6 BAB II KAJIAN PUSTAKA
2.1 Penyakit pada Kacang Panjang yang Disebabkan oleh Virus
Virus merupakan patogen yang potensial menurunkan hasil tanaman kacang panjang di daerah Asia, Amerika Latin, dan Afrika, salah satu virus yang menginfeksi kacang panjang adalah mosaik dan kuning pada kacang panjang. Virus yang menunjukkan gejala mosaik berasosiasi dengan tiga jenis virus yaitu Bean common mosaic (BCMV), Tobaco mosaic virus (TMV) dan Cucumber mosaic virus (CMV), sedangkan gejala kuning pada tanaman kacang panjang dilaporkan diinfeksi oleh Mungbean yellow mosaic india virus (MYMIV) (Damayanti et al. 2009).
2.1.1 Bean common mosaic virus ( BCMV)
BCMV merupakan salah satu virus anggota famili Potyviridae, genus Potyvirus dengan genom ssRNA (utas tunggal), positive sense, berbentuk filament dengan panjang 750 nm dan lebar 14 nm. Badan inklusi Potyvirus berbentuk cakra atau beberapa bentuk yang lain (Regenmortel et al. 2004).
Tipe gejala penyakit yang muncul pada pertanaman bergantung pada strain BCMV, temperatur, dan genotipe inang (Udayashankar et al. 2010). Gejala pertama kali terlihat pada daun-daun muda berupa pemucatan tulang daun yang mengakibatkan jaringan sekitarnya menjadi hijau muda, kemudian berkembang menjadi mosaik dengan pola warna hijau dan kuning disertai malformasi. Tulang daun akan mengerut sehingga daun terlihat bergelombang dan permukaan daun menjadi tidak rata. Gejala lanjut menunjukkan lepuhan-lepuhan sehingga bentuk
daun tidak teratur (pengurangan ukuran lamina daun), layu dan akhirnya gugur (Setyastuti, 2008). Menurut Mukeshimana et al. (2003), tanaman yang terserang BCMV memiliki daun yang menggulung, keriting, tanaman menjadi kerdil, dan polong serta biji yang dihasilkan lebih sedikit dibandingkan dengan tanaman sehat. Polong kacang panjang yang terserang BCMV menunjukkan gejala mosaik dan malformasi polong (Sutic et al.1999). BCMV bersifat terbawa benih dan dapat ditularkan secara mekanik oleh sap tanaman dan melaui alat-alat pertanian. Virus ini ditularkan oleh beberapa jenis kutudaun termasuk Myzus persicae, Aphis craccivora, A.fabae, A.gossypii dan A.medicaginis secara non persisten (Shukla et al. 1994).
2.1.2 Tobaco mosaic virus (TMV)
TMV (Tobacco mosaic virus) termasuk ke dalam genus Tobamovirus. TMV merupakan salah satu dari 14 spesies yang termasuk dalam genus Tobamovirus. TMV memiliki ciri berbentuk batang dengan panjang 300 nm dan diameter 15 nm. Proteinnya terdiri atas kira-kira 2130 protein subunit, dan setiap subunitnya terdiri 158 asam amino (Garry, 2002). Protein subunitnya tersusun pada sebuah helix. Asam nukleat TMV berbentuk untai tunggal RNA dan terdiri atas kurang lebih 6400 nukleotida. Untai RNA juga berbentuk helix sejajar dengan untai protein. Berat dari setiap partikel virus antara 3,9 x 107 dan 4 x 107 unit berat molekul.
TMV merupakan virus yang menyerang tanaman dan dapat menginfeksi lebih dari 35 spesies tanaman sehingga dapat menyebabkan kerugian yang besar pada tanaman tembakau. TMV dapat memperbanyak diri jika berada pada sel hidup, tapi
virus ini dapat tetap bertahan hidup pada fase dorman dan jaringan tanaman yang mati selama bertahun-tahun maupun di luar tanaman baik itu di dalam tanah, di permukaan tanah maupun pada peralatan yang telah terkontaminasi virus ini. TMV menyebar secara mekanis tetapi serangga seperti aphids tidak dapat menjadi vektor bagi virus ini (Garry, 2002). Tanaman yang terinfeksi TMV menunjukkan gejala yaitu : daun-daun muda berubah menjadi warna belang kuning hijau, keriting serta berkerut, tanaman kerdil, buah belang dan berwarna kuning. Gejala lain yang terlihat adalah munculnya garis nekrosis pada daun yang menyebabkan terjadinya gugur daun (Widodo dan Wiyono, 1995).
