendegradasi senyawa-senyawa toksik di sekitar perakaran. Barkovskii et al. (1994) melaporkan bahwa Azospirillum yang juga memiliki kemampuan menambat nitrogen banyak mengkolonisasi berbagai jenis tanaman dapat mendegradasi senyawa-senyawa fenol dan benzoat. Sehingga bakteri ini telah banyak digunakan secara komersial dalam bioremediasi tanah yang tercemar. Beberapa bakteri lain yang terdapat pada rizosfer, seperti: Achromobacter, Agrobacterium, Alcaligenes,
Acinetobacter, Azotobacter, Flavobacterium, Mycobaterium, Nitosomonas, Nocardia, Pseudomonas, dan Xanthobacter juga memiliki kemampuan
dalam metabolisme senyawa fenol, halogen, hidrokarbon, dan juga berbagai jenis pestisida.
Mikoriza sebagai suatu bentuk simbiosis mutualisme antara jamur dengan akar tanaman berperan dalam peningkatan ketersediaan nutrisi (terutama fosfat) bagi tanaman. Mikoriza juga dapat meningkatkan daya tahan tanaman terhadap serangan patogen tanah. Mikoriza dapat mengurangi toksisitas logam berat terhadap tanaman pada tanah-tanah tercemar.
Sehingga mikoriza juga memiliki peranan yang penting sebagai agen bioremediasi atau reklamasi bagi tanah-tanah yang tercemar oleh logam berat(Leyval et al., 1997), seperti pada lahan-lahan bekas tambang.
Sharples et al. (2000) melaporkan bahwa jamur pada daerah tambang berfungsi sebagai filter untuk menjaga agar konsentrasi As tetap rendah pada jaringan tanaman dan meningkatkan serapan P tanaman. Donelly and Fetcher (1994) melaporkan bahwa logam berat berikatan dengan gugus karboksil hemiselulosa pada matriks di antara sel tanaman dan jamur, sehingga tanaman terhindar dari keracunan. Selanjutnya, ia melaporkan bahwa beberapa jamur mikoriza seperti Rhizopogon vinicolor, Rhizopogon
vulgaris, Hymenoscyphus ericae, Oidiodendron griseum, dan Gautieria crispa memiliki kemampuan remediasi senyawa-senyawa toksik di tanah,
seperti dalam metabolisme berbagai senyawa aromatik: 2,4-D, atrazin, dan PCBs. Selanjutnya dinyatakan bahwa Radiigera atrogleba dan Hysterangium
gardneri mampu mendegradasi 2,2-diklorofenol sebesar 80% (Donelly and
Fetcher, 1994).
APAKAH MIKROBA DAPAT MENURUNKAN CEMARAN LOGAM DI LINGKUNGAN?
Beberapa logam tertentu memiliki peran penting dalam metabolisme mikroba, sedangkan yang lain tidak diketahui fungsinya. Akan tetapi, baik logam berat dan logam nonesensil akan bersifat toksik bila terdapat dalam jumlah yang sangat berlebihan. Karena sifat toksik logam, proses bioremediasi senyawa organik sering kali menjadi terhambat. Roane et al. (1998) menyatakan bahwa di antara logam-logam yang toksik tersebut terdiri dari kation-kation seperti merkuri, timbal, arsenat, boron, kadmium, kromium, tembaga, nikel, mangan, selenium, perak, dan seng. Proses bioremediasi logam di lingkungan berbeda dengan proses degradasi molekul-molekul hidrokarbon; logam bukan merupakan pembangun bagi komponen-komponen sel.
Peningkatan konsentrasi logam di lingkungan, terutama logam berat, menimbulkan efek yang cukup serius terhadap seluruh bentuk kehidupan. Bagi manusia gejala toksisitas logam berat dapat berupa kerusakan jantung, hati, kanker, kelainan dan kerusakan sistem syaraf, dan lain-lain. Pada tumbuhan keracunan logam dapat menyebabkan memendeknya akar, gugurnya daun, klorosis, kekurangan nutrisi, dan lain-lain. Bagi mikroba
kadar logam yang terlalu tinggi di lingkungan dapat menurunkan atau menghambat pertumbuhan mikroba.
Interaksi mikroba dengan logam berat menyebabkan perubahan-perubahan proses fisiologis yang sangat drastis dan dalam beberapa hal dapat membunuh mikroba. Mekanisme toksisitas di antaranya terjadi melalui pengikatan logam pada ligan-ligan sulfidril, karboksil, dan fosfat seperti protein dan asam nukleat. Untuk meminimalisasi toksisitas logam berat, jamur mengembangkan berbagai mekanisme pertahanan, seperti imobilisasi logam berat oleh molekul intrasel (fitokelatin dan metalotionin) dan imobilisasi oleh molekul ekstraseluler (asam-asam organik) yang dihasilkan oleh jamur (Baldrian, 2003).
Salah satu kelator yang dihasilkan oleh jamur dan sudah dikenal kemampuannya dalam mengikat logam adalah asam oksalat. Asam oksalat yang dihasilkan oleh mikroba dapat meningkatkan resistensi mikroba tersebut terhadap logam melalui pembentukan kompleks metal-oksalat yang bersifat tidak larut. Metal oksalat dapat terbentuk dengan Ca, Cd, Co, Cu, Mn, Sr, dan Zn (Sayer and Gadd, 1997). Selanjutnya juga telah banyak dilaporkan bahwa terdapat hubungan antara resistensi jamur terhadap logam dengan kemampuannya dalam menghasilkan asam oksalat.
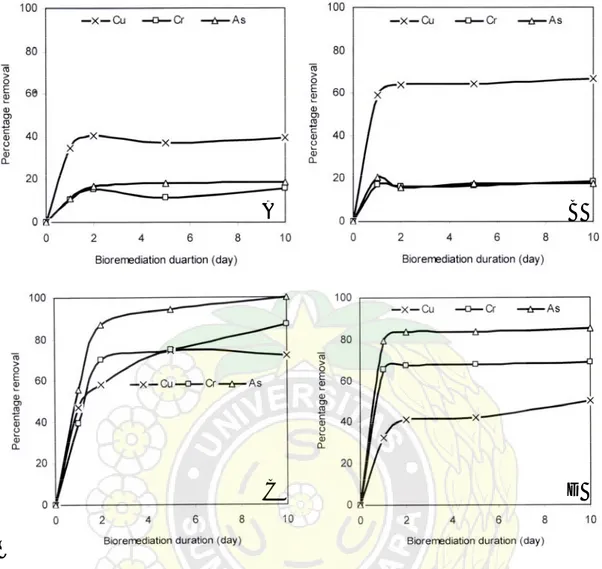
Munir et al. (2005) melaporkan bahwa biosintesis asam oksalat sangat penting untuk mendukung pertumbuhan jamur di bawah kondisi lingkungan yang tidak menguntungkan. Beberapa waktu yang lalu juga telah dilaporkan bahwa kultur pertumbuhan jamur busuk cokelat Tyromyces palustris,
Laetiporus suphureus, dan Coniphora puteana mampu menyerap Cu, Cr dan
As (CCA) dari kayu yang diawetkan, dan menurunkan kadar CCA dari kayu sampai di atas 75% (Kartal et al., 2003), seperti terlihat pada Gambar 6.
