TINJAUAN PUSTAKA
Biosintesis asam δ-aminolevulinat
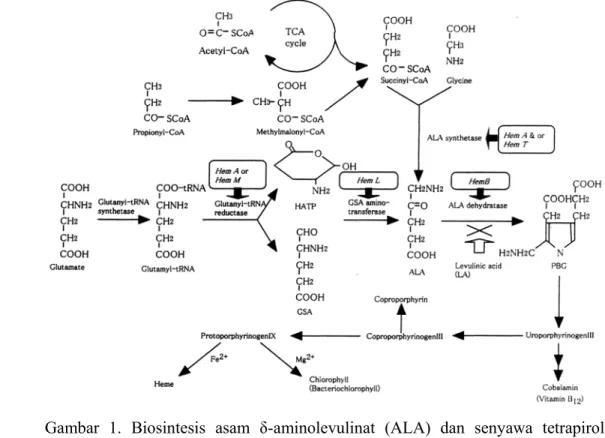
Cincin porfirin merupakan komponen dari berbagai molekul biologis penting termasuk diantaranya haemoglobin, sitokrom, vitamin B12 dan klorofil. Prekursor kunci dari cincin porfirin ini adalah asam δ-aminolevulinat (ALA). Biosintesis ALA dapat dilakukan melalui dua jalur: (1) dari suksinil-CoA dan glisin (Shemin pathway, C-4 pathway), dan (2) dari glutamat (C-5 pathway) seperti yang tersaji dalam Gambar 1.
Gambar 1. Biosintesis asam δ-aminolevulinat (ALA) dan senyawa tetrapirol (Sasaki et al. 2002).
Pada jalur Shemin, suksinil-CoA disediakan melalui siklus TCA. Suksinil-CoA dan glisin dikondensasi dengan bantuan enzim ALA sintase (ALAS). Jalur biosintesis ini digunakan oleh hewan, khamir dan sejumlah bakteri
dalam kelompok α-proteobacteria seperti Rhodobacter, Rhodospirillum dan Rhizobium. Di sejumlah bakteri, suksinil-CoA disintesis dari propionil-CoA melalui jalur biosintesis metilmalonil-CoA (Sasaki et al. 1990). ALAS merupakan enzim pembatas kunci untuk biosintesis senyawa tetrapirol, dan sintesis enzim ini sendiri diatur secara ketat (Sasikala and Ramana 1995) melalui pengaturan secara umpan balik pada gen penyandi HemA atau HemT (Neidle and Kaplan 1993).
Jalur biosintesis C-5 merupakan karakteristik dari tumbuhan tingkat tinggi, Briofita, Sianobakteri dan sejumlah Eubakteria. Jalur biosintesis ini melibatkan tiga buah enzim yaitu Glutamat-tRNA sintase, Glutamil-tRNA reduktase (gtr A/hemA), dan Glutamat 1-semialdehida aminotransferase (hemL). Pada tumbuhan tingkat tinggi, gen penyandi tRNAGlu terdapat di DNA kloroplas, sedangkan ketiga macam enzim yang terlibat dalam biosintensis ALA disandikan oleh DNA inti dan diimpor ke dalam stroma kloroplas setelah disintesis oleh ribosom sitoplasma. Pada fitoflagelata Euglena gracilis ditemukan adanya dua jalur biosintesis (C-4 dan C-5) yang dioperasikan di dalam organel yang berbeda. Jalur C-5 beroperasi di kloroplas dan secara eksklusif bertanggung-jawab untuk sintesis klorofil, sementara di mitokondria, ALA sintase bertanggung-jawab untuk sintesis heme a dari sitokrom c oksidase (Weinstein and Beale 1983).
Aplikasi ALA untuk pertanian
Aplikasi praktis ALA sebagai bioherbisida pertama kali di demonstrasikan oleh Rebeiz et al. (1984). Hasil yang dicapai sangatlah berarti karena ALA dapat bekerja secara selektif, tidak berbahaya dan merupakan bahan yang dapat diuraikan secara biologis. Mekanisme ALA sebagai “photodynamic herbicide” seperti diutarakan oleh Rebeiz et al. (1984) sebagai berikut: tanaman yang diberi
perlakuan dengan ALA dengan jumlah tinggi akan mengakumulasi secara berlebihan molekul protoporfirin IX (PPIX) pada tahap biosintesis klorofilnya. Ketika tanaman terkena cahaya, PPIX yang berlebih akan menghasilkan radikal bebas (O2-), yang akan mengoksidasi asam lemak tak jenuh pada membran sel dan pada akhirnya merusak tanaman. Oleh karena itu ALA dapat digunakan sebagai bahan pengganti yang aman untuk herbisida yang sangat toksik seperti Paraquat. Mekanisme yang sama juga ditunjukkan oleh ALA jika digunakan sebagai bioinsektisida untuk membasmi Trichopusia ni (Rebeiz et al. 1988).
