Judul :
FRAGMENTASI HUTAN :
Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan
ISBN : 978–602–1681–03-9
Penulis : Hendra Gunawan
Lilik Budi Prasetyo
Penelaah Ilmiah : Prof. Dr. Tukirin Partomiharjo, M.Sc.
Disain dan Tata Letak : Tatang Rohana Foto Sampul Depan : Hendra Gunawan
Diterbitkan oleh : Pusat Penelitian dan Pengembangan Konservasi dan Rehabilitasi
Badan Penelitian dan Pengembangan Kehutanan Kementerian Kehutanan
Jl. Gunung Batu No.5, Bogor 16610
©
Copyright 2013Hak cipta dilindungi oleh undang-Undang
Dilarang memperbanyak buku ini tanpa izin tertulis dari penerbit.
Boleh dikutip dengan menyebutkan sumbernya.
Saran pengutipan :
Gunawan, H. dan L.B. Prasetyo. 2013. Fragmentasi Hutan:
Teori yang mendasari penataan ruang hutan menuju pem bangunan berkelanjutan. Pusat Peneli tian dan Pengem bangan Konservasi dan Rehabi litasi. Bogor.
Saran tentang buku mohon disampaikan ke :
KATA PENGANTAR
KEPALA PUSAT PENELITIAN DAN PENGEMBANGAN
KONSERVASI DAN REHABILITASI, BADAN PENELITIAN
DAN PENGEMBANGAN KEHUTANAN
Salah satu tugas pokok lembaga penelitian adalah m enghasilkan dan mempublikasikan buku buku, baik yang bersifat teori tis maupun praktis, baik hasil penelitian maupun hasil pemikiran dan pengkajian literatur. Buku berjudul ”Fragmentasi Hutan : Teori yang mendasari penataan ruang hutan menuju pembangunan berkelanjutan” merupakan salah satu produk Pusat Penelitian dan Pengem bangan Konservasi dan Rehabi litasi yang diharapkan dapat membantu para prakitsi kehutanan, khususnya dalam pengaturan tata ruang kawasan hutan.
Kepada penulis disampaikan penghargaan dan terima kasih atas upayanya mem bukukan berbagai teori dalam sebuah buku yang mudah dimengerti, baik oleh para praktisi maupun akademisi sehingga dapat menjadi referensi, baik dalam penelitian maupun pengam bi lan keputusan. Kepada Penelaah Ilmiah juga disampaikan terima kasih atas telaah dan saran perbaikan untuk buku tersebut.
Akhirnya, semoga buku ini dapat bermanfaat untuk meningkatkan kualitas pengelolaan hutan dalam rangka pembangunan berkelanjutan dan berwawasan li ngkungan.
Bogor, Novem ber 2013 Kepala Pusat,
KATA PENGANTAR
Isu kepunahan keanekaragaman hayati tropika menjadi isu krusial yang menjadi perhatian internasional pada dua dekade terakhir. Fragmentasi hutan merupakan salah satu penyebab utama punahnya keanekaragaman hayati di beberapa lokasi. Fragmentasi hutan merupakan proses dan hasi l dari peri laku manusia dalam m emanfaatkan sumbedaya hutan, seperti konversi, penebangan liar, pem bakaran hutan, perladangan dan perambahan kawasan hutan.
Keberhasilan pengelolaan keanekaragaman hayati di kantong -kantong hutan yang telah terfragmentasi mem erlukan pemahaman konsep-konsep yang lahir dari teori biogeografi pulau, seperti fragmentasi, koridor dan single large or several small (SLOSS). Buku ini membantu para mahasiswa, praktisi konservasi, manajer satwaliar serta pengambi l kebijakan penataan ruang pem bangunan dalam memahami fragmentasi hutan dan kaitannya dengan pengelolaan dan konservasi keanekarag aman hayati.
Buku ini merupakan kom pilasi dari hasil terjemahan, penelaahan dan analisis dari berbagai buku teks dan website yang relevan. Untuk memahami dan m engetahui lebih detai l, para pembaca di pesilakan membaca buku- buku teks yang m enjadi sumber penu lisan buku ini.
Penulis menyadari, buku ini masih jauh dari kesempurnaan, oleh karena itu saran-saran dari peminat akan dipertimbangkan untuk perbaikan. Akhirnya, semoga buku ini bermanfaat bagi para peminat dan pem erhati masalah konservasi keanekaragaman hayati.
Bogor, Novem ber 2013
DAFTAR ISI
Halaman
BAB 1. FRAGMENTASI: SUATU PENDAHUL UAN ... 1
BAB 2. MASALAH DAN PROSES FRAGMENTASI ... 10
BAB 3. TEORI BIOGEOGRAFI PULAU vs FRAGMENTASI HUTAN ... 16
BAB 4. FRAGMENTASI HABITAT DIPANDANG DALAM SKALA LANSKAP ... 30 BAB 5. KONSEP KORIDOR DAN SI NGLE LARGE OR SEVERAL SMALL (SLOSS) ... 42 BAB 6. DINAMIKA POPULASI PADA SKALA LANSKAP ... 52
BAB 7. PENGARUH FRAGMENTASI TERHADAP SATWALIAR ... 71
BAB 8. PERPINDAHAN SATWA DALAM LANSKAP ... 95
BAB 9. FRAGMENTASI DAN DINAMIKA METAPOPULASI ... 103
BAB 10. IMPLIKASI PENGELOLAAN KONSERVASI ... 109
DAFTAR GAMBAR
Nomor Teks Halaman
Gambar 1. Model konseptual pengaruh fragmentasi
(Kupfer et al. 2004) ... 13
Gambar 2. Jum lah jenis relatif pada (a) pulau-pulau keci l, dekat, (b) pulau- pulau besar, jauh atau keci l, dekat, dan (c) pulau- pulau besar, dekat (Mac Arthur and
Wilson 1967) ... 21
Gambar 3. Mekanisme bekerjanya pengaruh isolasi ... 27
Gambar 4. Klasifikasi rejim gangguan berdasarkan pada luas yang terganggu dan luas lanskap (diadaptasi oleh Shugart
1984; dari Shugart and West 1981)………... 32
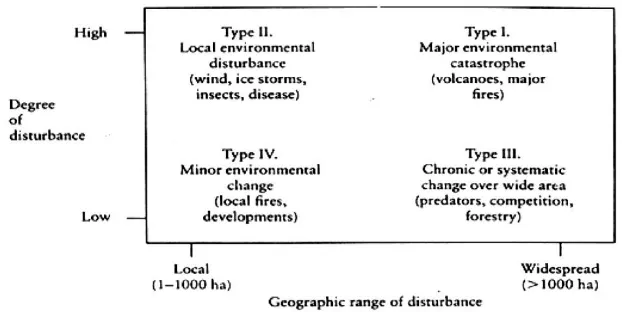
Gambar 5. Empat tipe gangguan yang ditunjukkan oleh derajat gangguan dan luas geografis yang dipengaruhinya
(Morrison et al. 1992) ... 32
Gambar 6. Fungsi insiden dari pelatuk totol besar (Picoides major) (Dari Moore dan Hooper dalam Wi lcove
et al. 1986) ... 35
Gambar 7. Contoh koridor perpindahan yang di buat di
pegunungan Costa Rica (Dari Stiles and Clark 1989) … 45
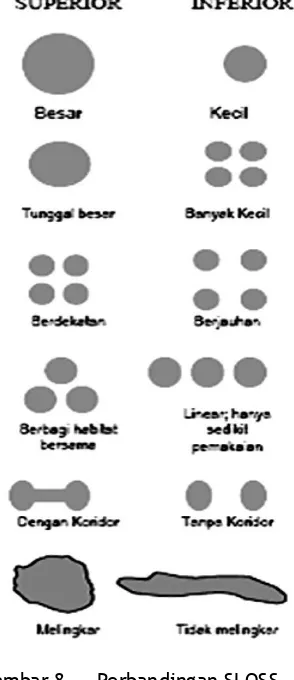
Gambar 8. Perbandingan SLOSS (Dari berbagai sumber) ... 50
Gambar 9. Gradien pertukaran di dalam metapoulasi dari
berbagai struktur internal (Morrison et al. 1992) ... 56
Gambar 11a. Batas Das dan sistem riparian (riverine)
(Morrison et al. 1992) ... 58
Gambar 11b. Kejadian sepuluh patches dari hutan tua
(Morrison et al. 1992) ...... 58
Gambar 11c. Kejadian vertebrata obligat dewasa di tujuh dari 10
Gambar 11d. Seleksi tiga patches untuk pemanenan tebang habis (Morrison et al. 1992) ...
59
Gambar 11e. Akibat langsung dari ganguan pemanenan : hilangnya
spesies pada tiga patches (Morrison et al. 1992) ... 60
Gambar 11f. Kehi langan spesies kemudian pada patch hutan yang jauh, terisolasi dan keci l (faunal relaxation) (Morrison et al. 1992) ... 60
Gambar 11g. Masih kehilangan spesies kemudian pada suatu patch hutan lebih besar yang kini terisolasi
(Morrison et al. 1992)... 61
Gambar 12. Contoh habitat yang terfragmentasi oleh: (1) jaringan listrik, (2) jalan raya, (3) tebang habis dan (4)
pertanian (Foto: Hendra Gunawan, 2008) ... 73
Gambar 13. Sekumpulan patch hutan yang ditempati oleh populasi-populasi penghuni hutan. Tebal tipisnya panah menunjukkan laju pertukaran individu antar populasi (panah tebal m enunjukkan lebih banyak pertukaran). Catatan: Laju pertukaran dikoreksi oleh seberapa dekat patch- patch dalam ruang (Sumber: http://chesapeake.towson.
edulandscape/forestfrag/effects.asp ... 76
Gambar 14. Penghunian (occupancy) sekumpulan patch hutan yang mendukung suatu metapopulasi spesies penghuni hutan pada dua waktu yang berbeda. Patch hitam ditempati dan patch putih tidak ditem pati. Catatan: penghunian pada patch individual berubah sepanjang waktu (seperti patch- patch dikolonisasi dan po pulasi-populasi di dalam patch punah), tetapi jum lah patch-patch yang ditem pati tetap sama sepanjang waktu (Sumber: http://chesapeake. towson.edu/landscape/
forestfrag/effects.asp ... 76
Gambar 15. Grafik menunjukkan hubungan antara kelembaban tanah dan edge dan core habitat dala patch hutan. Banyak spesies burung tidak dapat mentolerir kondisi tepi habitat (edge habitat) dan hanya ditemukan di core habitat (Sumber:http://chesapeake.towson.
edulandscape/forestfrag/effects.asp) ... 78
Gambar 16. Berbagai bentuk fragment dan gambaran luas
Gambar 17. Pengaruh kehi langan dan fragmentasi habitat pada ukuran patch dan isolasi (Fahrig 1997) ………..