2.1.3 Cucumber mosaic virus (CMV)
CMV (Cucumber Mosaic Virus) termasuk dalam kelompok Cucumovirus, bersama-sama dengan Peanut stunt virus (PStV) dan Cabaio aspermy virus (CAV) (Palukaitis et al.1997). CMV mempunyai tiga RNA genom beruntai tunggal (RNA 1, 2, 3), satu RNA subgenom (RNA 4). Masing-masing RNA ini mempunyai fungsi genomik yang berbeda (Kaper and Waterwoth, 2001). Virus ini mempunyai kisaran inang terluas di antara virus tanaman yang diketahui saat ini, dilaporkan dapat menginfeksi lebih dari 800 spesies tumbuhan, dapat menyebabkan kerugian besar pada berbagai jenis tanaman (Palukaitis et al. 1997).
Lebih dari 60 isolat CMV sudah diketahui sifat-sifatnya (Kaper and Waterwoth, 2001). CMV terdapat hampir di semua Negara, dengan strain dan sifat biologinya yang berbeda-beda. Kisaran inang dari CMV yang luas menyebabkan
gejala yang ditimbulkannya pun beragam. CMV mempunyai kisaran inang yaitu : terdapat pada tanaman sayuran, tanaman hias dan tanaman buah-buahan. CMV juga menyerang tanaman melon, labu, cabai, bayam, tomat, mentimun, seledri, bit, polong-polongan, pisang, tanaman famili crucifereae, delphinium, gladiol, lili, petunia, tulip, zinia, dan beberapa jenis gulma (Agrios, 2005).
Gejala infeksi yang diakibatkan oleh virus ini adalah, mula-mula tampak pada sebagian tulang daun menguning atau terjadinya jalur kuning sepanjang tulang daun. Daun berubah warna menjadi belang hijau muda dan hijau tua, serta daun menjadi kecil dan menyempit. Jika tanaman terinfeksi pada waktu masih muda tanaman akan terhambat pertumbuhannya dan menjadi kerdil. Tanaman yang sakit menghasilkan buah yang kecil dan sering tampak berjerawat (Semangun, 2000). Virus ini dapat menyebabkan penurunan hasil sebesar 30-60%, bahkan jika infeksi terjadi pada fase bibit dapat menyebabkan kerusakan sampai 100% (Duriat, 1996)
2.1.4 Mungbean yellow mosaic india virus (MYMIV)
Mungbean yellow mosaic india virus (MYMIV) merupakan famili
Geminiviridae, genus Begomovirus, yang sering menyerang tanaman budidaya dan
memiliki pasangan dua partikel virus isometrik dalam suatu susunan monogemini, bigemini, dan hibrigemini. Tiap partikel berukuran 28 - 30 nm dan tiap pasangan virus mengandung ss-DNA sirkuler tertutup, dengan ukuran antara 2500 - 3000 bp. Beberapa anggota Begomovirus mengandung genom yang berukuran sama, terpisah menjadi dua molekul DNA, tetapi urutan nukleotidanya tidak sama (Wahyuni, 2005).
Gejala yang ditimbulkan oleh MYMIV berbeda-beda, tergantung pada genus dan spesies tanaman yang terinfeksi. Gejala pada umumnya muncul pada daun muda atau pucuk berupa bercak kuning di sekitar tulang daun, kemudian berkembang menjadi urat daun berwarna kuning (vein clearing), cekung dan mengkerut dengan warna mosaik ringan atau kuning. Gejala berlanjut hingga hampir seluruh daun muda atau pucuk berwarna kuning cerah, dan ada pula yang berwarna kuning bercampur dengan hijau, daun cekung dan mengkerut berukuran lebih kecil dan lebih tebal (Wahyuni, 2005).
Virus ini ditularkan oleh kutu kebul (Bemisia tabaci). Penularan oleh serangga vektor B. tabaci sangat dipengaruhi oleh lamanya masa akuisisi serangga pada tanaman sakit, jumlah serangga dan lamanya periode inokulasi yang terjadi pada tanaman sehat. Vektor B. tabaci menularkan MYMIV secara persisten (tetap) artinya sekali B. tabaci makan tanaman yang mengandung MYMIV, maka selama hidupnya dapat menularkan virus. Periode makan akuisisi (makan tanaman sakit untuk memperoleh virus) selama 48 jam dapat menghasilkan tingkat penularan yang paling efisien (Gunaeni dkk. 2008).
2.2 Begomovirus Penyebab Penyakit Kuning
Begomovirus merupakan salah satu patogen penting yang menginfeksi
beberapa komoditas hortikultura utama di Negara tropis dan sub-tropis. Penularan Begomovirus di lapangan sebagian besar melalui vektornya Bemisia tabaci Gen.
ditularkan baik secara mekanis maupun benih. Kisaran inang yang luas dan penyebaran melalui vektor membuat kejadian penyakit Begomovirus tinggi dan sulit dikendalikan.
