1
Peptida Bioaktif Bahan Alam: RiPPs versus NRPSs
LINDA SUKMARINI
Pusat Penelitian Bioteknologi – LIPI Kompleks Cibinong Science Center
Jl. Raya Jakarta Bogor KM 46, Cibinong, Kab. Bogor, Jawa Barat 16911 Tel. 021 – 8754587/ Fax. 021 8754588
Email: [email protected]
Pendahuluan
Skrining senyawa bioaktif dari bahan alam merupakan salah satu sumber utama penemuan dan pengembangan obat baru. Banyak bahan baku obat berasal dari keragaman hayati seperti bakteri, fungi, metabolit sekunder tanaman ataupun dari sintetik
turunannya. Spektrum bioaktivitas bahan alam juga sangat luas, di antaranya sebagai antibakteri, antivirus, antikanker, peredam aktivitas sistem imun (immunosuppresant), sitotoksik, insektisida, antelmintik (anti-cacing parasitik) maupun
antialodinik (anti-nyeri pada saraf). Di antara jenis bahan alam, peptida termasuk senyawa yang cukup banyak ditemukan, baik yang disintesis di ribosom
maupun yang tidak disintesis di ribosom (non-ribosom) (McIntosh et al., 2009; Newman & Cragg, 2012).
Biosintesis Peptida
Bahan Alam
Pemahaman jalur sintesis senyawa bioaktif sangatlah penting agar dapat memanfaatkan perangkat biosintesis bawaan (endogenous) untuk memproduksi senyawa bioaktif tersebut. Pengetahuan dan kajian yang mendalam mengenai biosintesis peptida bahan alam dapat menfasilitasi rekayasa biosintesis dan/atau peptida bahan alam, baik yang bertujuan untuk meningkatkan jumlah produksi maupun tingkat bioaktivitasnya.Biosintesis peptida bahan alam secara umum dibagi menjadi dua kelompok utama (Gambar 1), yaitu (1) peptida yang disintesis oleh mesin ribosom yang disebut
ribosomally synthesized and post-translationally modified peptide(s) (=RiPP(s) atau
peptida yang disintesis di ribosom dan termodifikasi paska translasi) dan (2)
nonribosomal peptide(s) (=
NRP(s) atau peptida non-ribosom), yang merupakan produk multimodular synthetase peptida
non-ribosom (NRPS(s)) (McIntosh
et al., 2009; Dang &
Süssmuth et al., 2017).
Ribosomally synthesized
and post translationally
modified peptide(s)
(RiPP(s))
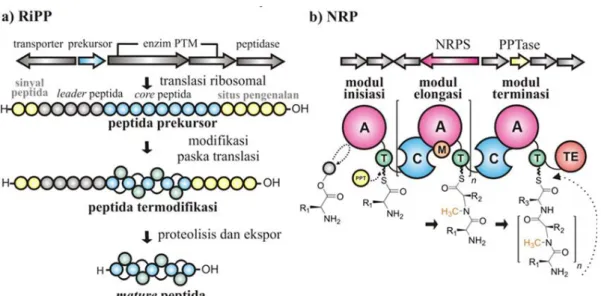
Secara umum, biosintesis RiPP (Gambar 1a) dimulai dengan produksi prekusor peptida (umumnya terdiri dari 20-110 asam amino) yang tidak aktif di ribosom. Prekursor peptida ini
biasanya terdiri dari sekuens awal (leader) peptida pada posisi N-terminus dan sekuens inti (core) peptida pada posisi C-terminus, disertai dengan sekuens opsional yaitu sinyal peptida dan situs pengenalan
peptida (Hallen et al., 2007; Arnison et al., 2013). Motif spesifik pada leader peptida dikenali dan diikat oleh enzim pemodifikasi paska translasi (post-translationally
modifying - PTM) yang
memfasilitasi proses modifikasi peptida pada daerah core peptida.
2
Gambar 1. Skema umum biosintesis peptida bahan alam. a) Peptida yang disintesis melalui
RiPP mengalami modifikasi paska translasi, sedangkan b) biosintesis NRP memanfaatkan mesin perakitan modular NRPS. Modul terdiri dari domain katalisis: adenilasi (A), thiolasi (T) (dimana paska translasi dimodifikasi oleh enzim PPTase (PPT, phosphopantetheine
transferase)), kondensasi (C), dan domain tambahan seperti: metilasi (M) dan terminal thioesterase (Modifikasi dari Dang & Süssmuth, 2017).
