EKOLOGI TERNATE
EDITOR
Ibnu Maryanto
Hari Sutrisno
PUSAT PENELITIAN BIOLOGI-LIPI
2011
ii
© 2011 Lembaga Ilmu Pengetahuan Indonesia (LIPI) Pusat Penelitian Biologi*
Katalog dalam Terbitan
Ekologi Ternate/Ibnu Maryanto dan Hari Sutrisno (Editor). – Jakarta: LIPI Press, 2011.
xiii + 371 hlm.; 14,8 x 21 cm
ISBN 978-979-799-609-3
1. Ekologi 2. Ternate
577
Editor Bahasa : Risma Wahyu Hartiningsih Penata Letak : Ibnu Maryanto
Penata Sampul : Fahmi Penerbit : LIPI Press
*Pusat Penelitian Biologi-LIPI
Gedung Widyasatwaloka, Cibinong Science Center Jln. Raya Bogor Km. 46, Cibinong 169111
Telp.: 021-8765056, 8765057
DAFTAR ISI
Ucapan Terimakasih iii
Kata Sambutan v
Kata Pengantar vii
DAFTAR ISI xi
GEOLOGI DAN IKLIM
Gunung Gamalama, Ternate, Maluku Utara: Dinamika Erupsi dan Potensi
Ancaman Bahayanya 1
Indyo Pratomo, Cecep Sulaeman, Estu Kriswati & Yasa Suparman Karakteristik Erupsi G Kie Besi dan Potensi Ancaman Bencananya Terhadap Lingkungan Kota Ternate: (Representasi dari karakter
gunungapi aktif di Busur Gunungapi Halmahera) 15 Estu Kriswati & Indyo Pratomo
Analisa Anomali Curah Hujan dan Parameter Laut-Atmosfer Periode
Januari - Agustus 2010 di Provinsi Maluku Utara 27 Dodo Gunawan
FAUNA
Kelimpahan dan Keragaman Kelelawar (Chiroptera) dan Mamalia Kecil
di Pulau Ternate 43
Sigit Wiantoro & Anang S Achmadi
Keanekaragaman Mamalia Kecil di Pulau Moti 55
Anang Setiawan Achmadi & Sigit Wiantoro
Kajian Ekologi Burung di Hutan Gunung Gamalama, Ternate, Maluku
Utara 69 Wahyu Widodo
Komunitas Burung Pulau Moti Ternate Maluku Utara 83 Eko Sulistyadi
Keanekaragaman Herpetofauna di Pulau Ternate dan Moti, Maluku Utara 105 Mumpuni
xii
Komunitas Keong Darat di Pulau Moti, Maluku Utara 121 Heryanto
Kajian keanekaragaman Ngengat (Insekta: Lepidoptera) di Gunung
Gamalama, Ternate 133
Hari Sutrisno
Tinjauan Keanekaragaman dan Sebaran Kupu Ternate 145 Djunijanti Peggie
Efektifitas Trap Warna Terhadap Keberadaan Serangga Pada Pertanaman
Budidaya Cabai di Kelurahan Sulamadaha Kecamatan P Ternate Ternate 159 Abdu Mas’ud
Eksplorasi Keragaman Serangga Coleoptera dan Lepidoptera di Pulau
Moti, Ternate, Maluku Utara 167
Warsito Tantowijoyo & Giyanto FLORA
Analisis Tutupan Lahan Kawasan Pulau Moti, Ternate, Maluku Utara 187 Hetty IP Utaminingrum & Roemantyo
Hutan mangrove di Pulau Moti 199
Suhardjono & Ujang Hapid
Keanekaragaman Anggrek di G Gamalama, Ternate 219 Izu Andry Fijridiyanto & Sri Hartini
Vegetasi Hutan Pulau Moti, Ternate, Maluku Utara 227 Edi Mirmanto
Keanekaragaman Jenis Pohon di Hutan Sekunder Pulau Moti,
Ternate-Maluku Utara 237
Razali Yusuf
Keanekaragaman Tumbuhan Berkhasiat Obat di Pulau Moti, Ternate,
Maluku Utara 251
Siti Sunarti
Eksplorasi Tumbuhan di Pulau Moti, Ternate, Maluku Utara 267 Deden Girmansyah & Siti Sunarti
MIKROBIOLOGI
Drug Discovery Antibiotik Berbasis Biodiversitas Aktinomisetes Lokal
Asal Ternate 283
Arif Nurkanto
Isolasi dan Identifikasi Kapang-Kapang Kontaminan Dari Biji Kenari
Kering (Canarium ovatum) 295
Nurhasanah &Sundari
Mikroba Laut Penghidrolisis Senyawa Nitril di Sekitar Pulau Moti,