82
Gambar 18. Ilustrasi zona pengaruh tepi (edge effect) suatu pulau
habitat………... 86
Gambar 19. Koridor harus cukup lebar untuk mem berikan manfaat positif lebi h banyak bagi satwaliar. Koridor riparian yang sempit seperti yang tampak dalam gambar berpeluang menghasi lkan damak negatif bagi
satwaliar ……… 101
Gambar 20. Metapopulasi adalah suatu populasi sumber (source)
dan populasi penerima (sink)(Barnes 2000) ……….……… 106
Gambar 21. Ukuran, bentuk, konfigurasi dan jum lah patch sem ua mempengaruhi jum lah habitat interior dalam patch kecil, tunggal, patch-patch persegi memberikan jumlah habitat interior yang keci l dan patch-patch lingkaran besar memberikan habitat interior terbesar
(Barnes 2000) ... 110
Gambar 22. Tahapan prosedur kajian evaluasi lanskap dan ana lisis
spasial ... 115
Gambar 23. Komposisi luas setiap kelas penutupan lahan di
kawasan lanskap Merapi Merbabu ... 118
Gambar 24. Jum lah patch setiap kelas penutupan lahan ……… 119
Gambar 25. Luas patch rata-rata setiap kelas penutupan lahan .... 120
Gambar 26. Total edge setiap kelas penutupan lahan ... 120
Gambar 27. Edge density setiap kelas penutupan lahan ... 121
Gambar 28. Mean patch edge setiap kelas penutupan lahan ………… 121
Gambar 29. Mean shape indexsetiap kelas penutupan lahan …... 121
Gambar 30. Mean patch fractal dimension untuk setiap kelas
penutupan lahan ………... 123
Gambar 31. Lokasi dugaan habitat macan tutul terpi lih (warna
DAFTAR TABEL
Nomor Teks Halaman
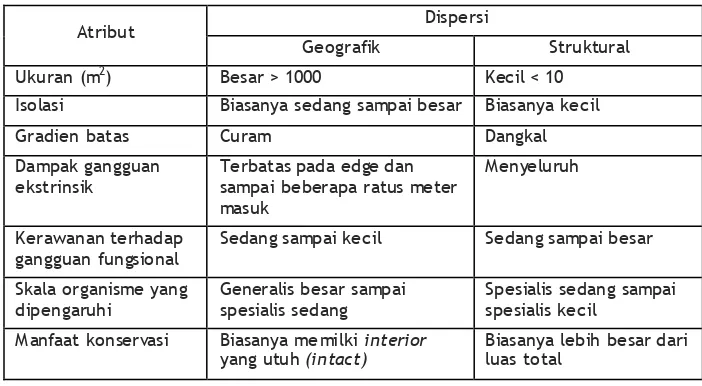
Tabel 1. Implikasi dispersi habitat pada skala berbeda untuk
berbagai atri but lanskap terfragmentasi ... 33
Tabel 2. Klasifikasi faktor-faktor untuk dievaluasi dalam suatu
studi gangguan lanskap ………... 54
Tabel 3. Komposisi pentutupan lahan AOI Kawasan Merapi-Merbabu 116
Tabel 4. Hasil patch analyses pada skala lanskap kawasan Merapi-
Merbabu ... 117
1
FRAGMENTASI :
SUATU PENDAHULUAN
PENDAH ULUAN
DEFINISI DAN PENGERTIAN
MENGAPA PERLU MEMPELAJARI FRAGMENTASI?
DAFTAR PUSTAKA
I. PENDAHULUAN
Mempelajari fragmentasi merupakan bagian dari i lmu ekologi
lanskap. Ekologi lanskap merupakan i lmu baru yang diturunkan dari disiplin
terdahulu sebagai suatu gabungan arsitektur lanskap, zoogeografi, geografi tumbuhan dan sinekologi. Ekologi lanskap merupakan i lmu yang
mempelajari respon spesies atau komunitas terhadap pola-pola (patterns)
patch dalam suatu lanskap (Morrison et al. 1992). Banyak prinsip-pri nsip
dasar ekologi lanskap diturunkan dari biogeografi pulau, zoogeografi dan
fitogeografi, yaitu i lm u yang mem pelajari penyebaran dan pergerakan
satwa dan tumbuhan melintasi pulau- pulau dan wi layah geografi yang lebih
luas (Morrison et al. 1992).
Ekologi lanskap banyak m emfokuskan pada dinamika tumbuhan dan
satwa di dalam patches (kantong habitat), khususnya di pulau-pulau dan
lingkungan terisolasi lainnya. Teori pulau k lasik menyatakan bahwa di
pulau, dinamika populasi dan struktur komunitas ditentukan oleh beberapa faktor : ukuran (pulau kecil memi liki resiko kepunahan lokal lebih besar);
jarak dari sumber spesies yang mengkolonisasi (semakin jauh, semakin
dispersal, demografik (survivorship, rekrutmen); dan spesialisasi
penggunaan habitat (Morrison 1992).
Pemukiman manusia dan kegiatan terkait lainnya, seperti hutan tanaman dan pertanian telah mengubah lanskap alami menghasilkan suatu mosaik dari habitat yang terfragmentasi. Fragmentasi habitat dapat
memberikan pengaruh merugikan pada flora dan fauna dari habitat alami yang sebelumnya masih utuh dan berkesinambungan. Ada dua pengaruh
utama dari fragmentasi yaitu : mengurangi total luas dari habitat asal dan
menciptakan wi layah tepi (edge area) di antara habitat asal dengan lanskap
yang terganggu oleh manusia, yang dikenal sebagai efek tepi (edge effects).
Dalam fragmentasi habitat ada enam proses terpisah yang dapat dipertim bangkan yaitu:
Berkurangnya luas total dari habitat
Meningkatnya jum lah wi layah tepi (edge)
Berkurangnya luasan habitat interior
Terisolasinya suatu fragment (potongan) habitat dari wilayah habitat
lainnya
Terpecahnya satu patch (kantong) habitat menjadi beberapa patch
(kantong) habitat yang lebih keci l
Berkurangnya ukuran rata-rata setiap patch (kantong) habitat
II. DEFINISI DAN PENGERTIAN
A. Ekologi Lanskap
Ekologi lanskap merupakan suatu bagian dari i lmu ekologi yang mempelajari bagaimana struktur lanskap mem pengaruh kelim pahan dan
distribusi organisme. Ekologi lanskap juga didefinisikan sebagai ilmu yang
mempelajari pengaruh pola (pattern) dan proses, dimana pola di sini
khususnya mengacu pada struktur lanskap. Dengan demikian secara
lengkap ekologi lanskap dapat di definisikan sebagai ilmu yang mempelajari bagaimana struktur lanskap mempengaruhi (memproses dan membentuk) kelimpahan dan distribusi organisme.
yang mempengaruhi proses-proses ekologi seperti distribusi, aliran energi,
materi dan individu dalam lingkungannya (yang pada gi lirannya mungkin
mempengaruhi ditri busi elemen- elemen lanskap itu sendiri). Teori biogeografi pulau dari MacArtur dan Wilson merupakan fokus dari ekologi lanskap yang oleh Levin digunakan untuk menjelaskan model metapopulasi.
Dalam ekologi lanskap dapat di pelajari bagaimana fragmentasi habitat
mempengaruhi daya hidup suatu populasi (population viability). Dalam
perkembangannya ekologi lanskap banyak m eng gunakan teknologi Sistem Informasi Geografis (SIG) dan banyak data habitat yang tersedia (seperti
citra satelit dan foto udara.
Sebagai bagian dari ekologi, ekologi lanskap ditujukan m empelajari sebab dan akibat dari heterogenitas spasial (Forman 1995). Heterogenitas
merupakan ukuran bagaimana bagian-bagian suatu lanskap berbeda satu
sama lain. Ekologi lanskap melihat pada bagaimana struktur spasial mempengaruhi kelimpahan organisme pada skala lansk ap, serta peri laku
dan fungsi lanskap secara keseluruhan. Hal ini berarti juga m empelajari
pola, atau keteraturan internal lanskap, proses atau operasi kontinu dari fungsi organisme (Turner 1989). Ekologi lanskap juga mencakup
geomorfologi dalam penerapannya untuk disain dan arsitektur lanskap (Allaby 1998). Geomorfologi merupakan i lmu yang mempelajari bagaimana
formasi geologi mempengaruhi struktur lanskap.