Dapatkah jamur digunakan sebagai alat untuk memonitor pencemaran logam di lingkungan? Karena potensinya dalam mengakumulasikan logam cukup besar, jamur pembusuk kayu dapat digunakan sebagai agen untuk monitor polusi logam di tanah atau di atmosfer atau sebagai alat analisis lingkungan yang cukup potensial. Gabriel et al. (1995) melaporkan bahwa terdapat hubungan yang erat antara polusi udara dengan kandungan logam dalam tubuh buah jamur (fruit body).
A
B
C
D
Gambar 6. Penurunan Kadar CCA Kayu yang Diawetkan Setelah Inkubasi dengan Kultur Pertumbuhan Jamur:
A, Kontrol; B, C. puteana; C, T. palustris; dan D, L. suphureus
Kemampuan bakteri dalam menyerap atau menurunkan kandungan logam berat dari lingkungan, baik dari tanah maupun dari perairan juga telah banyak dipelajari. Beberapa bakteri seperti Pseudomonas aeruginosa,
Acinetobacter calcoaceticus, Arthrobacter sp., Streptomyces viridans, dan
lain-lain menghasilkan senyawa biosurfaktan/bioemulsi yang dapat menyerap berbagai jenis logam berat seperti Cd, Cr, Pb, Cu, dan Zn dari tanah yang terkontaminasi. Desulfovibrio desulfuricans dapat mengendapkan uranium melalui proses reduksi.
Berbagai jenis Bacillus yang membentuk biofilm pada permukaan perairan dapat menyerap Cd, Cr, Cu, Hg, Ni, dan Zn dari dalam air. Mikroba yang membentuk film dalam ekosistem perairan juga memiliki peranan yang penting dalam bioremediasi logam. Saccharomyces cerevisiae dan Candida
sp. dapat mengakumulasikan Pb dari dalam perairan, Citrobacter dan
Rhizopus arrhizus memiliki kemampuan menyerap uranium (Roane et al.
1998).
BIOREMEDIASI SEBAGAI METODE ALTERNATIF PELESTARIAN LINGKUNGAN
Secara ekonomi dan fungsi, penggunaan teknik bioremediasi harus dapat berkompetisi dengan teknologi remediasi lainnya, seperti pembakaran (insinerasi) atau perlakuan kimia. Sebelum suatu teknik bioremediasi diaplikasikan, informasi tentang keadaan lokasi dan potensi mikroorganisme harus sudah diketahui. Untuk itu perlu dilakukan uji laboratorium untuk mengetahui kecepatan degradasi pada suatu fungsi lingkungan tertentu seperti pH, konsentrasi oksigen, nutrien, komposisi mikroba, ukuran partikel tanah, dan juga suhu.
Dibanding teknik remediasi lain, aplikasi bioremediasi jauh lebih murah. Levine and Gealt (1993) menyatakan bahwa bioremediasi untuk satu yard tanah yang terkontaminasi diperlukan dana sekitar 40 sampai 100 dolar. Sedangkan melalui proses lainnya, seperti dengan insinerasi, memerlukan biaya 250 sampai 800 dolar dan landfilling sekitar 150 sampai 250 dolar untuk kapasitas tanah yang sama. Bioremediasi dapat diaplikasikan pada lingkungan-lingkungan yang terpolusi melalui berbagai mekanisme. Litchfield (1991), bioremediasi dilakukan melalui lima pendekatan berikut: bioreaktor, perlakuan fase padat, pengomposan, landfarming, dan perlakuan in situ. Berbagai proses teknologi telah berkembang di masing-masing bidang.
FAKTOR YANG MEMPENGARUHI PROSES DEGRADASI POLUTAN
Masalah utama yang sering dijumpai dalam aplikasi mikroorganisme untuk bioremediasi adalah menurun atau hilangnya potensi mikroba. Walaupun dalam percobaan laboratorium mikroba menunjukkan aktivitas degradasi yang tinggi, ternyata tidak menunjukkan hasil yang menggembirakan dalam percobaan di lapangan (in situ).
Untuk meningkatkan keefektifan penggunaan mikroorganisme dalam bioremediasi dapat dilakukan dengan melakukan dua strategi berikut. Pertama; yang disebut sebagai biostimulation yaitu suatu teknik menambahkan nutrien tertentu dengan tujuan merangsang aktivitas
mikroba-mikroba tempatan (indigenous). Atlas and Berta (1992), teknik biostimulasi ini telah sukses dalam mengendalikan tumpahan minyak di perairan dan kontaminasi senyawa hidrokarbon (PAH) di tanah. Lieberg and Cutright (1999), nutrien yang sering ditambahkan adalah fosfor dan nitrogen. Strategi kedua; yang disebut sebagai bioaugmentasi, yaitu dengan mengintroduksi mikroba tertentu pada daerah yang akan diremediasi. Dalam beberapa hal, teknik bioaugmentasi juga diikuti dengan penambahan nutrien tertentu.
Di samping masalah di atas, lambatnya kecepatan degradasi polutan di lingkungan disebabkan oleh beberapa faktor sebagai berikut: enzim-enzim degradatif yang dihasilkan oleh mikroba tidak mampu mengkatalis reaksi degradasi polutan yang tidak alami, kelarutan polutan dalam air sangat rendah, dan polutan terikat kuat dengan partikel-partikel organik atau partikel tanah. Selain itu, pengaruh lingkungan seperti pH, temperatur, dan kelembapan tanah juga sangat berperan dalam menentukan kesuksesan proses bioremediasi.
Oleh karena itu, seleksi, baik yang dilakukan secara konvensional maupun melalui manipulasi genetika untuk mendapatkan mikroba-mikroba yang potensial, merupakan agenda yang cukup penting dalam mikrobiologi lingkungan. Di samping itu, proses degradasi komplit di lingkungan umumnya dilakukan oleh konsorsium mikroorganisme bukan oleh mikroorganisme sejenis.
