Summary Artificially increasing pollen supply (supplemen-tal mass pollination (SMP)) to conifer seed orchards has the potential to increase seed yields and improve the genetic worth of seed crops that would otherwise suffer from the detrimental effects of pollen contamination and unbalanced paternal con-tribution. However, success rates, measured as the proportion of seed fertilized by SMP, have been low. This review examines the concepts underlying SMP and presents data for two SMP field trials with Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) and white/Engelmann spruce (Picea glauca (Moench) Voss, Picea engelmannii Parry ex Engelm.). The trial results are discussed with respect to pollen fertility, pollination tech-nique, and competing pollen cloud density. I also summarize methods for ensuring the successful handling of pollen ex situ. Keywords: Douglas-fir, genetic improvement, interior spruce, seed yield, supplemental mass pollination.
Introduction
The concept of producing genetically superior conifer seed from orchards is well established (Feilberg and Søegaard 1975) and constitutes the link between tree breeding and plan-tation forestry. Considerable progress has been made in under-standing the reproductive biology of conifers, particularly the factors affecting seed production in orchards. Asexual tech-niques (e.g., rooted cuttings, somatic embryogenesis) are be-ing developed; however, seed orchards remain the most economical and robust method for managing genetic effi-ciency (the degree to which genetic gain and genetic diversity are optimized).
Although there are several factors that restrain the realiza-tion of full genetic efficiency in wind-pollinated seed orchards (Eriksson et al. 1973, Woessner and Franklin 1973), genetic efficiency of seed orchards can be maintained or enhanced by several techniques including orchard cooling (Fashler and El-Kassaby 1987, Blush et al. 1993), cone induction (Ebell 1972, Masters 1982, Wheeler et al. 1985, Ross et al. 1985), and artificially (supplemental mass pollination (SMP)) increasing pollen supply (Bridgwater et al. 1993). Despite the potential for manipulating the genetic structure of orchard crops through pollen management (Askew 1992, Lambeth 1993), the results for open-pollinated orchards have not been encouraging.
This review summarizes current knowledge on SMP in seed
orchards. Factors affecting SMP success are discussed with reference to two seed orchard species, Douglas-fir (Pseudot-suga menziesii (Mirb.) Franco) and white spruce/Engelmann spruce mixture (Picea glauca (Moench) Voss, Picea engel-mannii Parry ex Engelm.), hereafter referred to as interior spruce. Data from two field trials are presented and discussed with respect to pollen application technique, pollen viability and competing pollen cloud density.
Supplemental mass pollination
Supplemental mass pollination is the broadcast application of pollen to seed-cone buds that are not isolated (i.e., open-polli-nation) from airborne pollen (Bridgwater et al. 1993). It has been successfully used to improve seed yields (Daniels 1978, Bridgwater and Bramlett 1982, Wheeler and Jech 1986, Web-ber 1987, El-Kassaby and Reynolds 1990), to control paternal composition of orchard seed (El-Kassaby and Ritland 1986a, 1986b, El-Kassaby et al. 1986, El-Kassaby and Reynolds 1990), to introduce specific genotypes into the seed orchard (Woessner and Franklin 1973, Hadders 1984, Bridgwater et al. 1987, Eriksson et al. 1994), and to reduce self-fertilization and contamination (El-Kassaby and Ritland 1986b, Wheeler and Jech 1986).
Supplemental mass pollination success rates, defined as the proportion of viable seed fertilized by pollen parents arising from SMP, range from 4 to 90%. Wheeler and Jech (1986) reported an average 45% success rate for Douglas-fir (range 4--76%) and Blush (1987) reported values between 48 and 70% for pollen applied directly to receptive seed cones of loblolly pine (Pinus taeda L.). Conversely, Yazdani et al. (1986) could account for only 4% of the applied pollen when a remote-controlled helicopter was used in a Scots pine (Pinus sylvestris L.) seed orchard. Eriksson et al. (1994) found SMP success rates varied between 10 and 23% for Scots pine pollen dusted over the tree; however, when the pollen was applied directly to receptive seed cones, success rates ranged from 66 to 84%. Recently, Eriksson et al. (1995) have shown that the amount of pollen produced by orchard trees directly affects the success rate of SMP.
The timing of SMP also affects the success rate. El-Kassaby and Ritland (1986b) segregated a Douglas-fir seed orchard into early, mid and late reproductive phenological classes. Out-crossing rates were highest and contamination rates lowest
Pollen management for intensive seed orchard production
JOE E. WEBBER
B.C. Ministry of Forests, Glyn Road Research Station, 1320 Glyn Road, Victoria, B.C. V8W 3E7, Canada
Received August 10, 1994
within the mid-phenological period. Conversely, outcrossing rates were lowest and contamination rates highest in the clones flowering either early or late. Because competing pollen cloud density was lowest in these two periods, El-Kassaby and Rit-land (1986b) suggested restricting SMP treatments to these periods.