Namun dilain pihak, telah banyak penelitian yang melaporkan efek yang menguntungkan terhadap pertumbuhan berbagai jenis tanaman jika ALA diaplikasikan dalam jumlah rendah (30-100 ppm), seperti meningkatkan berat kering tanaman, peningkatan aktivitas fotosintesis dan efek penghambatan atas respirasi serta mempunyai aktivitas hormonal dalam induksi kallus dan mikropropagasi (Tanaka et al. 1992; Yoshida et al. 1996a; 1996b; Bindu and Vivekanandan 1998a; 1998b; Yoshida et al. 2004). Watanabe et al. (2000) menemukan bahwa aplikasi ALA pada konsentrasi rendah (100 ppm) meningkatkan toleransi terhadap salinitas pada tanaman kapas muda. Hal yang sama juga ditemukan pada tanaman bayam yang diberi perlakuan 0.6 dan 1.8 mM ALA pada media yang mengandung 50 dan 100 mM NaCl, ternyata bukan saja dapat tumbuh dengan baik, yang berarti tanaman menjadi toleran terhadap NaCl, tetapi juga menyebabkan meningkatnya laju fotosintesis (Nishihara et al. 2003). Pada barley, pemberian ALA 30-100 ppm dapat menyebabkan peningkatan hasil sampai 40% yang disebabkan oleh meningkatnya jumlah biji yang dihasilkan tanpa mempengaruhi berat biji. Hal yang sama juga diperlihatkan oleh tanaman
gandum dan padi. Tanaman kacang merah yang diberi perlakuan ALA pada fase awal seperti fase daun primer atau daun pertama, meningkat hasilnya sampai 20-30%, sedangkan perlakuan pada fase pembungaan menurunkan hasil sekitar 10%. Dalam hal ini hasil yang dicapai sangat tergantung pada fase pertumbuhan mana tanaman diberi perlakuan dengan ALA. Pada tanaman bawang dan kentang pemberian ALA pada fase pembentukan umbi dapat meningkatkan jumlah maupun berat umbi yang terbentuk. Wang et al. (2004) melaporkan bahwa jika kecambah melon (Cucumis melo L. Ximiya No. 1) yang ditumbuhkan dengan intensitas penyinaran 150 µmol m-2 s-1 dan diberi aplikasi larutan ALA dapat meningkatkan secara signifikan laju fotosintesis netto (Pn), jumlah CO2 yang difiksasi per jumlah foton (AQY), Efisiensi karboksilasi (CE) dan konduktansi stomata (Gs). Setelah pemberian dengan 10 ml larutan ALA (10 mg l-1 atau 100 mg l-1) per wadah yang diisi dengan 250 g pasir bersih selama 3 hari, Pn daun sekitar 40-200% lebih tinggi dibandingkan kontrol, dan AQY, CE and Gs meningkat secara berturut-turut sekitar 21-271%, 55-210% dan 60-335%. Lebih lanjut, perlakuan ALA meningkatkan kandungan klorofil daun dan tingkat gula terlarut demikian juga dengan laju respirasi dalam keadaan gelap, tetapi menurunkan laju respirasi dalam keadaan terang. Dilain pihak, Setelah kecambah melon yang ditumbuhkan di dalam ruang pertumbuhan diberi perlakuan pendinginan pada suhu 8°C selama 4 jam dan dikembalikan lagi ke suhu 25-30°C selama 2 dan 20 jam, Pn tanaman yang diberi air (kontrol) hanya sekitar 12-18% dan 37-47%, dibandingkan dengan Pn awal sebelum diberi perlakuan pendinginan. Jika kecambah dengan diberi perlakuan sama tetapi diberi ALA (10 mg l-1), Pnnya berturut-turut sekitar 22-38% and 76-101%, dibandingkan
dengan kontrol sebelum pemberian cekaman dingin. Jika lama pendinginan menjadi 6 jam, tanaman yang diberi perlakuan ALA hanya memperlihatkan sedikit gejala nekrosis pada daun sedangkan semua tanaman yang hanya diberi air akan mati, hal ini menyimpulkan bahwa ALA meningkatkan torelansi tanaman terhadap stres dingin dibawah kondisi cahaya rendah.
Ketahanan terhadap salinitas
Tanaman membutuhkan mineral nutrien esensial untuk tumbuh dan berkembang. Namun demikian, kelebihan garam-garam mineral yang terlarut di tanah dapat berbahaya bagi kebanyakan tanaman. Cekaman salinitas merupakan salah satu faktor lingkungan penting yang membatasi pertumbuhan dan produktivitas tanaman pertanian di seluruh dunia. Di perkirakan bahwa masalah salinitas mempengaruhi setidaknya 20% lahan yang dapat ditanami di seluruh dunia dan lebih dari 40% lahan beririgasi dengan berbagai tingkat kerusakan (Rhoades and Loveday 1990). Pada kasus yang ekstrem, produktivitas lahan pertanian tidak dapat lagi menunjang kelangsungan produksi pertanian dan harus ditinggalkan. Di daerah pantai invasi air laut secara periodik langsung menambahkan garam ke tanah. Tanah di daerah semi-arid atau arid, khususnya dengan sistem drainase yang jelek, akumulasi garam sebagai akibat evaporasi air irigasi, meninggalkan deposit garam garam terlarut.
Berdasarkan kemampuannya untuk tumbuh di tempat berkadar garam tinggi, secara tradisional tanaman dikelompokkan atas glikofita dan halofita (Flowers et al. 1977). Halofita toleran terhadap konsentrasi NaCl yang tinggi; beberapa dapat bertahan pada kadar garam dua kali konsentrasi garam pada air laut. Sebagian besar tanaman, termasuk mayoritas spesies tanaman pertanian
masuk ke dalam kelompok glikofita dan tidak dapat mentolerir salinitas yang tinggi. Untuk glikofita, salinitas menyebabkan cekaman ionik, osmotik dan cekaman lanjutan seperti penghambatan pengambilan ion dan cekaman radikal oksida (Zhu 2001a). Toksisitas oleh natrium merupakan cekaman ionik utama yang berhubungan dengan salinitas tinggi. Sebagai tambahan, sejumlah tanaman juga sensitif terhadap klorida, anion utama yang ditemukan pada tanah salin. Pada tanah salin tertentu, toksisitas ion diperparah oleh pH alkalin. Potensial osmotik yang rendah dari larutan salin menghambat pengambilan air dan menyebabkan kekeringan fisiologis. Untuk tanaman halofita yang toleran terhadap toksisitas natrium, cekaman osmotik merupakan penyebab utama terhambatnya pertumbuhan. Pemahaman atas mekanisme toleransi tanaman terhadap salinitas akan membantu secara efektif proses pemuliaan atau rekayasa genetika tanaman yang tahan terhadap salinitas. Beberapa aspek dalam respon tanaman terhadap cekaman salinitas berhubungan sangat erat dengan respon terhadap cekaman kekeringan dan dingin (Zhu 2001b). Studi tentang toleransi tanaman terhadap salinitas akan memberikan pemahaman tentang mekanisme toleransi silang pada cekaman abiotik.