PENUTUP
Indonesia yang beriklim tropis memiliki bioma hutan yang lebih padat dari bioma daerah subtropis. Diperkirakan terdapat tiga kali lebih banyak mikroba pada ekosistem tropis daripada di ekosistem lainnya, di mana sebagian besarnya adalah sebagai pengurai (dekomposer). Ini merupakan suatu anugerah bagi kita, karena kita memiliki kesempatan yang lebih besar untuk mempelajari dan menelitinya untuk mendapatkan mikroba-mikroba yang potensial, yang selanjutnya dapat dikembangkan baik untuk tujuan pelestarian lingkungan maupun untuk kesejahteraan manusia.
Bapak-bapak dan hadirin yang terhormat, karena luasnya kajian bioremediasi di lingkungan dan terbatasnya waktu, demikianlah yang dapat saya sampaikan dalam kesempatan ini.
UCAPAN TERIMA KASIH
Bapak Rektor, Bapak Dekan, Anggota Senat Akademik, Anggota Dewan Guru Besar, dan hadirin yang saya hormati.
Sebelum saya mengakhiri pidato pengukuhan ini, izinkanlah saya mengucapkan syukur dan puji ke hadirat Allah SWT atas rahmat dan hidayah yang telah diberikan-Nya, sehingga pada hari ini saya dapat dikukuhkan sebagai guru besar. Semoga Allah selalu memberikan kekuatan kepada saya untuk mengemban dan melaksanakan tugas guru besar dengan sebaik-baiknya.
Ucapan terima kasih disampaikan kepada Bapak Menteri Pendidikan Nasional atas kepercayaannya kepada saya untuk memangku jabatan Guru Besar Tetap dalam bidang Mikrobiologi pada Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara.
Ucapan terima kasih yang tulus saya sampaikan kepada Rektor Universitas Sumatera Utara, Bapak Prof. Chairuddin P. Lubis, DTM&H, SpA(K) yang telah memberikan semangat dan dorongan kepada saya untuk menjadi guru besar. Selanjutnya saya mengucapkan terima kasih kepada seluruh Pembantu Rektor dan mantan Pembantu Rektor, Tim Penilai Kenaikan Pangkat Universitas Sumatera Utara, dan Dewan Guru Besar yang telah memberikan dukungan dan persetujuan pengusulan saya sebagai guru besar.
Kepada Bapak Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Sumatera Utara, Dr. Eddi Marlianto, M.Sc. dan para Pembantu Dekan, para Ketua dan Sekretaris Departemen di lingkungan Fakultas Matematika dan Ilmu Pengetahuan Alam (FMIPA) USU, Ketua Departemen Biologi Bapak Dr. Dwi Suryanto, M.Sc. dan seluruh Dosen Departemen Biologi dan para Dosen departemen lainnya, serta seluruh tenaga administrasi Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Sumatera Utara, saya mengucapkan terima kasih yang sebesar-besarnya atas dukungan yang telah diberikan selama ini. Ucapan terima kasih juga saya sampaikan kepada Direktur dan para Asisten Direktur Sekolah Pascasarjana Universitas Sumatera Utara, Ketua dan Sekretaris Program Studi Pengelolaan Sumberdaya Alam dan Lingkungan Sekolah Pascasarjana Universitas Sumatera Utara.
Selanjutnya, saya ingin menyampaikan terima kasih yang tulus kepada Bapak Prof. Dr. Herman Mawengkang yang telah banyak memberikan
dorongan dan pelajaran kepada saya. Kemudian kepada Bapak Dr. Ir. A. Faiz Albar, M.Sc. yang banyak dan selalu memberikan nasihat-nasihat, saya mengucapkan terima kasih yang tulus dan sedalam-dalamnya. Demikian pula kepada kakanda Bapak dr. Chairul Yoel, Sp.A.(K) dan kakanda Bapak Prof. drg. Ismet Danial Nasution, Sp. Pros., Ph.D. yang banyak memberikan dorongan dan semangat kepada saya. Tidak lupa kepada Bapak drg. Saidina Hamzah Dalimunthe, Sp.Prio. atas bantuannya dalam pengusulan guru besar saya, saya mengucapkan terima kasih yang sebesar-besarnya.
Ucapan terima kasih yang tulus saya sampaikan kepada guru-guru yang telah mengajar saya dari guru mengaji, guru sekolah dasar sampai dosen-dosen tingkat doktoral. Terutama sekali kepada pembimbing-pembimbing penelitian saya, Prof. Jasmi Jusfah, M.S. di Laboratorium Mikrobiologi, Jurusan Biologi Universitas Andalas Padang, Prof. John P. Davidson dan Prof. Jiliang Chiu di Laboratory of Microbiology, Department of Biology, Tuskegee University, Alabama, USA, Prof. Ibrahim bin Chee Omar dan Prof. Darah Ibrahim di Laboratory of Enzyme and Fermentation Technology, Sains Kaji Hayat, Universiti Sains Malaysia, dan Prof. Mikio Shimada dan Dr. Takefumi Hattori di Laboratory of Biochemical Control, Department of Applied Life Science, Kyoto University, Kyoto, Japan, saya mengucapkan terima kasih yang sedalam-dalamnya atas bantuan dan bimbingan yang telah diberikan kepada saya. Kepada rekan-rekan dan kolega yang satu per satu tidak mungkin disebutkan di sini, saya mengucapkan terima kasih yang tulus atas dukungan dan semangat yang telah diberikan.
Selanjutnya saya mengucapkan terima kasih yang sebesar-besarnya kepada USAID (USA) dan MONBUSHO (Japan) yang telah memberikan beasiswa kepada saya selama belajar di Amerika Serikat dan di Jepang. Juga kepada JSPS (Japan Society for the Promotion of Science) yang memberikan bantuan-bantuan penelitian kepada saya. Tidak lupa kepada Yayasan Supersemar RI yang telah memberikan bantuan pendidikan dari sekolah menengah pertama sampai ke perguruan tinggi, saya mengucapkan terima kasih yang sebesar-besarnya.
Kepada kedua orang tua, almarhum ayahanda Munir dan almarhumah ibunda Elma, ananda mengucapkan terima kasih yang sedalam-dalamnya. Pengorbanan yang Ayah dan Ibu berikan kepada kami tidak ternilai harganya. Walau Ayah dan Ibu tidak dapat bersama kami pada kesempatan ini, semangat Ayah dan Ibu tetap bersama kami. Semoga Allah SWT memberikan tempat yang sebaik-baiknya di sisi-Nya, amin ya Rabbal ’Alamin.