Ternate 301
Nunik Sulistinah & Rini Riffiani
Isolasi dan Penapisan Bakteri Pendegradasi Dibenzothiophene,
Phenanthrene dan Fluoranthene Asal Perairan Laut Sekitar Pulau
Moti-Ternate 309 Rini Riffiani & Nunik Sulistinah
Penapisan dan Isolasi Bacillus Penghasil Amilase Dari Limbah Sagu
(Metroxylon sagu Rottb) 317
Deasy Liestianty1, Nurhasanah2 SOSIAL BUDAYA
Membangun Ternate Bermodal Kekayaan Sosio-Historis 329 Dhurorudin Mashad
Analisis Struktural Terhadap Mitos “Tujuh Putri” Pada Kebudayaan
Ternate, Maluku Utara 343
Safrudin Amin
301 Ekologi Ternate 301-308 (2011)
Mikroba Laut Penghidrolisis Senyawa Nitril di Sekitar Pulau Moti,
Ternate
Nunik Sulistinah & Rini Riffiani
Bidang Mikrobiologi, Pusat Penelitian Biologi Cibinong Science Center Jl. Raya Jakarta –Bogor Km 46, Cibinong, E-mail : [email protected]
ABSTRACT
Nitrile compounds are distributed widely in the environment as a result of anthropogenic activity or industrial synthesis. Nitrile-hydrolyzing microorganisms play an important role to degrade the compound. Microorganisms usually convert nitriles through two hydrolytic pathway. The potensial biotechnological application of the nitrile-hydrolysing enzymes are becoming commonplace in the synthesis of commodity chemical, pharmaceuticals, and also useful for bioremediation application. In this experiment 33 isolates of marine bacteria were isolated from sponge in Ternate Island. All isolates were tested for their ability to utilize acetamide (aliphatic amide) and bezamide (aromatic amide) as sole source of carbon and nitrogen. Screening of nitrile-hydrolysing bacteria was done using microplate titter with added Iodonitrotetrazolium. Based on their growth pattern, eleven isolates able to utilize acetamide (100 mM) as the source of carbon and nitrogen, and only six isolates able to grow on benzamide
(25 mM). One of them is Lysobacter sp showed the highest growth on the acetamide. Resting
cell of Lysobacter sp capable of hydrolyzing acetonitrile. Acetic acid and ammonia as the end
product of hydrolysis.
Key words: Nitrile-hydrolysing, enzyme, Ternate
PENDAHULUAN
Nitril dengan rumus molekul RCN merupakan salah satu senyawa toksik yang sulit untuk didegradasi. Sebagian besar senyawa nitril dilaporkan muta-genik dan karsinomuta-genik. Meskipun demikian senyawa ini mempunyai nilai komersial yang tinggi, misalnya nitril sintetik banyak diproduksi dan digunakan secara ektensif di industri-industri sebagai pelarut, pengekstrak, farmaka, dan pestisida (dichlobenil, bromoxynil). Industri kimia juga menggunakan sebagian besar senyawa nitril sebagai
polimer dan sintesa senyawa-senyawa kimia, seperti misalnya akrilonitril dan adiponitril diperlukan untuk produksi poliakrilamida dan polimer nilon 66 (Banerjee et al. 2002). Penggunaan
senyawa nitril yang sangat luas akan berdampak negatif terhadap kesehatan dan lingkungan. Dengan demikian detoksifikasi senyawa nitril di lingkungan perlu mendapat perhatian. Detoksifikasi senyawa nitril dapat dilakukan secara kimia dan biologis.