Arabidopsis thaliana merupakan tanaman glikofita yang sensitif terhadap cekaman salinitas dimana dapat terjadi penghambatan pertumbuhan dan kerusakan jaringan yang menuju kematian. Seperti kebanyakan glikofita, sensitivitas Arabidopsis thaliana terhadap cekaman salinitas diperlihatkan pada semua fase perkembangannya. Sebagai contoh, perlakuan selama 8 jam dengan 150 mM NaCl pada fase pembentukan biji menyebabkan terjadinya deposisi callosa dan perubahan yang abnormal pada struktur ovul dan embrio yang
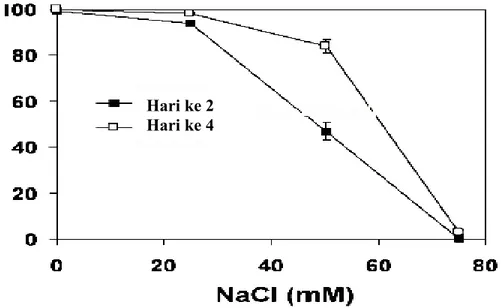
mengindikasikan kematian sel (Sun and Hauser 2001). Sensitivitas Arabidopsis thaliana terhadap cekaman salinitas sangat terlihat pada fase perkecambahan biji dan kecambahnya. Perkecambahan biji Arabidopsis thaliana sangat terganggu pada konsentrasi 75 mM NaCl atau lebih (Gambar 2). Pertumbuhan kecambah juga sangat sensitif terhadap NaCl. Bahkan pada konsentrasi NaCl yang lebih rendah (< 50 mM NaCl) dapat secara nyata mempengaruhi berat basah sedangkan pada konsentrasi yang lebih dari 50 mM NaCl akan menghambat pertumbuhan (Gambar 3) dan bahkan dapat menyebabkan kematian.
Gambar 2. Sensitivitas perkecambahan biji Arabidopsis terhadap NaCl. Biji ecotype C24 (Salah satu ecotype yang sensitif NaCl) ditumbuhkan pada kertas filter yang telah dibasahi dengan larutan NaCl dan diinkubasi pada suhu 4°C selama 2 hari sebelum dipindahkan ke suhu ruang (23°C) dibawah penyinaran cahaya putih untuk perkecambahan (Xiong and Zhu 2002).
Hari ke 2 Hari ke 4
Gambar 3. Sensitivitas tanaman Arabidopsis terhadap NaCl selama fase vegetatif. Kecambah berumur dua minggu (ecotype Columbia) ditumbuhkan ditanah yang di beri larutan 0, 50, 75, dan 100 mM NaCl. Dokumentasi diambil pada minggu ke tiga (Panel atas) dan minggu ke empat (Panel bawah) setelah perlakuan (Xiong and Zhu 2002)
Tanaman yang diadaptasikan dengan konsentrasi garam rendah dapat meningkatkan toleransi tanaman terhadap cekaman salinitas menunjukkan bahwa tanaman glikofita memiliki perangkat untuk membuatnya toleran terhadap salinitas yang mungkin tidak berkerja secara efektif pada keadaan yang tidak teradaptasi. Oleh karena itu, perbedaan toleransi terhadap salinitas antara glikofita dan halofita lebih pada tingkat kuantitatif daripada kualitatif, dan mekanisme dasar toleransi terhadap salinitas kemungkinan bersifat konservatif pada semua spesies tanaman (Xiong and Zhu 2002). Sebagai contoh, gen vakuolar Na+/H+ antiporter AtNHX1 tidak indusibel dibandingkan dengan gen yang homolog pada halofita, dan tingkat ekspresi yang tinggi gen AtNHX1 menggunakan promoter
Minggu ke-3
kuat 35S CaMV dapat secara nyata meningkatkan toleransi Arabidopsis terhadap salinitas (Apse et al.1999; Hamada et al. 2001).
Ion natrium sebenarnya tidak dibutuhkan dalam pertumbuhan oleh sebagian besar tumbuhan. Bahkan tumbuhan sepertinya tidak memiliki sistem transpor khusus untuk pengambilan Na+. Namun demikian, Na+ masih dapat memasuki sel tanaman melalui beberapa jalur. Karena konsentrasi Na+ di tanah biasanya lebih tinggi dibandingkan dengan yang ada di sitosol sel akar, pergerakan Na+ kedalam sel akar dapat terjadi secara pasif. Bukti yang didapat baru-baru ini menunjukkan bahwa sebagian besar Na+ memasuki sel akar melalui saluran kation tidak tergantung voltase (VIC) atau yang dikenal dengan nama lain saluran kation tidak selektif (NSCC) (Amtmann and Sanders 1999; Demidchik and Tester 2002).