B. Fragmentasi
Fragmentasi hutan terjadi karena hutan yang luas dan menyam bung terpecah menjadi blok-blok lebih kecil akibat pem bangunan jalan,
pertanian, urbanisasi atau pembangunan lai n. Akibatnya mengurangi fungsi
hutan sebagai habi tat berbagai spesies tumbuhan dan satwaliar. Fragmentasi juga mem pengaruhi struktur, temperatur, kelembaban dan
pencahayaan yang akan mengganggu satwa hutan yang adpatasinya telah terbentuk selama ribuan tahun. Fragmentasi didefinisikan sebagai
pemecahan habitat organisme m enjadi kantong-kantong (patches) habitat
yang mem buat organisme kesulitan melakukan pergerakan dari kantong habitat yang satu ke yang lainnya. Fragmentasi dapat disebabkan oleh
memisahkan habitat bahkan oleh jaringan kabel listrik (Rusak & Dobson
2007).
Fragmentasi adalah proses pemecahan suatu habitat, ekosistem
atau tipe landuse menjadi bidang-bidang lahan yang lebih keci l dan
fragmentasi juga merupakan sebuah hasil dimana proses fragmentasi
mengubah atribut-atribut habitat dan karakteristik suatu lanskap yang ada. Fragmentasi habitat mengubah konfigurasi spasial suatu kantong habitat
(habitat patches) besar dan menciptakan isolasi atau perenggangan
hubungan antara kantong-kantong (patches) habitat asli karena terselingi
oleh mosaik yang luas atau ti pe habitat lain yang tidak sesuai bagi spesies
yang ada (Wiens 1990).
Franklin et al. (2002) mengembangkan definisi baru tantang
fragmentasi sebagai hasil (outcome) dan proses. Hasil (outcome) dari
fragmentasi habitat adalah diskontinuitas yang di peroleh dari serangkaian mekanisme, di dalam distribusi spasial suatu sumberdaya dan kondisi yang
ada dalam suatu areal pada suatu skala tertentu yang mempengaruhi
okupansi, reproduksi atau survival suatu spesies. Fragmentasi habitat
didefinisikan sebagai serangkaian mekanisme yang mengakibatkan
diskontinuitas distribusi spasial suatu habitat. Ada empat komponen kunci dari dua definisi tersebut yai tu : (1) diskontinuitas, (2) m ekanisme, (3)
distribusi spasial dari suatu sumberdaya dalam suatu area, dan (4) atribut
demografik (Franklin et al. 2002).
Konsep fragmentasi habi tat diturunkan dari teori biogeografi pulau
(MacArthur & Wilson 1967), yakni jumlah spesies meningkat dengan
meningkatnya ukuran pulau (Haila 2002). Fragmentasi penting m endapat
perhatian karena berpengaruh pada kekayaan spesies dari komunitas, trend
populasi beberapa spesies dan keanekaragaman hayati ekosistem secara
keseluruhan (Morrison et al. 1992).
Menurut Wilcove (1987) dalam Morrison et al. (1992) ada em pat
cara fragmentasi dapat menyebabkan kepunahan lokal : (1) spesies dapat mulai keluar dari kantong habitat yang terli ndungi; (2) kantong habitat
gagal menyediakan habitat karena pengurangan luas atau hilangnya
variabilitas demografik, kemunduran genetik atau disfungsi sosial; (4)
fragmentasi dapat m engganggu hubungan ekologis yang penting sehingga
dapat menim bulkan sebab sekunder kepunahan dari hilangnya spesies kunci
dan pengaruh m erugikan dari lingkungan luar dan efek tepi (edge effect).
C. Patch
Patch, m erupakan suatu terminologi dasar dalam ekologi lanskap
didefinisikan sebagai sebuah area yang relatif homogen yang berbeda
dengan sekelilingnya (Forman 1995). Patch merupakan unit dasar dari
lanskap yang berubah dan berfluktuasi. Proses perubahan dan fluktuasi ini
disebut sebagai dinamika patch. Patch m emiliki bentuk tertentu dan
konfigurasi spasial, dan dapat digambarkan dalam komposisi
variabel-variabel internalnya seperti jum lah pohon, jum lah jenis pohon, tinggi pohon, atau variabel lainnya (Forman 1995).
D. Matrix
Matrix merupakan latar belakang (background) sistem ekologi dari
suatu lanskap dengan derajat konektifitas yang tinggi. Konektivitas adalah
ukuran bagaimana suatu koridor, jaringan (network) atau matrix terhubung
atau berkesinambungan (Forman, 1995). Sebagai contoh, suatu lanskap
berhutan (matrix) yang memi liki sedikit celah (gap) dalam tutupan
hutannya berarti memiliki konektifitas lebih tinggi. Koridor memi liki fungsi
penting sebagai jalur penghubung antara suatu tipe tutupan lahan yang berbeda dengan tetangga di kedua sisinya (Forman, 1995). Suatu jaringan
(network) merupakan suatu sistem hubungan antar koridor, sementara
mosaik menggambarkan pola (pattern) dari patch, koridor dan matrix yang
membentuk suatu lanskap dalam suatu kesatuan (Forman 1995).
E. Boundary dan Edge
Patches bisa memiliki batas (boundary) yang jelas atau tidak jelas
(kabur) (Sanderson & Harris 2000). Suatu zona yang tersusun atas ekosistem
edge di perbatasan disebut boundary (Forman 1995). Edge (tepi) berarti
menyebabkan perbedaan lingkungan antara interior suatu patch dengan
tepiannya (edge). Efek tepi (edge effect) ini meliputi perbedaan komposisi
spesies atau kelimpahan di bagian luar patch (Forman 1995). Sebagai
contoh, ketika suatu lanskap merupakan sebuah mosaik dari tipe- tipe
seperti hutan berdekatan dengan padang rum put, maka edge-nya adalah
Habitat adalah suatu ti pe komunitas biotik atau kesatuan komunitas biotik dimana spesies atau populasi hidup (Bai ley, 1984). Habitat adalah
suatu unit li ngkungan, alami maupun tidak (meliputi iklim, makanan, cover
dan air) dimana seekor satwa, tumbuhan atau populasi secara alami dan
normal hi dup dan berkembang (Helms, 1998). Defi nisi habitat terbaru yang
relevan untuk pengelola satwa liar datang dari Hall et al. (1997) yaitu
sumberdaya dan kondisi yang ada pada suatu tempat yang memberikan
tempat hidup (occupancy), termasuk survival dan reproduksi suatu
organisme. Definisi ini berimplikasi bahwa habitat adalah sejum lah
sumberdaya spesifik yang dibutuhkan oleh suatu spesies (Hall et al. 1997).
III. MENGAPA PERLU MEMPELAJARI FRAGMENTASI?
Sampai saat ini para ahli satwa m emiliki pandangan tradisioanal
tentang edges dan ekoton sebagai sesuatu yang positif. Dalam buku-buku
teks klasik (Leopold 1933; Thomas et al. 1979; Yoakum & Dasmann 1971
dalam Morrison et al. 1992) merekomendasikan bahwa pengelola satwa
seharusnya meningkatkan jum lah edges, karena satwa melim pah pada
pertemuan habitat (mereka menganggap habitat sebagai vegetasi atau lingkungan secara umum, bukan habitat spesifik suatu spesies yang
memerlukan syarat khusus). Anggapan bahwa pem bukaan dan pem bersihan
hutan m enguntungkan satwa ternyata tidak selalu benar. Dalam beberapa
berkesinambungan sekarang terancam karena di buka untuk mem buat
ekoton sehingga berdam pak negatif bagi satwa seperti meni ngkatnya
pemangsaan dan overbrowsing. Anggapan bahwa m enciptakan edges dan
membuka hutan itu bermanfaat perlu diteliti lagi dengan seksama (Morrison
et al. 1992).
Jum lah spesies satwa dan tumbuhan berhubungan dengan kondisi
yang ada dalam interior yang relatif luas dari vegetasi matang atau
dipengaruhi oleh vegetasi terganggu dan pengaruh tepi. Komposisi dan kelimpahan relatif dari asosiasi spesies dalam kom unitas biotik bervariasi
antar lanskap menurut jum lah dan sebaran habitat secara spasial. Dengan
demikian fragmentasi berpengaruh terhadap kekayaan spesies dari suatu komunitas, kecenderungan populasi spesies tertentu dan keanekaragaman
hayati secara keseluruhan dari suatu ekosistem. Apakah pengaruh tersebut
disukai atau tidak bergantung pada jum lah dan komposisi spesies yang ada pada suatu area. Hal ini memi liki implikasi kuat untuk pengelolaan lanskap
(Morrison et al. 1992).
Fragmentasi hutan juga menjadi perhatian di dalam hutan tropis.
Klein (1989) dalam Morrison et al. (1992) yang melakukan penelitian
terhadap kumbang bangkai dan organisme pada kotoran (feces) di hutan
bersinambung dan hutan terfragm entasi, menyim pulkan bahwa fragm entasi
menyebabkan berkurangnya spesies pengurai kotoran dan secara tidak
langsung fragmentasi hutan berpengaruh pada siklus hara dan proses -proses ekosistem terkait lainnya, dan mungkin mempengaruhi produktivitas
tapak dalam jangka panjang.
Fragmentasi hutan temperate di Amerika Serikat bagian barat
dapat mengurangi atau menghi langkan populasi pengerat keci l pemakan
jamur (mycophagus) yang menjadi agen kunci penyebaran sejum lah spesies
jamur hypogeous (di bawah tanah), khususnya mychorrizae yang penting
bagi hutan konifer (Maser et al. 1978 dalam Morrison et al. 1992).
Jamur-jamur michorri zae tum buh di rambut-ram but akar pohon konifer dan
membantu menyerap hara. Pengerat (rodent) kecil yang bertindak sebagai
agen kunci penyebaran mem butuhkan tegakan hutan yang tua atau pohon
mungkin secara tidak langsung mengurangi produktivitas hutan dalam
jangka panjang (Morrison et al. 1992).