Hidrolisis nitril secara kimiawi dinilai tidak menguntungkan karena memer-lukan kondisi yang ekstrim dan produk hidrolisis yang dihasilkan tidak ramah
Sulistinah & Riffiani
lingkungan. Sedangkan hidrolisis senya-wa nitril secara mikrobiologis dinilai merupakan cara yang efisien dan aman. Degradasi senyawa nitril secara biologi pada umumnya melibatkan enzim pedegradasi/penghidrolisis nitril (Nitril hidratase, Amidase, dan/atau Nitrilase) yang bisa disintesis oleh mikroba baik bakteri, jamur, maupun yeast (Kobayashi
1991). Pendegra-dasi senyawa nitril
melalui 2 alur reaksi, alur rekasi pertama nitril diubah menjadi senyawa amida dengan melibatkan enzim nitril hidratase, selanjutnya amida yang terbentuk oleh enzim amidase diubah menjadi asam karboksilat dan ammo-nium. Sedangkan alur reaksi kedua senyawa nitril akan diubah langsung menjadi asam karboksilat dan ammonium (Kobayashi 1991; Kobayashi & Shimizu 2000; Podar et al.
2006). Sebagian besar senyawa nitril aromatik melalui alur reaksi 2 (Nawaz
et al. 1992) yaitu hanya melibatkan
nitrilase saja.
Mengingat pentingnya dan aplikasi yang begitu luas dari enzim yang disinte-sis mikroba tersebut maka pencarian mikroba potensial sebagai pendegradasi nitril terus dilakukan dan mendapatkan perhatian karena enzim penghidrolisis nitril yang diproduksi oleh mikroba banyak diaplikasikan sebagai biokatalis untuk sintesa senyawa-senyawa organik ataupun sebagai agent bioremediasi lingkungan tercemar nitril/sianida (Banerjee et al. 2002; Hakansson et al.
2005; Manolov et al. 2005; Kohjama et al. 2006).
Sampai saat ini penelitian tentang biodegradasi/biotransformasi nitril dan pengungkapan potensi mikroba laut dan
terestrial penghasil enzim pendegradasi/ penghidrolisis nitril di Indonesia belum banyak dilakukan. Peluang untuk menda-patkan mikroba tersebut sangat besar karena Indonesia merupakan negara tropis dan sebagian besar kawasannya adalah perairan. Untuk memperoleh mikroba potensial tersebut harus diawali dengan kegiatan eksplorasi mikroba pendegradasi kemudian mengisolasi, seleksi/penapisan, dan selanjutnya mengkarakterisasi mikroba dan enzim yang terlibat dalam proses hidrolisisnya.
BAHAN DAN CARA KERJA Mikroba Penghidrolisis Nitril (RCN)
Isolasi mikroba dilakukan melalui beberapa cara, salah satu diantaranya melalui pengenceran bertingkat yaitu ± 1,0 gram sampel diinokulasikan ke dalam 9 ml larutan NaCl 0,85%, dihomogenkan dan kemudian ±50 μl sampel diinokulasi-kan ke dalam media Marine Agar dan
diinkubasi selama 24-48 jam. Koloni yang
tumbuh diisolasi, dimurnikan dan disimpan dalam media agar miring (slant agar) untuk pengujian lebih lanjut.
Seleksi mikroba untuk memperoleh isolat yang berpotensi sebagai penghidrolisis senyawa nitril (asetonitril) dilakukan dengan menumbuhkan isolat ke dalam
microtitter plate steril yang berisi media
mineral yang mengandung senyawa nitril. Selanjutnya diinkubasi di atas mesin pe-ngocok pada suhu ruang (28 oC) selama
72 jam. Pertumbuhan mikroba diamati dengan adanya kekeruhan pada media tumbuh, untuk memperkuat dugaan terse-but dilakukan uji larutan Iodonitrote-trazolium (INT) sebagai indikator.
