6 1.1Virus Influenza Tipe A
Virus influenza tipe A adalah virus RNA, famili Orthomyxoviridae dari genus Orthomyxovirus yang menyebabkan penyakit avian influenza. Virus ini merupakan virus yang mempunyai envelop, berpolaritas negatif, dan genomnya bersegmen. Masing-masing segmen dari virus influenza tipe A terdiri delapan segmen gen yang berbeda dan mengkode sekitar 10 protein virus yang berbeda (Palese dan Shaw, 2007).
Struktur protein dalam virion saat ini dapat dibagi kedalam protein permukaan yang meliputi Hemaglutinin (HA), Neuraminidase (NA) dan protein ion chanel membran (M2), sedangkan protein dalam, termasuk nukleoprotein (NP), protein matriks (M1), dan kompleks polimerase terdiri dari polimerase protein dasar 1 (PB1), polimerase protein dasar 2 (PB2), dan polimerase protein asam (PA) (Palese dan Shaw, 2007). Dua protein tambahan yang dihasilkan oleh virus AI adalah protein nonstruktural, protein nonstruktural 1 (NS1) dan protein nonstruktural 2 (NS2), yang juga dikenal sebagai nuclear export protein (NEP) (O’Neill et al., 1998).
Protein nonstruktural 1 (NS1) dianggap ada meskipun protein nonstruktural itu tidak ditemukan dalam partikel virus tetapi di produksi dalam jumlah besar di sel host (Birch-Machin et al., 1997; dan Subbarao et al., 1998). Protein nonstruktural 2 (NS2) terutama ditemukan dalam sel inang, tetapi
beberapa protein dapat ditemukan dalam virion (Palese dan Shaw, 2007). Salah satu protein yang tidak ditemukan di semua virus influenza tipe A adalah protein PB1-F2, yang merupakan protein 87-amino acid yang di transkripsi dari kerangka pembacaan yang berbeda dari protein PB1. Protein ini diduga terlibat dalam apoptosis dalam sel inang dan berperan dalam menentukan pathogenesis virus (Chen et al., 2001).
Virus influenza tipe A memiliki bermacam-macam subtipe, adanya variasi beberapa subtipe berdasarkan kombinasi dua antigen permukaan utama yaitu Hemaglutinin (HA) dan Neuraminidase (NA). Saat ini ada 17 subtipe HA (H1-H17) dan 10 subtipe NA (N1-N10) yang diketahui setelah penemuan terbaru dari genom virus influenza tipe A, subtipe baru ini diidentifikasi dari kelalawar, H17N10 (Tong S, et al., 2012). Daerah eksternal Hemaglutinin (HA) terdiri dari oligosakharida yang menyalurkan derivate neuraminik yang memainkan peran penting dalam masuknya virus serta berfusinya virus, dan Neuraminidase (NA) berperan dalam pelepasan virus (Liu et al., 1995; dan Russell et al., 2008).
Penyusunan virion virus AI di perankan oleh protein matriks yang mana protein M2 bersama dengan protein HA dan NA berfungsi sebagai saluran ion dan untuk menyusun struktur amplop virus, sedangkan protein M1 berfungsi dalam pemisahan protein M1 dari RNP untuk masuk ke sitoplasma sel tropisma (Reid et al., 2002). PB1, PB2, PA berfungsi untuk membentuk polimerase RNA-RNA aktif yang bertanggung jawab untuk replikasi dan transkripsi. Pengaturan dalam mendorong sintesis komponen-komponen virus dalam sel yang terinfeksi diperankan oleh protein NS1 dan NS2 (Gurtler, 2006).
Virus AI memiliki dua mekanisme utama untuk memberikan keragaman dalam populasi virus, mekanisme tersebut dapat berupa antigenic drift dan antigenic shift. Pada antigenic drift mengacu pada perubahan kecil (seperti subtitusi asam amino pada HA dan atau NA) yang mengakibatkan perubahan tempat antigenik. Sebaliknya, antigenic shift adalah bertanggung jawab dalam pembentukan virus subtipe baru dengan kombinasi dari HA dan NA dari subtipe yang berbeda. Seleksi dalam lingkungan memainkan peran yang sangat penting dalam terjadinya perubahan antigenik pada antigen bagian HA, seperti pada tempat-tempat yang mengalami perubahan adaptasi dan pada tempat antigenik yang mengalami subtitusi, sehingga terjadilah perubahan dalam antigenisitas virus, bahwa proses ini merupakan sisi pathogen untuk menghindari dari pertahanan host melalui co-evolusi dengan host (Blackburne et al., 2008).