Fragmentasi lingkungan merupakan suatu isu perencanaan dan konservasi habitat, yakni peningkatan fragmentasi dapat mempengaruhi ketahanan populasi dan keragaman spesies dan komunitas. Fragmentasi
dapat mengisolasi individu, unit berkem bangbiakan, dan sub populasi dari
spesies-spesies dalam patchinterior. Isolasi ini dapat meningkatkan resiko
kepunahan lokal karena peni ngkatan variasi ukuran populasi dalam peluang untuk hidup dan kesempatan berkem bangbiak; fluktuasi di lingkungan dan
kualitas dan kuantitas sumberdaya; peningkatan kerentanan untuk punah
dari patch yang lebih keci l, lebih terisolasi jika menghadapi bencana seperti
angin badai dan kebakaran (Morrison et al. 1992).
Saat ini di banyak negara, perhatian publik terhadap perencanaan
habitat telah meningkat karena adanya peningkatan isu terkait seperti penurunan keanekaragaman hayati, peningkatan fragmentasi lingkungan
yang dianggap berdam pak buruk bagi keanekaragaman hayati dan hilangnya
hutan-hutan tua serta semakin langka dan berkurangnya habitat. Oleh karena itu, baik dari perspektif s osial dan perhatian publik maupun
perspektif ilmiah, fragmentasi lingkungan dan pengelolaan habitat dalam skala lanskap perlu mendapat perhatian dan dipelajari secara benar.
DAFTAR PUSTAKA
Allaby, M. 1998. Oxford Di ctionary of Ecology. Oxford University Press, New York, NY.
Bailey, J.A. 1984. Princples of Wildlife Management. John Wiley and Sons. New York.
Forman R.T.T. 1995. Land mosaics: the ecology of landscapes and regions. Cambridge University Press, Cambridge, 632 pp.
Franklin, A.B., B.R. Noon, And T. L.George. 2002. What Is Habitat
Fragmentation? Studi es in Avian Biology No. 25:20-29.
http://www. hum boldt. edu/-tlg2/publications/whatis
Haila, Y. 2002. A conceptual genealogy of fragmentation research: from island biogeography to landscape ecolog y. Ecological Applications
12:321–334.
Hall, L. S., P. R. Krausman, And M. L.Morrison. 1997. The habitat concept and a plea for standard terminology. Wildlife Society Bulletin Diakses Tanggal 17 Oktober 2006.
http://en.wikipedia.org/wiki/Habitat_fragmentation. Diakses Tanggal 17 Oktober 2006.
http://en.wikipedia.org/wiki/Landscape ecology. Diakses Tanggal 24 Februari 2007.
http://www.carleton.ca/lands-ecol/whatisle.html. Diakses Tanggal 24 Februari 2007.
MacArthur, R. H., and E. O. Wilson. 1967. The theory of island biogeography. Princeton University Press, Princeton, New Jersey, USA.
Morrison, M.L., B.G. Marcot and R.W. Mannan. 1992. Wi ldlife-Habitat Relationships. The University of Wisconsisn. Madison, Wisconsin. Rusak, H. and C. Dobson. 2007. Forest Fragmentation. www.ontarionature
.org. Diakses tanggal 26 Februari 2007.
Sanderson, J. and L. D. Harris (eds.). 2000. Landscape Ecology: A Top-Down Approach. Lewis Publishers, Boca Raton, Flori da, USA.
Turner, M.G. and R. H. Gardner (eds.). 1991. Quantitative Methods in Landscape Ecology. Spri nger-Verlag, New York, NY, USA.
2
MASALAH DAN PROSES FRAGMENTASI
MASALAH FRAGMENTASI
PROSES FRAGMENTASI
PENGARUH FRAGMENTASI
MENGUKUR FRAGMENTASI
DAFTAR PUSTAKA
I. MASALAH FRAGMENTASI
Fragmentasi habitat merupakan masalah penting di seluruh dunia.
Penyempitan habitat secara luas memang cukup serius, tetapi ketika
dikombinasikan dengan fragmentasi maka dapat meruntuhkan kesatuan ekosistem secara keseluruhan. Jalan, urbanis asi dan pertanian merupakan
kegiatan utama manusia yang mem ecah-mecah wi layah alami yang seringkali diikuti oleh malapetaka bagi satwa. Wilayah yang sebelum nya
merupakan hutan yang kompak dan utuh menjadi terpecah-pecah,
berukuran keci l dan terisolasi, sehingga beberapa jenis satwa tidak dapat melakukan perpindahan atau pergerakan untuk mencari makan atau untuk
berkembang biak.
Di sisi lain, ada beberapa jenis satwa dan tumbuhan yang lebih
menyukai habitat interior berupa hutan yang rapat dan gelap, maka jika
50% hutan tersebut dibabat, misal untuk membuat jalan atau lapangan
parkir dan sisanya terbelah oleh jalan, maka hutan yang lebat, sejuk dan gelap berubah m enjadi terang benderang, kelem baban dan temperaturnya berubah dan tidak sesuai lagi sebagai habitat satwa dan tum buhan tersebut.
Dengan demikian, fragmentasi juga meningkatkan efek tepi (edge effect).
dari habitat lainnya di sekitarnya. Semakin keci l suatu habi tat, semakin
besar proporsi yang terkena efek tepi dan hal ini dapat menyebabkan
perubahan yang dramatis bagi komunitas satwa dan tum buhan.
Bila suatu populasi satwa menjadi kesulitan menyeberangi jalan raya yang menghalanginya untuk mencapai kantong habitat lainnya yang
berjarak cukup jauh, maka satwa tersebut hanya dapat berkembang biak terbatas di kantong habitat tempatnya tinggal yang dapat dikatakan sebagai
pulau dan populasinya akan menghadapi resiko inbreeding. Lebih jauh, jika
populasi diserang wabah penyakit atau bencana alam lainnya, maka satwa
tersebut akan mengalami kepunahan lokal dan sulit untuk rekolonisasi dari
populasi lainnya. Sejalan dengan itu, maka jelas bahwa kesinambungan atau konektivitas dari hutan sangat penting.
Suatu hutan yang sehat dan cukup besar akan mampu mendukung
organisme dengan jelajah luas, seperti jenis-jenis satwa besar atau satwa pemangsa. Pengurangan luas dapat memiliki dam pak langsung pada spesies
ini dan karena predator sering memainkan peran penting sebagai pengatur
populasi spesies lain, maka keseimbangan ekosistem dapat sangat terganggu. Dalam skala luas, perubahan ik lim juga mungkin m emaksa suatu
spesies untuk bermigrasi, jika habitat alaminya sangat terfragmentasi, banyak di antara mereka tidak dapat bermigrasi dan akan menghadapi
resiko kepunahan. Ini juga mem buktikan betapa pentingnya suatu lanskap
yang berkesinambungan.
II. PROSES FRAGMENTASI
Fragmentasi umumnya terjadi melalui hilangnya habitat (habitat
loss), sebaliknya hi langnya habi tat dapat dipandang sebagai akibat
fragmentasi. Tetapi fragmentasi dapat disertai hi langnya habitat
(berkurangnya jum lah) seiring dengan pemecahan atau pem bagian kantong habitat besar menjadi kantong-kantong habitat berukuran keci l dan lebih
terisolasi (Hunter 1997; Hai la 1999; Frank lin et al. 2002; Fahrig 2003). Jika
hilangnya habitat dan fragmentasi dipandang secara terpisah, maka hilangnya habitat memi liki dam pak lebih signifikan bagi kelangsungan hidup
karena fragmentasi dan hi langnya habitat terjadi bersamaan maka sangat
sulit untuk menentukan mana yang lebih penting bagi perubahan habitat
(Haila 1999).
Fragmentasi bekerja dalam empat cara ketika hilangnya habitat dan fragmentasi digabung untuk menggambarkan dan mengkategorikan
prosesnya (Frank lin et al. 2002; Fahrig 2003) : (1) habitat hi lang tanpa
fragmentasi; (2) pengaruh kombinasi hilangnya habitat dan pemecahan
habitat menjadi patches lebih keci l; (3) pemecahan habitat menjadi
patch-patch lebih keci l tanpa kehi langan habitat; dan (4) hilangnya habitat dan
pemecahan habitat menjadi patch-patch lebih kecil serta penurunan
kualitas habitat. Contoh ini berlaku untuk lanskap yang terdiri lebih dari satu habitat dan dikeli lingi oleh matriks di dalam suatu kesatuan lanskap.
Kasus pertama dan kedua berlaku ketika lanskap keseluruhan berisi satu
habitat dan tidak ada matriks di sekeli lingnya. Dalam kenyataan, kasus dua dan em pat merupakan cara yang paling umum dalam fragmentasi habitat.
Fragmentasi habitat merupakan satu aspek dari tahapan proses
yang secara spasial dan temporal mengubah habitat dan lanskap yang diakibatkan oleh sebab-sebab alami maupun antropogenik (Forman 1995).
Tetapi, perubahan habitat tidak dapat dihindari karena ti dak ada habitat atau lanskap yang tetap (Forman 1995). Lanskap berubah me lalui lima
proses spasial dengan berbagai derajat overlap sepanjang periode
perubahan lahan (Forman 1995), dan fragmentasi hanyalah satu outcome.
Proses ini dapat diakibatkan oleh penyebab alami dan antropogenik.
Perforasi (Perforation) merupakan proses membuat lubang di dalam
habitat. Pem otongan (Dissection) adalah pemotongan atau pem bagian area
menjadi habitat berbeda dengan lebar yang relatif sama. Fragmentasi
(Fragmentation) adalah pemecahan habitat menjadi potongan-potongan
yang lebih kecil. Penyusutan (Shrinkage) terjadi seiring potongan habitat
berlanjut dengan penurunan luas. Erosi habitat (Attrition) adalah proses
dimana kantong habitat yang tersisa berangsur hilang karena degradasi habitat atau suksesi.