Pada kesempatan ini, saya menyampaikan terima kasih yang dalam kepada kedua mertua, Bapak Drs. H. Jusran R.C. dan Ibu Dra. Hj. Murni Abdullah yang selalu memberikan pelajaran, dukungan, dorongan, dan nasihat kepada kami sekeluarga. Begitu juga kepada kakak-kakak dan adik yang selalu mendorong dan memberikan doa, serta kakak-kakak ipar yang selalu membantu kami sekeluarga, kami mengucapkan terima kasih yang tak terhingga.
Kepada istri tercinta Dra. Rossie Yusran yang selama ini telah siap sedia mendampingi, sulit rasanya menemukan kata-kata untuk menyampaikan rasa terima kasih dan penghargaan atas pengorbanan, perhatian, kesetiaan, dan doamu yang tulus ikhlas. Kepada ananda tersayang Mutia Chairani dan Muhammad Fadli, ayah menyampaikan terima kasih atas kesabaran dan perhatian yang kalian berikan. Ayah sadar, selama ini waktu kalian untuk bermain dengan ayah di rumah tidak begitu banyak, namun ini semua adalah untuk kalian. Mudah-mudahan kesempatan ini akan memberikan semangat bagi kalian untuk mencapai cita-cita, semoga kalian menjadi anak-anak yang beriman, bertakwa, dan saleh, amin ya Rabbal ’Alamin.
Akhirnya, kepada seluruh sivitas akademika Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Sumatera Utara dan kepada panitia pengukuhan ini saya mengucapkan terima kasih. Kepada seluruh hadirin yang telah bersedia meluangkan waktu untuk mengikuti rangkaian acara ini, saya mohon maaf atas segala kekurangan.
Wabillahi taufik wal hidayah.
DAFTAR PUSTAKA
Atlas, R.M. and Berta, R. (1992). Hydrocarbon biodegradation and oil spill bioremediation, Adv. Microbial Ecol. 12: 287-338.
Barkovskii, A.I., Boullant, M.L. and Balandreau, J. (1994). Polyphenolic compounds respired by bacteria. In: Bioremediation through rhizosphere technology, ed. T.A. Anderson & J.L. Coats, American Chemical Society, Washington DC, pp. 29-42.
Baldrian, P. (2003). Interaction of heavy metals with white-rot fungi,
Enzyme and Microbial. Technol. 23: 79-91.
Barr, D.P. and Aust, D.A. (1994). Mechanisms of white rot fungi use to degrade pollutants, Environ. Sci. Technol. 28: 78-87.
Bollag, J.M., Mertz, T. and Otjen, L. (1994). Role of microorganism in soil bioremediation. In: In: Bioremediation through rhizosphere technology, ed. T.A. Anderson & J.L. Coats, American Chemical Society, Washington DC, pp. 4-10.
Cerniglia, C.E. and Sutherland, J.B. (2001). Bioremediation of polycyclic aromatic hydrocarbons by ligninolytic and non-ligninolytic fungi. In: Fungi in Bioremediation, ed. G.M. Gadd, Cambridge University Press, Cambridge, pp. 136-187.
Chanway, C.P. (1997). Inoculation of tree roots with plant growth promoting bacteria: An Emerging technology for reforestation, Forest
Science 43: 96-112.
Cooke, R.G. and Rayner, A.D.M. (1984). Ecology of Saprophytic Fungi. Longman, New York.
de Jong, E., de Vries, F.P., Field, J.A,. van de Zwan, P.P. and Bont, J.A.M. (1992). Isolation of basidiomycetes with high peroxides activity,
Mycological Research, 96: 1098-1104.
Dixon, R.K., Brown, S., Houghton, R.A., Solomoan, A.M., Trexler, M.C. and Wisniewshki, J. (1994). Carbon pool and flux of forest ecosystem,
Science 26: 185-190.
Donelly, P.K. and Fetcher, J.S. (1994). Potential use of mycorrhizal fungi as bioremediation agents. In: Bioremediation through rhizosphere technology. eds. T.A. Anderson & J.R. Coats, American Chemical Society, Washington. pp. 93-99.
Evans, C.S., Dutton, M.V., Guillen, F. and Veness, R.G. (1994). Enzymes and small molecular mass agents involved with lignocelluloses degradation, FEMS Mircobiol. Rev. 13: 325-240.
Filed, J.A., de Jong, E., Feijoo-Costa, G. and de Bont, J.A.M. (1993). Screening for ligninolytic fungi applicable to the biodegradation of xenobiotics, Trens in Biotechnol. 11: 44-49.
Fitz, W.J. and Wenzel, W.W. (2002). Arsenic transformations in the soil-rhizosphere-plant system: fundamentals and potential application to phytoremediation, Journal of Biotechnology 99: 259-278.
Gabriel J., Rychlovsky, P. and Krenzelok, M. (1995). Beyllium content in some wood-rotting fungi in Czech Republic, Toxicol. Envinron. Chem. 50: 233-236.
Green, F. and Highley, T.L. (1997). Mechanism of brown-rot decay: Paradigm or paradox, Int. Biodet. Biodegrad. 39: 113-124.
Harvey, P.J., Floris, R., Lundell, T., Palmer, J., Schoemarker, H.E. and Wever, R. (1992). Catalytic mechanisms and regulation of lignin peroksidase, Biochem. Society Transact. 20: 345-349.
Hedger, J.N. (1985). Tropical agarics: resource relations and fruiting periodicity. In: Development Biology of Higher Fungi, eds. D.Moore, L.A. Casselton, D.A. Wood and J.C. Frankland, Cambridge University Press, pp. 41-86.
Higuchi, T. (1980). Lignin structre and morphological distribution in plant cell wall. In: Lignin Biodegradation, Microbiology, Chemistry, and Potentian Application, Vol. I. ed. K. Kirk, T. Higuchi & H. Chang. CRC Press. Boca Raton, Florida, pp. 1-19.
Kartal S.N., Munir, E., Kakitani, T. and Imamura, Y. (2004). Bioremediation of CCA-treated wood by brown-rot fungi Fomitpsis palustris,
Coniophora puteana, and Laetiporus sulfurous, J. Wood Sci. 50:
182-188.
Levine, M.A. and Gealt, M.A. (1993). Biotreatment of Industrial and Hazardous Easte, McGraw Hill, New York. p.4.
Leyval, C., Turnau, K. and Haselwandter (1997). Effect of heavy metal pollution on mycorrhizal colonization and function: physiolgical, ecological and applied aspects, Mycorrhiza 7: 139-153.