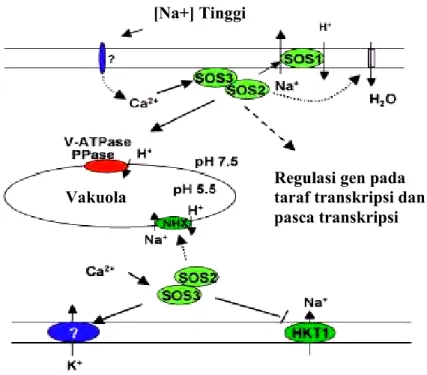
Selain itu, disebabkan oleh kemiripan antara ion Na+ and K+, transporter K+ HKT1 kemungkinan merupakan pintu masuknya ion Na+ ke dalam sel akar karena transporter ini mempunyai afinitas terhadap Na+ pada saat konsentrasi Na+ diluar sel tinggi (Rubio et al. 1995). Gen yang homolog pada Arabidopsis, AtHKT1, memperantarai pengambilan Na+ ketika diekspresikan di Saccharomyces cerevisiae atau oosit Xenopus (Uozumi et al. 2000). Mutan supresor hkt1 mempunyai kandungan Na+ yang lebih rendah, menunjukkan bahwa AtHKT1 memerantarai pengambilan Na+ kedalam tanaman. Studi ini juga menunjukkan bahwa SOS3 pada tipe liar, bekerja bersama-sama dengan SOS2, mungkin menekan aktivitas AtHKT1 dalam memungkinkan Na+ masuk kedalam sel akar (Gambar 4).
Tanaman mempunyai sejumlah cara untuk mengatasi cekaman ionik yang diakibatkan oleh salinitas yang tinggi termasuk diantaranya membatasi pengambilan, meningkatkan pengeluaran dan lokalisasi Na+ dalam vakuola dan
mengontrol transport jarak jauh ke daerah daun. Kelebihan Na+ pada permukaan
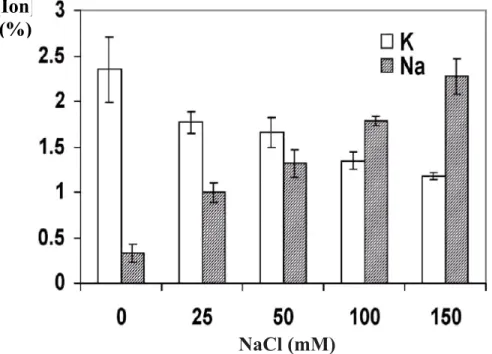
akar akan menganggu proses pengambilan K+ yang sangat penting dalam menjaga turgor sel, potensial membran dan aktivitas sejumlah enzim (Lazof and Bernstein 1999). Karena kemiripan sifat fisiko-kimia Na+ dan K+, Na+ pada konsentrasi tinggi mempunyai efek inhibisi yang kuat pada proses pengambilan ion kalium oleh akar. Sebagai contoh, pengambilan K+ melalui KUP1 Arabidopsis dihambat oleh konsentrasi 5 mM NaCl atau lebih tinggi (Kim et al. 1998; Fu and Luan 1998). Tanaman mengunakan sistem pengambilan K+ baik yang berafinitas tinggi maupun rendah. Ion natrium mempunyai efek yang lebih merusak pada sistem yang berafinitas rendah yang memiliki selektivitas K+/N+ yang rendah. Pada keadaan cekaman salinitas, merupakan hal yang penting bagi tanaman untuk mengoperasikan sistem pengambilan K+ yang berafinitas tinggi dalam rangka menjaga ketersediaan K+ yang dibutuhkannya. Sudah merupakan fenomena yang umum bahwa perlakuan NaCl pada tanaman menyebabkan menurunnya kandungan K+ (Gambar 5), yang secara parsial bertanggung jawab terhadap
Gambar 4. SOS pathway berperanan dalam homeostasis ion pada saat stres oleh salinitas. Konsentrasi NaCl ekstraselular yang tinggi akan memicu meningkatnya konsentrasi Ca2+ di sitosol. Sensor Ca2+ SOS3 yang menanggapi signal Ca2+ ini
akan berinteraksi dan mengaktifkan protein kinase SOS2. SOS2 yang teraktivasi kemudian akan mengatur aktivitas transporter ion atau aktivator transkripsional untuk menjaga kesetimbangan ion atau ekspresi gen. Target protein SOS2 termasuk Na+/H+ antiporter (SOS1), penukar Na+/H+ vakuola ( NHX), dan Na+ transporter (HKT1). Target potensial lainnya termasuk ATPase tonoplas dan pirofosfatase, saluran air, dan kalium transporter (Zhu 2003).
.Pengeluaran Na+ dari sel merupakan cara cepat untuk menghindari akumulasi ion natrium di dalam sitosol. Di perkirakan bahwa pengeluaran ion natrium mungkin lebih penting pada sel tertentu, misalnya sel epidermal akar. Hal ini disebabkan karena sebagian besar sel lain di tanaman dikelilingi sel tetangganya dan pengeluaran Na+ akan menimbulkan masalah bagi sel tetangganya. Pada analisis ekspresi promoter SOS1-GUS menunjukkan bahwa gen Na+/ H+ antiporter terekspresi dengan baik hanya pada sel epidermal yang mengelilingi ujung akar dan di sel parenkima yang menyelubungi xylem diseluruh tanaman (Shi et al. 2002a). Pada Arabidopsis thaliana, ekstrusi Na+ terjadi
[Na+] Tinggi
Regulasi gen pada taraf transkripsi dan pasca transkripsi Vakuola
melalui Na+/ H+ antiporter SOS1 yang terlokalisasi di membran plasma (Shi et al. 2000a; Shi et al. 2002b). Mutasi pada SOS1 menyebabkan tanaman mutan menjadi sangat sensitif terhadap ion natrium (Wu et al. 1996).