1.2Virus Avian Influenza Subtipe H5N1
Virus AI subtipe H5N1 merupakan penyakit hewan menular yang bersifat zoonosis. Virus AI subtipe H5N1 merupakan virus yang dapat menyebabkan penyakit influenza pada hewan piaraan jenis unggas dan mamalia seperti kuda, babi, dan manusia (Boyce et al., 2008).
Pada tahun 1997 dunia dikejutkan dengan wabah HPAI di Hong Kong yang disebabkan oleh virus AI subtipe H5N1, dan menular pada manusia dengan enam kasus meninggal (Suarez et al., 1998). Kejadian tersebut menyebabkan virus AI subtipe H5N1 menjadi perhatian utama kematian pada manusia sangat dikhawatirkan menjadi wabah pandemik yang berbahaya bagi umat manusia di
belahan bumi ini. Beberapa tahun terakhir perhatian dunia kesehatan hewan dan manusia terpusat kepada semakin mewabahnya penularan virus AI subtipe H5N1 yang telah menyebabkan kerugian perekonomian yang cukup besar baik pada industri peternakan unggas di dunia maupun di Indonesia pada khususnya. Selain itu, jumlah kasus kematian infeksi AI subtipe H5N1 pada manusia bertambah setiap tahun dan sampai saat ini, AI masih menjadi masalah yang serius yang perlu mendapatkan perhatian yang khusus mengingat korban meninggal akibat infeksi virus ini terus bertambah (WHO, 2012).
Virus AI subtipe H5N1 telah mengalami beberapa kali mutasi yang menyebabkan terjadinya perbedaan pathogenesis virus. Analisis genetik menunjukkan bahwa sebagian besar virus AI subtipe H5N1 dari unggas dan manusia di Asia termasuk dalam genotip Z, serupa dengan virus yang pertamakali diidentifikasi pada unggas di China Selatan (Guan et al., 2004). Virus AI subtipe H5N1 kemungkinan dapat menyebabkan beberapa pandemik influenza karena sifat virus yang mudah mengalami mutasi. Mutasi dari virus AI subtipe H5N1 kemungkinan besar dapat menghasilkan varian virus AI subtipe H5N1 baru yang dapat mengenali reseptor spesifik yang ada pada sel manusia (natural human α 2-6 glycan), sehingga jikalau hal ini terjadi, maka penularan virus AI subtipe H5N1 dari manusia ke manusia dapat terjadi dengan mudah dan dengan adanya kemungkinana tersebut virus AI subtipe H5N1 dapat mengancam kondisi global kesehatan manusia (Stevens et al., 2006).
1.3Pathogenesis
Virus AI memiliki dua tingkat pathogenitas berdasarkan genetic sequance dan kemampuan menimbulkan penyakit pada unggas yakni HPAI dan LPAI. Virus AI pathogenitas tinggi (HPAI) termasuk virus yang mampu menyebar dengan cepat dan dapat menyebabkan penyakit yang serius dengan tingkat kematian yang tinggi, sedangkan virus avian influenza unggas berpathogenitas rendah (LPAI) menyebabkan penyakit yang ringan yang kemungkinan besar tidak menunjukkan gejala jika terjadi infeksi pada unggas (Perdue dan Swayne, 2005).
Virus AI dapat ditularkan melalui berbagai cara sehingga dapat masuk ke tubuh host, tanpa adanya gejala klinis ataupun dengan menimbulkan gejala klinis bahkan terjadi kematian pada host yang terinfeksi. Saat virus menginfeksi host dan masuk kedalam sel host, maka akan terjadi pengkopian dari RNA berpolaritas negatif kedalam penyandian mRNA, dan proses ini adalah bagian terpenting dari segi siklus hidup virus. Ketika virus endositosis kedalam sel rentan, segmen RNA dilepaskan kedalam sel untuk memulai infeksi, dengan proses itu menyebabkan penurunana pH dalam vesikel endosit dan penggabungan membran virus dengan membran vesikel (Lamb et al., 2001; Vaccaro et al., 2005).