Lieberg, E.W. and Cutright, T.J. (1999). The investigation of enhanced biormediation through the addition of macro and micro nutrients in PAHs contaminated soil, Inter. Biodet. Biodegrad. 44: 55-64.
Litchfield, C.D. (1991) Practices, potential and pitfall in the application of biotechnology to environmental problem. In: Environmental Biotechnology for Waste Treatment, ed. G. Saylor et. al., Plenum Press, New York, pp. 147-157.
Mueller, J.G., Cerniglia, C.E., Pritchard, P.H. (1998). Bioremediation of environments contaminated with polycyclic aromatic hydrocarbon. In: Bioremediation: Principles and Application, ed. R.L. Crawford & D.L. Crawford, Cambridge University Press, Cambridge, pp. 125-194. Munir, E., Hattori, T. and Shimada, M. (2005). Role of oxalate biosynthesis
for the growth of the copper tolerant wood-rotting fungi under environmental stress. The 55th Annual Meeting of the Japan Wood
Research Society.
Munir, E., Yoon, J.J., Tokimatsu, T., Hattori, T. and Shimada, M. (2001a). A physiological role of oxalic acid biosynthesis in the wood-rotting basidiomycete Fomitopsis palustris, Proc. Natl. Acad. Sci. USA. 98: 11126–11130.
Munir, E., Yoon, J.J., Tokimatsu, T., Hattori, T. and Shimada, M. (2001b). New role for glyoxylate cycle enzymes in wood-rotting basidiomycetes in relation to biosynthesis of oxalic acid, J. Wood Sci. 47: 368-373. Munir, E. and Goenadi, D.H. (1999). Bioconversion of oil palm trunk derived
lignocellulose to sugars. Menara Perkebunan 67: 37-44.
Peraturan Pemerintah RI Nomor 18 Tahun 1999, Tentang Pengelolaan Bahan Berbahaya dan Beracun.
Reddy, M.S., Axelrood, P.E., Radley, R. and Rennie, R.J. (1994). Evaluation of bacterial strains for pathogen suppression and enhancement of survival and growth of conifer seedlings. In: Improving plant productivity with rhizosphere bacteria, Proc. of 3rd Internat, PGPR.
Adelide, Australia, pp. 75-76.
Roane, T.M., Pepper, I.L. and Miller, R.M. (1998). Microbial remediation of metals. In: Bioremediation: Principles and Application, ed. R.L. Crawford & D.L. Crawford, pp. 312-340.
Rosenberg, E. and Ron, E.Z. (1998). Bioremediation of petrolium contamination. In: Bioremediation: Principles and Application, ed. R.L. Crawford & D.L. Crawford, Cambridge University Press, Cambridge. pp. 100-124.
Sayer, J. and Gadd, G.M. (1997). Solubilization and transformation of insoluble inorganic metal compounds to insoluble metal oxalates by
Sharples, J.M., Meharg, A.A., Chambers, S.M. and Cairney, J.W.G. (2000). Symbiotic solution to arsenic contamination, Nature 404: 951-952. Shimada, M., Akamatsu, Y., Ohta, A. and Takahashi, M. (1991). Biochemical
relationship between biodegradation of cellulose and formation of oxalic acid in brown-rot wood decay. Intern. Res. Group. On Wood Preserv. Doc. No. IRG/WP 1427, pp. 1-12.
Sieghardt, H. (1990). Heavy metal uptake and distribution in Silene vulgaris and Minuartia verna growing on mining-dump material containing lead and zinc, Plant and Soil 123: 107-111.
DAFTAR RIWAYAT HIDUP
A. DATA PRIBADI
Nama Lengkap : Prof. Dr. Erman Munir, M.Sc. NIP : 131945354
Jabatan : Guru Besar Fakultas MIPA Pangkat/Golongan : Pembina/IV/a
Alamat : Komplek Villa Malina Indah, Jl. Permata Raya No. 34 Tanjung Sari, Medan
Tempat/Tanggal Lahir : Padang Panjang/1 November 1965 Jenis Kelamin : Laki-laki
Agama : Islam Nama Orang Tua : Munir (Ayah)
Elma (Ibu)
Nama Istri : Dra. Rossie Yusran
Nama anak : 1. Mutia Chairani (siswi kelas 5 SDN Percobaan, Jl. Sei Petani, Medan)
2. Muhammad Fadli (6 tahun)
B. RIWAYAT PENDIDIKAN
1979 : Lulus SDN 1, Gunung, Padang Panjang 1982 : Lulus SMPN, Gunung, Padang Panjang 1985 : Lulus SMAN, Padang Panjang
1989 : Lulus S1, Biologi, FMIPA, Universitas Andalas, Padang
1995 : Lulus S2, Department of Biology, Faculty of Science, Tuskegee University, Alabama, USA
2002 : Lulus S3, Department of Applied Life Sciences, Agriculture, Kyoto University, Kyoto, Japan
C. PENDIDIKAN TAMBAHAN 1991 1992 1992 1998 : : : :
Kursus Bahasa Inggris di Universitas Sriwijaya, Palembang (7 bulan)
Kursus Bahasa Inggris di The British Institute, Bandung (4 bulan)
Kursus di Economics Institute, Colorado University, Boulder, USA (4 bulan)
Kursus Bahasa Jepang di Kyoto University, Kyoto, Japan (4 bulan)
D. RIWAYAT PEKERJAAN 1991 s.d. sekarang 1991 s.d. sekarang 1995-1997 1996-1997 : : : :
Dosen Fakultas MIPA USU
Staf Ahli Laboratorium Mikrobiolgi FMIPA USU Kepala Laboratorium Biologi Dasar LIDA USU Sekretaris Program Studi Biologi, FMIPA USU 2006 s.d. sekarang : Sekretaris Program Magister Pengelolaan
Sumberdaya Alam dan Lingkungan, Sekolah Pascasarjana USU
E. PUBLIKASI ILMIAH DAN PROSIDING
1. Munir, E., Yoon, J.J., Tokimatsu, T., Hattori, T. and Shimada, M. (2001). A physiological role of oxalic acid biosynthesis in the wood-rotting basidiomycete Fomitopsis palustris. Proc. Natl. Acad. Sci.
USA. 98 (20): 11126–11130.
2. Munir, E., Yoon, J.J., Tokimatsu, T., Hattori, T. and Shimada, M. (2001). New role for glyoxylate cycle enzymes in wood-rotting basidiomycetes in relation to biosynthesis of oxalic acid. J. Wood Sci. 47(5): 368-373.