Gambar 5. Stres oleh salinitas menghambat pengambilan ion K pada Arabidopsis. Dengan meningkatnya NaCl di dalam medium kultur, kandungan Na+ di dalam tanaman meningkat sedangkan kandungan K+ menurun. Kecambah Arabidopsis (ecotype Columbia) ditumbuhkan di larutan ½MS (pH 5.3) selama 2 minggu yang diberi perlakuan NaCl. Kecambah ditumbuhkan selama 3 hari sebelum dipanen dan dianalisa kandungan ionnya (berdasarkan berat kering) (Xiong and Zhu 2002)
Toleransi terhadap cekaman salinitas merupakan suatu karakter yang komplek melibatkan berbagai respon selular terhadap cekaman osmotik dan ionik, demikian juga efek cekaman sekunder. Banyak penelitian yang mempelajari berbagai efek cekaman salinitas dan melindungi tanaman dari reactive oxygen species (ROS) sepertinya merupakan salah satu komponen penting dari karakter tanaman yang toleran. Konsentrasi NaCl yang tinggi biasanya akan mengganggu transpor elektron selular pada berbagai organel subselular dan menyebabkan terbentuknya ROS seperti singlet oxygen, superoksida, hidrogen peroksida dan
[Ion]
(%)
radikal hidroksil (Noctor and Foyer 1998). Kelebihan ROS memicu reaksi fitotoksik seperti peroksidasi lipid, degradasi protein dan mutasi DNA. Sumber utama ROS adalah radikal superoksida, yang dibentuk diberbagai organel subselular seperti mitokondria, kloroplas dan sitoplasma melalui sejumlah jalur metabolik selama kondisi cekaman oksidatif (Noctor and Foyer 1998). Tingkat kerusakan sel akibat proses peroksidasi ditentukan oleh potensi sistem enzim antioksidatif (Gambar 6).
Untuk mengontrol jumlah ROS dan melindungi sel dibawah kondisi cekaman, sel tanaman mempunyai sejumlah enzim untuk mendetoksikasi ROS (superoksida dismutase, catalase, peroksidase dan glutathion peroksidase), detoksikasi produk peroksidasi lipid (glutathion S-transferase, phospholipid-hidroperoksida glutathion peroksidase dan askorbat peroksidase) dan antioksidan berberat molekul rendah (antosianin, α-tokoferol, askorbat, glutathion dan senyawa polifenol) serta seluruh enzim yang dibutuhkan untuk meregenerasi bentuk aktif dari antioksidan (monodehidroaskorbat reduktase, dehidroaskorbat reduktase dan glutathion reduktase, Gambar 7 (May et al. 1998; Blokhina et al. 2003)). Di dalam sel, superoksida dismutase (SOD) merupakan pertahanan pertama terhadap ROS yang mengkatalisis dismutasi anion superoksida menjadi oksigen dan hidrogen peroksida (H2O2). O2- dihasilkan disetiap tempat dimana terdapat rantai transfer elektron dan oleh karena itu aktivasi O2 dapat terjadi di organel yang berbeda (Elstner 1991), termasuk mitokondria, kloroplas, mikrosom, glikosisom, peroksisom, apoplas dan sitosol. Oleh karena itu SOD dapat ditemukan di semua lokasi subselular. Kloroplas, mitokondria dan peroksisom merupakan tempat utama bagi pembentuk ROS (Fridovich 1986).
SOD APX GPX
Berdasarkan penggunaan kofaktor logam oleh enzim, SOD dapat dikelompokkan menjadi tiga: Fe-SOD, Mn-SOD dan Cu-Zn SOD dan ketiga SOD ini terletak di kompartemen sel yang berbeda. Fe-SOD terdapat di kloroplas, Mn-SOD di mitokondria dan peroxisome, dan Cu-Zn SOD terdapat di kloropls, sitosol dan kemungkinan di ruang ekstraselular (Alscher et al. 2002). Untuk mendetoksikasi H2O2 yang terbentuk, tanaman mengembangkan enzim antioksidatif seperti peroksidase dan katalase. Peroksidase tanaman menggunakan substrat yang berbeda seperti guaikol, askorbat dan glutathion untuk mendetoksikasi H2O2 intraselular. Berdasarkan spesifisitas substrat peroksidase dikelompokkan menjadi guaikol peroksidase, askorbat peroksidase and glutathion peroksidase. H2O2 yang dihasilkan di glyoxysom dan peroxisom dalam proses β-oksidasi didetoksikasi menjadi H2O oleh enzim katalase, sedangkan di organel subselular lain H2O2 dikonversi menjadi H2O oleh enzim askorbat peroksidase dan glutathion peroksidase (Halliwell and Gutteridge 1989; Sen-Gupta et al. 1993). .
Gambar 6. Model bagi induksi reactive oxygen species (radikal superoksida, hydrogen peroksida dan hidroksil) yang terjadi pada saat perlakuan dengan NaCl dan peranan enzim antioksidatif superoksida dismutase (SOD), askorbat peroksidase (APX) dan glutathion peroksidase (GPX) dalam mendetoksikasi radikal bebas superoksida, hidrogen peroksida dan hidrosil (Xiong and Zhu 2002).
Radikal Superoksida
Gambar 7. Halliwell-Asada pathway (Siklus Askorbat-glutathione). APX, ascorbat-peroksidase; MDHAR, monodehidroaskorbat reduktase; DHAR, dehidroaskorbat reduktase; GR, glutathion reduktase (May et al 1998)
Chlorophyll A Oxygenase (CAO)
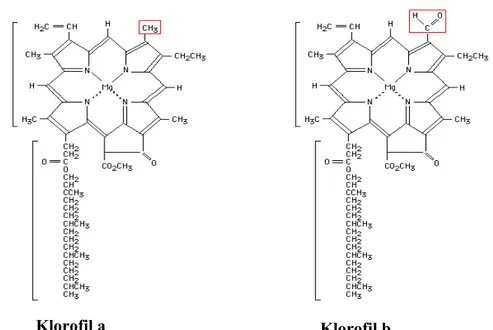
Klorofil merupakan inti dari proses fotosintesis dalam pemanenan energi cahaya dan pengkonversiannya menjadi energi kimia. Tumbuhan darat, algae hijau dan proklorofita menghasilkan klorofil a dan klorofil b (Gambar 8). Klorofil a sendiri terdapat di pusat reaksi dari komplek fotosistem yang memiliki komposisi dan organisasi yang sangat konservatif. Di lain pihak, klorofil a dan klorofil b merupakan komponen antena penangkap cahaya periferal. Komplek antena penangkap cahaya memperlihatkan perubahan yang terkontrol dalam beradaptasi terhadap berbagai kondisi pertumbuhan, yang memungkinkan penggunaan yang optimal dari cahaya yang tersedia. Diketahui bahwa rasio klorofil a dan b tinggi pada kondisi pertumbuhan dengan cahaya terang dibandingkan pada kondisi pertumbuhan cahaya redup, dimana terjadi pembesaran ukuran komplek antena penangkap cahaya pada kondisi cahaya redup (Gambar 9) (Tanaka and Tanaka 2005). Oleh karena itu regulasi biosintesis klorofil b menjadi sangat penting dalam memahami mekanisme adaptasi tanaman terhadap berbagai intensitas cahaya.