Wheeler and Jech (1986) also found that the effectiveness of SMP applied to 12--14-m tall Douglas-fir trees varied with reproductive phenology within the tree. The average success rate of SMP was 45%. It was highest (54%) when the seed cone was at its prime receptive period (defined as 25--75% of bud scales exposed for less than 3 days) and lowest (28%) at earlier stages (only a few bud scales exposed). When pollen was applied late (defined as 75% of bud scales exposed for more than 3 days), the success rate averaged 38%, indicating that cones need not be pollinated immediately after bud burst to achieve positive results.
In a second study, Wheeler and Jech (1988) compared phe-nological periods of specific Douglas-fir clones to the relative success of a single SMP. All seed cones were at the same stage of receptivity (cones at an early stage of receptivity were removed), but competing pollen cloud density was not consid-ered as a source of variation. In early flowering clones, SMP success rate was 80%, whereas the rate varied between 60 and 90% in late flowering clones.
Supplemental mass pollination trials Douglas-fir
I compared pressurized direct- and mist indirect-SMP tech-niques, each with single and double pollen applications, on eight clones (single tree treatments) in each of two mature Douglas-fir orchards (17 and 10--11 years old). A single pollen lot with an isozyme (IDH) allele that occurs in less than 5% of the coastal Douglas-fir population (Yeh and O’Malley 1980, Webber and Yeh 1987) was used. The pollen lot exhibited moderate to low viability (Webber and Bonnet-Masimbert 1993). Table 1 shows the average (eight clones by two or-chards) filled seed per cone (FSPC) and percent SMP success
rate (%SP). Supplemental mass pollination had no effect on seed yields compared to the open-pollinated controls; how-ever, paternal composition was affected. In both orchards, the average success rate for SMP by the pressurized direct-polli-nation technique was 19.1% for a single pollen application and 22.0% for two applications, indicating that a single application is sufficient, especially if pollination is timed to the optimal stage of receptivity (i.e., 2--4 days after bud burst). The mist indirect-pollination technique was less effective than the di-rect-pollination technique; the corresponding success rates for one and two pollen applications were 11.6 and 17.1%, respec-tively. The corresponding percent of seed with the IDH marker from open-pollinated cones on the same tree but not subjected directly to SMP was 4.6%. The paternal source of this seed was attributed to residual pollen from the SMP treatments. One tree in Orchard 2 carried the rare IDH allele and was excluded from the paternity analysis.
Average seed yield from a single controlled cross-pollina-tion using SMP was 20.6 FSPC compared to 30.3 FSPC for open-pollinated controls. Seed yields from one and two direct-pollinations (21.3 and 20.3 FSPC, respectively) were similar to those obtained from a single controlled cross-pollination (20.6 FSPC), whereas seed yields from one and two mist-pol-linations (26.2 and 25.2 FSPC, respectively) were more similar to those obtained from open-pollinated control cones (30.3 FSPC) than those obtained from a single controlled cross-pol-lination (20.6 FSPC). These results support the hypothesis that direct-pollination delivers pollen to the receptive sites of the seed cone and loads the stigmatic tips more effectively than the mist system. I conclude that the mist-pollination technique left more receptive sites exposed to be pollinated by the higher viability, orchard pollen cloud (untreated, open-pollinated yields were the highest at 30.6 FSPC).
Figure 1 shows the average (by clone) filled seed per cone and percent SMP success rate for each of the two orchards. Although seed yields from direct-pollination were similar in both orchards, the seed yields from mist-pollination and open-pollination were less in Orchard 2 than in Orchard 1 as a result of differences in pollen cloud density between the two or-chards.
Pollen cloud density (PCD) was determined with either a 7-day clock mechanism mounted on a rotating platform (Or-chard 1) or a double-sided sticky tape on a dowel (Or(Or-chard 2). The 7-day clock mechanism is a modification of the Ebell and Schmidt (1964) and Sarvas (1968) recording monitors which used a 7-day recording clock mechanism with a piece of petroleum-jelly-coated celluloid film. The celluloid film was marked into seven sections each equaling a 24-h period (Web-ber and Painter 1995). Pollen grains were counted over a defined area and expressed as grains per mm2 per day.
Figure 2 shows the average (six stations in Orchard 1 and four in Orchard 2) daily pollen cloud density values (± stand-ard errors) for the seed cone receptivity period of the two orchards. The daily PCD values were summed over the recep-tivity period of each tree within each of the two orchards to give the pollen load for each tree’s receptivity period. The receptivity period of each tree was defined as the 2-day period Table 1. Mean filled seed per cone (FSPC) and percent success rate
(%SP), with standard errors in parentheses, for an SMP trial averaged over eight clones in each of two orchards using pressurized direct-pol-lination and mist indirect-poldirect-pol-lination applied either as a single or double application. Corresponding values for controlled cross-pollina-tion (CC) and open-pollinated (OP) seed yields are also shown (n = 16).
Pollination treatment FSPC (± SE) %SP (± SE)
before and after pollination including the day of pollination (5 days total). Table 2 shows the total PCD (pollen load) values for seven clones in Orchard 1 and eight clones in Orchard 2.