Peptida yang telah termodifikasi lalu akan mengalami lisis (proteolisis) oleh peptidase pada bagian
leader peptida. Adapun
proses PTM tambahan dapat ditemukan pada beberapa kasus untuk menghasilkan peptida termodifikasi dengan modifikasi tertentu (Ortega & van der Donk, 2016). Pada akhirnya,
mature peptida diekspor dari
sel sebagai senyawa bioaktif melalui protein transporter, seperti adenosine trifosfat (ATP)-binding cassette (ABC) transporter (Schmidt, 2010).
Terdiri dari kelompok bahan alam yang beragam secara struktur maupun fungsinya, RiPPs dibagi menjadi beberapa famili. Hingga saat ini, diketahui ada lebih dari 20 anggota
famili dengan bobot molekul < 10 kDa berdasarkan
karakteristik struktur dan mesin biosintesisnya (McIntosh et al., 2009; Arnison et al., 2013; Ortega & van der Donk, 2016). Di antara famili RiPPs (Gambar 2), lasso peptida (misalnya capistruin), cyanobactin (misalnya trunkamida), thiopeptida (misalnya thiostrepton A) dan
lanthipeptida (misalnya nisin A) adalah beberapa RiPPs dari bakteri yang memiliki karakter biokimia yang kuat dan jalur biosintesis yang umum (Ortega & van der Donk, 2016; Hudson & Mitchell, 2018). Capistruin diketahui memiliki aktivitas sebagai inhibitor RNA polymerase dan agen antibakteri (Knappe et al.,
2008; Kuznedelov et al., 2011). Cyanobactin trunkamida bersifat sitotoksik terhadap karsinoma kolon dan sel melanoma (Burkhart et al., 2017). Selain memiliki aktivitas antibakteri yang kuat, thiostrepton A
menunjukan potensi sebagai antimalaria dan antikanker (Aminake et al., 2011; Hegde
et al., 2011; Arnison et al.,
2013). Sedangkan nisin A merupakan lanthipeptida yang telah banyak dipelajari dan sejak lama digunakan sebagai antibakteri untuk pengawet produk makanan dan juga untuk produk-produk perawatan hewan ternak (Dischinger et al., 2014). Selain itu, ada pula banyak contoh RiPPs dari fungi, seperti amatoxins,
3 phallotoxins, dan omphalotin
A. Tumbuhan mensintesis RiPPs cyclotida, orbitida,
sedangkan metazoan (cone
snails) mensintesis RiPPs
conopeptida (Arnison et al., 2013; Ramm et al., 2017).
Gambar 2. Struktur kimia peptida dari beberapa famili RiPPs beserta masing-masing bakteri penghasil dan bioaktivitasnya.
4
Non-ribosomal peptide
synthetase(s) (NRPS(s))
Tidak seperti RiPPs dimana satu set asam amino proteinogenik atau non-kanonik digunakan sebagai penyusun utama, jalur non-ribosom untuk biosintesis NRP (Gambar 1b)
memanfaatkan set unit penyusun (building block) yang lebih besar dari hampir 500 asam amino
non-proteinogenik. Komponen penyusunnya termasuk asam amino N-termetilasi, D -asam amino, -asam lemak, dan asam hidroksi, yang disintesis oleh modular enzim yang kompleks dan multifungsi (Pupin et al., 2008; Walsh et al., 2013). Pada dasarnya, perangkat perakitan (assembly) NRPS (Gambar 1b) terdiri dari modul inisiasi, elongasi dan terminasi. Setiap modulnya paling tidak memiliki tiga domain katalitik esensial yaitu, domain A (A), domain thiolasi atau protein peptidyl
carrier (T atau PCP) dan
domain kondensasi (C). Domain A menseleksi monomer asam amino spesifik yang diaktivasi oleh ATP menjadi bentuk amino
acyl adenylate. Protein carrier domain T yang
memiliki gugus
4’-phosphopantetheine (PPant) secara kovalen akan
mengikat monomer
teraktivasi. Sebelum sintesis
peptida terjadi, enzim PPTase
(phosphopantetheine transferase) mentransfer PPTant dari koenzim A ke domain T dari masing-masing modul untuk