Fragmentasi dimulai dengan dissection ketika jalan, jaringan
transmisi, sungai dan fitur linear lainnya menjadi penghalang pergerakan.
(habitat patches) keci l yang dibuat oleh manusia atau sebab alami dan efek
tepi menjadi nyata. Fragmentasi merupakan tahap ketiga yang terjadi
ketika kantong habitat yang lebih keci l meningkat frekuensinya dan berkurang luasnya sampai pada tingkat di mana habitat yang terfragmentasi
mulai m endominasi lanskap. Attrition m erupakan tahap akhir dimana lahan
alami atau habitat asli tersisa sebagai kantong yang kecil dan terisolasi di tengah-tengah lanskap yang sekarang didominasi oleh suatu mosaik habitat
yang telah berubah dan terfragmentasi. Proses fragmentasi membuat habitat menjadi tidak sesuai atau memi liki kesesuaian rendah bersamaan
dengan berkurangnya kualitas habitat satwaliar. Sebaliknya, jika proses
gangguan mengubah mosaik habitat tetapi tidak ada perubahan kualitas habitat berarti ti dak terjadi fragmentasi, atau habitatnya berubah teta pi
tidak terfragmentasi (Hunter 1997)
III. PENGARUH FRAGMENTASI
Ada empat cara primer fragmentasi hutan dapat mempengaruhi
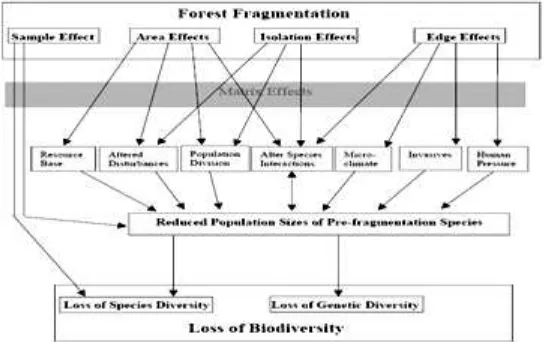
keanekaragaman hayati, yaitu: (1) keterwakilan (sample effect); (2) luas
area (area effect); (3) isolasi (isolation effect) dan (4) pengaruh tepi (edge
effect) (Gambar 1). Masing-masing pada gilirannya akan berpengaruh pada
sebaran populasi, komunitas dan proses ekosistem (Kupfer et al. 2004).
Mekanisme dan proses fragmentasi menghasi lkan tiga tipe
pengaruh: (1) ukuran patch; (2) pengaruh tepi (edge effect); dan (3)
pengaruh isolasi (Fahrig, 2003). Ahli satwa harus memperhatikan semua karena ketiganya terjadi pada fragmentasi habitat dan masing-masing
memerlukan penanganan yang berbeda (Frankli n et al. 2002; Fahrig 2003).
Fragmentasi habitat dapat dipandang dari segi positif dan negatif.
Pengaruh positifnya adalah m eningkatkan keragaman habitat, m enciptakan
penjajaran habitat yang bermanfaat, dan meningkatkan edge yang disukai
spesies satwaliar generalis. Fragmentasi mem berikan pengaruh negatif ketika: (1) ada habitat yang hi lang; (2) terbentuk kantong habitat lebih kecil yang mendorong pada kepunahan lokal dan isolasi; (3) habitat-habitat tidak lagi bersambungan, khususnya jika fragmentasi disebabkan oleh
aktifitas non kehutanan; dan (4) jum lah edge meningkat sehingga
fragmentasi habitat merugikan spesies interior (Barnes 2000).
IV. MENGUKUR FRAGMENTASI
Mengukur fragmentasi sangat menarik, ada banyak ukuran yang dapat digunakan untuk mengkuantifikasikan fragmentasi habitat. Sebagai
contoh dalam program FRAGSTATS tersedia 100 matrix fragmentasi habitat
(McGarigal and Marks 1995), tetapi hanya sedikit kesepakatan matrik mana yang paling sesuai dan bisa diterjemahkan ke dalam tindakan manajemen konservasi (Garrison 2005).
Indeks-indeks struktur lanskap seringkali digunakan untuk
mengkuantifikasikan fragmentasi habitat. Indeks -indeks telah
dikembangkan untuk m engukur tiga aspek struktur lanskap: (1) kom posisi
lanskap; (2) konfigurasi lanskap; dan (3) bentuk patch di dalam lanskap.
Komposisi menunjukkan jumlah dari tipe penutupan (cover) yang berbeda
yang ditemukan dalam lanskap. Konfigurasi menunjukkan bagaimana patch
dari tipe cover yang sama atau berbeda ters usun di dalam lanskap dan
hubungannya satu sama lain. Lanskap dengan komposisi sama dapat memiliki konfigurasi yang berbeda, sehi ngga diperlukan beberapa indeks
Program Patch Anlayst yang kom patibel dengan ArcView 3.x cukup handal untuk menghitung statistik fragmentasi, karena merupakan modifikasi dari program Fragstats dan dapat digunakan untuk m enghitung
statistik spasial, baik file poligon (seperti shape files) dan file raster
(seperti Arc grids) (Elkie et al., 1999). Penjelasan dan contoh aplikasi
program Patch Analyst dibahas secara terpisah pada Bab 11.
DAFTAR PUSTAKA
Barnes, T.G. 2000. Landscape Ecology and Ecosystems M anagement. Cooperative Extension Services, University of Kentucky, College of Agriculture. UK. http://www.ca.uky.edu. Diakses Tangal 24 Februari 2007.
Elkie, P.C., R.S. Rempel and A.P. Carr. 1999. Patch Analyst User’s Manual.
Ontario Ministry of Natureal Resources, Northwest Science & Technology. Thunder Bay. Ontario.
Fahrig, L. 2003. Effects of habitat fragmentation on biodiversity. Annual
Reviews of Ecology and Systematics 34:487–515.
Forman, R. T. T. 1995. Some general principles of landscape and regional
ecology. Landscape Ecology 10:133–142.
Franklin, A.B., B.R. Noon, and T. L.George. 2002. What Is Habitat
Fragmentation? Studies in Avian Biology No. 25:20-29. http://www. humboldt. edu/-tlg2/publications/whatis habitat20fragmentation.Pdf. Diakses Tanggal 11 Mei 2007.
Garrison, B.A. 2005. Fragmentation of Terrestrial Habitat : An Overview for Wildlife Biologists. Trans.W.Sect.Wildl. Soc. 41:2005.
Haila, Y. 2002. A conceptual genealogy of fragmentation research: from island
biogeography to landscape ecology. Ecological Applications 12:321–334.
http://chesapeake.towson.edu/landscape/forestfrag/what_habitat.asp. Diakses Tanggal 1 Mei 2007.
Hunter, M. L., JR. 1997. The biological landscape. Pages 57–67 in K. A. Kohm
and J. Franklin (eds). Creating a Forestry for the 21st Century. Island Press. Washington, D.C.
Kupfer, J.A., G.P. Malanson and S.B. Franklin. 2004. Identifying the
Biodiversity Research Needs Related to Forest Fragmentation. A report prepared for the National Commission on Science for Sustainable Forestry (NCSSF) and funded by the National Council for Science and the Environment (NCSE).
McGarigal, K., and B. J. Marks. 1995. Fragstats: spatial pattern analysis program for quantifying landscape structure. USDA For. Serv. Gen. Tech. Rep. PNW-351. http://www.
3
TEORI BIOGEOGRAFI PULAU vs
FRAGMENTASI HUTAN
PENDAH ULUAN
TIPE-TIPE PULAU
HUBUNGAN SPESIES-AREA
KEPUNAHAN LOKAL DAN PERUBAHAN KOMUNITAS
EFEK ISOLASI
PERBEDAAN ANTARA BIOGEOGRAFI PULAU SEJATI DENGAN PULAU
HABITAT
DAFTAR PUSTAKA
I. PENDAHULUAN
Teori Biogeografi pulau pertama dikemukakan oleh Mac Arthur dan Wilson (1967). Dalam teorinya mereka m emprediksi jumlah spesies yang
mungkin akan bertahan pada suatu pulau yang baru ter bentuk. Dalam
biogeografi pulau dipelajari dan dijelaskan faktor-faktor yang
mempengaruhi keragaman spesies dari sauatu komunitas tertentu. Dalam
konteks ini, “pulau” dapat berupa areal habitat yang dikelilingi oleh areal lain yang tidak sesuai untuk spesies dalam “pulau” tersebut; bukan hanya
pulau sesungguhnya yang dikeli lingi lautan, tetapi juga gunung yang
dikeli lingi oleh gurun pasir, danau yang dikelilingi daratan, dan fragment
hutan yang dikeli lingi oleh lanskap yang terganggu oleh manusia.
Model biogeografi pulau adalah suatu model kaidah umum m engenai
penyebaran keanekaragaman hayati yang menjelaskan hubungan antara
luas areal dan jum lah spesies (species-area relationship). Pulau- pulau yang
luas memi liki jum lah spesies yang lebih banyak dari pada pulau-pulau kecil.
dan komunitas yang lebih banyak, menyediakan kemungkinan isolasi
geografis dan jumlah populasi yang lebi h banyak untuk setiap spesies
sehingga memperbesar kemungkinan spesiasi dan memperkeci l
kemungkinan kepunahan dari spesies yang baru terbentuk atau dari spesies
yang baru datang (Primack et al. 1998). Model biogeografi pulau
memperkirakan jika 50% dari wi layah pulau rusak, sekitar 10% spesies yang hidup di pulau tersebut akan punah. A pabi la spesies ini endemik di wilayah
tersebut, maka spesies akan punah. Apabi la 90% dari habitat rusak, pulau akan kehilangan 50% spesiesnya, dan jika 99% habitatnya hi lang, maka 75%
spesies alami akan hilang (Primack et al. 1998).