303 Mikroba Laut Penghidrolisis Senyawa Nitril
Aktivitas mikroba ditentukan dengan metode Nessler (Oliver et al. 1989).
Kultivasi, Medium Isolasi dan Medium Seleksi.
Media isolasi dan kultivasi mikroba laut digunakan Marine Agar (Difco),
sedangkan media seleksi mikroba penghi-drolisis digunakan media mineral (Sunarko et al. 2007)
Analisis produk degradasi nitril.
Penentuan produk degradasi pada umumnya ditentukan dengan menggu-nakan GC-Shimadzu 14 B , detektor FID, kolom Porapax Q , suhu kolom 225oC,
suhu injektor dan detektor 240 oC
(Sunarko et al. 2000) atau dengan
menggunakan HPLC. Penentuan ammonium dilakukan dengan metode Nessler
Identifikasi dilakukan secara molekuler dengan 16S rDNA.
HASIL
60 isolat bakteri telah diisolasi dari material sponge yang diperoleh dari perairan laut Ternate, 33 isolat diantaranya telah diuji kemampuan tumbuhnya pada senyawa amida (asetamida dan benzamida). Dari hasil pengujian pertumbuhan terhadap ke 33 isolat tersebut pada asetamida dan benzamida dengan teknik microplate
titter dan penambahan indikator INT
serta pengujian aktivitas dengan metode Nessler menunjukkan bahwa 30 % isolat dapat tumbuh pada asetamida, sedang-kan pada benzamida hanya ± 20 %. Dari 30% isolat yang tumbuh tidak semuanya memberikan aktivitas yang baik, demikian juga yang terjadi pada benzamida
0 10 20 30 40 50 60 Aset amida Benz amid a Tida k tum buh P ro sen tase i s o lat ( % )
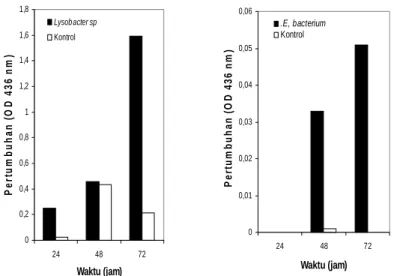
Sulistinah & Riffiani 0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 24 48 72 Waktu (jam) P e rt um buha n ( O D 436 n m ) Lysobacter sp Kontrol 0 0,01 0,02 0,03 0,04 0,05 0,06 24 48 72 Waktu (jam) P e rt um buha n ( O D 4 3 6 nm ) .E, bacterium Kontrol
Gambar 2. Pertumbuhan Lysobacter sp (A) dan Enterobacteriaceae bacterium (B) pada
asetamida 100 mM
(Gambar 1). Prosentase isolat yang tidak tumbuh lebih besar dibandingkan isolat yang tumbuh pada senyawa amida uji.
Identifikasi secara molekuler dilakukan terhadap 6 isolat yang mempuyai kemampuan menghidrolisis senyawa amida. Isolat-isolat tersebut adalah Micrococcus luteus, Lysobacter
sp., Enterobacteriaceae bacterium,
Sphingomonas sp, Sponge bacterium.
Lysobacter sp dan E. bacterium tumbuh
cukup baik pada 100 mM asetamida (Gambar 2A dan 2B). Tampak pada Gambar 2A dan 2B tersebut Lysobacter
sp menunjukkan kemampuan tumbuh dan mempunyai aktivitas yang lebih baik dibandingkan E. bacterium.
Hal ini diperkuat dengan hasil degradasi asetonitril oleh isolat tersebut.