Dilaporkan oleh Pratap et al. (2011) dan Green et al. (2003) bahwa tanaman terung di India dan Thailand terinfeksi oleh berturut-turut Tomato leaf curl virus (ToLCV) dan Tomato yellow leaf curl virus (TYLCV). Kedua virus tersebut termasuk anggota Begomovirus, famili Geminiviridae. Navas- Castillo, et al (1999) menyebutkan, terdapat pula beberapa spesies Begomovirus yang menyebabkan penyakit kuning pada tanaman hortikultura lainnya yaitu, African cassava mosaic virus (ACMV), Bean dwarf mosaic virus (BDMV), Bean golden mosaic virus
(BGMV), Abutilon mosaic virus (AbMV), Cotton leaf crumple virus (CLCV), Squash leaf curl virus (SLCV), Tomato golden mosaic virus (TGMV), Potato yellow mosaic
virus (PYMV), Mungbean yellow mosaic virus (MYMV), Euphorbia mosaic virus
(EuMV), dan Indian cassava mosaic virus (ICMV).
2.2.1 Karakter molekuler begomovirus
Begomovirus merupakan salah satu genus dari family Geminiviridae .
Geminiviridae ini merupakan kelompok kelompok virus patogen tumbuhan terbesar.
Nama Geminviridae berasal dari karakteristik morfologi partikel virus berupa isometrik kembar yang selalu berpasangan (twinned-geminate) dengan ukuran berkisar 20-30 nm. Secara genetik Geminiviridae memiliki genom berupa DNA utas tunggal (ssDNA) yang berbentuk sirkuler (Harrison dan Robinson 1999).
Harison and Robinson (1999) menjelaskan bahwa Begomovirus diklasifikasikan ke dalam famili Geminiviridae yang dibagi ke dalam tiga genus yaitu Mastrevirus, Curtovirus, dan Begomovirus yang didasarkan atas perbedaan kisaran
inang, serangga vektor dan genomnya. Mastrevirus adalah Begomovirus yang menginfeksi tanaman inang monokotil, ditularkan oleh serangga vektor wereng daun dan memiliki struktur genom monopartit. Curtovirus, menginfeksi tanaman dikotil dengan vektor dan struktur genom sama dengan genus pertama. Begomovirus, menginfeksi tanaman dikotil dan ditularkan oleh kutukebul (Bemisia tabaci) memiliki struktur genom monopartit atau bipartit dengan sifat penularan persisten, sirkulatif dan non-propagatif.. Menurut Van Regenmortel (2004) selain ketiga genus tersebut, ada satu genus lainnya yang termasuk ke dalam famili Geminiviridae yaitu Topocuvirus yang menginfeksi tanaman dikotil, ditularkan oleh wereng pohon dan
memiliki genom monopartit.
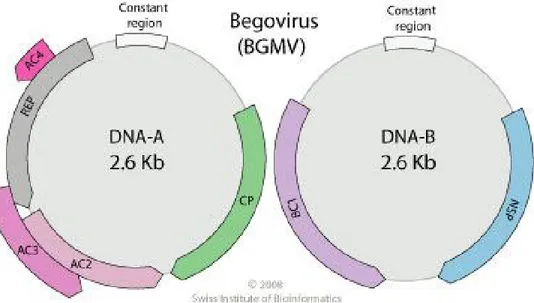
Diantara genus-genus tersebut di atas, Begomovirus merupakan genus dengan jumlah anggota terbesar. Genus Begomovirus terdiri dari virus-virus dengan genom bipartit atau monopartit. Sebagian besar anggota genus Begomovirus memiliki genom bipartit yang terdiri dari dua molekul DNA utas tunggal sirkuler yang berbeda yaitu DNA A dan DNA B dengan masing-masing berukuran 2,7-2,8 kb (Gambar 1). Begomovirus dengan genom monopartit, semua gennya terletak pada satu DNA utas
tunggal sirkuler yang berukuran 2,8 kb. Komponen DNA Begomovirus baik monopartit maupun bipartit mengandung gen-gen yang menyandikan protein dengan fungsi yang khusus (Tabel 1). Gen penyandi protein selubung virus merupakan
daerah genom yang mempunyai runutan DNA dengan derajat kesamaan yang tinggi antara anggota Begomovirus dalam satu genus (Rojas et al. 1997).
Gambar 2.2. Organisasi genom DNA-A dan DNA-B Begomovirus
Keterangan : DNA-A memiliki enam open reading frame (ORF), yaitu CP (gen AR1; protein selubung, CP) dan MP (gen AR2; protein untuk perpindahan virus, MP) pada salah satu untai; REP (gen AL1; protein replikasi, Rep); AC2 (gen AL2; protein activator transkripsi, TrAP); AC3 (gen AL3, peningkat replikasi, REn) dan AC4 (gen AL4; protein AC4) pada untai komplementer. DNA-B mengandung 2 protein pengkode ORF yang terlibat dalam perpindahan virus, yaitu NSP (gen BL1; protein selubung inti) pada salah satu untai dan BC1 (gen BL1; protein untuk perpindahan virus) pada untai komplementer (Fauquet et al. 2005; Seal et al. 2006).