Gambar 8. Struktur klorofil A dan klorofil B. Perbedaan terletak pada gugus dalam kotak merah. Pada klorofil a berupa gugus metil sedangkan klorofil b aldehida
Gambar 9. Struktur fotosistem II pada keadaan cahaya redup (atas) atau cahaya terang (tengah) pada tanaman tingkat tinggi tipe liar dan mutan tanpa klorofil b (bawah).
A. Protein-protein pusat reaksi (Chl a) B. Protein-protein antena utama (Chl a) C. Protein-protein antena periferal minor (Chl a, Chl b)
D. Protein-protein antena periferal mayor (Chl a, Chl b) Kondisi intensitas cahaya rendah Kondisi intensitas cahaya tinggi Mutan minus Chl b Klorofil a Klorofil b
Terdapat dua hipotesis mengenai regulasi ukuran antena penangkap cahaya: regulasi oleh ekspresi gen-gen Lhc dan regulasi oleh biosintesis klorofil b. Hipotesis pertama berdasarkan penemuan bahwa pola ekspresi gen-gen tersebut berhubungan erat dengan ukuran antena (Maxwell et al. 1995; Escoubas et al. 1995). Namun demikian, terdapat sejumlah bukti yang berlawanan dengan hipotesis pertama dan cenderung mendukung hipotesis kedua. Pertama, pada sejumlah mutan chl b-less, lebih banyak protein LHC yang hilang ketika jumlah klorofil b menurun. Pada Arabidopsis thaliana mutan yang tidak bisa sama sekali membentuk klorofil b, tidak ada protein LHC yang terbentuk (Espineda 1999), sementara itu di barley mutan yang masih mengandung sejumlah kecil klorofil b, satu atau lebih protein masih dapat ditemui (Bossmann et al 1997). Proses transkripsi, translasi dan transpor protein LHC berjalan normal, menunjukkan bahwa stabilitas protein LHC yang mengalami gangguan (Bellemare, 1982).
Kedua, prekursor protein LHC membutuhkan klorofil b untuk kestabilan insersi ke dalam membran (Kuttkat et al. 1997). Ketiga, telah dibuktikan bahwa mRNA Lhc dengan jumlah kurang dari 5% dari keadaan normalnya sudah cukup untuk menghasilkan ukuran antena yang terbesar sekalipun. Flachman and Kühlbrandt (1997) dan Flachman (1995) melakukan transformasi tanaman tembakau dengan antisense gen Lhc. Di tanaman ini ekspresi gen Lhc mengalami tekanan oleh ekspresi antisensenya. Uniknya, tingkat protein LHC tidak mengalami perubahan bahkan pada tanaman transgenik yang hanya mengekspresikan 5% mRNA dengan orientasi sense. Hasil ini menunjukkan bahwa klorofil b dibutuhkan untuk kestabilan dan/atau insersi protein LHC ke
dalam membran dan transkripsi gen-gen Lhc bukanlah faktor utama yang mengatur ukuran antena penangkap cahaya.
Transformasi Arabidopsis dengan gen CAO dibawah kontrol promotor 35S CaMV menunjukkan bahwa ukuran antena pada fotosistem II meningkat antara 10-20% jika dibandingkan dengan tanaman tipe liarnya, hal ini memperlihatkan bahwa biosintesis klorofil b dapat mengatur ukuran antena penangkap cahaya pada tanaman (Tanaka et al. 2001).
Sebelum gen yang bertanggung-jawab dalam biosintesis klorofil b ditemukan, sangat sedikit yang diketahui tentang jalur biosintesis klorofil ini. Identifikasi gen untuk biosintesis klorofil b dilakukan dengan mutagenesis insersi pada Chlamydomonas reinhardtii yang menghasilkan sejumlah chl b-less mutan. Hasil analisis genom yang termutasi berhasil mengidentifikasi sebuah gen yang diperkirakan suatu monooxygenase yang memiliki [2Fe-2S] Rieske center motif dan mononuclear iron-binding motif (Tanaka et al. 1998). Kloning gen homolog yang dilakukan menggunakan PCR dan pencarian dari database pada alga laut prokariotik, Prochlorothrix dan Prochlorococcus, dan tanaman tingkat tinggi, Arabidopsis thaliana menunjukkan bahwa gen ini konservatif dari prokariot sampai tanaman tingkat tinggi (Tomitani et al. 1999).