membangun domain holo-T paska translasi. Selanjutnya, domain C mengkatalisis gugus amida dari ikatan peptida antara residu yang terkait dengan residu yang berdekatan sehingga terjadi elongasi peptida. Pada perangkat perakitan NRPS yang sederhana, domain C tidak ditemukan pada modul inisiasi karena ketiadaan fungsi untuk kondensasi dengan monomer upstream. Domain thioesterase (TE) hanya terdapat pada modul terminasi yang menjadi karakteristik NRPS dari bakteri. TE menyebabkan pemutusan rantai mature oligopeptida sehingga peptida terlepas dari mesin NRPS. Adapun produk yang dilepaskan dapat berupa senyawa yang linear atau senyawa yang termodifikasi menjadi peptida siklik melalui reaksisiklisasi intramolekuler. Domain tambahan dapat juga ditemukan pada saat
elongasi rantai peptida yang menyebabkan modifikasi struktur ataupun diversifikasi substrat. Beberapa domain tambahan di antaranya, siklisasi (Cy), epimerasi (E), formilasi (F), metilasi (M),
oksidasi (Ox), dan reduksi (R). Setelah proses terminasi NRPS, paska modifikasi dan dekorasi dapat juga terjadi dan merekrut enzim yang berkaitan dengan asilasi, glikosilasi atau halogenasi (Marahiel et al., 1997; Nolan & Walsh, 2009; Süssmuth & Mainz, 2017).
Banyak peptida yang digolongkan sebagai NRPs, terutama peptida yang berbentuk siklik dengan beragam struktur dan ukuran, yang sudah terbukti ataupun berpotensi sebagai bahan baku obat, di
antaranya teixobactin, cyclosporin A, echinocandin B, dan PF1022A (Gambar 3). Teixobactin ditemukan menggunakan teknologi iChip yang dikembangkan untuk bakteri yang sulit dikultivasi (Ling et al., 2015). Antibakteri teixobactin yang bekerja menghambat biosintesis dinding sel bakteri target melalui pengikatan dengan lipid telah diaplikasikan untuk menghambat penolakan transplantasi, sedangkan echinocandin B sebagai antifungi berpotensi digunakan untuk
pengobatan infeksi kronis fungi (Hüttel et al., 2016). Selain itu, PF1022A diketahui memiliki aktivitas
antelmintik yang berpotensi sebagai obat cacing (Sasaki
5
Gambar 3. Struktur kimia peptida NRPs beserta masing-masing bakteri penghasil dan bioaktivitasnya.
Keragaman struktur
RiPPs versus NRPs
Seperti telah disebutkan di atas, secara ringkas RiPPs dan NRPs memiliki beberapa perbedaan mendasar (Tabel 1) dalam membentuk keragaman struktur yang berdampak pada fungsi aktivitas biologinya. Beberapa karakteristik struktur menjadi landasan peran penting peptida bahan alam dalam kimia farmasi obat, di antaranya rigiditas makrosiklik memfasilitasi
target pengikatan, rangka N-metilasi meningkatkan permeabilitas membran dan
D- atau β-asam amino
berkontribusi dalam resistensi proteolitik. Oleh karenanya, kedua peptida bahan alam ini menjadi bahan baku obat dan antibiotik yang potensial (Imani & Freeman, 2018; Huang & Kries, 2019).
Kedua jalur ribosom dan non-ribosom dalam
mensintesis peptida bioaktif memiliki kelebihan dan keterbatasan, namun
beberapa bisa saling
melengkapi. Struktur peptida non-ribosom memiliki
keragaman yang sangat tinggi karena kemampuan enzim synthetase untuk menginkorporasi unit penyusun yang tidak lazim (unusual building block) dan sangat beragam, sedangkan peptida yang disintesis di ribosom mengalami modifikasi paska translasi. Walaupun RiPPs terbatas hanya pada penggunaan 20 asam amino proteinogenik, namun modifikasi paska
6 translasi melengkapi
keterbatasan keragaman strukturnya sehingga mampu mengakses tingkat
keragaman struktur yang serupa dengan NRPs. Banyak motif pada peptida non-ribosom juga ditemukan pada peptida ribosom (Nolan & Walsh, 2009). Salah satu kelebihan utama RiPPs adalah sekuens atau susunan
basanya dapat dimodifikasi melalui manipulasi genetik sederhana dari beberapa kodon. Hal ini memudahkan RiPPs untuk dimodifikasi menjadi senyawa bioaktif baru. Sebaliknya, NRPs memerlukan rekayasa genetik yang cukup kompleks untuk
menginkorporasi modifikasi (McIntosh et al., 2009).