Monopartit Bipartit Protein dan Fungsi
V1 AV1 Protein selubung virus (coat protein), berperan dalam penyebaran virus, pergerakan virus di dalam inangnya dan berperan dalam penularan yaitu melindungi partikel virus dari degradasi pada saat masuk sistem pencernaan kutukebul (Briddon et al.1989; Morin et al. 2000; Hull, 2002; Harrison & Robinson, 1999)
V2 AV2 virus dalam tanaman terinfeksi (Hull 2002; Harrison & Robinson 1999)
C1 AC1 Replication-associated protein (Rep), berperan
dalam proses replikasi virus (Desbiez et al. 1995; Hull, 2002)
C2 AC2 Transcriptional activator protein (TrAP), protein yang terlibat dalam pengaktifan transkripsi dari promoter protein selubung. Protein ini ditemukan pada inti dan berperan dalam patogenisitas virus (van Wezel et al. 2001)
C3 AC3 Replication enhancer protein (REn), protein ini berinteraksi dengan protein C1 dan meningkatkan akumulasi DNA virus (Hanleybowdoin et al. 2000) C4 AC4 Berinteraksi dengan C1 dan V2, berperan dalam
penentu gejala dan terlibat dalam inisiasi pembelahan sel (Krake et al. 1998), pergerakan DNA virus dari sel ke sel (Rojas et al. 2001), mematahkan mekanisme pertahanan tanaman (van Wezel et al. 2001),
- BV1 Nuclear shuttle protein (NSP) dan menyandikan virion DNA B (Hull, 2002; Salati, 2002)
- BC1 Movement protein (MP), berperan dalam pergerakan virus di dalam tanaman terinfeksi (Hull, 2002; Salati, 2002).
2.2.2 Gejala infeksi Begomovirus
Sumber inokulum Begomovirus berada pada tanaman inang, sisa-sisa tanaman dan inang alternatif. Secara alamiah Begomovirus dapat sampai ke tanaman atau berpindah dari satu tanaman ke tanaman lainnya apabila ada serangga vektor
Tabel 2.1.
kutukebul (B. tabaci Gen) karena virus ini tidak ditularkan melalui biji ataupun secara mekanik. Setelah virus sampai pada inang yang sesuai maka virus akan melepaskan selubung protein kemudian memanfaatkan DNA tanaman untuk bereplikasi kemudian berpindah dari satu sel ke sel lainnya mengikuti aliran nutrisi dan air tumbuhan sehingga gejala akan bersifat sistemik (Hull 2002).
Gejala yang ditimbulkan Begomovirus bervariasi tergantung pada strain virus, jenis tanaman, fase pertumbuhan tanaman dan beberapa faktor lainnya. Gejala infeksi virus berupa daun menggulung, penebalan tulang daun,bercak-bercak klorotik pada daun, klorosis di antara tulang daun, malformasi daun, belang dan menguning (Lotrakul et al. 2000). Menurut Sulandari (2006) gejala awal yang ditimbulkan pada daun berupa penjernihan tulang daun (vein clearing) yang kemudian berkembang menjadi warna kuning, penebalan tulang daun, dan penggulungan daun (cupping). Infeksi lanjut Begomovirus menyebabkan daun-daun mengecil, berwarna kuning cerah dan tanaman menjadi kerdil.
Sedangkan infeksi pada gulma A. conyzoides yang terinfeksi Begomovirus menunjukkan gejala vein clearing atau penjernihan tulang daun (Sukamto et al. 2005). Haerani dan Hidayat (2003), menyebutkan infeksi Begomovirus menghasilkan gejala yang beragam pada studi penularan Begomovirus asal A. conyzoides terhadap beberapa tanaman Solanaceae (tomat, tembakau dan cabai rawit). Daun tanaman tomat yang terinfeksi mengeriting ke arah bawah, tulang daun menebal, dan tangkai daun melengkung ke bawah. Tanaman tembakau yang terinfeksi menunjukkan gejala berupa daun yang menggulung terutama daun muda, tulang daun menebal dan daun
melengkung ke arah bawah. Tanaman cabai rawit yang terinfeksi menunjukkan gejala daun yang melepuh, tulang daun menebal dan daun melengkung ke atas. Gulma yang terinfeksi Begomovirus menunjukkan gejala yang bervariasi tetapi gejala yang banyak ditemukan pada gulma adalah penguningan tulang daun (netting) (Sukamto et al. 2005).
2.2.3 Kisaran inang Begomovirus
Begomovirus memiliki kisaran inang yang cukup luas baik pada tanaman
budidaya maupun gulma. Tomat, cabai, tembakau, mentimun, terung, ubi kayu dan kacang-kacangan adalah inang Begomovirus dari tanaman budidaya. Babadotan (A. conyzoides) merupakan gulma yang telah dilaporkan sebagai inang Begomovirus di
daerah tropis dan subtropis (Sukamto et al. 2005). Gulma lainnya yang dapat menjadi inang Begomovirus adalah Sida spp., Macroptilium lathyroides, dan Wissadula amplissima yang ditemukan di Jamaika.