In a separate study, PCD data were determined for cones on 15 Douglas-fir trees exposed to the orchard pollen cloud for five periods in each of three phenological periods. There was an asymptotic relationship between average PCD and seed yield (Figure 3). As pollen cloud density values increased, there was a general linear increase in seed yield; however, there was a point beyond which further increases in pollen cloud density were not associated with an increase in seed yield. I defined this point as 100% pollen load. In Douglas-fir, pollen cloud density values in the order of 4--8 pollen grains mm−2 day−1 summed over the receptivity period of the seed cone (or
tree) approximated 100% pollen load. Sorenson and Webber (unpublished data) have developed this relationship to give:
y = 41.2 exp(−1.7 /x) (r2 = 0.792), (1)
where y is the filled seed per cone, and x is the summed pollen cloud density over the exposure period for the seed cone.
Seed yields (FSPC) for open-pollinated cones in each of the two orchards as well as their corresponding predicted seed yield (FSPC) calculated from Equation 1 are shown in Table 2. In this relationship, the PCD values were summed over the receptivity period of the exposed cones and related to their corresponding seed yields. Equation 1 accurately predicted the seed yields for Orchard 1, especially when the PCD values
Figure 1. Filled seed per cone and pcent SMP success rate (with standard er-ror bars) for two pollination techniques (D = direct and M = mist), each with a single and double application in each of two Douglas-fir orchards. Filled seed per cone values for control cross-pollina-tion (CC) and open-pollinated seed cones (OP) are also shown (n = 16 for Orchard 1, and n = 14 for Orchard 2).
were averaged over the clones, but the prediction was less accurate for Orchard 2 (Table 2). The discrepancy in the accuracy of prediction between the two orchards is attributed
to the differing pollen capturing efficiencies of the 7-day clock and sticky tape devices.
The effects of application technique and competing pollen cloud density (pollen load) largely explain the results of this SMP trial. However, if the pollination hypothesis is correct, the results for percent SMP success are lower than expected, especially in Orchard 2 where seed yields for open-pollinated cones and pollen load were less than in Orchard 1. According to the pollination hypothesis, direct-pollination loads the re-ceptive sites of seed cones more effectively than indirect mist-pollination. Thus, direct-pollination should produce a higher success rate for SMP than indirect-pollination. The highest success rate for SMP was from two direct-pollinations but the percent efficiency was only 22%. Because only the controlled cross-pollinations were isolated, whereas the treated cones were exposed to the orchard pollen cloud throughout their receptive period, one explanation for the low success rate may be that high viability orchard pollen reached the receptive sites before the low viability supplemental pollen was applied. Pollen arriving during the early phases of receptivity of seed cones of Douglas-fir has a higher probability of successfully completing the steps toward fertilization than pollen arriving during the later phases (Webber and Yeh 1987). These trial results confirm the importance of pollen viability and timing in optimizing SMP success and suggest that the pollination efficiency hypothesis is dependent not only on application technique but also on pollen viability. If high viability pollen had been used instead of the moderate to low viability pollen, seed yields might have remained the same because of the high pollen load, but success rates might have been higher.
El-Kassaby et al. (1993) reported similar results for the same orchard (but with a different set of clones), pollinated between one and three times with a hand-operated insecticide sprayer, similar to our mist applicator. Two pollen applications were as effective as three and both were better than one application. The success rate of the best SMP treatment averaged 18%.
Interior spruce
Douglas-fir has a dry pollination mechanism (Owens et al. 1981) whereas Picea and Pinus species have a pollination mechanism that involves a pollination drop (Owens and Blake 1985). Because the drop occurs a few days after bud burst, the hypothesis that pollen arriving early has a competitive advan-tage over pollen arriving later may not apply to interior spruce. Pollen applied early (before the emergence of the drop) will produce seed, but the best yields are obtained at the mid-2 stage of receptivity, defined when the cone axis has fully elongated from the bud scales and the ovuliferous scales begin reflexing and curling at the edge (Webber 1991). Phenological development in interior spruce is sensitive to temperature, and the number of days to reach optimal seed cone receptivity varies between 3--4 days in warm weather to 10--14 days in cool, wet weather. Under average field conditions (day/night temperatures about 20/5 °C), the optimal time to pollinate interior spruce corresponds to about 4--7 days after bud burst. Figure 4 shows the number of filled seed per cone (with standard error bars) obtained from a SMP trial for interior Table 2. Total daily pollen cloud density (PCD) values (grains mm−2
day−1) summed over the receptive period (5 days) of each treated Douglas-fir tree. The filled seed per cone (FSPC) for open-pollinated cones and their corresponding predicted FSPC (using an exponential function1) are also shown. The total PCD values shown represent the average daily PCD for six stations in Orchard 1 and four stations in Orchard 2.