Untuk membuat virus baru, RNA virus dilepas sebagai kompleks Nukleoprotein-RNA (NP), kemudian bersama protein lain menuju nukleus. Tiga protein polimerase besar (hampir 50 % dari total informasi genetik) mulai berfungsi setelah RNA virus mencapai inti, dengan membuat RNA positif atau mRNA maupun dari template untuk membuat molekul RNA antisense baru yang
akan di kemas dalam pembentukan partikel virus yang baru. Menurut Noda et al. (2006), untuk mendapatkan titer virus yang tinggi dalam waktu singkat kompleks NP-RNA merupakan kunci utama.
Fungsi gen lainya adalah membantu mengatur pembuatan dan perakitan dari partikel virus setelah memulai proses dari produksi mRNA. Sintesis protein struktural virus ditentukan oleh empat gen yakni dua gen penyandi protein permukaan (HA dan NA), serta dua gen penyandi protein dalam (M1dan NP). Tiga kategori utama dari gen dalam virus avian influenza dapat diklasifikasikan secara luas sebagai (1) gen sintesis RNA, (2) gen regulatory (pengaturan), dan (3) gen protein struktural. Kategori ini penting sebagai penentu variasi genetik untuk pathogenesis dari virus avian influenza. Yang paling penting adalah jenis variabilitas genetik yang terjadi pada berbagai gen yang mempengaruhi berbagai tahapan siklus hidup virus serta berpengaruh pada pathogenesitas virus (Swyne, 2008).
2.4 Avian Influenza Pada Ayam
AI dilaporkan pertamakali sebagai kejadian yang sangat mematikan dan bersifat sistemik pada ayam di Itali pada tahun 1878, yaitu HPAI (Perroncito, 1878). AI merupakan penyakit sistemik hebat yang sering disebut dengan istilah fowl plague atau fowl pest, akan tetapi nama lain juga telah digunakan, termasuk peste aviaire, Geflugelpest, tyhpus exudatious gallinarium, Brunswick bird plaque, fowl atau bird grippe (Stubbs, 1926).
Di Indonesia kasus AI terjadi pertamakali pada ayam ras komersial, kasus ini menyebabakan kematian pada banyak ayam. Di Indonesia AI pertamakali dilaporkan telah terjadi di Jawa Barat dan Jawa Tengah, kemudian secara cepat menyebar ke berbagai daerah di Jawa, Kalimantan, Sumatra, Lampung, dan Bali (Kandun, 2006; Mahardika et al., 2004; Smith et al., 2006).
Virus AI merupakan virus yang sangat berbahaya, bersifat zoonosis dan mematikan, baik pada unggas (ayam) ataupun pada manusia. Virus ini telah dilaporkan dapat menginfeksi manusia dan ditransmisikan secara langsung dari unggas (ayam) ke manusia di Hongkong, dan menyebabkan enam orang meninggal dari 18 orang yang terinfeksi virus tersebut (Claas et al., 1998; Shortridge et al., 1998; Subbaro et al., 1998; Katz et al., 2000). Pada ayam, virus AI menyebabkan gejala klinis yang sangat bervariasi, mulai dari infeksi yang bersifat asimptomatik sampai infeksi yang berakibat fatal dan bersifat multisistemik (Swayne dan Suarez, 2000).
2.5 Avian Influenza Pada Unggas Air (Itik)
Unggas air dapat terinfeksi oleh semua subtipe virus influenza tipe A, dengan sedikit atau tanpa menunjukkan gejala (Woebser, 1997). Unggas air, salah satunya itik memainkan peranan penting dalam pemeliharaan dan penyebaran virus avian influenza yang termasuk golongan LPAI (Olsen et al., 2006), tetapi penelitian eksperimental menunjukkan bahwa itik mungkin memainkan peran dalam pemeliharaan virus HPAI subtipe H5N1. Itik yang terinfeksi dapat tidak menunjukkan tanda-tanda klinis namun dapat mengekskresikan virus dengan
konsentrasi tinggi yang bersifat pathogen pada spesies unggas lainya (Hulse-Post et al., 2005).