Malvastrum coromandelianum merupakan gulma yang dapat menjadi inang
Begomovirus di Guangdong, Cina (Wu et al. 2007). Achyranthes aspera, Euphorbia
heterophylla, Nicandra physaloides, Commelina erecta, Amaranthus spinosus,
Erigeron floribundus, A. conyzoides, Bidens pilosa, Sida acuta, Ipomoea batatas,
Amaranthus viridis, Portulaca oleracea, Cassia obtusifolia, Euphorbia hirta,
Calopogonium mucunoides, Clotalaria retusa, Trianthema portulacastrum,
Alternanthera sessilis, Celosia trigyna, Commelina diffusa, Chromolaena odorata,
Boerhavia diffusa, Physalis angulata dan Acanthospermum hispidis adalah 28 spesies
gulma yang potensial sebagai sumber tomato yellow leaf curl Begomovirus (TYLCV) di Tanzania. H. brevipes, P. floridana, C. juncea, A. conyzoides bunga putih dan ungu adalah gulma yang rentan terhadap Begomovirus isolat Segunung pada pengujian di rumah kaca (Sulandari et al. 2005). Brown and Nelson (1988) melaporkan terdapat beberapa spesies tanaman termasuk gulma, famili Solanaceae, Leguminosae, Malvaceae dan Asclepiadaceae yang merupakan inang TYLCV di Meksiko.
2.2.4 Penularan Begomovirus
Brown and Nelson (1988) melaporkan bahwa penyakit pada tanaman tomat di Meksiko yang disebabkan oleh Begomovirus tidak dapat ditularkan secara mekanik dengan cairan perasan, tetapi melalui serangga vektor. Hal ini didukung juga oleh Hull (2002) yang menyatakan bahwa Begomovirus tidak dapat menular secara mekanik atau pun melalui benih, hanya dapat menular dengan cara penyambungan tanaman sakit pada tanaman sehat serta melalui serangga vektor B. tabaci.
Penularan di alam secara alami diketahui hanya melalui vektor B. tabaci sehingga perannya menjadi sangat penting dalam penyebaran Begomovirus. Berdasarkan hasil penelitian Rusli et al. (2000), Begomovirus isolat Segunung yang ditularkan secara mekanik pada tanaman cabai besar dan cabai rawit tidak menghasilkan gejala sedangkan penularan dengan cara penyambungan menghasilkan gejala dengan kejadian penyakit sebesar 71,4% pada tanaman cabai besar dan 57,1% pada cabai rawit. Keefektifan penularan tertinggi diperoleh pada perlakuan penularan
melalui serangga vektor yaitu mencapai 70-80%. Hubungan antara B. tabaci dan Begomovirus berdasarkan lamanya virus bertahan pada vektor bersifat persisten
sirkulatif non propagatif yaitu virus tersebut berada dalam tubuh serangga untuk kemudian akan ditularkan pada tanaman sehat melalui proses makan (Harrison dan Robinson 1999).
Efisiensi penularan Begomovirus dengan B. tabaci melalui proses makan sangat dipengaruhi oleh lamanya masa akuisisi serangga tersebut, selain oleh jumlah serangga yang menularkan Begomovirus pada tanaman sehat (Rachmawati, 2003). Periode akuisisi minimum B. tabaci untuk menularkan TYLCV adalah selama 15 menit dan terus meningkat hingga mencapai tingkat maksimum setelah akuisisi selama 24 jam (Mehta et al.1994). Menurut Aidawati (2006) B. tabaci yang melalui periode makan akuisisi (PMA) dan periode makan inokulasi (PMI) masing-masing selama 15 menit mampu menularkan Begomovirus walaupun dengan efisiensi penularan yang berbeda-beda untuk tiap kombinasi biotipe B. tabaci dan strain Begomovirus yang berbeda. Pada PMA dan PMI tiga dan enam jam strain
Begomovirus isolat Bogor menghasilkan efisiensi penularan 80 − 100% dengan masa
inkubasi 9 hari.
B. tabaci termasuk ke dalam ordo Hemiptera dengan famili Aleyrodidae
(Henneberry dan Castle 2001). Serangga ini memiliki kisaran inang meliputi berbagai tanaman budidaya dan gulma, dapat berkembang dengan baik di daerah tropis dan subtropis (Kalshoven, 1981). Menurut Henneberry dan Castle (2001) ada 500 jenis tanaman yang dapat menjadi inang B. tabaci dengan preferensi yang berbeda. Salah
satu faktor yang mempengaruhinya adalah permukaan daun, serangga tersebut umumnya memiliki preferensi yang tinggi pada permukaan daun yang berambut (hirsute) dibandingkan dengan daun yang permukaannya tidak berambut (glabrous). Basu (1995) melaporkan ada 540 spesies dari 77 famili tanaman yang dapat menjadi inang B. tabaci. Menurut Bezerra et al. (2004) gulma Acanthospernum hispidum paling banyak terinfestasi B. tabaci pada lahan tomat di daerah Brazil dibandingkan dengan Amaranthus reflexus, Datura stramonium dan Euphorbia heterophylla.