Tree Receptivity Total PCD FSPC Predicted
period FSPC1
Orchard 1
Tree 1 April 2--6 3.4 24.3 24.9
Tree 2 April 3--7 3.7 53.2 26.0
Tree 3 April 4--8 4.8 38.3 28.9
Tree 4 April 8--12 8.6 40.7 33.8
Tree 5 April 5--9 5.4 32.5 30.1
Tree 6 April 10--14 9.3 25.0 34.3
Tree 7 April 13--17 15.2 16.5 36.8
Average 7.2 32.9 32.5
Orchard 2
Tree 1 April 8--12 5.0 32.7 29.3
Tree 2 April 9--13 4.0 28.2 26.9
Tree 3 April 14--18 0.7 27.7 3.6
Tree 4 April 12--16 1.5 30.5 13.3
Tree 5 April 10--14 3.2 31.5 24.2
Tree 6 April 14--18 0.7 12.5 3.6
Tree 7 April 12--16 1.5 27.2 13.3
Tree 8 April 14--18 0.7 30.0 3.6
Average 2.2 27.5 19.0
1 The predicted values shown were calculated using an exponential
equation: FSPC = 41.2exp(−1.7/PCD) (Sorenson and Webber, unpub-lished data). The average predicted value for each orchard was calculated using the average PCD values of the receptivity periods of individual trees within the orchard.
Figure 3. The relationship between total pollen cloud density (n = 2) and filled seed per cone (n = 15) for a Douglas-fir seed orchard. The relationship is best explained (r2 = 0.792) by an exponential function:
spruce at the Kalamalka Seed Orchards, Vernon, B.C., Canada. The pressurized direct-pollination produced higher seed yields than mist indirect-pollination, and two pollen applications were only marginally better than one. Paternity analyses of these seed lots were not done. However, because pollen pro-duction in this and surrounding orchards was low, and conse-quently, competing pollen cloud density was low, the seed yields obtained from SMP were principally from the applied pollen.
The results for the SMP trials in Douglas-fir and interior spruce indicate that pollen quality, timing of application, fre-quency of application, application technique, and competing pollen cloud density all affect the success rate of SMP. The following discussion considers the factors affecting pollen viability ex situ.
Pollen collection
An important factor affecting pollen fertility ex situ is pollen maturation stage at harvesting. There is considerable variabil-ity in seed set among years, presumably as a result of variation in environmental conditions during pre- and post-meiotic de-velopment of the pollen cone bud. Temperature effects on pollen development have been described for Norway spruce (Picea abies (L.) Karst.) (Eriksson et al. 1970b, Jonsson 1974, Ross 1988), larch (Larix) (Eriksson 1970, Eriksson et al. 1970a) and western hemlock (Tsuga heterophylla (Raf.) Sarg.) (Colangeli and Owens 1991). Although little is known about cultural effects on pollen and seed cone maturation, it is as-sumed that treatments favoring growth will also facilitate pol-len and seed cone development. Several studies of angiosperm plants (Snow and Spira 1991) suggest that the cultural and environmental conditions prevailing during vegetative and re-productive development have important effects on both yields and genetic adaptation of progeny (Ottaviano and Mulcahy 1989).
A substantial source of yearly variation in fertility of pollen can be attributed to collection time. Under ideal conditions,
pollen cones should be harvested for extraction as close to shedding as possible (Beers et al. 1981). Under operational conditions, this is difficult because the time between optimal stage for harvesting and shedding can be as little as a few hours. The problem is further compounded by variation in maturity of pollen buds both at the branch and tree levels.
We have investigated the effects of harvesting interior spruce pollen at two stages of post-meiotic maturation and attempted to artificially mature pollen ex situ. Maturation effects were measured by pollen yields, extraction efficiency (dry weight of pollen extracted to the dry weight of original pollen buds), and fertility (defined as seed set). Early maturation was defined as the stage when pollen-cone buds had enlarged sufficiently to break the bud scales but the microsporangia were still tightly packed. The late maturation stage was defined as the preshed-ding bud. Pollen-cone buds were harvested at the defined maturation stage and either extracted immediately or condi-tioned in an atmosphere of 100% RH at 10 °C for 48 h.
Table 3 shows the extracted dry weight of pollen, the extrac-tion efficiency, and the resulting filled seed per cone (with standard errors) for pollen-cone buds harvested early and late and preconditioned at 100% RH for 48 h. Extracted pollen dry weight, extraction efficiency and seed set (FSPC) of pollen-cone buds harvested early were all reduced compared to pol-len-cone buds harvested and extracted closer to natural shedding. Exposing the pollen-cone buds harvested early to 100% RH at 10 °C for 48 h improved the amount of pollen extracted as well as seed set, whereas treating pollen-cone buds harvested at a later stage of maturity did not affect pollen dry weight or extraction efficiency and seed set was reduced. We conclude that harvesting pollen-cone buds as close to shedding as possible is the best method.
An alternative to harvesting and extracting pollen-cone buds ex situ is to collect naturally shedding pollen. Both the system described by Philippe and Baldet (1992) for collecting pollen from larch orchards and the vacuum system used by Copes et al. (1991) for Douglas-fir orchards have merit. In addition to facilitating pollen harvesting, there are two additional benefits to vacuum collection: the quality of vacuum-collected pollen is equal to or greater than that obtained by harvesting mature pollen-cone buds, and the water content of the vacuum-col-lected pollen is reduced so that direct freezer storage may be possible (Copes et al. 1991).