Biogeografi pulau dapat diaplikasikan pada fragmentasi lanskap sebagai model bagaimana “pulau-pulau” habitat kecil dapat berpengaruh buruk pada keragaman hayati habitat as linya (Harris 1984). Teori ini sangat
penting dalam mendisain kawasan konservasi karena memberikan panduan kuantitatif tentang luas kawasan dan kesinambungan antara kawasan
konservasi yang bertetangga, berdasarkan karakteristik ekologi di wilayah
tersebut (Diamond 1975).
Menurut teori biogeografi pulau (Mac Arthur & Wi lson 1967),
kekayaan spesies suatu pulau bergantung pada:
Isolasi pulau, karena isolasi mempengaruhi laju kolonisasi. Pulau yang terisolasi atau jauh, memiliki spesies yang lebih sedikit dari pada pulau
yang dekat dengan sumber spesies yang mengkolonisasi. Pulau yang lebih jauh, lebih sedikit didatangi pengkoloni dibandingkan pulau yang
lebih dekat. Jika ada pulau- pulau di antara sumber kolonisasi (daratan utama) dengan pulau, maka dapat berperan sebagai batu loncatan yang besar memiliki jumlah s pesies yang lebih banyak dibandingkan
lebih sedikit refugia (area untuk mengungsi) dan memiliki laju
kepunahan lebih tinggi.
Dinamika kolonisasi dan kepunahan, kolonisasi menggantikan spesies
yang punah (species turnover).
Kekayaan spesies mencerminkan suatu keseimbangan (equilibrium)
antara kolonisasi dan kepunahan.
II.
TIPE-TIPE PULAU
Pulau adalah daratan yang dikeli lingi air. Kondisi fisik pulau biasanya sempit, ti pe habitatnya sedikit, sedangkan pengaruh laut sangat
besar. Pulau digunakan oleh para ahli ilmu pengetahuan alam sebagai
laboratorium lapangan untuk m enelaah masalah biogeografi. Darwin dan
Wallace mem bedakan antara pulau benua (continental island) dan pulau
laut (oceanic island) (Haris 1984). Pulau laut adalah pulau yang belum
pernah berhubungan dengan daratan lainnya (misalnya : P. Nias, P.
Bawean, P. Natuna, P. Belitung, P. Sum ba dan P. Aru). Pulau Benua adalah pulau yang pada masa lam pau mem punyai hubungan dengan daratan atau benua lainnnya karena turunnya permukaan laut (misalnya : Pulau Sumatra,
P. Jawa, P. Kalimantan, P. Papua, P. Simeulue, P. Enggano, P. Buru, P. Kai
dan P. Tanimbar) (Primack et al. 1998).
Ahli lain mem bagi pulau m enjadi tiga tipe yaitu (Ripley 1985):
1. Pulau benua (continental island) yang dulunya merupakan bagian
daratan benua tetapi sekarang terpisah dari daratan induknya, ciri -ciri kehidupan serupa dengan benua/daratan utamanya dan dekat dengan
benua.
2. Pulau gunung berapi (volcanic island) yang berasal dari gunung berapi dan tidak terbentuk sebagai bagian benua. Pulau- pulau gunung berapi
berada di dekat pulau-pulau besar membentuk busur pulau gunung berapi di laut. Pulau-pulau gunung berapi di bagian sisi yang
menghadap benua biasanya dikeli lingi perairan dangkal sedangkan di
3. Pulau samudera (oceanic island) sejati yang terbentuk dari proses
geologi. Biasanya berukuran keci l dan seringkali letaknya jauh dari
daratan utama. Pulau ini dapat berbentuk pulau tunggal atau membentuk kepulauan kecil. Suksesi yang terjadi di pulau ini adalah suksesi primer
Disamping ketiga tipe pulau tersebut juga ada tipe pulau lain yaitu
(Whitten et al., 1988):
1. Pulau atol. Pulau atol terbentuk dari binatang karang yang telah mati, berukuran keci l, biasanya berbentuk cincin dan banyak terdapat di
Lautan Pasifik (Contoh : Pulau Takabonerate dan Tukang Besi).
2. Pulau purba (ancient island). Pulau purba telah terpisah dari daratan utama sejak lama sekali (lebih dari 100 juta tahun) oleh karenanya
memiliki tingkat endemisme tinggi (misalnya P. Madagaskar dan
Kepulauan Selandia Baru).
Pulau-pulau benua memperoleh keanekaragaman hayatinya ketika
pulau-pulau tersebut berhubungan dengan benua. Selanjutnya pulau- pulau
ini memperoleh tam bahan dari hasil migrasi hewan/tum buhan (Primack et
al. 1998). Pulau laut asal mulanya m erupakan substrat kosong, komunitas
hewan berkembang dari kolonisasi awal yang kemudian berkembang semakin kaya dan kompleks (Haris 1984). Dengan perkataan lain,
pulau-pulau laut hanya dapat mem peroleh keanekaragaman hayatinya dari
kemampuan migrasi hewan dan tumbuhan yang berasal dari tem pat-tem pat
lain (Primack et al. 1998). Laju imigasi dan kolonisasi serta faktor-faktor
lingkungan laut sangat mempengaruhi karakteristik biota pulau- pulau laut, setidakya pada tingkat hunian awalnya (Haris 1984).
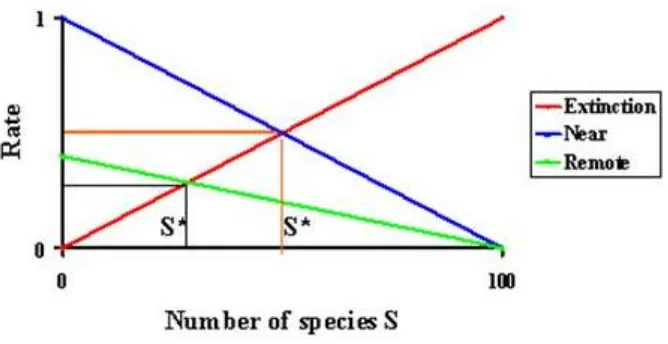
III. HUBUNGAN SPESIES-AREA
Menurut teori biogeografi, jum lah spesies (jum lah equilibrium)
yang terdapat di suatu pulau ditentukan oleh dua faktor, yaitu jarak dari
daratan utama dan ukuran pulau. Keduanya akan mem pengaruhi laju
pulau yang lebih kecil peluang kepunahan lebih besar dari pada pulau- pulau
besar. Pulau-pulau besar memi liki jumlah spesies lebih banyak dari pada
pulau-pulau keci l (Mac Arhtur & Wilson 1967).
Kekayaan spesies suatu pulau juga tergantung pada: (1) isolasi, karena isolasi mempengaruhi laju kolonisasi dan (2) ukuran pulau karena
ukuran pulau m empengaruhi laju kepunahan. Kolonisasi dan kepunahan adalah dinamis, kolonisasi menggantikan spesies yang punah, atau disebut
turn over. Kekayaan spesies suatu pulau menunjukkan keseimbangan
(equilibrium) antara kecepatan kolonisasi dan kecepatan kepunahan (Mac
Arhtur & Wilson 1967).
Hubungan antara jum lah spesies dan luas pulau digambark an
dengan rumus sebagai berikut (Mac Arhtur & Wi lson 1967):
Dimana S = jum lah spesies, A = luas pulau, z dan c adalah konstanta yang
diperlukan untuk menyesuaikan data luas (dalam m2, km2, dll) dengan
jumlah spesies. Hubungan ini dapat dibuat li nier dengan menggunakan log
sehingga rumusnya menjadi (Mac Arhtur & Wilson 1967):
Log S = Log c + z log A
Laju kolonisasi lebih tinggi pada pulau yang dekat dengan daratan
utama karena lebih banyak spesies yang dapat menyeberang laut yang
relatif dekat. Laju kepunahan lebih besar pada pulau yang lebih keci l karena populasi-populasi berukuran lebih keci l dan kemungkinan terkena penyakit dan kejadian merugikan lainnya yang dapat m enghabiskan populasi
atau menurunkannya sampai tingkat yang ti dak viable. Hubungan ini
merupakan prinsip dasar teori biogeografi pulau seperti dtunjukkan pada
Gambar 2.
Hubungan antara ukuran pulau dan jum lah jenis relatif konstan
untuk kelom pok-kelompok hewan atau tum buhan. Secara umum
berkurangnya sepuluh kali ukuran pulau mengurangi setengah jum lah jenis. Jika suatu pulau m endukung lebih sedikit jenis dari yang diharapkan sampai
di bawah garis (Gambar 2), mungkin disebabkan oleh (Whitten et al. 1988):
2. Keseimbangan jumlah jenis belum tercapai.
3. Pulau terdiri atas jumlah habitat yang relatif terbatas atau habitat
tidak mendukung jum lah jenis yang banyak.
4. Pulau sangat jauh dan sulit dikolonisasi.
Jika suatu pulau m endukung lebih banyak jenis dari pada yang
diharapkan sampai di atas garis (Gambar 2), mungkin disebabkan oleh
(Whitten et al. 1988):
1. Jum lah melebihi keseimbangan, namun beberapa jenis pada waktunya
nanti akan hi lang.