Replikasi virus AI pada itik terjadi terutama pada saluran pencernaan (usus), dengan konsentrasi virus low pathogenic dalam feses (Webster et al., 2002). Akibatnya, terjadilah transmisi lokal LPAI pada unggas air yang tergantung pada transmisi melalui tinja (Hinshaw et al., 1979,1980), sedangkan penyebaran secara geografis dari virus AI sebagian besar tergantung pada pola dari pelepasan virus dan prilaku migrasi unggas yang terinfeksi (Lebarbenchon et al., 2009).
Burung liar juga merupakan reservoir alami dari virus AI, kemungkinan besar spesies burung liar menumpahkan sebagian besar virus HPAI subtipe H5N1 tanpa menunjukkan gejala klinis (Tumpey et al., 2012), kejadian tersebut memungkinkan dapat menambah tingkat terjadinya transmisi penyebaran virus AI subtipe H5N1 pada unggas air dan menyebabkan ketidakseimbangan diantara virus influenza A dengan unggas air (itik) (Sturm-Ramirez, 2005).
Ketidakseimbangan antara unggas air (itik) dengan virus AI subtipe H5N1 memungkinkan dapat menyebabkan terjadinya perubahan virulensi virus AI subtipe H5N1 karena sifat virus AI yang mudah mengalami mutasi. Mutasi, termasuk subtitusi, delesi, dan insersi adalah salah satu mekanisme paling penting dalam menghasilkan variasi virus AI (Holland et al., 1982).
2.6 Imunohistokimia (IHK)
IHK telah muncul sebagai suatu teknik penelusuran kuat yang dapat memberikan informasi tambahan untuk penilaian rutin morfologi jaringan. IHK digunakan untuk mempelajari penanda seluler yang menentukan fenotipe tertentu yang dapat memberikan suatu kepentingan diagnosis, prognosis, dan prediksi sebuah informasi yang berkaitan tentang status dan biologis penyakit. Penggunaan antibodi untuk kajian molekuler dan patologi jaringan dapat di sempurnakan dengan teknik IHK (Key, 2009).
Menurut Snider, (2013) pendekatan dalam deteksi antigen-antibodi sebagai dasar dari IHK terbagi menjadi dua, yaitu dengan deteksi langsung dan tidak langsung, dan dalam pemilihan pendekatan yang digunakan tergantung pada tingkat target ekspresi antigen. Dalam deteksi antibodi secara langsung, antibodi terhadap antigen target (antibodi primer) adalah penghubung dengan sebuah enzim. Enzim yang paling sering digunakan adalah Horseradish Peroxidase (HRP) atau Alkaline Phosphatase (AP). Kerja enzim ini yang akan diaktifkan dengan penambahan substrat. Pada deteksi langsung tidak memerlukan langkah tambahan untuk penambahan antibodi sekunder. Kelemahan teknik ini kemungkinan sulit untuk dideteksi jika protein yang ditemukan dalam jumlah sedikit.
Pada metode IHK tidak langsung, deteksi menggunakan antibodi skunder yang bereaksi dengan antibodi primer berlabel untuk memperkuat sinyal, dengan demikian sensitifitas unji meningkat, antibodi skunder lazimnya berlabel enzim HRP/AP kemudian direaksikan dengan subtrat untuk menghasilkan kromogenik
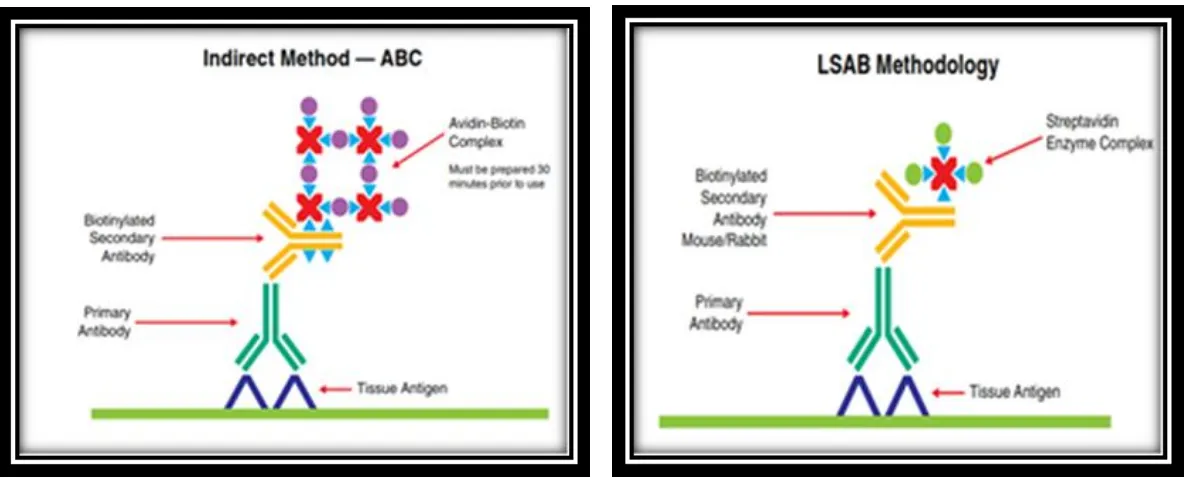
(Snider, 2013). Metode IHK tidak langsung dapat dilakukan dengan beberapa cara misalnya: Avidin-Biotin Compleks (ABC), Labeled Streptavidin Biotin (LSAB), dan Phosphatase-Anti-Phospatase (PAP) (Boenisch, 2001).