2.3 Metode Identifikasi Begomovirus
Metode serologi merupakan cara yang paling sering digunakan untuk mendeteksi virus tumbuhan, baik menggunakan antibodi poliklonal maupun antibodi monoklonal. Kekurangan pada metode serologi untuk mendeteksi kelompok Begomovirus. Hal ini disebabkan sulitnya mendapatkan titer virus yang cukup untuk
digunakan dalam deteksi serologi. Sifat fisik dan kimia partikel Begomovirus sulit dimurnikan dalam bentuk stabil, sifat imunogenik dari virion yang lemah, dan protein selubung terutama untuk virus-virus yang ditularkan B. tabaci tidak dapat dibedakan melalui antiserum poliklonal maupun monoklonal (Robert et al. 1984).
Pendekatan secara molekuler telah banyak dilakukan untuk menentukan infeksi Begomovirus yang terjadi di lapang dan mengidentifikasi Begomovirus secara umum. Penggunaan polymerase chain reaction (PCR) merupakan teknik yang sangat sensitif dan spesifik untuk deteksi dan identifikasi patogen tanaman. Metode ini dapat digunakan untuk menunjukkan dengan tepat komposisi populasi patogen dan
keragaman genetik virus. PCR dan degenerate oligonucleotide primer telah digunakan untuk deteksi dan identifikasi genus Begomovirus (Farag et al. 2005).
Polymerase chain reaction (PCR) merupakan reaksi invitro untuk
menggandakan jumlah molekul DNA pada target tertentu dengan cara mensintesis molekul DNA baru yang berkomplemen dengan molekul DNA target dengan bantuan enzim dan oligonukleotida sebagai primer, serta dilakukan di dalam thermocycler. Panjang target DNA berkisar antara puluhan sampai ribuan nukleotida yang posisinya diapit sepasang primer. Primer yang berada sebelum target disebut primer forward dan primer yang berada setelah target disebut primer reverse. Enzim sebagai pencetak rangkaian molekul DNA baru disebut sebagai enzim polymerase. Untuk mencetak rangkaian tersebut dalam teknik PCR, diperlukan juga dNTPs yang mencakup dATP (nukleotida berbasa Adenine), dCTP (nukleotida berbasa Cytosine), dGTP (nukleotida berbasa Guanin) dan dTTP (nukleotida berbasa Thymine) (Muladno, 2002). PCR merupakan suatu metode yang menggunakan komponen‐komponen replikasi DNA untuk mereplikasi suatu fragmen DNA yang spesifik di dalam tabung reaksi.
PCR dikembangkan untuk mempercepat isolasi DNA spesifik tanpa membuat dan melakukan pustaka genom. Dua primer oligonukleotida pendek digunakan untuk mengapit daerah DNA yang akan diamplifikasi. Primer menguatkan dan mencangkok target sekuen, satu dari setiap rantai DNA. Primer menentukan fragmen yang akan diamplifikasi dan DNA polymerase mereplikasi DNA dengan memanfaatkan empat deoksiribonukelotida (dGTP, dATP, dCTP, dTTP) yang disediakan di dalam tabung
reaksi (Nurulita, 2011). Pada sebuah siklus amplifikasi, DNA didenaturasi pada temperatur tinggi, annealing primer dilakukan dengan menurunkan temperatur dan DNA polymerase memperpanjang DNA dari primer.
Pengulangan siklus denaturasi, annealing primer, dan sintesis DNA menghasilkan DNA melalui amplifikasi secara eksponensial. Sekitar 25 sampai 40 siklus pada umumnya digunakan di dalam thermalcycler, yaitu sebuah alat yang secara otomatis mengontrol temperatur dan waktu. Suatu DNA polymerase khusus yaitu Taq polymerase stabil pada suhu tinggi, yang diisolasi dari suatu bakteri thermofilik, Thermus aquaticus, yang hidup di sumber air panas. Produk hasil PCR dianalisis menggunakan elektroforesis gel (Barnum, 2005).
Identifikasi secara tepat spesies yang menginfeksi tanaman sangat penting untuk tindakan yang akan diterapkan dalam hal mengendalikan penyakit tersebut. Analisis perunutan nukleotida dan asam amino saat ini memiliki peranan yang tidak kalah penting didalam melakukan deteksi dan karakterisasi virus. Berdasarkan hasil analisis perunutan nukleotida dan asam amino dapat diketahui tingkat kesamaan nukleotida dan dapat menentukan kelompok suatu virus maupun strain-strain dari virus yang sama (Shukla et al. 1994). Analisis tersebut digunakan sebagai pelengkap proses identifikasi dan karakterisasi virus. Tujuan paling penting DNA sequencing adalah mencari pattern yang diketahui di dalam sekuen. Pattern ini bisa terlibat di fungsi biologis yaitu mengkode protein dan RNA serta mengontrol eskpresi gen dan replikasi DNA.
