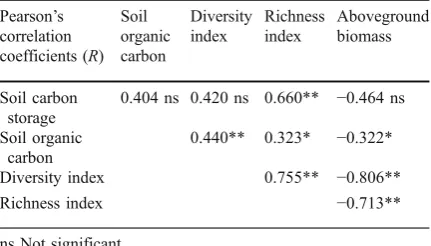
Effects of artificial grassland establis
Teks penuh
Gambar
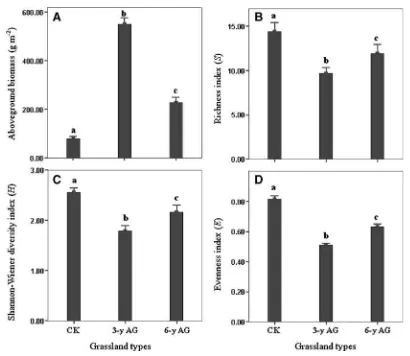
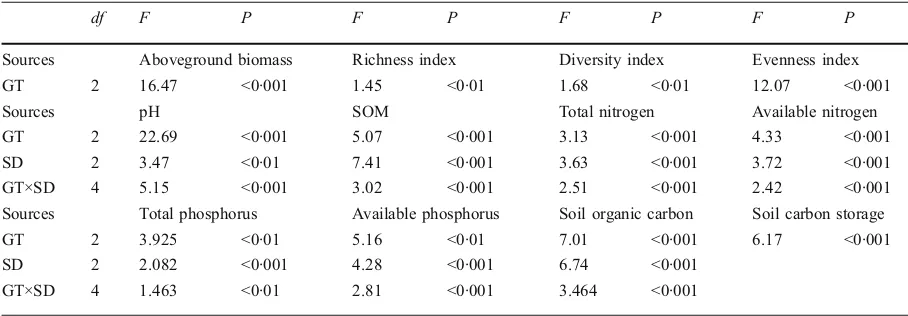
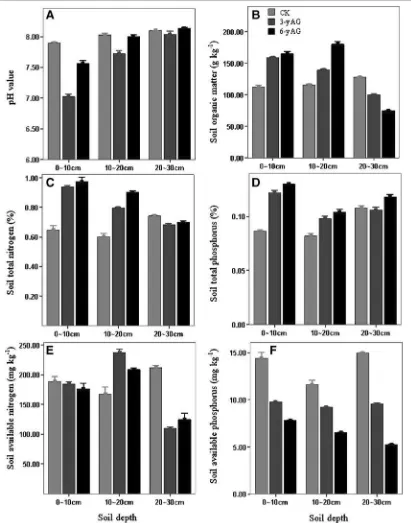
Garis besar
Dokumen terkait
Perbedaan total waktu penyelesaian penjadwalan perusahaan dengan usulan dengan metode LRPT berbeda sebesar 414 menit dan perbedaan waktu tersebut dapat digunakan
Konfirmasi tersebut membutuhkan waktu selama 2.129,5 detik untuk setiap unit truck sehingga terjadi kegiatan menunggu pada proses membuat surat jalan dan form rute
Tujuan penelitian adalah untuk mengetahui dan mengevaluasi kualitas kimia daging itik Peking meliputi massa lemak, massa protein dan massa kalsium daging yang diberi
Jika Peserta didik dapat menuliskan syarat dan rukun shalat jenazah yang akan dimandikan dengan lengkap, skor 3 c.. Jika Peserta didik dapat menuliskan syarat dan
The post-training assessment indicated that there was increase in participants’ knowledge on business record keeping, profit/loss calculation, product pricing, savings, loans as
Termasuk kategori thaghut adalah setiap orang yang memposisikan dirinya sebagai musyarri' (pembuat hukum dan perundang-undangan) bersama Allah, baik dia itu sebagai pemimpin
Pembagian tugas terhadap dosen wali untuk melaksanakan bimbingan terhadap mahasiswa bukan hanya terbatas pada kontrak studi, tetapi harus dapat memberikan pemahaman
BERITA ACARA PENETAPAN PEMENANG Nomor: 09/INDAGKOP/LU.BAPP/II/ULP-P1/V-2012 Tanggal 31 MEI 2012 maka UNIT LAYANAN PENGADAAN KELOMPOK KERJA PEKERJAAN KONSTRUKSI Kabupaten kepulauan