2. Pulau sangat kaya akan ti pe-tipe habitat.
3. Pulau merupakan pusat penyebaran jenis kelom pok tertentu.
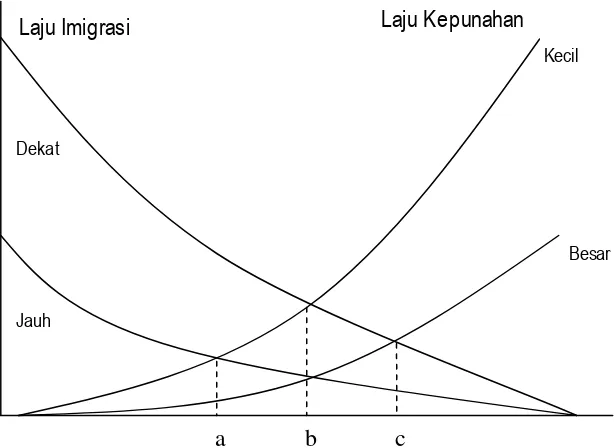
Gambar 2. Jum lah jenis relatif pada (a) pulau-pulau kecil, dekat, (b)
pulau-pulau besar, jauh atau keci l, dekat, dan (c) pulau- pulau besar, dekat (Mac Arthur and Wilson 1967).
Laju Imigrasi
Laju Kepunahan
Jumlah spesies pada sebuah pulau
JauhDekat
Kecil
Besar
Sebagai contoh, untuk total jenis, Sulawesi berada di bawah garis
dari jum lah yang seharusnya untuk hewan dan tumbuhan, tetapi berada di
atas garis untuk mamalia, hal ini mungkin disebabkan oleh sebaran jenis -jenis tikus yang luar biasa dan juga ular. Sulawesi juga berada di atas garis
untuk jenis endemik, hal ini disebabkan oleh sejarah geologinya (Whitten et
al. 1988).
Pengaruh ukuran pulau pada tingkat endemisitas juga sangat tinggi.
Pulau-pulau besar mempunyai jum lah spesies dengan tingkat endemisitas yang jauh lebih tinggi, tetapi korelasinya tidak begitu tampak dengan
kekayaan spesies. Spesies endemik berkorelas i negatif dengan kekayaan
spesies dan berkorelasi positif dengan tingkat isolasi. Disamping itu juga ada perbedaan antara tumbuhan dan hewan dalam tingkat endemisitas.
Tingginya jumlah hewan endemik tidak selalu diikuti oleh tingginya
tumbuhan endemik, tetapi seringkali tingginya tumbuhan endemik diikuti
oleh tingginya hewan endemik (Primack et al. 1998).
Tumbuhan endemik lebi h terpengaruh oleh ukuran pulau dari pada
isolasi geografik. Sebaliknya endemisitas burung sangat bergantung pada isolasi geografik. Pulau- pulau yang jauh dan keci l dapat mempunyai tingkat
endemisitas burung yang ti nggi, tetapi tingkat endemisitas tumbuhannya
dapat rendah (Primack et al. 1998).
IV. KEPUNAHAN LOKAL DAN PERUBAHAN KOMUNITAS
Istilah pulau bisa diartikan mulai dari seb uah pulau keci l, pulau karang (atol) terpenci l yang dikeli lingi laut sampai pada kawasan hutan
yang ditetapkan secara hukum. Suatu komunitas alami yang ada sebagai
sebuah pusat dari suatu habitat regional yang lebih besar akan berisi banyak spesies langka yang tergantung pada sistem yang lebih besar untuk
eksistensinya. Seiring dengan cepatnya perubahan penggunaan lahan di
sekitarnya, maka akan tercipta suatu kantong pulau habitat (habitat patch)
yang terisolasi. Seiring dengan pulau habitat yang semakin terisolasi
Kepunahan mengurangi satwa di setiap pulau habitat sehingga
memainkan peran penting dalam menentukan kom posisi spesies. Tidak
adanya kompensasi kolonisasi, proses tersebut menghasilkan kemiringan
(slope) yang lebih tajam pada kurva species-area. Dengan demikian
perbedaan ni lai kemiringan antara pulau-pulau samudera dan pulau- pulau
benua dapat dijelaskan oleh ketidakseimbangan relatif dari proses kepunahan dan kolo nisasi (Harris 1984).
Picton (1979) dalam Harris (1984) menganalisis perubahan populasi
10 spesies mamalia besar pada 24 wi layah pegunungan kontinental yang
semi terisolasi di Rocky Mountains bagian utara. Luas arealnya bervariasi
dari 11 – 4.480 mi l persegi (29 – 11.600 km2). Persentasi spesies asli yang
telah hi lang selama periode pembagunan pem ukiman, pertanian dan
peternakan ranch, perburuan subsisten, pembalakan dan penam bangan
berbanding terbalik dengan ukuran luas areal. Sementara a real yang lebih kecil kehi langan lebih dari 50% spesies aslinya, areal yang lebih besar
kehilangan sekitar 4% dari spesies yang pernah ada (Pi cton 1979 dalam
Harris 1984). Dengan perkataan lain, faktor-faktor yang meningkatkan perbedaan antara kepunahan dan kolonisasi menyebabkan kemiringan
hubungan spesies-area meningkat seperti ni lai yang ditunjukkan oleh pulau-pulau sebenarnya.
Kepunahan lokal atau hi langnya spesies dari taman nasional, cagar
alam dan pulau- pulau habitat m empengaruhi kelompok spesies tertentu lebih dari lainnya. Secara umum, spesies yang paling rawan untuk hilang
ditunjukkan oleh populasi yang keci l. Populasi kecil dapat dihasilkan oleh keterbatasan sumberdaya spasial dan tem poral (seperti habi tat yang sangat spesialis atau sumberdaya pakan), eksploitasi berlebih atau areal yang
relatif lebi h keci l dari pada dae rah jelajah spesies tersebut. Spesies yang
sangat spesialis, yang menunjukkan jum lah terbesar dari kelas makanannya
(seperti raptor, karnivora darat, pemakan serangga di batang pohon) dan
yang berada pada tropic level yang lebih tinggi biasanya yang pertama
mengalami kepunahan. Walaupun spesies dengan jelajah luas memi liki
potensi kolonisasi yang lebih tinggi, laju kepunahan lokalnya lebih tinggi
Untuk berbagai alasan, fenom ena yang digambarkan di atas sangat
penting untuk mamalia. Beruang grizzly, srigala abu-abu, dan fisher telah
hilang dari Oregon Barat dan wolverine (sejenis anjing hutan) dan Lynx
menjadi sangat jarang. Semua jenis tersebut adalah karnivora puncak.
Olterman & Verts (1982) dalam Harris (1984) meninjau status 41 spesies
mamalia di Oregon. Tujuh dari 8 spesies (88%) yang pernah ada di fragment
bagian barat dan dinyatakan ”punah”, ”jarang” atau ”terancam punah
(endangered)” adalah karnivora. Sebaliknya, lima dari delapan spesies
(62%) dalam kategori ”tidak jarang atau ti dak endagered” adalah herbivora.
Picton (1979) dalam Harris (1984) membatasi analisisnya untuk wialyah
Rocky Mountains pada herbivora besar karena karnivora sudah sangat serius
menurun sehingga analisisnya tidak dapat dijamin. Di Great Basin, spesies
herbivora dengan kebutuhan habitat yang umum (generalis) dan berukuran
tubuh keci l sampai sedang masih bertahan pada sebagian besa r pulau habitat, herbivora dengan ukuran tubuh besar dan/atau mem erlukan
habitat khusus (spesialis) serta karnivora memiliki laju kepunahan yang
lebih tinggi dan hanya bertahan di sedik it areal dari 19 pulau habitat
(Brown 1978 dalam Harris 1984).
V. EFEK ISOLASI
Baik komunitas tropika maupun tem perate sama-sama menghadapi
masalah inbreeding dan hi langnya keragaman genetik yang disebabkan oleh
sub populasi tumbuhan dan satwa yang terisolasi satu sama lain akibat
fragmentasi. Jika jarak yang memisahkan antara dua fragment terlalu
besar dan suatu spesies tidak dapat menyeberanginya, populasi di kedua
fragment tersebut benar-benar terpisahkan. Inbreeding biasa terjadi jika
sub populasi di fragment tersebut keci l. Walaupun hal ini belum ada
catatannya tetapi potensial terjadi (Harvey and Lyles 1986).
Hilangnya keragaman genetik dapat terjadi bahkan tanpa
inbreeding, dan homozigositas yang dihasilkan dalam gen-gen tertentu
dapat membawa suatu spesies pada kepunahan secara perlahan (evolusioner) (Soule 1986). Untuk meningkatkan pertukaran antar
fragment, banyak pengelola satwa m emanfaatkan koridor yang
kekayaan spesies burung-burung yang sedang berkem bangbiak (MacClintock
et al. 1977) dan meningkatkan mobi litas satwa penyebar biji (Haris 1984).
Koridor mungkin bisa menghambat hi langnya keragaman genetik dan
memungkinkan penyebaran spesies antar fragment, tetapi pengurangan
ekosistem alami ke dalam fragment-fragment terus saja terjadi. Penurunan
luas ekosistem alami ini hanya dapat merugikan spesies yang bergantung padanya untuk bertahan hidup.
Salah satu pri nsip biogeografi yang telah lama dianut adalah pentingnya isolasi dalam menentukan karakteristik komunitas biotik.
Walaupun jarak dianggap sebagai ukuran utama dari isolasi, tetapi
konsepnya jauh lebi h kompleks. Mungkin ukuran jarak absolut lebi h mudah dipahami oleh pengelola, tetapi kenyataannya tidak selalu demikian karena
yang dihadapi oleh spesies -spesies tumbuhan dan satwa berbeda. Ada
ribuan perbedaan antara jarak tem puh normal rutin bulanan antara
mamalia keci l seperti m ole (sejenis tikus mondok) dan beruang grizzly atau
srigala. Dengan demikian jarak 0,6 mil bisa 75 kali radius jelajah seekor
tikus mondok, tetapi hanya 1% dari radi us jelajah seekor coughar (Harris
1984).