3. Munir, E., Hattori, T. and Shimada, M. (2002). Purification and characterization of isocitrate lyase from the wood-destroying basidiomycete Fomitopsis palustris grown on glucose. Arch.
Biochem. Biophys. 399 (2): 225-231.
4. Munir, E., Hattori, T. and Shimada, M. (2002). Purification and characterization of malate synthase from the glucose-grown wood-rotting basidiomycete Fomitopsis palustris. Biosci. Biotech. Biochem. 66 (3): 576-581.
5. Munir, E., Hattori, T. and Shimada, M. (2001). A new concept of oxalic acid biosynthesis in physiology of copper-tolerant fungi. Intern. Res. Group on Wood Preserv. Doc. No. IRG/WP 01-10394 (Stockholm), pp. 1–8.
6. Munir, E., Hattori, T. and Shimada, M. (2003). A possible role of unique TCA cycle in wood-rotting basidiomycetes. Intern. Res. Group on Wood Preserv. Doc. No. IRG/WP 01-10394 (Stockholm). pp. 1-7.
7. Munir, E. and Goenadi, D.H. (1999). Bioconversion of oil palm trunk derived lignocellulose to sugars. Menara Perkebunan 67 (2): 37-44. 8. Munir, E. (2002). The importance of sulfhydryl compounds for the
9. Munir, E., Darah, I. and Omar, I. (1997). Solid-state fermentation processes for lignin degrading enzyme production by Phanerochate
chrysosporium on rice husks as substrates. Proceedings of the First
Colloquium on Lignocellulose at University Sains Malaysia.
10. Munir, E. (1995). Superkoiling DNA pada Salmonella thypimurium gyrB mutant. Prosiding Seminar Ilmiah LUSTRUM VI FMIPA USU, Medan, pp. 83-97.
11. Munir, E. (2002). Chromatographic profile of isocitrate lyase during purification. Jurnal Media Farmasi 10 (2): 162-173.
12. Munir E., Hattori, T. and Shimada, M. (2001). Distribution of isocitrate lyase and malate synthase among wood-rotting fungi. The Third International Wood Science Symposium, Wood Research Institute, Kyoto University, Kyoto. pp. 380–385.
13. Munir, E., Hattori, T. and Shimada, M. (2002). A new glucose metabolism in wood-rotting fungi. The Fourth International Wood Science Symposium. LIPI Jakarta. pp. 336-340.
14. Munir, E., Hattori, T. and Shimada, M. (2004). Profile of enzyme activity and growth of wood rotting fungi in metal ion containing media. The Fifth International Wood Science Symposium. Kyoto Japan. pp. 281-286.
15. Munir, E. and Shimada, M. (2005). An Inhibitory effect of acetate added to the culture on growth of wood rotting basidiomycetes. The sixth International Wood Science Symposium. Bali Indonesia. pp. 405-410.
16. Munir, E. (2004). Isositrat liase sebagai enzim target perancangan obat. Jurnal Media Farmasi. 12 (1): 13-21.
17. Munir, E. (2003). A new aspect of oxalic acid biosynthesis in biochemistry of basidiomycetes. Workshop and Expose of Fundamental Research Scientific Results of LIPI-JSPS Cooperation Program. pp. 81 – 87.
18. Munir, E. (2005). Journal Impact Factor sebagai standar mutu jurnal ilmiah. Perkasa: Periodikal Pascasarjana USU 9 (1): 35-37.
19. Yoon, J. J., Munir, E., Miyasou H., Hattori, T., Terashita, T. and Shimada, M. (2002). A possible role of the key enzymes of the glyoxylate and gluconeogenesis pathways for fruit body formation of the wood-rotting basidiomycete Flammulina velutipes.
Mycoscience. 43 (4): 327-332.
20. Darah, I., Omar, I. and Munir, E. (1997). The role of agitation on the production of lignin peroxidase and manganese peroxidase by
on Lignocellulose at University Sains Malaysia, Penang.
21. Shimada, M., Sakai, F., Kuroda, H., Hattori. T., Setiadi, Y., Primaturi, R., Munir, E., Hanafiah, A.S., Tengku, S., Goenadi, D.H., Taniwiriyon, D. and Widiastuti. (2001). Biochemical analysis of organic acid metabolism of symbiotic and saprophytic basidiomycetes occurring in forest ecosystems. In: Science for Sustainable Utilization of Forest Resources in the Tropics. Published by the International Committee of Academic Exchange, Wood Research Institute Kyoto University. pp. 62-65.
22. Shimada, M., Yoon, J.J., Munir, E., Hattori, T. (2002). Metabolic physiology of wood decay fungi: Copper tolerance, oxalic acid and biochemistry of wood decay. Wood Preservation 28 (3): 86-97. 23. Davidson, J.P., Chiu, J., Session, W., Suppiramaniam, V. and Munir, E.
(1995). The effect of temperature on DNA super coiling in a gyrB mutant in Salmonella tyhphimurium. Journal of Cellular
Biochemistry 19A, 103.
24. Nurwahyuni, I., Munir, E. dan Riyani, Y. (1996). Perbanyakan anggrek Dendrobium sp. Secara kultur jaringan. Jurnal Komunikasi
Penelitian USU 8 (4): 330-336.
25. Agustien, A. dan Munir, E. (1997). Purifikasi penisilinasilase dari
Bacillus. Prosiding seminar PPI Malaysia. Penang. pp. 270-277.
26. Kuwahara, M., Shimada, M., Watanabe, T., Honda, Y., Kondo, R., Prasetya, B., Basuki, T., Idiyanti, T., Goenadi, D.H., Away, Y., Pasaribu, R.A., Munir, E. and Darma, IGKT. (2001). Production of pulp and paper by using biological methods from tropical wood resources. In: Science for Sustainable Utilization of Forest Resources in the Tropics. Published by the International Committee of Academic Exchange, Wood Research Institute Kyoto University. pp. 29-33.
27. Kartal S.N., Munir, E., Kakitani, T. and Imamura, Y. (2004). Bioremediation of CCA-treated wood by brown-rot fungi Fomitpsis
palustris, Coniophora puteana, and Laetiporus sulfurous. J. Wood Sci. 50 (1): 182-188.
28. Munir, E., Yoon. J.J., Tokimatsu, T., Hattori, T. and Shimada, M. (2000). Novel occurrence of the glyoxylate cycle enzymes in wood-rotting basidiomycetes growing in glucose rich-media. 50th Annual
Meeting of the Japan Wood Research Society at Kyoto University, Kyoto, Japan.