Sel Lysobacter sp yang ditumbuhkan
pada 100 mM asetamida mampu menghidrolisis asetonitril 190 mM dengan
asetat dan ammonium sebagai produk metabolitnya (Gambar 3)
PEMBAHASAN
Berdasarkan hasil pengujian pertumbuhan menggunakan teknik mikroplate tiiter dengan penambahan larutan indilakor INT menunjukkan ternyata hanya sedikit isolat yang mampu tumbuh dengan baik pada kedua senyawa amida tersebut. Isolat yang tumbuh pada asetamida lebih banyak dibandingkan benzamida. Hal ini terlihat pada saat penambahan indikator
Iodonitrotetra-zolium chloride (INT) pada kultur cair
tampak perubahan warna menjadi pink. Warna yang timbul merupakan indikasi adantya pertumbuhan mikroba, semakin pekat warna yang terbentuk pertumbu-han semakin baik (Sunarko et al. 2008)
Dilaporkan beberapa mikroba sulit tumbuh pada benzamida dan /atau
Sulistinah & Riffiani 0 10 20 30 40 50 60 15 60 90 120 180 Waktu (menit) K o n s en tra s i aset o n it ri l, a set at , d a n a m m oni um (m M ) AsCN Asetat Amonium
mempunyai kelarutan yang rendah. Sedangkan asetonitril dan/atau asetamida merupakan senyawa alifatik nitril yang paling sederhana, mudah larut sehingga banyak mikroba yang mampu menggunakan senyawa ini sebagai sumber karbon dan nitrogen untuk substrat tumbuhnya.
Berdasarkan hasil yang diperoleh 1 isolat yaitu Lysobacter sp menunjukkan
pertumbuhan paling baik pada asetamida 100 mM, sedangkan pada benzamida
Lysobacter sp tidak mampu untuk
tumbuh. Dengan demikian Lysobacter
sp. merupakan isolat penghidrolisis asetonitril yang diisolasi dari lingkungan laut. Pengujian degradasi asetonitril dengan menggunakan sel Lysobacter sp
yang ditumbuhkan pada asetamida menunjukkan bahwa isolat tersebut ternyata mampu menghidrolisis asetonitril 190 mM dengan produk hidrolisis berupa asam asetat dan ammonium. Degradasi asetonitril dengan menggunakan sel utuh
(whole cell) tampaknya melalui 2 alur
reaksi yang melibatkan enzim nitril hidratase dan amidase.
Kemampuan enzim penghidrolisis asetonitril dari Lysobacter sp relatif
rendah bila dibandingkan dengan isolat-isolat hasil penelitian yang telah dilakukan, seperti misalnya Rhodococcus
erythro-polis BL1 yang diisolasi dari sedimen laut
mampu tumbuh dan menghidrolisis asetonitril 1M ( Langdahl & Ingvorsen, 1996). Rendahnya aktivitas enzim mungkin disebabkan karena belum dilakukan optimasi terhadap kinerja enzim
yang dihasilkan oleh Lysobacter sp.,
sehingga enzim belum bekerja pada kondisi yang optimal.
Secara umum dapat dikatakan bahwa jumlah mikroba yang diisolasi dari sponge yang berpotensi sebagai penghidrolisis senyawa nitril sangat rendah. Rendahnya isolat berpotensi yang diperoleh dan kemampuan tumbuh isolat mungkin juga disebabkan karena teknik isolasi yang digunakan kurang
307 Mikroba Laut Penghidrolisis Senyawa Nitril
tepat. Pada umumnya untuk memperoleh atau mengisolasi mikroba target (pende-gradasi senyawa toksik) tertentu idealnya dilakukan teknik aklimatisasi yaitu sampel yang diperoleh ditumbuhkan pada media yang mengandung substrat toksik tersebut dan diinkubasi dalam waktu yang relatif panjang kemudian dilakukan isolasi (Matsuda et.al. 2007). Berdasarkan
teknik aklimatisasi ini biasanya diperoleh mikroba target yang diinginkan dengan aktivitas yang relatif cukup tinggi untuk mendegradasi senyawa target.