Alasan kedua mengapa ukuran jarak absolut tidak begitu m enjadi
perhatian, karena peri laku menetap (sedentary) versus berpindah-pindah
(migratory) setiap spesies berbeda. Suatu spesies migratory dengan
anggota yang menjelajahi ribuan ki lometer secara m usiman memi liki peluang lebih tinggi untuk mengkolonisasi pulau terisolasi dalam jarak
tempuhnya dari pada penghuni yang memiliki sifat menetap. Untuk spesies
seperti pika, jarak 1.000 feet (300 m) antara singkapan talus merupakan
rintangan yang sulit dan sangat tidak mungkin menyebar dengan jarak lebih
dari beberapa kilometer dari li ngkungannya (Smith 1974 dalam Harris
1984).
Pertimbangan ketiga adalah spesialisasi habitat dan toleransi
terhadap variasi klimatik, edafik dan perubahan (gradient) vegetasi.
Spesies tertentu (misalnya beruang hitam) menjelajah seluruh wilayah
ketinggian, seluruh g radient kelem baban dan seluruh tahapan suksesi
vegetasi. Spesies lain seperti red tree vole (sejenis tikus keci l) mungkin
1984), hanya ada di tajuk beberapa spesies konifer (kecuali dalam situasi
tidak umum), dan hanya terbatas di Oregon bagian barat dan California
Barat Laut. Dengan demikian tebang habis yang luas dan hutan sekunder, bagi spesies tertentu bisa tidak berpengaruh tetapi bisa mengisolasi secara total bagi spesies lainnya (Harris 1984).
Derajat isolasi harus di lihat sebagai suatu kontinum bagi spesies secara spesifik dan harus di lihat dari aspek biologi spesies tersebut dan
kondisi lingkungannya. Findley & Anderson (1956) dalam Harris (1984)
menyatakan bahwa spesies arboreal seperti tupai dan spesies lain seperti
marten dan fisher yang sangat tergantung pada habi tat hutan, akan sangat
terbatasi oleh sungai besar dan ngarai. Mereka juga menyatakan bahwa penyebaran spesies mamalia di Colorado Rockies merupakan kebalikan dari
ketergantungannya pada hutan. Keberadaan hutan riparian yang dapat
digunakan sebagai koridor penyebaran sangat penting bagi penyebaran
banyak spesies (Findley & Anderson 1956 dalam Harris 1984).
Pengaruh ti dak langsung tambahan dari isolasi adalah sangat
penting bagi satwa, karena sebaran tumbuhan di habitat terisolasi juga masalah bagi sebaran satwa. Isolasi mungkin memiliki pengaruh
mengurangi keragaman sumberdaya tum buhan dan kualitas habitat
(Johnson 1975 dalam Harris 1984). Dengan demikian meskipun suatu
spesies satwa dapat mencapai suatu pulau habitat, habitat tersebut
mungkin tidak cukup beragam dan kompleks untuk mendukung spesies
tersebut. Johnson (1975) dalam Harris (1984) menyimpulkan bahwa
pemiskinan spesies burung dan kerapatan yang lebih rendah spesies di Pegunungan Great Basin diakibatkan oleh pemiskinan tumbuhan dan serangga yang merupakan makanan dan habitatn ya.
Pengaruh gabungan dari semua faktor tam paknya bekerja secara
signifikan pada amfibia dan repti lia, diikuti oleh mamalia, burung penghuni tetap dan burung migran. Dengan kata lain, burung secara proporsional
lebih melimpah di komunitas terisolasi seperti Mary’s Peak dan Steens
Mountains dari pada rata-rata di habitat terfragmentasi lainnya. Isolasi
juga kurang penting untuk menentukan di ti pe habitat apa burung berada di
keberadaan mamalia (Brown 1978 dalam Harris 1984). Secara grafis, efek
isolasi diperlihatkan pada Gambar 3.
VI. PERBEDAAN ANTARA BIOGEOGRAFI PULAU SEJATI
DENGAN PULAU HABITAT
Disamping kemiripan antara biogeografi pulau samudera sejati
dengan pulau-pulau habitat (habitat islands) dan pegunungan, terdapat
beberapa perbedaan yang harus dijelaskan (Wilcox 1980 dalam Harris
1984). Pulau-pulau sejati dikeli lingi oleh media yang bisa berfungsi sebagai
agen penyebaran tetapi tidak akan cukup sebagai habitat spesies daratan.
Puncak pegunungan Mesic dikelilingi oleh lingkungan yang sangat ekstrim
perbedaannya seperti gurun pasir bisa menjadi analogi baik sebagai
perbandingan. Tebang habis secara total di sekeli ling suatu pulau tua juga berbeda dari habitat tua yang tidak sesuai bagi penghuni yang lama tinggal
disana. Hal ini digunakan untuk mem benarkan analogi, tetapi analogi ini tidak memiliki konsistensi kuat (Harris 1984).
Gambar 3. Mekanisme bekerjanya pengaruh isolasi.
sehati selalu dianggap dekat dengan benua yang berperan sebagai sumber
imigran. Benua secara implisit berisi sejumlah besar dan pemasok
jenis-jenis potensial yang tak pernah habis menjadi pengkoloni. Imigrasi dan kolonisasi dari sumber spesies selalu mungkin. Tetapi, beberapa dekade mendatang ketika hutan terdiri dari tanaman m onokultur rotasi pendek dan
ketika banyak spesies terbatasi hanya di cagar alam yang terpisah jauh,
maka tidak ada “benua” sumber spesies yang dapat mengkoloni. Sama halnya, ti dak akan ada benua sumber spesies untuk memperkaya
pulau-pulau tua. Seperti halnya Burgess & Sharpe (1981) dalam Harris (1984) yang
menganggap pulau-pulau tua akan terjadi di sebuah lautan tanpa “benua”
di Amerika Serikat Bagian Timur. Pulau-pulau tersebut m enggambarkan kantong-kantong habitat yang harus berperan sebagai satu-satunya sumber
spesies. Tidak akan ada lagi cadangan spesies yang dapat menjadi sumber
imigrasi (Harris 1984).
DAFTAR PUSTAKA
Amos, W.H. 1980. Wildlife of Islands. H.arry N. Abrams, Incorporated. New York.
Brown, J.H. and M.V. Lomolino. 1998. Biogeography. Sinauer Associates, Inc. Sunderland, Massachusetts.
Diamond, J.M. 1975. The Island Di llema : Lessons of Modern Biogeograpgic Studies for the Design of Natural Reserves. Biological Conservation 7:129-146.
Haris, L.D. 1984. The Fragmented Forest : Island Biogeography Theory and the Preservation of Bioti c Diversity. The University of Chicago Press. Chicago.
Harvey, P.H. and A.M. Lyles. 1986. Inbreeding in natural population of birds and mammals. In M.E. Soule (ed). Conservation Biology : The Science of Scarci ty and Diversity. Sinauer Assoc. Inc., Sunderland, M.A.
http://learning.turner.com/efts/rforest/habfrag.htm. Eco-Link Habitat Fragmnentation. Diakses Tanggal 17-10-2006.
Mac Arthur, R.H. and E.O. Wilson. 1967. The Theory of Island Biogeography. Princeton University Press. Princeton, New Jersey.
MacClintock, L., R.F. Whitcomb and B.L. Whitcom b. 1977. Island Biogeography and Habitat Island of Eastern Forests. II. Evidence for the Value of Corridors and Minimization of Isolation in Preservation of Biotic Diversity. American Birds 31: 6-16.
Primack, R.B., J. Supriatna, M. Indrawan dan P. Kramadirata. 1998. Biologi Konservasi. Yayasan Obor Indonesia. Jakarta.
Ripley, S.D. 1985. Tropical Asia. Time-Life Books Inc. U.S.A.
Whitten, A.J., M. Mustafa dan G.S. Henderson. 1988. The Ecolog y of Sulawesi. Gadjahmada University Press. Yogyakarta.
Wikipedia. 2007. Island Biogeography. http://en.wikipedia.org/ wiki/Island_ biogeography. Diakses Tanggal 30-01-2007.
4
FRAGMENTASI HABITAT DIPANDANG
DALAM SKALA LANSKAP
DINAMIKA PATCH
DESKRIPSI POLA PATCH
PENGARUH UKURAN PATCH
PENGARUH FA KTOR-FAKTOR LAIN
DAFTAR PUSTAKA
I. DINAMIKA
PATCH
Kejadian (occurance) dan distribusi patch- patch dalam suatu
lanskap adalah tidak statis. Mereka terus berubah sepanjang waktu
dibawah kekuatan sistematis seperti suksesi dan erosi serta di bawah kekuatan bencana alam seperti badai dan gangguan jangka pendek oleh
manusia. Gangguan bencana alam umumnya menciptakan patch-patch
dengan suksesi awal di dalam suatu lanskap dan mengurangi patch-patch
dengan suksesi lebih tua yang sudah ada. Hal ini merupakan ciri khas
proses dimana hutan yang sudah matang terfragmentasi menjadi blok -blok.
Gangguan bencana alam juga mempengaruhi ukuran patch, biasanya
meningkatkan patch-patch dengan suksesi baru dan m engurangi ukuran
patch-patch dengan suksesi lama. Tetapi beberapa gangguan bisa lebih
berpengaruh di dalam patch itu sendiri dari pada antar patch.
Heterogenitas dalam patch vegetasi dapat m eningkat dengan m embuka
kanopi, menciptakan pohon tum bang, dan m eningkatkan pancaran cahaya dan panas ke strata kanopi yang lebih rendah dan perm ukaan tanah.