29. Munir, E., Yoon. J.J., Tokimatsu, T., Hattori, T. and Shimada, M. (2002). A new role of glyoxylate cycle in wood-rotting
basidiomycetes in relation to the biosynthesis of oxalic acid. 50th
Annual Meeting of the Japan Wood Research Society at Kyoto University, Kyoto, Japan.
30. Munir, E., Yoon. J.J., Tokimatsu, T., Hattori, T. and Shimada, M. (2002). Purification and characterization of isocitrate lyase from a wood-rotting fungus Fomitopsis palustris. 51st Annual Meeting of the
Japan Wood Research Society at the University of Tokyo, Tokyo, Japan.
31. Munir, E., Hattori, T. and Shimada, M. (2002). Purification and characterization of malate synthase from the brown-rot fungus
Fomitopsis palustris. 52nd Annual Meeting of the Japan Wood
Research Society at Gifu University, Gifu, Japan.
32. Munir, E., Yoon. J.J,, Tokimatsu, T., Hattori, T. and Shimada, M. (2001). Purification and characterization of isocitrate lyase from a wood-rotting basidiomycete Fomitopsis palustris grown in a glucose-rich medium. Annual Meeting of Japan Society for Bioscience, Biotechnology and Agrochemistry. Ritsumeikan University, Kyoto, Japan.
33. Munir, E., Hattori, T. and Shimada, M. (2003). Purification and characterization of malate synthase from the oxalic acid-producing basidiomycete Fomitopsis palustris. The 423rd Kansai Regional
Meeting of the Japan Society for Bioscience, Biotechnology, and Agrochemistry, Kyoto, Japan.
34. Munir, E. (2002). Oxalate fermentation by wood-rotting fungi vs. normal respiration. Semirata bidang MIPA BKS-PTN Wilayah Barat Indonesia XV, Medan.
35. Munir, E. (2002). Chromatographic profile of isocitrate lyase during purification. Seminar Nasional Biologi XVII Perhimpunan Biologi Indonesia, Padang.
36. Munir, E. (2003). A possible role of glyoxysomal enzymes for the growth of wood-rotting basidiomycetes metal ion containing media. Seminar Nasional Kimia, Medan.
37. Munir, E. (2003). Peranan Asam Oksalat dalam Sitem Biologi dan Lingkungan. Orasi Ilmiah. Dibacakan pada DIES Natalis USU ke-51, Medan.
38. Munir, E., Hattori, T. and Shimada, M. (2005). Role of oxalate biosynthesis for the growth of the copper tolerant wood-rotting fungi under environmental stress. The 55th Annual Meeting of the
39. Munir, E. (2005). Peranan asam oksalat dalam degradasi lignoselulosa. Seminar Nasional Kimia II, Medan.
40. Yoon. J.J., Munir, E., Hattori, T. and Shimada, M. (2002). A physiological role of glyoxylate and TCA cycles in fruit body formation of the wood-rotting fungus Fomitopsis palustris. 46th
Annual Meeting of the Mycological Society of Japan at Shinsu University, Shinsu, Japan.
41. Yoon, J.J., Munir, E., Tokimatsu, T., Hattori, T. and Shimada, M. (2000). A possible role of glyoxylate cycle in fruit body morphogenesis of edible mushrooms. 44th Annual Meeting of the
Mycological Society of Japan at Kinki University, Nara, Japan.
42. Yoon, J.J., Miyaso, H., Munir, E., Hattori, T., Terashita, T. and. Shimada, M. (2001). Carbon metabolism during growth and development of mycelium and fruit body in the white-rot fungus Flamulina velutipes. 45th Annual Meeting of the Mycological Society
of Japan, Tokyo, Japan.
43. Shimada, M., Hattori, T., Munir, E., Yoon, J.J., Tokimatsu, T. and T. Nishide. (2001). A new mechanism for carbon metabolism coupling with oxalate biosynthesis in wood-rotting fungi. The 1st Conference
on Fungal Genetics and Molecular Biology. Tokyo University, Tokyo, Japan.
44. Nagai, Y., Munir, E., Hattori, T., Tokimatsu, T. and Shimada, M. (1999). Intramolecular electron transfer in glyoxylate dehydrogenase from brown-rot fungus Tyromyces palustris. 49th
Annual Meeting of the Japan Wood Research Society at Tokyo University of Agriculture, Tokyo, Japan.
45. Kartal, N.S., Munir, E. and Imamura, Y. (2003). Removal of copper, chromium, and arsenic from CCA-tretaed wood by bioremediation with brown-rot fungi. The 53rd Annual Meeting of the Japan Wood
Research Society. Sizuoka, Japan.
46. Nurtjahja, K., Munir, E. and Nugroho, R.P. (2004). The distribution of tropical mushrooms in Sibolangit conservation forest, North Sumatra, Indonesia. The Fifth International Wood Science Symposium, Kyoto, Japan.
47. Sakai, S., Nishide, T., Munir, E., Hattori. T., Baba, K., Inui, H., Nakano, Y. and Shimada, M. (2004). Subcellular localization of isocitrate lyase in the wood rotting destroying basidiomycetes
Fomitopsis palustris. The Fifth International Wood Science
48. Shimada M, Hattori T, Munir, E., Yoon J.J., Nishide T., Watanabe T. and Sakai, S. (2004). A novel physiological aspect of wood-destroying Basidiomycetes. Proceedings of the 4th Regional
coordination of wood science international symposium, pp 467-468, St. Petersburg, Rusia.
49. Suryanto, D., Ginting, D. Munir, E. dan Yurnaliza (2005). Isolasi bakteri kitinolitik dan uji aktivitas kitinolitik kasar beberapa bakteri tanah. Seminar Nasional ke XVII dan Kongres X Perhimpunan Biokimia & Biologi Molekuler Indonesia, Pekan Baru.
50. Suryanto, D., Kelana, T.B., Munir, E. dan Nani, N. (2005). Uji brine-shrimp dan pengaruh ekstrak metanol daun tumbuhan pradep
Psychothria stipulacea Wall (Familia: Rubiaceae) terhadap mikroba.
Simposium Nasional Bahan Alam, Bogor.
F. PEMAKALAH PADA BEBERAPA SEMINAR
1. The 55th Annual Meeting of the Japan Wood Research Society at
Kyoto University, Kyoto, 2005. 2. Seminar Nasional Kimia II, 2005.