Dengan demikian, bukan hal yang tidak mungkin bahan baku obat dengan struktur NRP dapat diperoleh melalui rekayasa sintesis pada jalur ribosom RiPPs. Pada
akhirnya gap antara RiPPs dan NRPs dengan
keterbatasan maupun kelebihan masing-masing dapat dijembatani melalui kemajuan sintetik biologi.
Tabel 1. Perbandingan RiPPs versus NRPs (Nolan & Walsh, 2009)
RiPPs NRPs
mRNA sebagai cetakan pada ribosom Cetakan HS-pant-P sebagai T domain pada protein modul
20 proteinogenik asam amino
Pertumbuhan rantai peptida (rangka) sebagai peptidyl-O-tRNA
Rantai peptida dilepaskan melalui mekanisme hidrolisis (peptida-CO2-)
>200-500 nonproteinogenik asam amino
Pertumbuhan rantai (rangka) sebagai peptidyl-S-pant-NRPS
Rantai peptida dilepaskan melalui mekanisme hidrolisis, makrosiklisasi ataupun reduksi
Modifikasi paska translasi Tailoring paska perakitan
Pant= phosphopantetheinyl
Daftar Pustaka
Aminake MN, Schoof S, Sologub L, Leubner M, Kirschner M, Arndt HD dan Pradel G. (2011): Thiostrepton and Derivatives Exhibit
Antimalarial and
Gametocytocidal Activity by Dually Targeting Parasite Proteasome and Apicoplast. Antimicrobial Agents and Chemotherapy, 55, 1338-1348.
Arnison PG, Bibb MJ, Bierbaum G, Bowers AA, Bugni TS, Bulaj G, Camarero JA, Campopiano DJ, Challis GL, Clardy J, Cotter PD, Craik DJ, Dawson M, Dittmann E, Donadio S, Dorrestein PC,
7 Entian K-D, Fischbach MA,
Garavelli JS, Göransson U, Gruber CW, Haft DH, Hemscheidt TK, Hertweck C, Hill C, Horswill AR, Jaspars M, Kelly WL, Klinman JP, Kuipers OP, Link AJ, Liu W, Marahiel MA, Mitchell DA, Moll GN, Moore BS, Müller R, Nair SK, Nes IF, Norris GE, Olivera BM, Onaka H, Patchett ML, Piel J, Reaney MJT, Rebuffat S, Ross RP, Sahl H-G, Schmidt EW, Selsted ME, Severinov K, Shen B, Sivonen K, Smith L, Stein T, Süssmuth RD, Tagg JR, Tang G-L, Truman AW, Vederas JC, Walsh CT, Walton JD, Wenzel SC, Willey JM dan van der Donk WA. (2013): Ribosomally Synthesized and Post-translationally Modified Natural Products: Overview and recommendations for a universal nomenclature, Natural Products Reports, 30, 108-160.
Burkhart BJ, Schwalen CJ, Mann G, Naismith JH dan Mitchell DA. (2017):
YcaO-Dependent Posttranslational Amide Activation:
Biosynthesis, structure and function. Chemical Reviews, 105, 685-714.
Dang T dan Süssmuth RD. (2017): Bioactive Peptide Natural Products as Lead Structures for Medicinal Use, Accounts of Chemical Research, 50, 1566-1576. Dischinger J, Chipalu SB dan
Bierbaum G. (2014): Lantibiotics: Promising Candidates for Future Applications in Health Care. International Journal of Medical Microbiology, 304, 51-62.
Hallen HE, Luo H, Scott-Craig JS dan Walton JD. (2007): Gene Family Encoding the Major Toxins of Lethal Amanita Mushrooms. Proceedings of the National Academy of Sciences of the United States of America, 104, 19097-19101.