KESIMPULAN
33 isolat bakteri diisolasi dari sponge yang dikoleksi dari Ternate. 30% dari keseluruhan isolat tersebut mampu tumbuh pada asetamida dan hanya 20% isolat tumbuh pada benzamida. 6 isolat diantaranya telah teridentifikasi sebagai
Arthrobacter sp. WPCB190,
Entero-bacteriaceae bacterium B12,
Lysobac-ter sp , Micrococcus luteus strain
G3-6-08 , Sphingomonas sp, dan Sponge
bacterium IS8. Lysobacter sp
menunjuk-kan kemampuan menghidrolisis asetonitril 190 mM. Asam asetat dan ammonium merupakan produk hidrolisisnya.
UCAPAN TERIMA KASIH
Penulis mengucapakan terima kasih kepada Dr. Ibnu Maryanto selaku koordinator Projek IPTEKDA Khusus 2009 yang telah mendanai kegiatan penelitian ini.
DAFTAR PUSTAKA
Banerjee, A., R. Sharma, UC. Banerjee. 2002. The nitrile degrading enzymes: current status and future
prospect. Appl. Microbiol.
Biotechnol., 60 : 33-34
Hakansson, K., U. Welander & B. Mattiason. 2005. Degradation of acetonitrile through a sequenceof
microbial reactors. Water Res. 39
: 648-654
Kobayashi, M. 1991. Studies on Enzymes Involved in Nitrile Metabolism in
Rhodococcus rhodochrous. PhD
Thesis.
Kobayashi & Shimizu. 2000. Nitrile
hydrolases. Curr. Opin Chem.
Biol. 4 : 95-102
Kohyama, L, A. Yoshimura, D. Aoshima, T. Yoshido, H. Kawamoto & Nagasawa. 2006. Convenion treat-ment of acetonitrile-containing wastes using the tandem combina-tion of nitrile hydratase and amidase producing
microorga-nisms. Appl. Microbiol.
Biotech-nology 92: 600-606.
Langdahl, BR., P. Bisp & K. Ingvorsen.
1996. Nitrile hydrolysis by
Rhodo-coccus erythropolis BL1, an
acetonitrile-tolerant strain isolated
from a marine sediment.
Microbio-logy Letter. 142: 145-154
Manolov, T., K. Hakansson & G. Benoit. 2005. Continuous acetonitrile degradation in packed bedreactor.
Appl. Microbiol. Biotechnol. 66 :
567-574
Masuda, M. Y. Yamasaki, S. Ueno & A. Inone. 2007. Isolation of gisphenol
Sulistinah & Riffiani
a tolerant/degrading Pseudomanas
monteilii strain N-402. Extremo-philes 11 (2): 355-362
Nawaz, MS., TM. Heinze & CA. Cerniglia. 1992. Metabolism of benzonitrile and butyronitrile by
Klebsiella pneumoniae. Appl. Environ. Microbiol., 38 : 27-31
Oliver, MH., NK. Harison, JE. Bishop,
PJ. Cole & GJ. Lauren. 1989. A
rapid and convenient assay for counting cells cultured in microwell plates : Application for assessment
of growth factors. Journal of Cell
Science, 92: 513-518.
Podar, M., JR. Eads & TH. Richardson. 2006. Evolution of a microbial
nitrilase gene family: a comparative and environmental genomic study.
BMC Evol. Biol. 5 : 42
Sunarko, B., Adityarini, USF. Tambunan & N. Sulistinah. 2000. Isolasi, seleksi dan karakterisasi mikroba pendegradasi asetonitril. Berita
Biologi 5(2) : 177-185.
Sunarko, B., TU. Harwati, AT. Utari, D. Setianingrum & L. Nurhayani. 2007. Penapisan mikroba potensial untuk biokatalis produksi senyawa obat antiinflamasi nonsteroid (AINS). Laporan Teknik
Kegia-tan Pusat Penelitian