3. The Fifth International Wood Science Symposium, Kyoto Japan, 2004.
4. Seminar Hasil Penelitian Program PPD HEDS, Medan, 2003. 5. Seminar Nasional Kimia, Medan, 2003.
6. Orasi Ilmiah dibacakan dalam DIES Natalis USU ke-51, Medan, 2003.
7. Workshop and Expose on Fundamental Research Scientific Results of Indonesia – Japan Cooperation Program, LIPI Jakarata, 2003. 8. The fourth International Wood Science Symposium, LIPI Jakarta,
2002.
9. The 423rd Kansai Regional Meeting of the Japan Society for
Bioscience, Biotechnology, and Agrochemistry. Kyoto, Japan, 2002. 10. Seminar Nasional Biologi XVII Perhimpunan Biologi Indonesia,
Padang, 2002.
11. Semirata bidang MIPA BKS-PTN Wilayah Barat Indonesia XV, Medan, 2002.
12. 44th Annual Meeting of the Mycological Society of Japan at Kinki
University, Nara, Japan, 2000.
13. Annual Meeting of Japan Society for Bioscience, Biotechnology and Agrochemistry. Ritsumeikan University, Kyoto, 2001.
Preserv), Nara Jepang, 2001.
15. The 50th Annual Meeting of the Japan Wood Research Society at
Kyoto University, Kyoto, Japan, 2000.
16. The third International Wood Science Symposium, Wood Research Institute, Kyoto University, Kyoto, 2000.
17.
First Colloquium on Lignocellulose at University Sains Malaysia,Penang, Malaysia, 1997.
G. SEBAGAI PESERTA BEBERAPA SEMINAR/WORKSHOP
1. Seminar Kerjasama Kemitraan Riset Pemerintah dengan Swasta, BPPT Jakarta, 2005.
2. Perkembangan Teknologi Informasi di Dunia Pendidikan, Microsoft Indonesia, Medan, 2004.
3. Perkembangan Terkini Ilmu Kimia dan Pembelajarannya, Medan, 2004. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18.
International Seminar on Sustainable Utilization of Acacia Mangium, Kyoto, Japan, 2003.
The second symposium on Chemical Biology of Metal Sensors with Switching Functions, Kyoto, Japan, 2002.
Seminar Peran Biokimia Asam Oksalat Hasil Simbiosis Cendawan Basidiomycetes dalam Ekologi dan Konservasi Hutan, Medan, 2000. Seminar PPI Jepang, Hamamatsu, Jepang, 2000.
Seminar PPI Jepang, Kyoto, Japan, 1999.
The 49th Annual Meeting of the Japan Wood Research Society at
Tokyo University of Agriculture, Tokyo, Japan, 1999.
The 17th symposia on the Life Science, Kyoto, Japan, 1998.
Lokakarya BKS FMIPA Wilayah Barat, Universitas Palangkaraya, Palangkaraya, 1996.
Seminar IMT-GT, Penang, Malaysia, 1996. Seminar Evaluasi HEDS Project, Batam, 1996.
Seminar Manajemen Sumber Daya Air, Puslit SDAL-USU, Medan, 1995.
Seminar Microbial Ecology and Biotechnology and Course Development, USU, Medan, 1995.
Seminar Citizens Working to Protect Our Environment, Puslit SDAL-USU, Medan, 1995.
Keystone Symposia on Bacterial chromosome, Santa Fe, New Mexico, USA, 1995.
H. KEGIATAN PENELITIAN Ketua 1997 1997 1998 1997 s.d. 1998 1999 s.d. 2002 2002 Anggota 1997 s.d. 1998 1997 s.d. 1999 2000 s.d. 2002 2003 s.d. 2004 2003 s.d. 2004 2005 2005 s.d. 2006
Isolasi kristal protein Bacillus thuringensis untuk kajian bioinsektisida. (Penelitian OPF USU)
Isolasi enzim pendegradasi lignin dari Phanerochaete
Chrysosporium. (Penelitian PPD DIKTI)
Optimasi produksi enzim ligninolitik oleh
Phanerochaete chrysosporium dan aktivitasnya
dalam Mendegradasi limbah lignoselulosa. (Penelitian BBI DIKTI)
Biokonversi lignoselulosa batang kelapa sawit untuk menghasilkan gula. (Dana Penelitian USM Malaysia) Metabolisme asam oksalat pada jamur-jamur pembusuk kayu. (Dana JSPS, Kyoto University)
Kemampuan jamur kayu dalam detoksifikasi ion-ion logam (JSPS-Japan)
Pembuatan poliblem mampu terdegradasi menggunakan teknik reaktif polyolefin dengan serat limbah kelapa sawit. (Hibah Bersaing DIKTI)
Production of pulp and paper by using biological methods from tropical wood resources. (Penelitian kerja sama LIPI Indonesia dengan JSPS Jepang) Biochemical analysis of organic acid metabolism of symbiotic and saprophytic basidiomycetes occurring in forest ecosystem. (Penelitian kerja sama LIPI Indonesia dengan JSPS Jepang)
Localization of oxalate synthesizing enzymes and cDNA cloning of the key enzyme for oxalate biosynthesis. (JSPS-Japan)
Sustainable production and utilization of tropical forest resources for establishment of a recycling based-society. (Penelitian kerja sama LIPI Indonesia dengan JSPS Jepang)
Isolasi dan uji aktivitas antimikroba beberapa tumbuhan asal Cagar Alam Tangkahan Sumatera Utara yang berpotensi sebagai obat. (Proyek SP4) Eksplorasi bakteri kitinolitik: Keragaman genetik gen penyandi kitinase pada berbagai jenis bakteri dan kemungkinan pemanfaatannya. (Hibah Bersaing DIKTI)
2005 s.d. 2006 Screening of phosphate solubilizing basidiomycetes from tropical forest. (Penelitian kerja sama dengan Kyoto University)
I. ANGGOTA ORGANISASI PROFESI
1994 s.d. 1995 American Society for Microbiology 1994 s.d. 1995 National Geographic of the USA 1995 s.d. 1996 American Phytopathological Society 1998 s.d. sekarang Japan Mycological Society
1998 s.d. sekarang Core University Program antara LIPI (Indonesia) – JSPS (Japan)
1999 s.d. sekarang Japan Wood Research Society
2000-2004 Japan Society for Bioscience, Biotechnology, and Agrochemistry
2002 s.d. sekarang Perhimpunan Mikrobiologi Indonesia 2004 s.d. sekarang Masyarakat Peneliti Kayu Indonesia
J. PENGHARGAAN
1996
2003
1. Dosen Teladan I FMIPA USU 2. Dosen Teladan III USU
Peneliti terbaik kerjasama LIPI dengan JSPS Jepang di bidang Wood Sciences