2.4 Analisis Filogenetika
Filogenetika adalah bidang ilmu yang berkaitan dengan biologi yang menyediakan fasilitas dalam bidang genetika, ekologi, dan evolusi biologi. Filogenetik merupakan cara melihat sejarah evolusi populasi, alat untuk memahami proses biologis, menggambarkan hipotesis hubungan antar taksa (genus, spesies, individu) yang diilustrasikan sebagai pohon.
Dharmayanti (2011) menyatakan, pohon filogenetik adalah diagram berbentuk pohon berakar atau tanpa akar yang terdiri atas cabang luar atau daun (berarti taksa (jamak, takson untuk tunggal), titik-titik dan cabang mewakili hubungan di antara taksa. Pohon filogenetik merupakan grafik dua dimensi yang menunjukkan hubungan antar organisme berdasarkan data genetik dan lainnya yang dihasilkan melalui analisis filogenetika.
Proses evolusi melibatkan mutasi genetik dan proses rekombinan dalam spesies untuk membentuk spesies yang baru. Sejarah evolusi organisme dapat diidentifikasi dari perubahan karakternya. Karakter yang sama adalah dasar untuk menganalisis hubungan satu spesies dengan spesies lainnya. Pohon filogenetik adalah pendekatan logis untuk menunjukkan hubungan evolusi antara organisme (Dharmayanti, 2011). Filogenetika diartikan sebagai model untuk merepresentasikan sekitar hubungan nenek moyang organisme, sekuen molekul atau keduanya (Brinkman and Leipe, 2001). Salah satu tujuan dari penyusunan filogenetika adalah untuk mengkonstruksi dengan tepat hubungan antara organisme dan mengestimasi perbedaan yang terjadi dari satu nenek moyang kepada keturunannya (LI et al. 1999).
Menurut Hidayat dan Pancoro (2006), terdapat tiga tahap yang dilakukan dalam melakukan proses analisis filogenetika molekuler, yaitu sequence alignment, rekonstruksi pohon filogenetika, dan evaluasi pohon filogenetika dengan uji statistik.
2.4.1 Sequence alignment
Tahap ini merupakan tahap pennetuan tingkat homolog dari satu sekuen DNA atau protein dengan pembanding lainnya yang ada pada Gen Bank. Tahap ini melibatkan dua sekuen yang homolog disebut pairwise alignment, sedangkan yang melibatkan banyak sekuen yang homolog disebut multiple alignment. Keberhasilan analisis filogenetika sangat tergantung kepada akurasi proses alignment. Pada tahap alignment sering ditemukan adanya gap, yang ditandai oleh garis putus-putus. Gap
terjadi karena adanya insersi dan atau delesi. Dalam prakteknya, gap bisa dianggap sebagai data yang hilang, walaupun dalam banyak kasus gap dapat dilibatkan dalam analisis karena bisa bersifat informative (Dharmayanti, 2011).
.
2.4.2 Rekonstruksi pohon filogenetika
Membangun sebuah pohon filogenetika berdasarkan karakter (menggunakan urutan nukleotida atau asam amino secara langsung dalam rekonstruksi pohon), dapat menggunakan empat metode yaitu Distance method (DM), Maximum Likelihood (ML), Bayessian Inference (BI), MP (Maximum parsimony).
2.4.2.1 Distance method
Metode jarak pertama kali ditemukan oleh Feng dan Doolitle; pengelompokan program oleh penulis tersebut menghasilkan sebuah penjejeran dan pohon dari set sekuen protein (Feng and Doolitle, 1996). Program CLUSTALW, digunakan untuk neighborjoining distance method sebagai panduan untuk multiple sequence
alignment. Program PAUP versi 4 merupakan pilihan untuk membentuk sebuah
analisis filogenetika dengan distance method. Program PHYLIP package yang membentuk analisis distance termasuk program yang secara otomatis dibaca dalam sekuen dalam PHYLIP infile format dan secara otomatis menghasilkan file yang disebut dengan tabel distance. Metode jarak bekerja pada jumlah perubahan diantara masing-masing pasangan dalam kelompok untuk mengkonstruksi pohon filogenetika dalam kelompok.
Pasangan sekuen yang mempunyai jumlah perubahan terkecil diantara mereka disebut neighbors. Pada pohon, sekuen-sekuen ini menggunakan secara bersama-sama satu titik atau posisi common ancestor dan masing-masing dihubungkan titik oleh sebuah cabang. Tujuan dari metode jarak adalah metode untuk mengidentifikasi pohon pada posisi neighbors dengan benar, dan juga mempunyai cabang yang menghasilkan data orisinil sedekat mungkin. Penemuan neighbors terdekat diantara kelompok sekuen dengan metode jarak biasanya langkah pertama dalam memproduksi sebuah multiple sequence alignment.