Protein dari gen yang diklon dari Arabidopsis thaliana dan diproduksi di Escherichia coli mengkatalisis dua tahapan reaksi dari chlorophyllide (Chlide) a menjadi Chlide b (Chlorophyllide merupakan prekursor tidak teresterifikasi dari klorofil). Berdasarkan aktivitas ini, enzim ini dinamakan CAO (Chlorophyllide a monooxygenase) (Tanaka and Tanaka 2005). Hal yang menarik adalah bahwa enzim ini hanya mengenali chlorophyllide a, artinya bahwa klorofil a
pertama-tama harus di de-esterifikasi kembali menjadi clorophyllide a dan kemudian dikonversi menjadi clorophyllide b dan kemudian diesterifikasi kembali menjadi klorofil b (Oster et al. 2000).
Penemuan yang tidak terduga lainnya adalah bahwa CAO mengkatalisis dua tahapan reaksi. Tahapan pertama adalah oksigenasi gugus metil menjadi gugus hidroksil menghasilkan 7-hydroxymethyl-chlorophyll. Pada tahap kedua gugus hidroksil dioksigenasi lagi menjadi gugus formil menghasilkan klorofil b. Kedua tahapan ini membutuhkan NADPH dan ferredoxin sebagai tenaga pereduksi (Tanaka and Tanaka 2005).
Walaupun sudah secara jelas dibuktikan dari eksperimen menggunakan tanaman yang mengekspresikan secara berlebihan gen CAO bahwa biosintesis klorofil b memegang peranan penting dalam mengatur ukuran antena, namun jalur degradatif klorofil b tetap merupakan hal yang menarik untuk diketahui. Selama ini jalur degradatif klorofil hanya diketahui untuk klorofil a saja, karena produk degradatif klorofil a dapat teramati. Alasan mengapa tidak terdapat produk degradatif klorofil b baru dapat diketahui setelah ditemukannya jalur konversi klorofil b menjadi klorofil a (Ito et al. 1993). Sekarang telah diketahui bahwa klorofil b pertama-tama dikonversi menjadi 7-hydroxymethyl-chl dan kemudian direduksi menjadi klorofil a (Ito et al. 1994; Ito et al. 1996; Ito and Tanaka 1996; Scheumann et al. 1996a; 1996b; Scheumann et al, 1998). Tahap pertama dan kedua membutuhkan NADPH dan ferredoxin sebagai tenaga pereduksi (Scheumann et al. 1998). Aktivitas dari tahap pertama meningkat selama proses senescen pada daun (Scheumann et al. 1999). Penelitian yang dilakukan menunjukkan bahwa klorofil a yang dibentuk dari klorofil b dapat diinsersikan
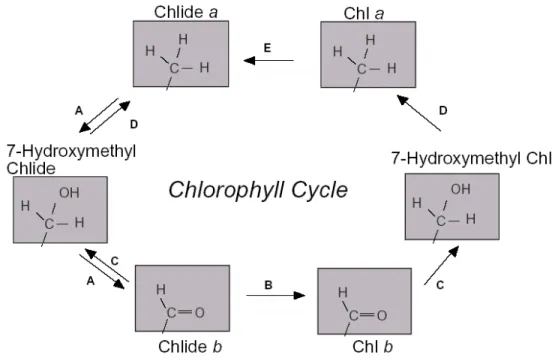
kembali ke dalam protein pengikat klorofil, yang berarti bahwa jalur ini juga berfungsi untuk menggunakan kembali klorofil b (Ohtsuka 1997). Mungkin saja konversi klorofil b menjadi klorofil a dan aktivitas CAO dikoordinasi untuk mengatur kesetimbangan antara klorofil a dan klorofil b. Oleh karena itu interkonversi klorofil a dan klorofil b disebut "chlorophyll cycle" dan diperkirakan siklus ini memainkan peranan penting dalam mengatur ukuran antena (Gambar 10) (Tanaka et al. 1998; Oster et al. 2000).
Gambar 10. Sebuah model yang diusulkan untuk siklus klorofil. Klorofil a (atas kanan) di dephytilasi menjadi chlorophyllide (Chlide) a (atas kiri) chlorophyllase (E) dan kemudian dikonversi menjadi Chlide b (bawah kiri) melalui 7-hydroxymethyl-chlorophyllide (tengah kiri) oleh CAO (A). Chlide b dapat di konversi kembali menjadi Chlide a melalui 7-hydroxymethyl-chlorophyllide oleh Chl b reductase (C) dan 7-hydroxymethyl-chlorophyllide reductase (D). Alternatif lain, Chlide b di phytilasi menjadi klorofil b (bawah kanan) oleh chlorophyll synthase (B) dan kemudian dikonversi menjadi klorofil a melalui hydroxymethyl-Chl (tengah kanan) oleh Chl b reduktase (C) and 7-hydroxymethyl-chlorophyllide reduktase (D) (Tanaka and Tanaka 2005).
Salah satu tantangan kedepan dalam penelitian tentang klorofil b adalah identifikasi protein atau gen yang terlibat dalam konversi klorofil b menjadi klorofil a. Dengan selesainya sekuensing genom Arabidopsis thaliana memungkinkan hal ini dapat dilakukan lebih mudah (The Arabidopsis Genome Initiative 2000). Dengan mengklon gen yang terlibat dalam konversi klorofil b menjadi klorofil a, memungkinkan dilakukannya pengujian hipotesis bahwa degradasi klorofil memicu terjadinya proses senescen (Vincentini et al. 1995; Matile et al. 1996). Jika hipotesis ini benar, memungkinkan ditundanya proses senescen tanaman melalui supresi aktivitas konversi klorofil b menjadi klorofil a dengan ekspresi berlebih antisense gen yang mengkode enzim pereduksi klorofil b.