Hegde NS, Sanders DA, Rodriguez R dan
Balasubramanian S. (2011): The transcription Factor FOXM1 is A Cellular Target of the Natural Product Thiostrepton. Nature Chemistry, 3, 725-731. Huang H-M dan Kries H. (2019):
Unleashing the Potential of Ribosomal and
Nonribosomal Peptide Biosynthesis, Biochemistry, 58, 73-74.
Hudson GA dan Mitchell DA. (2018): RiPP Antibiotics: Biosynthesis and Engineering Potential, Current Opinion in Microbiology, 45, 61-69. Hüttel W, Youssar L, Grüning
BA, Günther S dan Hugentobler KG. (2016): Echinocandin B
Biosynthesis: A Biosynthetic Cluster from Aspergillus nidulans NRRL 8112 and Reassembly of the Subclusters Ecd and Hty from Aspergillus
pachycristatus NRRL 1140 Reveals a Single Coherent Gene Cluster, BMC Genomics, 17, 1-8. Imani AS dan Freeman MS.
(2018): RiPPing Apart the Rules for Peptide Natural Products, Syntetic and Systems Biotechnology, 3, 81-82.
Knappe TA, Linne U, Zirah S, Rebuffat S, Xie X dan
Marahiel MA. (2008): Isolation and Structural Characterization of
Capistruin, A Lasso Peptide Predicted from the Genome Sequence of Burkholderia thailandensis E264, Journal of the American Chemical Society, 130, 11446-11454. Kuznedelov K, Semenova E, Knappe TA, Mukhamedyarov D, Srivastava A, Charttejee S, Ebright RH, Marahiel MA dan Severinov K. (2011): The antibacterial Threaded-Lasso Peptide Capistruin Inhibits Bacterial RNA Polymerase, Journal of Molecular Biology, 412, 842-848.
Ling LL, Schneider T, Peoples AJ, Spoering AL, Engels I,
Conlon BP, Mueller A, Schäberle TF, Hughes DE, Epstein S, Jones M, Lazarides L, Steadman VA, Cohen DR, Felix CR, Fetterman KA, Millett, WP, Nitti AG, Zullo AM, Chen C dan Lewis K. (2015): A New Antibiotic Kills Pathogens without Detectable Resistance, Nature, 517, 455-459.
Marahiel MA, Stachelhaus T dan Mootz HD. (1997): Modular Peptide Synthetases Involved in Nonribosomal Peptide Synthesis. Chemical Reviews, 97, 2651-2674. McIntosh JA, Donia MS dan
Schmidt EW. (2009): Ribosomal Peptide Natural Products: Bridging the Ribosomal and
Nonribosomal Worlds, Natural Product Reports, 26, 537-559.
8 Newman DJ dan Cragg GM.
(2012): Natural Products as Sources of New Drugs over the 30 Years from 1981 to 2010, Journal of Natural Products, 75, 311-335. Nolan EM dan Walsh CT.
(2009): How Nature Morphs Peptide Scaffolds into Antibiotics, ChemBioChem, 10, 34-53.
Ortega MA dan van der Donk WA. (2016): New Insights into Biosynthetic Logic of Ribosomally Synthetized and Post-Translationally
Modified Peptide Natural Products. Cell Chemistry and Biology, 23, 31-44.
Pupin M, Fontaine A, Jacques P dan Kucherov G. (2008): NORINE: A Database of Nonribosomal Peptides, Nucleic Acids Research, 36, 326-331.
Ramm S, Krawczyk B, Mühlenweg A, Poch A, Mösker E dan Süssmuth RD. (2017): A Self-Sacrificing N-methyltransferase is the Precursor of the Fungal Natural Product
Omphalotin, Angewandte Chemie International Edition, 56, 1-5.
Sasaki T, Takagi M, Yaguchi T, Miyadoh S, Okada T dan Koyama M. (1992): A New
Anthelmintic Cyclodepsipeptide, PF1022A. Journal of Antibiotics, 45, 692-697. Schmidt EW. (2010): The
hidden Diversity of
Ribosomal Peptide Natural Products, BMC Biology, 8, 1-4.
Walsh CT, Brien RVO dan Khosla C. (2013):
Nonproteinogenic Amino Acid Building Blocks for Nonribosomal Peptide and Hybrid Polyketide Scaffolds, Angewandte Chemie International Edition, 52, 7098-7124.