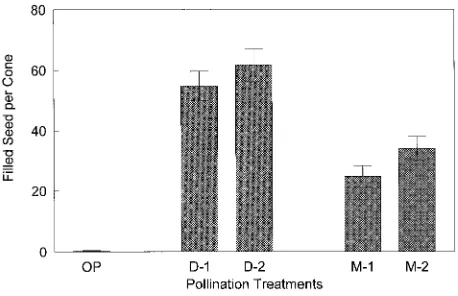
Figure 4. Filled seed per cone (with standard error bars) for two pollination techniques (D = direct and M = mist), each with a single and double application in an interior spruce orchard. Corresponding values for open-pollinated (OP) seed cones are also shown (n = 16).
Table 3. The effect of maturation stage and preconditioning of interior spruce pollen-cone buds. Buds were harvested early (post-meiotic) and late (preshedding) and extracted (0 h) or preconditioned at 100% RH at 10 °C for 48 h. Treatment response was measured on extracted pollen dry weight (g), percent extraction efficiency (EE), and seed set (FSPC) with standard errors in parenthesis (n = 24).
Stage Dry weight EE FSPC
Early-0 2.8 (0.72) 35.5 (1.10) 52.3 (4.1) Early-48 3.8 (0.21) 46.0 (0.86) 89.9 (4.4)
Late-0 4.9 (0.24) 49.5 (0.55) 87.1 (3.1)
Pollen drying and storage
Optimal conditions for drying pollen are species specific. In general, pollen is dried at temperatures between 20 and 30 °C and a relative humidity of 20 to 40% (Sprague and Snyder 1981). Although there are variations among species with re-spect to the optimal pollen storage conditions, all species respond similarly (Bramlett and Matthews 1991, Webber 1991, Jett et al. 1993, Webber and Painter 1995). Pollen water content must be below 10% (range 4--8%). The dried pollen should be stored in an air-tight container, preferably with the air (oxygen) evacuated or replaced with nitrogen, at −20 °C or under liquid nitrogen (Copes 1985, 1987).
Pollen testing
Several in vitro pollen viability tests are applicable to conifer pollen (Stanley and Linskens 1974, Ching and Ching 1976), and occasionally they have been successfully related to seed set (Moody and Jett 1990, Webber and Bonnet-Masimbert 1993). In vitro pollen germination is a widely used test; how-ever, in vitro germination is also the most sensitive to assay conditions. Effects of the assay medium on pollen germination have been demonstrated for Douglas-fir (Webber and Bonnet-Masimbert 1993), western larch, Larix occidentalis Nutt. (Webber and Ross 1995) and Scots pine (Bonnet-Masimbert and Webber 1995).
Hydration effects on pollen germination both in vivo and in vitro are also important. Pollen hydration is essential for in vivo germination to proceed (Heslop-Harrison 1987), and the same is apparently true for in vitro germination. Improved germina-tion response after pollen hydragermina-tion has been demonstrated in Douglas-fir (Charpentier and Masimbert 1983, Webber and Bonnet-Masimbert 1993), loblolly pine (Jett and Frampton 1990) and western larch (Webber and Ross 1995). Although pollen hydration improves the in vitro assay response, there is no effect on seed set (Mellerowicz and Bonnet-Masimbert 1986, Webber 1991).
Respiration and conductivity (see Webber and Bonnet-Masimbert 1993) are also useful in vitro assays. Respiration is the least sensitive test to assay conditions and consistently gives the best indication of pollen fertility potential in Douglas-fir. Respiration is relatively insensitive to prehydra-tion, whereas conductivity tests are very sensitive to hydration state (Webber 1991, Webber and Bonnet-Masimbert 1993).
If pollen viability assays are used to estimate potential fertility, then correlation analyses between assay response and seed set must be completed. This was done for loblolly pine (Moody and Jett 1990) and Douglas-fir (Webber and Bonnet-Masimbert 1993), and for Douglas-fir, threshold values were established using a particular pollen lot for controlled cross-pollination. These threshold values are useful for estimating potential fertility of a single pollen lot, but they are less useful for determining paternal contribution from pollen mixes (i.e., used for SMP) or under open pollination when competition from other pollen sources is high.
An important source of variability in fertility trials (meas-ured by seed set) is the pollen lot (male) by clone (female)
interaction. Variability in paternal contribution from pollen mixes has been demonstrated for several conifer species (Mo-ran and Griffin 1985, Schoen and Cheliak 1987) and may be caused by differential pollen viability (Webber and Yeh 1987). Apsit et al. (1989) suggest that the male--female interaction is significant and has a genetic basis. If substantial variability occurs for pollen lots used in pollen mixes, then Fowler (1987) suggests diluting pollen mixes and maintaining the number of pollen parents comprising the mix to about 20 (Fowler and Wiselogel 1993).