Transformasi yang diperantarai Agrobacterium:
Agrobacterium tumefaciens mempunyai peranan penting dalam perkembangan rekayasa genetika tanaman dan penelitian dasar dalam biologi molekular. Hampir 80% tanaman transgenik dihasilkan melalui cara ini. Pada awalnya, orang percaya bahwa hanya tanaman dikotil, gymnosperma dan beberapa spesies monokotil saja yang dapat ditransformasi menggunakan bakteri ini; namun hasil yang dicapai baru-baru ini merubah secara total pandangan ini yang diperlihatkan oleh beberapa spesies “rekalsitran” yang tidak termasuk inang alami seperti monokotil dan fungi ternyata dapat ditransformasi (Chan et al. 1993;Bundock et al. 1995). Sebagai tambahan, sel yang tertransformasi biasanya membawa jumlah kopi T-DNA terintregrasi dalam jumlah rendah didalam genomnya dengan sedikit atau tanpa penantaan kembali, dan DNA berukuran sangat besar dapat ditransformasikan ke dalam tanaman.
Dasar molekular transformasi genetik sel tanaman oleh Agrobacterium tumefaciens adalah transfer dari bakteri dan integrasi ke dalam genom tanaman suatu bagian dari plasmid Ti (tumor-inducing) atau Ri (rhizogenic-inducing) (Gambar 11). Transfer T-DNA diperantarai oleh produk yang disandikan oleh suatu daerah vir yang berukuran 30-40 kb pada plasmid Ti. Daerah ini terdiri sedikitnya 6 operon esensial (vir A, vir B, vir C, vir D, vir E, vir G) dan dua non esensial (vir F, vir H). Operon yang konstitutif hanya operon vir A dan vir G, yang menyandikan sistem dua komponen (VirA-VirG) yang akan mengaktifkan transkripsi gen-gen vir lainnya. Vir A merupakan suatu protein sensor dimer transmembran yang dapat menditeksi molekul signal, terutama senyawa fenolik kecil yang dilepaskan oleh tanaman yang terluka (Pan et al. 1993). Signal lain bagi aktivasi Vir A diantaranya pH asam, senyawa fenolik seperti acetosyringon (Winans et al. 1992) dan monosakarida jenis tertentu yang bekerja secara sinergis dengan senyawa fenolik (Ankenbauer et al. 1990; Cangelosi et al. 1990; Shimoda et al. 1990a; Doty et al. 1996).
Protein VirA secara struktural dapat dibagi menjadi tiga domain: domain periplasma (input) dan dua domain transmembran (TM1 dan TM2). Domain TM1 dan TM2 bertindak sebagai suatu transmiter (signaling) dan penerima (sensor) (Parkinson 1993). Domain periplasma sangat penting untuk deteksi monosakarida (Chang and Winans 1992). TM2 merupakan suatu domain kinase dan mempunyai peranan yang penting dalam aktivasi VirA yaitu memfosforilasi dirinya sendiri pada residu His-474 (Huang et al. 1990; Jin et al. 1990a; 1990b) sebagai respon atas molekul signal dari tanaman yang terluka. Deteksi monosakarida oleh VirA merupakan sistem amplifikasi penting dan respon terhadap jumlah senyawa
fenolik yang rendah. Induksi sistem ini hanya mungkin melalui protein pengikat gula (glukosa/galaktosa) periplasma ChvE (Ankenbauer and Nester 1990; Cangelosi et al. 1990), yang berinteraksi dengan VirA (Shimoda et al. 1990b; Chang and Winans 1992; Turk et al. 1993). VirA yang teraktivasi mempunyai kemampuan untuk mentransfer gugus fosfatnya ke residu aspartat dari suatu protein pengikat DNA sitoplasma VirG (Jin et al. 1990a; 1990b; Pan et al., 1993). VirG berfungsi sebagai faktor transkripsional yang mengatur ekspresi gen-gen vir lainnya ketika terfosforilasi oleh VirA (Jin et al. 1990a; 1990b). Daerah C-terminal bertanggung-jawab untuk aktivitas pengikatan DNA sedangkan daerah N-terminal adalah domain fosforilasi dan mempunyai kemiripan dengan domain penerima signal VirA (sensor). Aktivasi sistem vir juga tergantung pada faktor luar seperti suhu dan pH. Pada suhu yang melebihi 32°C, gen-gen vir tidak akan terekspresi karena perubahan konformasi dalam proses pelipatan protein VirA yang menyebabkan protein ini tidak aktif (Jin et al. 1993).
VirD1 dan VirD2 bertanggung-jawab untuk pembentukan strand, T-DNA utas tunggal, dengan mengenali dan memotong secara spesifik utas bagian bawah T-DNA pada kedua pembatas, yang mana batas kanan merupakan titik awal dan sangat penting dalam proses ini. Setelah pemotongan, VirD2 tetap terikat secara kovalen pada ujung 5’ utas T, membentuk suatu komplek dengan karakter polar dimana ujung 5’ akan bertindak sebagai bagian kepala dalam proses transfer. Komplek VirD2/utas T dan protein pengikat DNA utas tunggal lain yang disebut VirE2 dipercaya ditransferkan secara terpisah ke dalam sel tanaman melalui suatu struktur yang mirip pilus yang terdiri atas suatu komplek protein VirB (Fullner 1996) and VirE2 selama didalam sel bakteri tidak dapat
menempel ke utas T karena adanya protein chaperonin yang disebut VirE1 (Deng et al. 1999; Sundberg and Ream 1999). Sekali memasuki sel tanaman, molekul VirE2 akan bersama-sama menempel ke utas T, membentuk komplek T (Sundberg et al. 1996), yang kemudian ditargetkan ke dalam nukleus oleh nuclear target signals (NLS) yang terdapat pada VirD2 dan VirE2, dimana T-DNA diintegrasikan secara acak ke dalam genom tanaman dengan kopi tunggal atau ganda. Mekanisme yang terlibat dalam integrasi T-DNA belum terkarakterisasi, namun diperkirakan melibatkan proses rekombinasi yang tidak sah (Gheysen et al. 1991; Lehman et al. 1994; Puchta 1998).