Pewarnaan IHK dengan metode avidin-biotin-peroksidase kompleks (ABC) yang melibatkan penggunaan antibodi skunder berlabel biotin diikuti oleh penambahan dari avidin-biotin-peroksidase kompleks, memberikan hasil lebih baik ketika di bandingkan dengan metode antibodi yang tidak berlabel. Terdapatnya tempat ikatan dari biotin secara kompleks dihasilkan karena avidin dengan peroksidase berlabel biotin (Boenisch, 2001).
Dalam pembentukan kompleks, avidin berfungsi sebagai jembatan antara molekul peroksidase berlabel biotin, molekul peroksidase berlabel biotin, yang mengandung beberapa gugus biotin, berfungsi sebagai penghubung antara molekul avidin. Akibatnya, sebuah pola-pola kompleks yang mengandung beberapa molekul peroksidase kemungkinan terbentuk. Ikatan dari kompleks ini pada gugus biotin berhubungan dengan antibodi sekunder hasil atas pewarnaan dengan intensitas yang tinggi (Hsu et al., 1981).
Dalam metode yang sama, metode LSAB juga menggunakan antibodi sekunder biotinylated. Dimana antibodi skunder akan menghubungkan antibodi primer untuk menghubungkan streptavidin-peroksidase, akan tetapi kompleks yang terbentuk lebih kecil dari pada kompleks yang terbentuk oleh metode ABC, dan itu sulit dijangkau epitop, jadi metode LSAB memiliki tingkat sensitifitas lebih baik dari pada metode ABC (Chilosi et al., 1994). Kedua metode tersebut
dilihat perbedaanya berdasarkan ikatan dari avidin-biotin kompleks dan streptavidin-biotin (Gambar 2.1).
Gambar 2.1 A) Metode Avidin-Biotin Complex (ABC), B) Metode Labeled Streptavidin Biotin (LSAB). Sumber: Handbook IHC Staining Method, DAKO (2009).
Avidin adalah glikoprotein dan memiliki titik isoelektrik (pl) berkisar 10, ini memiliki kecenderungan untuk mengikat secara khusus seperti lektin dan komponen jaringan yang bermuatan negatif pada pH fisiologis. Streptavidin memiliki titik isoelektrik yang lebih netral dan tidak memiliki gugus karbohidrat. Perbedaan-perbedaan ini mengakibatkan kurang spesifik dalam pengikatan jaringan. Molekul biotin adalah konjugasi yang mudah untuk antibodi dan enzim. Dalam metode ABC antibodi sekunder adalah penghubung untuk biotin dan berfungsi sebagai penghubung diantara antibodi primer jaringan yang terikat dan sebuah avidin-biotin-peroksidase kompleks (Heras et al., 1995).
Pada teknik imunohistokimia antibodi yang biasanya digunakan adalah antibodi monoklonal, dimana antibodi monoklonal adalah antibodi monospesifik yang dapat mengikat satu epitope saja. Bahwasanya antibodi monoklonal memiliki antigen binding surface yang sama (Sudiana, 2005).