Pollen application
Data presented for the two SMP trials emphasized the impor-tance of pollination technique, pollen viability and the timing of pollen application (Owens and Blake 1985, Owens 1993). We have described the importance of direct-pollination using dry pollen. For those species with a wet pollination mechanism (e.g., Picea and Pinus), aqueous pollinations may be beneficial especially when drought conditions occur during pollen drop formation. In western hemlock, pollen viability may be as important as pollination timing and application technique. Colangeli and Owens (1989) describe a mechanism by which the bracts of receptive western hemlock seed cones capture the pollen and show how the germinating pollen tube grows over the bract surface to the ovules, through the micropyle, and into the nucellus. This mechanism would favor those pollen grains that possess high viability and are capable of rapid germ tube growth. The optimal time to pollinate western hemlock may be less important than loading the bracts early with high viability pollen.
Conclusions
Factors limiting the success of SMP include competing pollen cloud density and the technical difficulties associated with administering an effective pollen delivery system. The success of SMP can be improved somewhat by the use of high viability pollen and effective application techniques; however, average success rates greater than about 20% do not seem feasible under current orchard conditions, although alternative orchard designs (i.e., clonal rows) and intensive management practices (i.e., crown pruning) may facilitate higher success rates. For mature orchards containing low breeding value parent mate-rial, SMP may not be cost effective, whereas for orchards that are at risk from contamination or are established from ad-vanced generation breeding material, a 20% increase in genetic value may be acceptable (Askew 1992).
References
Apsit, V.J., R.R. Nakamura and N.C. Wheeler. 1989. Differential male reproductive success in Douglas-fir. Theor. Appl. Genet. 77:681--684.
Beers, W.L., J. Biven and J.E. Mocha. 1981. Pollen collection. In Pollen Management Handbook. Agriculture Handbook 587. Ed. E.C. Franklin. USDA Forest Service, Washington DC, pp 30--32. Blush, T.D. 1987. An operational trial of supplemental mass
pollina-tion in a loblolly pine seed orchard. In Proc. 19th South. For. Tree Improve. Conf. College Station, Texas, USA, pp 240--246. Blush, T.D., D.L. Bramlett and Y.A. El-Kassaby. 1993. Reproductive
phenology of seed orchards. In Advances in Pollen Management. USDA Handbook 698. Eds. D.L. Bramlett, G.R. Askew, T.D. Blush, F.E. Bridgwater and J.B. Jett. USDA Forest Service, Wash-ington DC, pp 15--23.
Bonnet-Masimbert, M. and J.E. Webber. 1995. From flower induction to seed production in forest tree orchards. Tree Physiol. 15:419--426.
Bramlett, D.L. and F.R. Matthews. 1991. Storing loblolly pine pollen. South. J. Appl. For. 15:153--157.
Bridgwater, F.E. and D.L. Bramlett. 1982. Supplemental mass pollina-tion to increase seed yields in loblolly pine seed orchards. South. J. Appl. For. 6:100--104.
Bridgwater, F.E., D.L. Bramlett and F.R. Matthews. 1987. Supplemen-tal mass pollination is feasible on an operational scale. In Proc. 19th South. For. Tree Improve. Conf. College Station, Texas, USA, pp 216--222.
Bridgwater, F.E., T.D. Blush and N.C. Wheeler. 1993. Supplemental mass pollination. In Advances in Pollen Management. USDA Handbook 698. Eds. D.L. Bramlett, G.R. Askew, T.D. Blush, F.E. Bridgwater and J.B. Jett. USDA Forest Service, Washington DC, pp 69--77.
Charpentier, J.P. and M. Bonnet-Masimbert. 1983. Influence d’une réhydration préalable sur la germination in vitro du pollen de Douglas (Pseudotsuga menziesii) après conservation. Ann. Sci. For. 40:309--317.
Ching, T.M. and K.K. Ching. 1976. Rapid viability tests and aging study of some coniferous pollen. Can. J. For. Res. 6:516--522. Colangeli, A.M. and J.N. Owens. 1989. Post-dormancy seed-cone
development and the pollination mechanism in western hemlock (Tsuga heterophylla). Can. J. For. Res. 19:44--53.
Colangeli, A.M. and J.N. Owens. 1991. Effects of accelerated pollen-cone development on pollen cytology and fertilizing potential in western hemlock (Tsuga heterophylla). For. Ecol. Manage. 40:151--162.
Copes, D.L. 1985. Fertility of Douglas-fir pollen after one year storage in liquid nitrogen. For. Sci. 31:569--574.
Copes, D.L. 1987. Long-term storage of Douglas-fir pollens. For. Sci. 33:244--246.
Copes, D.L., N.C. Vance, W.K. Randall, A. Jasumback and R. Hall-man. 1991. Vacuum collection of Douglas-fir pollen for supplemen-tal mass pollinations. USDA Forest Service, Res. Note PNW-RN-503, 8 p.
Daniels, J.D. 1978. Efficacy of supplemental mass-pollination in a Douglas-fir seed orchard. Silvae Genet. 27:52--58.
Ebell, L.F. 1972. Cone-induction response of Douglas-fir to form of nitrogen fertilizer and time of application. Can. J. For. Res. 2:317--326. Ebell, L.F. and R.L. Schmidt. 1964. Meteorological factors affecting conifer pollen dispersal on Vancouver Island. Canadian Dept. of Forests, Publ. No. 1036, 28 p.