2.4.2.2 Maximum likelihood dan bayessian inference
Metode maximum likehood menampilkan kesempatan penambahan untuk mengevaluasi pohon dengan variasi dalam rata-rata mutasi dalam lineage yang berbeda. Metode ini dapat digunakan untuk mengekplorasi hubungan antara sekuen yang lebih beragam, dimana kondisi ini tidak dapat dilakukan dengan baik jika menggunakan metode maximum persimony (LI et al. 1999; Dharmayanti, 2011). Kekurangan metode maximum likehood adalah membutuhkan pekerjaan komputer yang sangat intensif. Jika menggunakan komputer yang lebih cepat, metode maximum likehood dapat digunakan untuk model evolusi yang lebih komplek. Metode ini juga
dapat digunakan untuk menganalisa mutasi pada overlapping reading frame pada virus
(SCHADT et al. 1998). Metode ini mirip dengan metode maximum parsimony dalam analisis yang dibentuk pada masing-masing kolom dalam multiple
sequence alignment. Semua kemungkinan pohon yang terbentuk dipertimbangkan,
sehingga metode ini hanya cocok untuk sekuen dalam jumlah kecil. Metode ini mempertimbangkan untuk masing-masing pohon, jumlah perubahan sekuen atau mutasi yang terjadi yang memberikan variasi sekuen maximum likehood.
Untuk Bayessian Inference merupakan metode yang mengartikan bahwa perubahan-perubahan diantara semua basa nukleotida adalah sebanding. Masalah serius dari metode ini adalah waktu perhitungan yang lama, walaupun telah dikembangkan algoritma baru yang dianggap dapat mempercepat proses perhitungan (Hidayat dan Pancoro, 2006). Pada dasarnya metode ini adalah sama dengan
likelihood methode, hanya berbeda dalam penghitungan distribusi prior untuk
membangun pohon filogenetika. Salah satu metode untuk menghitung distribusi prior adalah metode MCMC (Markov chain Monte Carlo).
2.4.2.3 Maximum parsimony
Parsimony atau metode minimum evolution pertama kali digunakan dalam
filogenetik oleh Camin and Sokal pada tahun 1965 (Felsenstein, 1978). Metode ini memprediksikan pohon evolusi/ evolutionary tree yang meminimalkan jumlah langkah yang dibutuhkan untuk menghasilkan variasi yang diamati dalam sekuen. Untuk alasan ini, metode ini juga sering disebut sebagai metode evolusi minimum/minimum evolution method (Dharmayanti, 2011). Sebuah multiple sequence alignment dibutuhkan untuk memprediksi posisi sekuen yang sepertinya
berhubungan. Posisi ini akan menampilkan kolom vertikal dalam multiple sequence alignment. Untuk masing-masing posisi yang disejajarkan, pohon filogenetika
membutuhkan perubahan evolusi dalam jumlah terkecil untuk menghasilkan pengamatan perubahan sekuen yang diidentifikasi (Mount, 2001). Analisis ini terus menerus dilakukan terhadap masing-masing posisi dalam penjejeran sekuen. Akhirnya, pohon yang menghasilkan jumlah perubahan terkecil secara keseluruhan dihasilkan untuk semua posisi sekuen yang diidentifikasi (Dharmayanti, 2011).
Dari keempat metode di atas, PM sangat sering dipilih, antara lain karena pohon yang dibentuk lebih menggambarkan perubahan evolusioner yang terjadi setiap waktu, mengandung asumsi bahwa proses evolusi akan menempuh jalan yang
paling singkat (parsimonious), dan perhitungan relatif lebih sederhana dan cepat dengan tingkat realibilitas yang tinggi.
2.4.3 Evaluasi pohon filogenetika
Menurut Hidayat dan Pancoro (2006), evaluasi pohon filogeni ini bertujuan untuk memastikan tingkat kepercayaan dari pohon tersebut. Proses ini dilakukan dengan menerapak beberapa metode yaitu interior branch test (IB) dan Felsentein’s bootstrap test (FB). Secara umum prinsip kerja dari metode evaluasi IB adalah
estimasi pohon dengan menguji reliabilitas setiap cabang sebelah dalam (interior branch).
Felsentein’s bootstrap test (FB), menguji tingkat reliabilitas dengan
menggunakan metode Efron’s bootstrap. Evaluasi pohon dilakukan menggunakan analisis bootsrap sebanyak 1.000 ulangan, dimana sebuah set dari site basa nukleotida diambil secara acak dan dilakukan secara berulang, kemudian dilakukan konsensus, sehingga hanya satu pohon filogenetika yang dihasilkan. Pada dasarnya pola perubahan basa nukleotida sangat rumit dan sering berubah sejalan dengan waktu evolusi, sehingga metode FB sangat baik digunakan dalam mengevaluasi pohon filogenetika (Hidayat dan Pancoro, 2006).