El-Kassaby, Y.A. and K. Ritland. 1986a. Low level of pollen contami-nation in a Douglas-fir seed orchard as detected by allozyme mark-ers. Silvae Genet. 35:224--229.
El-Kassaby, Y.A. and K. Ritland. 1986b. The relation of outcrossing and contamination to reproductive phenology and supplemental mass pollination in a Douglas-fir seed orchard. Silvae Genet. 35:240--244.
El-Kassaby, Y.A. and S. Reynolds. 1990. Reproductive phenology, parental balance, and supplemental mass pollination in a Sitka spruce seed-orchard. For. Ecol. Manage. 31:45--54.
El-Kassaby, Y.A., R. Davidson and J.E. Webber. 1986. Genetics of seed orchards: a Douglas-fir case study. In Proc. IUFRO Conf. on Breeding Theory, Progeny Testing, and Seed Orchards. Wil-liamsburg, VA, USA, pp 440--456.
El-Kassaby, Y.A., S. Barnes, C. Cook and D.A. MacLeod. 1993. Supplemental mass pollination success rate in a mature Douglas-fir seed orchard. Can. J. For. Res. 23:1096--1099.
Eriksson, G. 1970. Low-temperature induced irregularities in pollen mother cells of Larix leptolepis. Stud. For. Suec. 85:1--14. Eriksson, G., I. Ekberg and A. Jonsson. 1970a. Further studies on
meiosis and pollen formation in Larix. Stud. For. Suec. 63:1--131. Eriksson, G., I. Ekberg and A. Jonsson. 1970b. Meiotic investigations
in pollen mother cells of Norway spruce cultivated in a plastic house. Hereditas 66:1--20.
Eriksson, G., A. Jonsson and D. Lindgren. 1973. Flowering in a clonal trial of Picea abies (L.) Karst. Stud. For. Suec. 110:1--45. Eriksson, U., R. Yazandi, L. Wilhelmsson and Ö. Danell. 1994.
Suc-cess rate of supplemental mass pollination in a seed orchard of
Pinus sylvestris L. Scand. J. For. Res. 9:60--67.
Eriksson, U., G. Jansson, R. Yazdani and L. Wilhelmsson. 1995. Effects of supplemental mass pollination (SMP) in a young and a mature seed orchard of Pinus sylvestris. Tree Physiol. 15:519--526. Fashler, A.M.K. and Y.A. El-Kassaby. 1987. The effect of spray cool-ing treatment on reproductive phenology in a Douglas-fir seed orchard. Silvae Genet. 36:245--249.
Feilberg, L. and B. Søegaard. 1975. Historical review of seed orchards.
In Seed Orchards. Ed. R. Faulkner. For. Comm. Bull. 54:1--8.
Fowler, D.P. 1987. In defense of the polycross. Can. J. For. Res. 17:1624--1627.
Fowler, D.P. and A.E. Wiselogel. 1993. Pollen polymixes. In Advances in Pollen Management. USDA Handbook 698. Eds. D.L. Bramlett, G.R. Askew, T.D. Blush, F.E. Bridgwater and J.B. Jett. USDA Forest Service, Washington DC, pp 79--82.
Hadders, G. 1984. Supplemental and controlled mass pollination in seed orchards of Scots pine (Pinus sylvestris L.). In Foreningen skogstradsforadling. Institutet for skogsforbattring, Arsbok, pp 47--64.
Heslop-Harrison, J. 1987. Pollen germination and pollen-tube growth. Int. Rev. Cytol. 107:1--78.
Jett, J.B. and L.J. Frampton Jr. 1990. Effect of rehydration on in vitro germination of loblolly pine pollen. South. J. Appl. For. 14:48--51. Jett, J.B., D.L. Bramlett, J.E. Webber and U. Eriksson. 1993. Pollen collection, storage and testing. In Advances in Pollen Management. USDA Handbook 698. Eds. D.L. Bramlett, G.R. Askew, T.D. Blush, F.E. Bridgwater and J.B. Jett. USDA Forest Service, Wash-ington DC, pp 41--46.
Jonsson, A. 1974. A study on the temperature response of pollen mother cells in Norway spruce. Stud. For. Suec. 116:1--32. Lambeth, C.C. 1993. Overview of pollen management in tree
breed-ing. In Advances in Pollen Management. USDA Handbook 698. Eds. D.L. Bramlett, G.R. Askew, T.D. Blush, F.E. Bridgwater and J.B. Jett. USDA Forest Service, Washington DC, pp 97--99. Masters, C.J. 1982. Weyerhaeuser’s seed orchard program. In Proc.
18th Can. Tree Improve. Assoc. Meeting, Part 2. Eds. D.F.W. Pollard, D.G.W. Edwards and C.W. Yeatman. Canadian Forest Service, Ottawa, Ontario, Canada, pp 60--70.
Moody, W.R. and J.B. Jett. 1990. Effects of pollen viability and vigor on seedproduction of loblolly pine. South. J. Appl. For. 14:33--38. Moran, G.F. and A.R. Griffin. 1985. Non-random contribution of
pollen in polycrosses of Pinus radiata D. Don. Silvae Genet. 34:117--121.
Ottaviano, E. and D. Mulcahy. 1989. Genetics of angiosperm pollen. Adv. Genet. 26:1--64.
Owens, J.N. 1993. Pollination biology. In Advances in Pollen Manage-ment. USDA Handbook 698. Eds. D.L. Bramlett, G.R. Askew, T.D. Blush, F.E. Bridgwater and J.B. Jett. USDA Forest Service, Wash-ington DC, pp 91--13.
Owens, J.N. and M.D. Blake. 1985. Forest tree seed production. Canadian Forest Service, Information Rep. Pl-X-53, 161 p. Owens, J.N., S.J. Simpson and M. Molder. 1981. The pollination
mechanism and the optimal time of pollination in Douglas-fir (Pseudotsuga menziesii). Can. J. For. Res. 11:36--50.
Philippe, G. and P. Baldet. 1992. Mechanized pollen harvesting with a view to hybrid larch seed production. Ann. Sci. For. 49:297--303. Ross, S.D. 1988. Pre- and post-pollination polyhouse environment
effects on pollen and seed development in potted Picea engelmanii grafts. Can. J. For. Res. 18:623--627.
Ross, S.D., J.E. Webber, R.P. Pharis and J.N. Owens. 1985. Interaction between gibberellin A4/7 and root-pruning on the reproductive and vegetative process in Douglas-fir. I. Effects on flowering. Can. J. For. Res. 15:341--347.
Sarvas, R. 1968. Investigation on the flowering and seed crop of Picea
abies. Commun. Inst. For. Fenn. 67(5):1--84.
Schoen, D.J. and W.M. Cheliak. 1987. Genetics of the polycross. 2. Male fertility variation in Norway spruce, Picea abies (L.) Karst. Theor. Appl. Genet. 74:554--559.
Snow, A. and T.P. Spira. 1991. Pollen vigour and the potential for sexual selection in plants. Nature 352:796--797
Spraque, J.R. and E.B. Synder. 1981. Extracting and drying pine pollen. In Pollen Management Handbook. Agriculture Handbook 587. Ed. E.C Franklin. USDA Forest Service, Washington DC, pp 33--36.
Stanley, R.G. and H.F. Linskens. 1974. Pollen: biology, biochemistry and management. Springer-Verlag, New York, pp 66--86. Webber, J.E. 1987. Increasing seed yield and genetic efficiency in
Douglas-fir seed orchards through pollen management. For. Ecol. Manage. 19:209--218.
Webber, J.E. 1991. Interior spruce pollen management manual. B.C. Ministry of Forests, Land Manage. Rep. 70, 25 p.
Webber, J.E. and F.C.H. Yeh. 1987. Test of the first-on, first-in polli-nation hypothesis in coastal Douglas-fir. Can. J. For. Res. 17:63--68.
Webber, J.E. and M. Bonnet-Masimbert. 1993. The response of dehy-drated Douglas-fir (Pseudotsuga menziesii) pollen to three in vitro viability assays and their relationship to actual fertility. Ann. Sci. For. 50:1--22.
Webber, J.E. and R.A. Painter. 1995. Douglas-fir pollen management manual. 2nd Edn. Res. Branch, B.C. Ministry of Forests, Victoria, B.C., Canada, Work. Paper 02/1995. In press.
Webber, J.E. and S.D. Ross. 1995. Flower induction and pollen viabil-ity for western larch. In Proc. IUFRO Symp. on the Ecol. and Manage. of Larix Forests. Eds. W.C. Schmidt and K.J. McDonald. USDA Forest Service, Whitefish, MT, USA. In press.
Wheeler, N.C. and K.S. Jech. 1986. Estimating supplemental mass pollination (SMP) success electrophoretically. In Proc. 20th Can. Tree Improve. Assoc. Meeting. Eds. F. Caron, A.G. Corriveau and T.J.B. Boyle. Canadian Forest Service, Quebec City, Quebec, Can-ada, pp 111--120.
Wheeler, N.C. and K.S. Jech. 1988. Supplemental mass pollination (SMP) in Douglas-fir seed orchards: biological efficacy and eco-nomic evaluation. Weyerhaeuser Internal Tech. Rep. 050-3210/25, 26 p.
Wheeler, N.C., C.J. Masters, S.C. Cade, S.D. Ross, J.W. Keeley and L.Y. Hsin. 1985. Girdling: an effective and practical treatment for enhancing seed yields in Douglas-fir seed orchards. Can. J. For. Res. 15:505--510.
Woessner, R.A. and E.C. Franklin. 1973. Continued reliance on wind-pollinated southern pine seed orchards----is it reasonable? In Proc. 12th South. For. Tree Improve. Conf. Louisiana State University, Baton Rouge, LA, USA, and USDA Southern Forest Experimental Station, pp 64--73.
Yazdani, R., G. Hadders and A.E. Szmidt. 1986. Supplemental mass pollination in a seed orchard of Pinus sylvestris L. investigated by isozyme analyses. Scand. J. For. Res. 1:309--315.